Title 3’-末端にグルコサミン類を有するオリゴヌクレオチド誘導体の合成とその生物活性の評価( 本文(Fulltext) ) Author(s) 羅, 雄 Report No.(Doctoral Degree) 博士(薬科学) 連創博甲第23号 Issue Date 2013-09-30 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/47863 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
3'-末端にグルコサミン類を有するオリゴヌクレオチド
誘導体の合成とその生物活性の評価
Synthesis of oligonucleotides with glucosamine at the 3'-position and
evaluation of their biological activities
2013
目次 第 1 章 緒言 1 第 2 章 研究の背景及び目的 3 2 - 1 RNA 干渉の歴史 3 2 - 2 RNA 干渉の機構 5 2 - 3 RNA 干渉の特徴 6 2 - 4 siRNA の医薬品化への問題点 8 2 - 5 siRNA の DDS 研究 9 第 3 章 グルコサミン誘導体による siRNA の 3'-末端化学修飾 21 3 - 1 塩基部非置換型グルコサミンモノマー誘導体の合成 23 3 - 2 塩基部置換型グルコサミンモノマー誘導体の合成 26 3 - 3 3'-末端にグルコサミン誘導体を含む siRNA の合成 29 第 4 章 3'-末端にグルコサミン誘導体を含む siRNA の生物学的活性評価 31 4 - 1 siRNA 二本鎖熱的安定性評価 31
4 - 2 Dual luciferase reporter assay によるタンパク発現抑制評価 32
4 - 3 Snake Venom Phosphodiesterase によるヌクレアーゼ耐性評価 33
4 - 4 細胞膜透過性評価 34 第 5 章 結語 37 実験の部 43 参考文献 68 謝辞 74 参考資料 75
第 1 章 緒言 2000 年から 2011 年までの 12 年間、世界の医薬品市場は 2 倍以上の規模に 拡大しており、年平均成長スピードは約 6%で堅調に伸びている (Fig. 1-1) 。 また、世界中における人口増、高齢化などの要因から、その成長率はまだまだ 保持できると予想されている。 *日本製薬工業協会 DATA BOOK 2013 この大きな産業の発展を推進するのは医薬品の開発、いわゆる創薬である。 これまでの創薬は有機合成によるランダムな化合物から新薬の可能性を調べ 尽くす、もしくは微生物発酵物や動植物からの抽出物の分析による新薬の成分 を探すという方法であった。しかし、このような創薬方法は資金、労力と時間 がかなりかかるため、近年バイオ医薬品を中心とした創薬事業が進んでいる。 バイオ医薬品の開発は特定疾患を引き起こす要因となっている遺伝子・タン Fig. 1-1 医薬品市場規模の推移 (販売額) *
パク質の解明を基に生物によって生産される物質 (タンパク質、核酸等) 、治 療法を探索することである。利点としては、以前の大量の候補化合物からのラ ンダムスクリーニングに対して、メカニズムを解明することで、明確な創薬標 的を有する新薬の創製が可能な点が挙げられる。このバイオ医薬品の発展は大 きく 3 つの段階に分けられ、現在すでに第三世代の核酸医薬時代に入っている (Table 1-1) 。 Table 1-1 バイオ医薬品の発展 世代 医薬品名 タイプ 作用 第一世代 (1980 年代から) ホルモン サイトカイン タンパク質 ホルモンなどの投入により、生体活 性化を促進する。 第二代 (1990 年代から) 抗体医薬 タンパク質 特定の標的分子の抗原抗体結合に より、がんなどの疾病を治療する。 第三世代 (2000 年代から) 核酸医薬 DNA / RNA 遺伝子レベルの調整により、疾病の 治療と予防に関与する。 核酸医薬は第一、第二世代と比べ、治療のターゲットをタンパク質からさら に前の段階に遡り、遺伝子レベルに至る。主な核酸医薬品は mRNA を標的と し、その配列を特異的に認識し、標的タンパク質の発現を抑制する。例えば、 アンチセンス法、リボザイム法と siRNA 法である。その中で、著者は siRNA 法に注目し、RNA 創薬の観点で研究を行った。
第 2 章 研究の背景及び目的
2 - 1 RNA 干渉の歴史
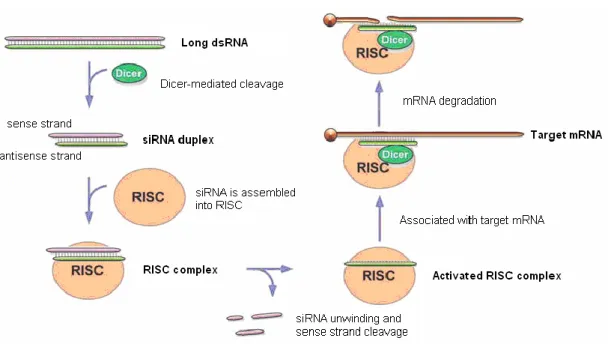
RNA 干渉 (RNA interference: RNAi) とは、二本鎖 RNA (double strand RNA : dsRNA) が相補的な標的 mRNA を特異的に分解し、標的タンパク質の発現を抑制する 現象である。この手法は 2001 年 5 月に開催された RNA 学会 (RNA Society) で 初めて発表されたが、最初にこの現象が発見されたのは、1990 年の植物実験 であった。Rich Jorgensen 氏はペチュニアに紫色の色素合成に関与する外来遺 伝子を導入することで強い紫色を発色させようとしたが、得られたのは予想に 反して斑入りの花であった1) (Fig. 2-1) 。この現象は、外来遺伝子が何らかの 原 因 で 外 来 遺 伝 子 及 び 内 在 性 の 遺 伝 子 の 発 現 を 抑 制 し た と 推 察 さ れ 、 Co-suppression と名付けられた。さらに同じ時期に他の植物研究所において、 植物が RNA ウイルスの感染により RNA が特異的に分解されることが発見さ れた 2,3,4,5)。それ以降、転写後遺伝子サイレンシング (post-transcriptional gene silencing; PTGS) あ る い は 、 ウ イ ル ス 誘 発 性 遺 伝 子 サ イ レ ン シ ン グ (viral-induced gene silencing; VIGS) と呼ばれた。
*http://www.easypedia.gr/el/images/shared/2/24/Rnai_phenotype_petunia_crop.pngより引用
1995 年、Guo 氏と Kemphues 氏によって線虫 (C. elegans) 遺伝子発現の抑制 実験により、センス RNA がアンチセンス RNA と同程度に遺伝子発現を抑制 することを発見した6)。続いて 1998 年、Fire 氏らは線虫を利用し、dsRNA が センス RNA やアンチセンス RNA 単独よりも 10 倍強く遺伝子発現を抑制する ことを明らかにした7)。この発見で、植物以外において RNAi が初めて見出さ れた。線虫での RNAi の報告に相次いで、プラナリア8)、ヒドラ9)、ショウジ ョウバエ 10)などの無脊椎動物、また、ゼブラフィッシュ 11)のような脊椎動物 においても観察されることが報告された。哺乳類では、長鎖 dsRNA を導入す るとインターフェロン応答が起こり、哺乳類細胞への応用はできないと考えら れていたが、2001 年にElbashir 氏らによって短い dsRNA を導入することにより、 インターフェロン応答を回避できることを明らかにした12) (Fig. 2-2) 。RNA 干 渉は、医学や生物学などの研究に大きな影響を与え、2006 年のノーベル医学 生理学賞の受賞対象となった。 プラナリア 線虫 ヒドラ ショウジョウバエ ゼブラフィッシュ マウス胚 Fig. 2-2 RNA 干渉に研究される多種の動物
2 - 2 RNA 干渉の機構
RNAi は dsRNA により引き起こされる配列特異的に遺伝子発現を抑制する 現象である。細胞内に導入された長鎖 dsRNA は、Dicer と呼ばれる RNase III ファミリーに属する酵素により、3'-末端側に 2 塩基の突出 (ダングリングエン ド) を持つ 21~23 塩基の短鎖 dsRNA である siRNA へと分解される。その後、 siRNA は複数のタンパク質と RNA 誘導型サイレンシング複合体 RISC (RNA induced silencing complex) に組み込まれ、RISC の組成たんぱく質 Argonaute (AGO) が持つ Slicer 活性により短鎖 dsRNA のセンス鎖が切断される。RISC に残ったアンチセンス鎖はガイド分子として相補的な配列を含む mRNA 鎖を 特異的に認識し13)、最終的に標的 mRNA が速やかに分解され、タンパク質の 発現を抑制する14,15)
(Fig. 2-3) 。
2 - 3 RNA 干渉の特徴
2 - 3 - 1 RNAi の高効率性
Elbashir 氏らは、dsRNA による遺伝子抑制に関する研究において、dsRNA 濃 度が 25 nmol/L と 100 nmol/L の場合はサイレンシング効果はほぼ変わらなかっ たと報告している。また、dsRNA 濃度を 1.5 nmol/L まで下げても、変化も殆 どなく、サイレンシング効果が完全に消えるのは 0.05 nmol/L の低濃度であっ た16)。Holen 氏らも dsRNA が 1~100 nmol/L の濃度下、遺伝子サイレンシング 効果が一致することを明らかにした 17)。つまり、アンチセンス法による遺伝 子サイレンシングと異なり、dsRNA を介して RNAi は化学量論的ではなく、 遺伝子サイレンシング現象が触媒的に高効率で進行することを示している。
2 - 3 - 2 RNAi の特異性
Elbashir 氏らと Brummel Kamp 氏らにより、21~23 塩基の siRNA 配列の中に 標的 mRNA と 1~2 塩基の mismatch が存在する場合、その mRNA の分解効果 が著しく減少することが報告された。RNAi により遺伝子サイレンシング現象 は siRNA の相補的な配列を含む標的 mRNA を特異的に分解し、タンパク質の 発現を抑制することが明らかになった。
2 - 3 - 3 RNAi の位置効果
Holen 氏らはヒト組織因子 (Tissue Factor: TF) 発現の抑制効果実験で配列の 違う位置を標的とする 4 種類の dsRNA (hTF167i、hTF372i、hTF478i、hTF562i) を用意したが、その中で最も活性高いのは hTF167i と hTF372i で、約 85%~90% の遺伝子発現を抑制した。一方、hTF478i の活性は殆どなかった 18)。また、 hTF167i を中心として三塩基づつ左右へずらし、新たに用意した dsRNA の活 性もかなり減少した。すなわち、RNAi の位置効果が存在し、配列に少しでも
位置のずれが生じた場合、siRNA の遺伝子サイレンシング効果に大きな影響を 与える。
2 - 3 - 4 RNAi の競合性
Holen 氏らによる 10 nmol/L 及び 30 nmol/L の hTF167i による TF 発現の抑制 効果には差が殆どなかったが、10 nmol/L の hTF167i と 20 nmol/L の低い抑制 活性 PSK314i を混合した後では hTF167i の抑制効果も大きく減少することが報 告された。これは RNAi の競合性とも言える。
2 - 3 - 5 RNAi の伝播性
Feinberg 氏と Hunter 氏は線虫に SID-1 という膜貫通タンパクを発見した。こ のタンパクを介して細胞内に導入された dsRNA を細胞外へ転送すると、線虫 の全身まで拡散できる19)。しかし、ショウジョウバエに SID-1 の同源タンパク が発見されなく、ショウジョウバエにおいては、RNAi の伝播ができないとい う報告もあった。
2 - 3 - 6 RNAi の ATP 依存性
RNAi 現象は ATP を除去した環境で起こらないことにより、RNAi は少なく とも二つの段階で ATP 依存的であることが明らかになった。一つ目は Dicer により切断された短鎖 dsRNA が RISC に取り込まれ、ATP 依存的に Dicer 側か ら巻き戻しを受け一本鎖化する段階と、二つ目は RISC 複合体に切断された mRNA が、ATP 依存的に RISC から追い出される段階である20)。
2 - 4 siRNA の医薬品化への問題点 RNAi 現象を引き起こす要因の一つである siRNA は、特異的な遺伝子抑制効 果を有するため、生物学及び医薬分野の基礎研究に応用されているとともに、 臨床への応用も期待されている。しかし、今まで一般的な siRNA は効果的に RNA 干渉機能 (ノックダウン効果) を発揮させるには充分ではなく、RNA 創 薬の実現には以下に示すような幾つかの大きな問題点を抱えている。 2 - 4 - 1 オフターゲット効果
siRNA のオフターゲット効果 (off-target effect) とは、導入された siRNA が 標的とする遺伝子以外の遺伝子の発現も同時に抑制してしまう現象である。こ のオフターゲット効果は大きく 2 種類に分けられる21)。一つ目は siRNA 配列 の選択性が弱く、他の遺伝子に配列相同性を示す場合に起こる。二つ目は、免 疫系防御機構によるサイトカイン分泌に起因する免疫反応である (Fig. 2-4) 。 Fig. 2-4 siRNA のオフターゲット効果 参考文献 21 より一部改変
2 - 4 - 2 酵素的安定性 siRNA は DNA 分子と同様に化学的に比較的安定であるが、生体内でのリボ ヌクレアーゼ (RNase) などにより分解され、短時間に腎臓から排出される。 2 - 4 - 3 作用点への送達 siRNA は生体内に投与した後、RNase などの酵素による分解される他に、投 与部位から標的とする目的細胞に到達するまでに多くの障壁がある。例えば、 タンパク質への吸着、腎臓の他肝臓や脾臓での取り込みによる分解、静脈投与 での血管壁の透過、目的細胞膜の透過など、さまざまな障壁を越えなければな らない。 2 - 4 - 4 作用の持続性 siRNA による治療ではアンチセンス法やアプタマー法などと異なり、 siRNA/RISC 複合体で標的 mRNA を持続的に分解できるため、その作用は一回 投与すると一週間程度に維持することが可能である。しかし、一回投与で一ヶ 月間以上の長期効果を持続することは未だ困難である。 上記四つの問題点が存在するため、siRNA などのオリゴ核酸の医薬品化まで、 まだ多くの研究が必要である。現在最も注目されている研究分野の一つが、 siRNA の薬物送達システム (Drug Delivery System: DDS) である。すなわち、 問題点 2, 3, 4 の解決を目指している。
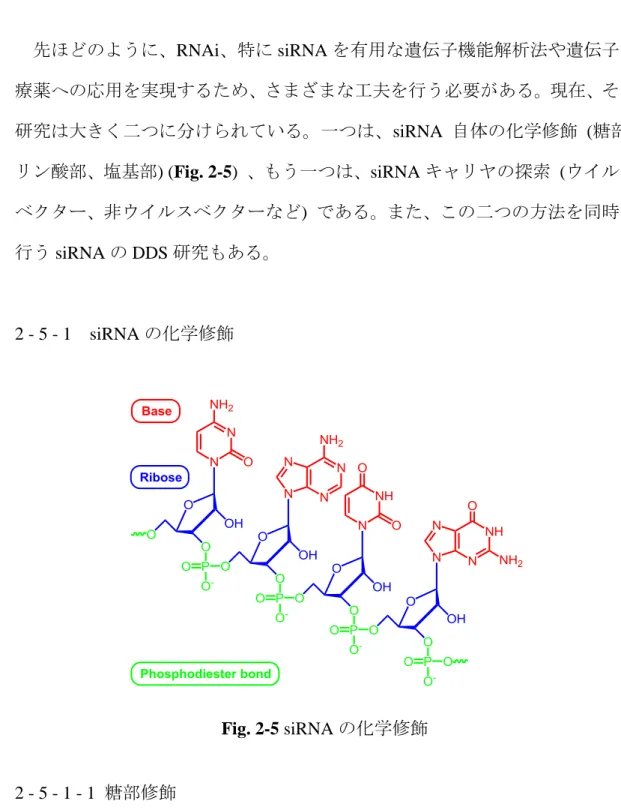
2 - 5 siRNA の DDS 研究 先ほどのように、RNAi、特に siRNA を有用な遺伝子機能解析法や遺伝子治 療薬への応用を実現するため、さまざまな工夫を行う必要がある。現在、その 研究は大きく二つに分けられている。一つは、siRNA 自体の化学修飾 (糖部、 リン酸部、塩基部) (Fig. 2-5) 、もう一つは、siRNA キャリヤの探索 (ウイルス ベクター、非ウイルスベクターなど) である。また、この二つの方法を同時に 行う siRNA の DDS 研究もある。 2 - 5 - 1 siRNA の化学修飾 2 - 5 - 1 - 1 糖部修飾 siRNA の化学修飾に関して、最も多く報告されていたのは糖部の修飾法であ る22-29)
(Fig. 2-6) 。siRNA 糖部の 2′-OH の有無は RNAi パスウェイに入ること に必要がない30)ため、遺伝子抑制活性に影響を与えないと考えられ、2′位の修 飾が多く報告されている。2′-OH をメチル化した 2′-O-Me 修飾により、相補鎖
との結合親和力やヌクレアーゼ耐性が向上し、化学的に安定な二本鎖構造を保 持できる。さらなる高いヌクレアーゼ耐性を有し、効果的な遺伝子抑制活性を 維持するためには、2′-O-Me 修飾のアナログ体の導入数を制限し、また導入部 位の選択が必要となる。
2′-OH をデオキシ化した 2′-H の DNA 型も、RNA アナログと考えられる。 最初の3′-末端のダングリングエンド部位に DNA 型を導入すること22)から、現 在、DNA 型の導入数と導入部位を制限すれば、siRNA に応用できることまで 発展してきた。また、Ui-Tei 氏らにより、siRNA の 5′-末端側から幾つかの塩 基を DNA 型に置換すると、オフターゲット効果が減少することが報告されて いる31)。 Fig. 2-6 siRNA 糖部の化学修飾
アニオン性の siRNA は、細胞膜に透過することが困難であるため、細胞膜 との親和性や電荷の中和などを考慮し、アミノ基を導入することにより、プロ トン化されたアミノ基の正電荷が隣接するリン酸ジエステルの負電荷を中和 し 、 細 胞 膜 の 透 過 性 が 向 上 で き る と 考 え ら れ る 。 2′- 位 に ア ミ ノ エ チ ル (2′-O-AE) 基を初め、より炭素鎖の長いアミノプロピル (2′-O-AP) 基や複数の アミノ基を有するグアニジノエチル (2′-O-GE) 基は、より高い安定性が得られ ている23-25)。
また、2′-OH をフッ素及びアジド基に置換した 2′-F-RNA と 2′-N3-RNA のよ うな糖部修飾型ヌクレオシドも報告されている 26, 27)。これらの修飾により、 二本鎖の結合親和力や血中での安定性が増強することが報告されている。 天然型核酸リボースのパッカリングにより、N 型と S 型の 2 種類の構造をと り得るため、化学構造上形の自由度が大きい。そこで、リボース環の 2′-位と 4′-位をメチレンで架橋することによって、自由度が制限された LNA (Locked Nucleic Acid) の修飾法も研究が進められている。完全に N 型に固定される LNA は二本鎖の安定性が大幅に向上することにより、血中滞留時間及びヌク レアーゼ耐性の向上、さらにオフターゲット効果の減少が期待される28, 29)。 一方、リボース環の酸素原子を硫黄原子に置換した4′-S-RNA も報告された 32)。熱力学的に安定な二本鎖を形成し、高いヌクレアーゼ耐性も保持できると 共に、標的分子との相互作用、三次構造の多様性、及び触媒活性の向上などの 利点がある。 2 - 5 - 1 - 2 リン酸部修飾 siRNA の構造変化を最小限に抑え、ヌクレアーゼなどの酵素に耐性を上げる ため、リン酸部 (リン酸ジエステル) の修飾もよく研究されている (Fig. 2-7) 。 ただし、修飾を施すのに伴い、リン原子にキラリティが発生し、立体異性体が 生じてしまうことに注意が必要である。その中に最も有名な修飾法は、リン酸
ジエステル結合の非結合性酸素原子を硫黄原子に置換したチオリン酸エステ ル (PS) 結合である。この修飾法で合成された RNA は加水分解酵素に強い耐 性をもつことで、血清中での安定性が向上することが報告されている 33)。 siRNA 鎖の末端に PS 結合の導入型と未修飾型は遺伝子抑制能が同等であった が、鎖全体に PS 結合を導入すると抑制能低下と細胞毒性上昇という問題点も 引き起こされる。 また、ヌクレアーゼ耐性と安定性を向上するため、硫黄原子以外にリン酸ジ エステル結合の非結合性酸素原子をボラン (-BH3) 、メチル基 (-CH3) 及びア Fig. 2-7 siRNA リン酸部の化学修飾
ミノ基 (-NH2) などの官能基に置換される修飾法も報告されている34)。 さらに、リン酸ジエステル結合の非結合酸素原子だけの置換ではなく、リン 酸ジエステル自体をアミド結合に変更した siRNA の修飾も報告されている。 未修飾型と比べ、3′-位により安定な構造をとるため、相補鎖 RNA とより安定 な二本鎖を形成する35)。 その他に、3′,5′-位のリン酸ジエステル結合を 2′,5′-位間の結合へ変化させた siRNA 修飾法が報告されている。天然型より 2′,5′-結合型 siRNA は核酸分解に 耐性が強くなるが、二本鎖 siRNA 中に、センス鎖のみに容認でき、アンチセ ンス鎖が容認できないという問題点もある36)。 2 - 5 - 1 - 3 塩基部修飾 siRNA 塩基部分の修飾に関する報告例は少なくないが、遺伝子抑制能の低下 を伴うため、多くの場合実用に至っていない (Fig. 2-8) 。代表的なものとして、 5-ブロモウリジン、5-ヨードウリジン、及びアデニン誘導体である 2,6-ジアミ ノプリンの塩基部修飾体は、A-U 塩基対を安定化させることにより siRNA へ の導入が実施されている。しかし、これらの修飾体が導入された siRNA は、 いずれも未修飾型 siRNA より、遺伝子抑制能が大幅に低下する傾向にあった。 その他に、2-チオウリジン、シチジンと塩基対を形成するシュードウリジン の修飾は、二本鎖の熱力学的安定性、遺伝子抑制ならびにターゲット特異性を 向上させた 37)。一方、5-位にプロピニルを導入されウリジン置換 siRNA 修飾 体は二本鎖の熱力学的安定性の向上を伴い、遺伝子抑制能を低下することも報 告されている38)。
2 - 5 - 2 siRNA キャリア 直接に siRNA の糖部、リン酸部及び塩基部を化学的に修飾する以外に siRNA キャリアを利用し、低分子 siRNA の分子量を多くすることにより、血中滞留 時間の増加を目指す手法もある。それは、ウイルスキャリアと非ウイルスキャ リヤの大きく 2 種類に分けられている。 ウイルスベクターにヘアピン構造をコードする遺伝子 (shRNA or miRNA) が導入され、宿主細胞内に RNA 分子を発現し、最終的に siRNA へ分解される ことが報告された39)。ウイルスキャリアの DDS は一回の投与が比較的長時間 siRNA の発現が保持でき、かつ、一般的にデリバリー困難とされる神経細胞ま で届くことも可能という利点がある。すなわち、慢性疾患である HIV 感染症 や神経変性病であるハンチントン病、アルツハイマー病などの治療に向いてい ると考えられる40, 41)。現在、ウイルスキャリアの DDS は in vivo での応用も多 数報告されている 42, 43)が、強い免疫反応のリスク、肝臓毒性 44)及び弱いベク ター安定性といった欠点はウイルスキャリヤの実用化に障壁となっている45)。 一方、非ウイルスキャリヤの場合は、免疫反応などの安全性面に優れている Fig. 2-8 siRNA 塩基部の化学修飾
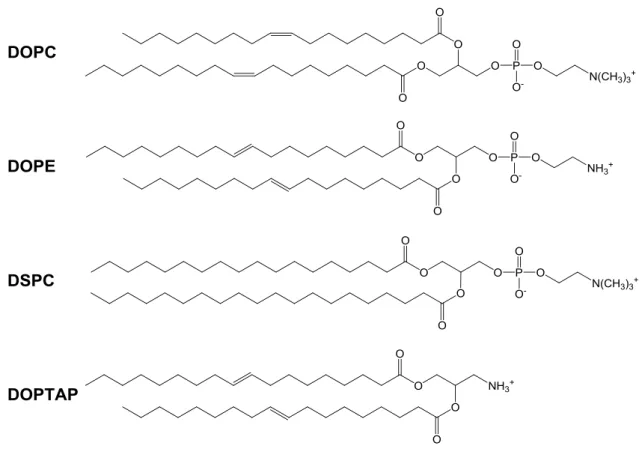
ことが明らかになった 46)。近年、ナノ技術を活用し、さまざまなカチオン性 物質と複合体を形成させた siRNA の DDS 報告例が多くなってきた 48-52) (Fig. 2-9) 。カチオン性脂質はアニオン性 siRNA と自発的に結合し、また細胞膜と の親和性も良く、代表的な非ウイルスキャリアとして、多く研究が実施されて きた。また、ポリエチレン・グリコール (PEG) でのコーティング化により、 毒 性 を 抑 制 す る こ と も 可 能 で あ る 53) 。 さ ら に 、 1, 2-dioleoyl-3-trimethylammonium-propane (DOTAP) 54)は in vivo での実験で、 siRNA の DDS による腫瘍壊死因子 (Tumor Necrosis Factor: TNF) と血管内皮細 胞増殖因子 (Vascular Epithelial Growth Factor: VEGF) の発現を抑制することが 報告された55, 56)。
Fig. 2-9 siRNA のカチオン性脂質キャリア
カチオン性脂質以外に、ポリマーを siRNA の DDS へ応用する研究もよく知 られている (Fig. 2-10) 。カチオン性ポリマーと siRNA のアニオン性リン酸エ ステルの自己集合 (Fig. 2-11) により、ゲル状粒子を形成し、siRNA を効率的 に移送することができる。 Fig. 2-11 siRNA とポリカチオン性物質の自己集合 参考文献 57 より一部改変 Fig. 2-10 siRNA のポリマーキャリア 参考文献 47 より一部改変
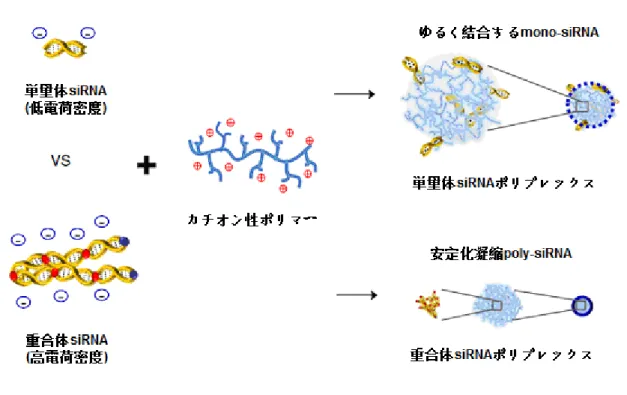
また、最近の研究は負電荷密度の低い単量体 siRNA 分子はカチオン性ポリ マーとの自己集合が弱いため、siRNA 分子がナノ粒子ポリプレックスの表面に 浮きやすく、生体内にヌクレアーゼなどによる分解されるリスクが高い。この 問題点を解決するため、低分子 siRNA から、ハイブリダイゼーション siRNA59) や重合体 siRNA60-64)への開発が挙げられている。これらの手法により、siRNA 分子の負電荷密度を増やし、カチオン性ポリマーとの自己集合を安定させるこ とと共に、そのポリプレックスの直径を 100 nm 程度 (200 nm 以上になると、 速やかに肝臓及び脾臓の細網内皮系 RES による排出される 65) 。) に維持し、 排出作用も抑制できる (Fig. 2-12) 。 カチオン性ポリマーは、細胞膜透過性を高めるため直径 100nm 以下のナノ 粒子になるが、デンドリマーのような巨大分子を利用することも可能である (Fig. 2-13) 。デンドリマーとは、ポリアミドアミン (Polyamidoamine: PAMAM)
Fig. 2-12 単量体 siRNA 分子及び重合体 siRNA 分子の自己集合
に代表される規則的な分岐構造をもつ樹状高分子を母核に、末端にはアミノ基 などの分子と結合しやすい側鎖を有する構造を基本骨格とするものである。同 一構造の分子が集合し、親和力を増強するクラスター効果が得られるという利 点が注目されている66)。 一方、ペプチドの細胞取り込みを増進する特徴を利用した siRNA の DDS 応 用も進められている (Fig. 2-14) 。 その中に、細胞透過性ペプチド (CPP : Cell Fig. 2-13 siRNA のデンドリマーキャリア 参考文献 47 より一部改変
Penetrating Peptides) というカチオン性ペプチドはデリバリーキャリアとして、 複合体の形で目標分子を細胞内へ容易に運ぶことがよく知られている。例えば、 HIV-1 由来の TAT67)及びインフルエンザウイルス由来の INF-1 と INF-768)はウ イルスタンパク質由来の CPP である。また、CPP の siRNA デリバリへの応用 は 2 種類に分けられている。一つ目は、CPP と siRNA が共有結合で連結した ものである (主にジスルフィド結合及びチオエーテルリンカー) 。二つ目は、 静電作用によって CPP と siRNA を複合体にしたものである。細胞内への輸送 効率が高いため、現在、共有結合法は多く利用されているが、遺伝子抑制能の 低下も問題点となる69)。これは共有結合が強すぎ、RISC への取り込みを阻害 すると考えられる。ただし、ペプチドの共有結合法で siRNA の遺伝子抑制能 を保持できるとの報告もある70)。 GLFEAIAGFIENGWEGMIDGGGC (INF-1) GLFEAIEGFIENGWEGMIDGWYG (INF-7) GALFLGFLGAAGSTMGA-WSQP-KKKRKV (MPG) Ac-GALFLAFLAAALSLMGLWSQPKKKRKV-Cya (MPGα) PRRRRSSSRPVRRRRRPRVSRRRRRRGGRRRR (Protamine) WEAALAEALAEALAEHLAEALAEALEALAA (GALA) GRKKKRRQRRRPPQ (TAT) IRQRRRR (IRQ) RQIKIWFQNRRMKWKK (Penetratin-ABT) Fig. 2-14 siRNA のペプチドキャリア 参考文献 47 より一部改変
第 3 章 グルコサミン誘導体による siRNA の 3'-末端化学修飾 著者が所属する研究室では、siRNA の 3'-末端のダングリングエンド部位の 化学修飾に着目し、さまざまな高機能性 RNA 分子の開発研究を行っている 71-73) (Fig. 3-1) 。 その様な過程で、著者はカチオン性ポリマーであるキトサンを注目し、 siRNA の 3'-末端修飾を行った。キトサンは生分解能を有しており、その分解 過程で生産されるグルコサミンは、人体重要な構成成分である糖タンパクに多 く存在するため、他のカチオン性物質と比べ、低い毒性、高い生体適合性とい う利点が示されている。また、細胞膜親和性、抗菌性及び親水性などの特徴も 有している。3'-末端の化学修飾には高分子のキトサンを直接利用するのが困難 であるため、構成糖のグルコサミンを選択し、導入することにした。 最初に開発したグルコサミンモノマー誘導体の合成法は、Scheme 3-1 に示し ている。まず、グルコサミン塩酸塩 1 から水酸基を O-アセチル基で保護、ア ミノ基をフタルイミド基で保護してから、1'-位に SPh 基に置換させ、化合物 3 を得た。次に、1'-位にリンカーを導入してから、3'-, 4'-, 6'-位を脱保護し、化合 物 5 を得た。そして、1 級と 2 級の水酸基の活性差を利用し、TBDMS 基とベ Fig. 3-1 siRNA の 3'-末端のダングリングエンドに導入される化合物
ンゾイル基でそれぞれ 6'-位と 3'-, 4'-位を保護し、化合物 7 を得た。最後に、リ ンカー末端のベンジル基を DMTr に置換させ、6'-位を脱保護してから、スクシ ニル化、CPG 樹脂と結合、合わせて全 10 ステップで CPG 樹脂体 11 まで合成 できた。
Scheme 3-1 グルコサミンモノマー誘導体の合成法
Reagents and conditions: (a) (i) MeONa, MeOH, rt; (ii) Phthalic anhydride, MeOH, rt; (iii) Acetic anhydride, pyridine, rt, 68%; (b) PhSH, Et2O·BF3, CH2Cl2, rt, 93%; (c) benzyloxyethanol, NIS, TfOH, CH2Cl2, 4Å MS, rt, 60%; (d) MeONa, MeOH, rt, 85%; (e) TBDMSCl, pyridine, rt, 97%; (f) BzCl, pyridine, rt, 95%; (g) Pd(OH)2/C, H2, THF, rt, 95%; (h) DMTrCl, pyridine, rt, 88 %; (i) TBAF, THF, 0ºC, 35%; (j) (i) Succinic anhydride, DMAP, pyridine, rt; (ii) CPG, EDC, DMF, rt, 28
しかし、この合成法では、DNA/RNA 自動合成機により、オリゴ核酸に導入 した後、グルコサミンの保護基を完全に外すこと及びオリゴ核酸の精製が困難 であった。具体的には、アンモニア処理の時に、長時間処理によるオリゴ核酸 の分解を回避するため、12 時間と定められていたが、グルコサミン 2'-位のフ タルイミド基は 100%で脱保護ができず、脱保護が不完全な siRNA の存在が MALDI-TOF/MS で確認された (Fig. 3-2) 。続いてのアッセイ試験でも新規に 合成された siRNA の純度の影響で正確な結果が得られなかった (未発表) 。 そこで、本研究ではその合成法を改善し、グルコサミン 6'-位に塩基部非置 換型、又は塩基部置換型 2 種類のモノマー誘導型 CPG 樹脂を用意した。 3 - 1 塩基部非置換型グルコサミンモノマー誘導体の合成 先ほど述べた合成法の問題点が 2'-位のフタルイミド基の脱保護であるが、 最初から無水フタル酸での保護ステップをとばし、アミノ基がフリーのままで 合成されると、1'-位のアノマー炭素が α 型と β 型の混合物になってしまうため、 合成中に別の段階でフタルイミド基を変えるしかないと考えられる (Scheme 3-2) 。 Fig. 3-2 従来法による合成された siRNA の混合物
そこで、グルコサミン塩酸塩 1 を出発原料として、MeOH に溶解し、MeONa を加え、2'-位に遊離のアミノ基にした後、無水フタル酸にて保護した。2'-位の アミノ基が消えたことを TLC で確認した上、溶媒を減圧留去し、一晩乾燥さ せた。MeOH を完全に除去してから、乾燥ピリジン中、無水酢酸と反応させ、 1'-, 3'-, 4'-, 6'-位の水酸基を全保護した化合物 2 が 68%収率で得られた。 続いて、1'-位アセトキシ基をルイス酸で容易に脱離可能なチオフェニル基に 変換することとした。化合物 2 は Et2O·BF3存在下で PhSH を作用させ、93%収 Scheme 3-2 塩基部非置換型グルコサミンモノマー誘導体の合成
Reagents and conditions: (a) (i) MeONa, MeOH, rt; (ii) Phthalic anhydride, MeOH, rt; (iii) Acetic anhydride, pyridine, rt, 68%; (b) PhSH, Et2O·BF3, CH2Cl2, rt, 93%; (c) Ethylenediamine, MeCN, reflux, 60%; (d) TIPSCl, pyridine, rt, 86%; (e) (CF3CO)2O, pyridine, rt, 79%; (f) BzCl, pyridine, rt, 83%; (g) benzyloxyethanol, NIS, TfOH, CH2Cl2, 4Å MS, rt, 78%; (h) Pd(OH)2/C, H2, THF, rt, 89%; (i) DMTrCl, pyridine, rt, 98%; (j) TBAF, THF, rt, 68%; (k) (i) Succinic anhydride, DMAP, pyridine, rt; (ii) CPG, EDC, DMF, rt, 19 μmol/g.
率でβ 型のみのチオフェニル体 3 を得た。 次に、従来法と異なり、エチレンジアミンの強い塩基性条件下に高温還流さ せ、一気に化合物 3 の 2'-位のフタルイミド基と 3'-, 4'-, 6'-位のアセチル基を外 し、全脱保護のチオフェニル体 12 が 60%収率で得られた。この化合物は極性 が高いため、一般のクロロホルム-メタノール系とヘキサン-酢酸エチルエス テル系の展開溶媒を用いたシリカゲルカラムクロマトグラフィーでは困難で あり、酢酸エチル:メタノール:水 = 8 : 1.5 : 1 を用いて精製した。 化合物 12 から、一連の水酸基とアミノ基の保護反応を行い、化合物 15 まで 進んだ。まずは、6'-位に TIPSCl にてシリル基で保護、86%収率で化合物 13 が 得られた。そして、フタルイミド基及びアセチル基と比較し塩基性条件下容易 に脱離するトリフルオロアセチル基を無水トリフルオロ酢酸にて 2'-位アミノ 基を保護し、79%収率で化合物 14 を得た。続いて、3'-, 4'-位の水酸基を従来法 と同様に BzCl で保護し、83%収率で化合物 15 を得た。 その後、固相担体 CPG 樹脂と結合するため、介するリンカーを導入した。 4Å のモレキューラシーブの存在下で、化合物 15 にベンジルオキシエタノール、 NIS 及びルイス酸 TfOH を順に加え、反応させた。化合物 16 を 78%収率で得 た。 リンカー末端のベンジル基をトリチル基に変更するため、まず接触水素化反 応を行い、THF 溶媒中に Pd(OH)2/C を触媒として、化合物 16 は水素にて、化 合物 17 に還元され、89%収率で反応を終えた。そして、DMTrCl でリンカー末 端の水酸基にトリチル基を導入し、98%収率で化合物 18 が得られた。この段 階から、トリチル基の脱離を防ぐため、シリカゲルカラムクロマトグラフィー による精製ステップは、溶媒に少量のピリジンを加え、弱塩基性環境を保持す る必要がある。 次に、6 位の TIPS 基を TBAF で脱保護し、68%収率で化合物 19 を得た。 最後に、ピリジン中に DMAP の存在下、無水コハク酸と反応させ、定量的
にスクシニル体を導いた。この段階は、シリカゲルカラムクロマトグラフィー による精製は行わず、抽出のみを行った後、完全に乾燥させ、DMF 中に、CPG 樹脂及び EDC-HCl を加え、樹脂体 20 を得た。CPG 樹脂 1g に結合しているグ ルコサミンモノマーの量 (μmol/g) (CPG 樹脂の活性) は、樹脂体 20 を過塩素 酸 : エタノール = 3 : 2 溶液で脱保護し、そのろ液の波長 498 nm でのトリチ ル基の吸光度から算出し、樹脂体 20 の活性は 19.3 μmol/g であった。 この樹脂体 20 は、先に合成した樹脂体 11 と比べ、2'-位のフタルイミド基を より容易に脱離するトリフルオロアセチル基に変更した。したがって、核酸オ リゴマーの合成後のアンモニア処理による保護基の脱離が効率的に進むこと が期待される。 3 - 2 塩基部置換型グルコサミンモノマー誘導体の合成 siRNA の 3'-末端ダングリングエンドへ導入されるグルコサミン分子は化学 修飾の観点から見ると、糖部とその架橋部の修飾とも言える。つまり、リボー スからグルコサミンへ、また、リン酸ジエステルからリンカーへの変更である。 しかし、この化学修飾は、塩基部がない、糖部のみの変更なので、塩基部が存 在するグルコサミンモノマー誘導体の合成も行った。 まずは、保護した塩基の用意が必要となる。今回選択した塩基はチミン、保 護基はベンゾイル基であった。 (Scheme 3-3) 。 Scheme 3-3 塩基の保護
Pyridine の存在下、チミンを直接 1.0 当量の BzCl とベンジル化した際、
N1-Bz-Thymine、N3-Bz-Thymine、また N1,N3-Bz-Thymine 多種の生成物ができて しまい、収率は低く、精製も困難であったため、2 段階反応に変更した。Thymine を 過 剰 量 のベ ン ゾイル ク ロ リ ド (BzCl) (3.0 当 量 ) と 反応 させ 、 す べ て N1,N3-Bz-Thymine にした後、選択的に 1-位を脱保護し、N3-Bz-Thymine を 45% 収率で結晶として得た。 本研究では、グルコサミンモノマー誘導体合成の従来法を改善すると共に、 6'-位にチミン置換型グルコサミンモノマー誘導体も合成した (Scheme 3-4) 。 Scheme 3-4 塩基部置換型グルコサミンモノマー誘導体の合成
Reagents and conditions: (a) Benzyloxyethanol, Me2O·BF3, MeCN, reflux, 67%; (b) MeONa, MeOH, rt, 88%; (c) N3-Benzoyl-thymine, PPh3, DEAD, THF, rt, 84%; (d) Ethylenediamine, EtOH, reflux, 87%; (e) (CF3CO)2O, pyridine, rt, 90%; (f) Pd(OH)2/C, H2, THF, rt, quant; (g) DMTrCl, Pyridine, rt, 79%; (h) (i) Succinic anhydride, DMAP, pyridine, rt; (ii) CPG, EDC, DMF, rt, 37 umol/g.
3 - 1 で合成された化合物 2 を原料として、Me2O·BF3の存在下、リンカーの ベンジルオキシエタノールと MeCN 中に高温還流させ、チオフェニル化ステ ップを省略し、リンカーを 1'-位に直接導入した。化合物 21 は 67%収率で得ら れた。そして、3 - 1 のように同時に 2'-位のフタルイミド基と 3'-, 4'-, 6'-位のア セチル基を外すことなく、MeONa の比較的に弱い条件で 2'-位のフタルイミド 基を残したまま、3'-, 4'-, 6'-位のアセチル基を除去し、88%収率で化合物 22 を 得た。この段階で 2'-位のフタルイミド基も除去した場合は、次の Mitsunobu 反応による 6'-位に塩基の導入は副生成物ができ、精製困難であるため、収率 も下がることになった (Fig. 3-3) 。 続いて、Mitsunobu 反応を行い、化合物 22 は PPh3と DEAD 存在下、THF 中 に N3 -Benzoyl-thymine と反応させたところ、副生成物が生じることなく、84% 収率で化合物 23 を得た。ここから、3 - 1 での合成法とほぼ同様に、エチレン ジアミンでフタルイミド基を外し、トリフルオロアセチル基に変えたところ、 各々87%、90%収率で化合物 24 と 25 を得た。次に、リンカー末端ベンジル基 をトリチル基に変更する反応も 3- 1 のように行い、化合物 26 は定量的に、ま た、化合物 27 は 79%収率で得られた。 最後に、3'-, 4'-位に無水コハク酸にてスクシニル化し、CPG 樹脂と結合し、 固相担体 28 が得られ、その活性は 36.7 μmol/g であった。 Fig. 3-3 2'-位フリーのアミノ基での Mitsunobu 反応の生成物と副生成物
3 - 3 3'-末端にグルコサミン誘導体を含む siRNA の合成
今回は Renilla luciferase の配列を選択し、合成した塩基部非置換型、及び塩 基部置換型 2 種類のグルコサミンモノマー誘導体の固相担体 20 と 28 を用いて、 それぞれ DNA/RNA 自動合成機にかけ、3-末端に X, tX, ttX, Y, tY, ttY を有する siRNA を合成した (Fig. 3-4) 。また、天然型である 3-末端に tt チミジン-チ ミジン骨格を有する siRNA も比較用として用意した。
Renilla sense strand: 5'-CUUCUUCGUCGAGACCAUG-3'
Renilla antisense strand: 5'-CAUGGUCUCGACGAAGAAG-3'
合成終了後、常法に従い、合成カラムから樹脂体を取り出し、28 %アンモニ ア水溶液 : エタノール = 3 : 1 混合溶液にて室温で 12 時間反応させ、siRNA の CPG 樹脂からの切り出し及び脱保護を行った。そして、1M TBAF/THF 溶液 にて室温で 12 時間にシリル基の脱保護をした後、反応を 0.1M の TEAA バッ ファ (pH 7.0) で終結させ、Sep-Pak C18 逆相カラムクロマトグラフィーを用い 脱塩を行った。得られた siRNA は、20% PAGE 電気泳動により粗精製した。ま Fig. 3-4 合成された siRNA の 3-末端構造
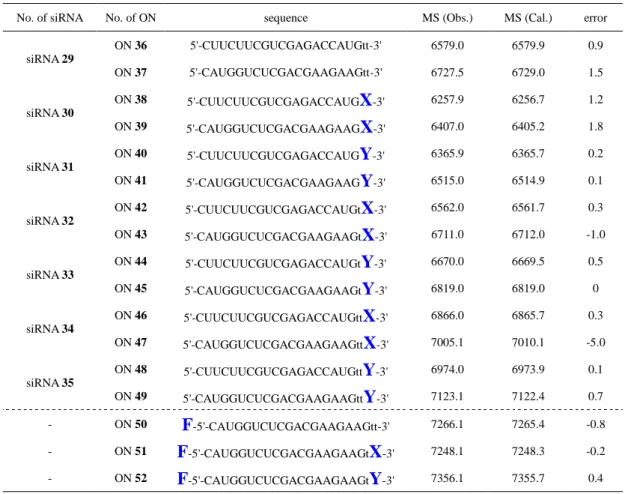
た 、 逆 相 高 速 液 体 ク ロ マ ト グ ラ フ (HPLC: high performance liquid chromatograph) を用いてさらに精製し、MALDI-TOF/MS (matrix-assisted laser desorption / ionization time-of-flight mass spectrometry) により 3'-末端にグルコサ ミンモノマーを有する siRNA 誘導体の分子量を確認した (Table 3-1) (Fig. 3-5)。
Table 3-1 合成した siRNA の配列の確認
No. of siRNA No. of ON sequence MS (Obs.) MS (Cal.) error
siRNA 29 ON 36 5'-CUUCUUCGUCGAGACCAUGtt-3' 6579.0 6579.9 0.9 ON 37 5'-CAUGGUCUCGACGAAGAAGtt-3' 6727.5 6729.0 1.5 siRNA 30 ON 38 5'-CUUCUUCGUCGAGACCAUGX-3' 6257.9 6256.7 1.2 ON 39 5'-CAUGGUCUCGACGAAGAAGX-3' 6407.0 6405.2 1.8 siRNA 31 ON 40 5'-CUUCUUCGUCGAGACCAUGY-3' 6365.9 6365.7 0.2 ON 41 5'-CAUGGUCUCGACGAAGAAGY-3' 6515.0 6514.9 0.1 siRNA 32 ON 42 5'-CUUCUUCGUCGAGACCAUGtX-3' 6562.0 6561.7 0.3 ON 43 5'-CAUGGUCUCGACGAAGAAGtX-3' 6711.0 6712.0 -1.0 siRNA 33 ON 44 5'-CUUCUUCGUCGAGACCAUGtY-3' 6670.0 6669.5 0.5 ON 45 5'-CAUGGUCUCGACGAAGAAGtY-3' 6819.0 6819.0 0 siRNA 34 ON 46 5'-CUUCUUCGUCGAGACCAUGttX-3' 6866.0 6865.7 0.3 ON 47 5'-CAUGGUCUCGACGAAGAAGttX-3' 7005.1 7010.1 -5.0 siRNA 35 ON 48 5'-CUUCUUCGUCGAGACCAUGttY-3' 6974.0 6973.9 0.1 ON 49 5'-CAUGGUCUCGACGAAGAAGttY-3' 7123.1 7122.4 0.7 - - - ON 50 F-5'-CAUGGUCUCGACGAAGAAGtt-3' 7266.1 7265.4 -0.8 ON 51 F-5'-CAUGGUCUCGACGAAGAAGtX-3' 7248.1 7248.3 -0.2 ON 52 F-5'-CAUGGUCUCGACGAAGAAGtY-3' 7356.1 7355.7 0.4
Error = MS (Obs) – MS (Cal)
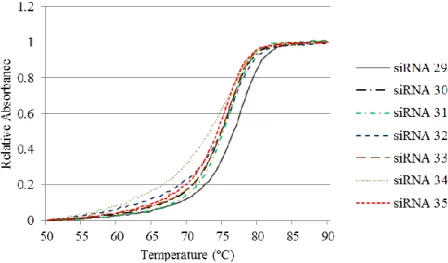
- 31 - 第4 章 3'-末端にグルコサミン誘導体を含む siRNA の生物学的活性評価 4 - 1 siRNA 二本鎖の熱的安定性評価 合成したsiRNA オリゴヌクレオチドのセンス鎖とアンチセンス鎖をそれぞ れ600 pmol を混合させ、測定用緩衝液 (200 μL) に溶解し、アニーリングに より、二本鎖を形成させた。各二本鎖 siRNA の 50% 融解温度 (Tm) を測定 することにより、二本鎖熱的安定性を評価した (Fig. 4-1) (Table 4-1) 。 Table 4-1 siRNA のTm値 No. of siRNA Tm (°C) Tm (°C) siRNA 29 76.1 - siRNA 30 75.4 -0.7 siRNA 31 75.2 -0.9 siRNA 32 75.5 -0.6 siRNA 33 75.1 -1.0 siRNA 34 74.5 -1.6 siRNA 35 74.0 -2.1
Tm = Tm (modified siRNA) -Tm (siRNA
29)
3'-末端ダングリングエンド部位が tt の天然型 siRNA 29 のTm値は76.1°C、 そして、今回合成した 3'-末端にグルコサミンモノマーを有する siRNA 30 ~ 35 はいずれもほぼ同等の値を示しており、大きな差は見られなかった (Tm は最大-2.1°C である) 。すなわち、siRNA に修飾したグルコサミンは二本鎖 熱的安定性に殊んど影響を与えないものと考えられる。
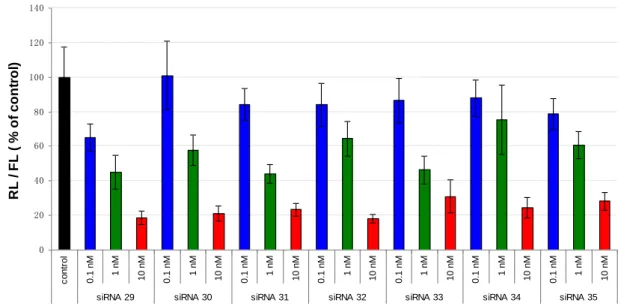
4 - 2 Dual-Luciferase reporter assay によるタンパク発現抑制評価
ヒト子宮頸癌由来の HeLa 細胞を用い、3'-末端にダングリングエンドを持 つsiRNA 29 ~ 35 のタンパク発現抑制能を Dual-Luciferase reporter assay 法 で評価した。本手法は発光タンパクである Firefly Luciferase と Renilla Luciferase の生物発光反応を識別している。なお、今回合成した siRNA は Renilla Luciferase を標的とした配列であるため、Renilla Luciferase の発現 抑制効果をFirefly ルシフェラーゼの発光量と比較し、タンパク抑制能を評価 できる。また、siRNA をトランスフェクションしていない状態を control とし、 100%とした (Fig. 4-2) 。 0 20 40 60 80 100 120 140 c o n tr o l 0 .1 n M 1 n M 1 0 n M 0 .1 n M 1 n M 1 0 n M 0 .1 n M 1 n M 1 0 n M 0 .1 n M 1 n M 1 0 n M 0 .1 n M 1 n M 1 0 n M 0 .1 n M 1 n M 1 0 n M 0 .1 n M 1 n M 1 0 n M
siRNA 29 siRNA 30 siRNA 31 siRNA 32 siRNA 33 siRNA 34 siRNA 35
R L / F L ( % o f c o n tr o l)
結果としては、0.1 nM の低濃度で天然型の siRNA 29 はグルコサミンモノ マーで修飾したsiRNA 30 から siRNA 35 のいずれよりもタンパク質発現抑制 能が優れていた。1 nM の場合は、siRNA 31 と siRNA 33、また、10 nM の場 合は、siRNA 30 と siRNA 32 はほぼ同程度の抑制能を示していた、また Y 修 飾型 siRNA は X 修飾型より優れた (siRNA 31 > siRNA 30、siRNA 33 > siRNA 32、siRNA 35 > siRNA 34) 。しかし、siRNA 34 と siRNA 35 では抑 制能が減少したことが分かった。
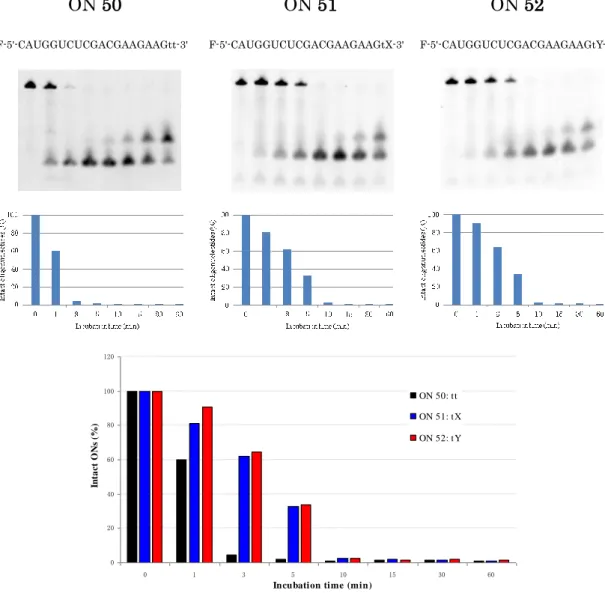
4 - 3 Snake Venom Phosphodiesterase によるヌクレアーゼ耐性評価
天然型のtt ダングリングエンドと同じ長さの tX と tY を選び、5'-末端に蛍 光色素であるフルオレセインを有するON 50、ON 51 および ON 52 を合成し た。 エキソヌクレアーゼの一つであるヘビ毒ホスホジエステラーゼ (SVPD) を 用い、3'-エキソヌクレアーゼに対する耐性を評価した。ON 50、ON 51 及び ON 52 をそれぞれ一本鎖状態で SVPD にて処理した RNA の経時変化を、電 気泳動を行い解析した (Fig. 4-3)。 天然型siRNA は SVPD により、3 分間で 95%以上分解されてしまった。一 方、グルコサミンモノマー修飾型 (tX、tY) のいずれも 60%以上残っており、 5 分間経っても、30%程度残存できることが確認され、修飾型 siRNA (tX、tY) は天然型siRNA (tt) と比べ、はるかに高いヌクレアーゼ耐性を示した。
ON 50 ON 51 ON 52
F-5'-CAUGGUCUCGACGAAGAAGtt-3' F-5'-CAUGGUCUCGACGAAGAAGtX-3' F-5'-CAUGGUCUCGACGAAGAAGtY-3'
0 20 40 60 80 100 120 0 1 3 5 10 15 30 60
Incubation time (min)
In ta c t O N s (%) ON 50: tt ON 51: tX ON 52: tY 4 - 4 細胞膜透過性評価74) 天然型のtt、または tX 及び tY ダングリングエンドをもつ siRNA のセンス 鎖(ON 36、ON 42、ON 44) と 5'-末端に蛍光色素であるフルオレセインを有 するアンチセンス鎖 (ON 50、ON 51、ON 52) をそれぞれ二本鎖にしトラン スフェクション試薬を混合し、HeLa 細胞と 4 時間の培養した後共焦点レーザ ースキャン顕微鏡の観察により、細胞膜透過性を評価した (Fig. 4-4) 。
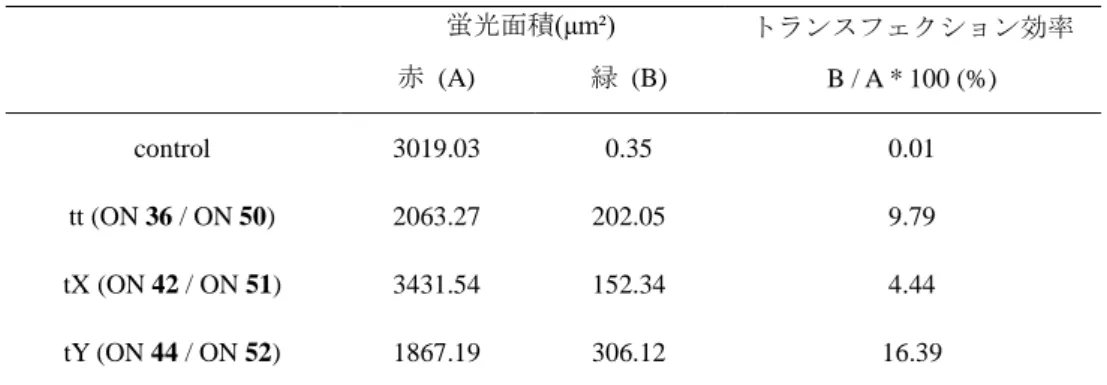
Red (Mito Tracker) Green (Fluorescein) Merge Control 3'-tt siRNA 3'-tX siRNA 3'-tY siRNA 赤色蛍光 (Mito Traker) は染色されたミトコンドリア、緑色蛍光は導入さ れたsiRNA を示している。トランスフェクション効率は付属ソフト Zen2012 により、緑色蛍光面積対赤色蛍光面積の比で評価した (Table 4-2) 。 Fig. 4-4 4 時間インキュベーション後 Hela 細胞の共焦点顕微鏡画像
Table 4-2 siRNA のトランスフェクション効率 蛍光面積(μm²) トランスフェクション効率 B / A * 100 (%) 赤 (A) 緑 (B) control 3019.03 0.35 0.01 tt (ON 36 / ON 50) 2063.27 202.05 9.79 tX (ON 42 / ON 51) 3431.54 152.34 4.44 tY (ON 44 / ON 52) 1867.19 306.12 16.39
天然型siRNA (3'-tt 末端) と比べ、3'-tY 末端を有する siRNA のトランスフ ェクション効率は約1.7 倍に向上した。一方、3'-tX 末端を有する siRNA は天 然型siRNA の半分程度しかなかったことが明らかになった。
第5 章 結語 RNA 干渉は、発見されてから僅か十数年の短い間に、生物学や医学の研究 に大きなインパクトを与え、核酸医薬の新星として世界中で注目されている。 しかし、血中滞留時間が短い、細胞膜透過性が低い、および生体内のヌクレア ーゼによって分解されやすいなどの問題があり、in vitro 実験が成功しても、 in vivo 実験、また臨床上の応用まで至らなかった例が多い。 著者が所属する研究室では、siRNA の 3′-末端ダングリングエンド部位の化 学修飾に着目している。オフターゲット効果の回避とヌクレアーゼ耐性の向上 を目指し、ヌクレオシドの糖部あるいはヌクレオシド全体を芳香族化合物に置 換したもの、もしくは塩基部欠損型ヌクレオシドなど、さまざまなアナログを siRNA の 3′-末端に導入しその機能結果を検証した (Fig. 5-1) 。多くの修飾型 siRNA は天然型 siRNA より優れたヌクレアーゼ耐性と遺伝子抑制能をもつこ とが確認されたが、いずれも細胞膜透過性などの siRNA の DDS の検証には 十分に触れていなかった71-74)。 カチオン性物質キトサンの構成糖であるグルコサミンは、人体に低毒性、高 Fig. 5-1 これまでの siRNA の 3′-末端ダングリングエンド部位の化学修飾 75)
細胞膜透過性などの利点を有するため、本研究では、siRNA の機能向上、か つ、DDS への応用を目指し、天然型グルコサミン X (塩基なし) 、また修飾型 グルコサミン誘導体Y (塩基あり) を siRNA の 3′-末端への導入、それらの化 学的・生物学的機能の検証を行った (Fig. 5-2) 。以下、得られた知見を要約 する。 これまでに、著者の研究室ではグルコサミンモノマーとダイマーを siRNA の 3′-末端へ導入する試みがあったが、従来法で保護されたグルコサミン誘導 体を DNA/RNA 自動合成機によってオリゴ核酸合成した場合、アンモニア処 理による脱保護基ステップが効率的に進まず、3-末端に導入されたグルコサ ミンの保護基はさまざまな構造に変化していた。著者はグルコサミン 2-アミ ノ基の保護基を塩基性条件下で容易に除去できるトリフルオロアセチル基に 変更したところ、後処理の際に副生成物を生じず、効率的にグルコサミン部分 の脱保護が可能となった (第 3 章 3-1) 。 また、グルコサミンをヌクレオシドの糖部と見なし、塩基を導入することで、 塩基を有するグルコサミン誘導体の合成法を開発した。Mitsunobu 反応によ り、1 級水酸基と 2 級水酸基の活性差を利用し、部分的に修飾された塩基を Fig. 5-2 本研究の siRNA の 3′-末端ダングリングエンド部位の化学修飾
グルコサミンの 6-位に導入した (第 3 章 3-2) 。この合成法はチミン以外に、 すべての天然型塩基あるいは人工修飾塩基にも適用可能で、今後さまざまな塩 基を有するグルコサミン誘導体が合成できることを示唆した (Fig. 5-3) 。
ダングリングエンド部位は、RISC の構成タンパク Argonaute の PAZ ドメ インに存在する疎水性ポケットに入り込み、認識されることがすでに解明され た。今回3′-末端ダングリングエンド部位に X と Y を導入した siRNA (X、Y、 tX、tY、ttX、ttY) は、天然型 siRNA (tt) と比べ、遺伝子抑制能の向上が見 られなかったが、いずれも濃度依存的にタンパク質の発現を抑制できる。遺伝 子抑制能が非常に劣っているのはダングリングエンド部位に三塩基突出のttX 型とttY 型であることも分かった。また、1 nM の濃度で塩基部置換型のグル コサミンタイプは塩基部非置換型のグルコサミンタイプより、優れた遺伝子抑 制能を示した (Y>X、tY>tX、ttY>ttX) 。これらの結果から、グルコサミン誘 導体の導入は PAZ ドメインの疎水性ポケットへの認識に障害となり、天然型 より比較的に入り込みにくい、特に天然型の二塩基突出よりダングリングエン ドが長くなる三塩基の場合 (ttX、ttY) は、ダングリングエンドの大きさがさ らにPAZ ドメインへの入り込みに阻害し、RNA 干渉効果が劣化すると考えら Fig. 5-3 塩基部置換型グルコサミン誘導体合成法の確立
れる (Fig. 5-4) 。また、塩基部置換型のグルコサミンタイプは塩基部非置換 型タイプと比べ、天然型のヌクレオシドと構造的に近いため、PAZ ドメイン に比較的に入り込みやすい。すなわち、グルコサミン修飾型siRNA にとって、 塩基の存在は RISC の PAZ ドメインとの認識に関与すると考えられる (第 4 章 4-2) 。
Fig. 5-4 (a) RISC の PAZ ドメインへ 3′-末端 tt 型 siRNA の入り込み
次に、3′-末端ダングリングエンド部位に X と Y の導入にも関わらず、tt 末 端の天然型siRNA と比べ、高いヌクレアーゼ耐性をもっていた。この結果か ら、3'-エキソヌクレアーゼである蛇毒ホスホジエステラーゼ (SVPD) は、3'-末端にグルコサミン誘導体を導入することにより、天然型核酸塩基を優先的に 認識し、オリゴヌクレオチドを切断することが示唆された。また、塩基部置換 型のY 修飾体は塩基部非置換型の X 修飾体より若干優れたヌクレアーゼ耐性 をもつことも確認できた。つまり、塩基部置換型グルコサミン誘導体をsiRNA の3'-末端に導入すると、更なる SVPD との認識を阻害することが明らかとな った (第 4 章 4-3) 。 細胞膜透過性については、ダングリングエンド部位に塩基部非置換型グルコ サミンでの修飾 (3′-tX 末端) は向上が見られなかったが、塩基部置換型グルコ サミンでの修飾 (3′-tY 末端) は天然型より優れたことが確認された。この結果 から、細胞膜の電荷中和などを考慮するカチオン性分子で修飾しても、誘導体 塩基部の欠損は細胞膜透過性に影響を与えることを示した。今回の細胞膜透過 性結果はRNAi 効果の結果 (tY>tX) と一致した。すなわち、細胞膜透過性の
高いsiRNA はより多く細胞内に入り込み、RISC 複合体と結合し RNAi 効果 を現すことに繋がる。また、3′-末端ダングリングエンド部位に塩基部置換型グ ルコサミンモノマーでなく、ダイマーでの修飾 (YY) は細胞膜透過性がさらに 向上することが期待できる (第 4 章 4-4) 。 本研究は、五炭糖のリボースおよび芳香環置換に基づくsiRNA の一般糖 部修飾法の他に、同じく生体内に存在する六炭糖のグルコサミンに着目し、 siRNA の 3'-末端ダングリングエンド部位に化学修飾を行った。修飾型二本鎖 siRNA の熱的安定性が天然型と同程度に保持したまま、遺伝子抑制能の上昇 が見られなかったが、ヌクレアーゼ耐性および細胞膜透過性が予想通り向上し た。また、塩基置換型グルコサミン誘導体の合成法は、6'-位にさまざまな塩基 が導入できることを示した。そこで、siRNA の 3'-末端ダングリングエンド部 位への導入のみならず、リボースの代わりに、グルコサミンを母核としてグリ コシル反応によるsiRNA 分子全鎖をグルコサミンまたはその誘導体で高機能 化することにより、塩基配列の自由に選択できる新規人工核酸の合成も可能と なり、ヌクレアーゼ耐性や細胞膜透過性の更なる向上が期待できる。さらに、 グルコサミン誘導体は人体に毒性が低いと期待されるため、グルコサミン修飾 を用いるDDS 化された siRNA 分子は RNA 創薬に繋がる可能性が非常に高く、 今後臨床的にも意義のある研究テーマであると考えられる。
実験の部
略語
APS ammonium peroxodisulfate
BF3·OEt2 boron trifluoride - ethyl ether complex BF3·OMe2 boron trifluoride - methyl ether complex Boc2O di-tert-butyl dicarbonate
BzCl benzoyl chloride CPG controlled pore glass DEAD diethyl azodicarboxylate DMAP 4-dimethylaminopyridine DMF N,N-dimethylformamide DMSO dimethyl sulfoxide
DMTrCl 4,4′-dimethoxytrityl chloride
EDC-HCl 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride HPLC high performance liquid chromatograph
MALDI-TOF/MS matrix-assisted laser desorption / ionization time-of-flight mass spectrometry
MS 4Å molecular sieves 4Å
NIS N-iodosuccinimide
NMR nuclear magnetic resonance PPh3 triphenylphosphine
SDS-PAGE sodium dodecyl sulfate-polyacrylamide gel electrophoresis SVPD snake venom phosphodiesterase
TBDMSCl tertbutyldimethylsilyl chloride TEA triethylamine
TEAA triethylammonium acetate
TEMED N,N,N′,N′-tetramethylethylenediamine TfOH trifluoromethanesulfonic acid
TIPSCl triisopropylsilyl chloride THF tetrahydrofuran TLC thin-layer chromatography TMSCl trimethylchlorosilane Tris tris(hydroxymethyl)aminomethane 使用機器 1) 測定機器
核酸自動合成機 Applied Biosystems Model 3400 NTS H-6
GC/MS SHIMADZU GCMS-QP 2010A
DART/MS JEOL The AccuTOF Lc-plusJMS-T100LP MALDI/TOF-MS SHIMADZU AXIMA-CFR plus
NMR スペクトル JEOL JNM-AL400
Tm測定機 SHIMADZU UV2400
ルミノ・イメージアナライザー FUJIFILM LAS4000
吸光度計 HITACHI U-2001 spectrophotometer 共焦点レーザースキャン顕微鏡 Carl Zeiss LSM710
TLC Silicagel 60 F254 plate (Merck, Art 5715) 中性シリカゲル Silicagel 60 N (spherical, neutral)
(関東化学; 63-210 mesh) Sep-Pak C18 Waters Corporation HPLC 用逆相カラム YMC J’sphere ODS-M80
3) 各種フィルター
水系 ADVANTEC DISMIC-13JP (PTFE 0.5μm) MILLIPORE MILLEX®-HV
有機系 PALL Acrodisc® Syringe Filter 0.2μm HT Tuffryn® Membrane
水系・有機系 MILLIPORE MILLEX®-LH
4) 使用試薬
〈有機合成用試薬及び溶媒〉
【Aldrich】 20% Pd(OH)2/C, DMTrCl, 1.0M TBAF/THF, Thymine, PPh3
【関東化学】 THF (脱水、安定剤無添加)
【東京化成】 TIPSCl, BzCl, TfOH, NIS, DMAP, EDC-HCl 【ナカライテスク】 Celite, Na2S2O3, THF, TEA, AcOH, DMF*2
【和光純薬】 MeONa (28% in MeOH), Phthalic anhydride, Thiophenol, BF3·OEt2, BF3·OMe2, DEAD, Acetic anhydride, 2-Benzyloxy ethanol, Ethylenediamine, Trifluoroacetic anhydride, DMTrCl, Succinic anhydride, CPG resin, EDC-HCl, Pyridine*1, 脱水 溶媒 (MeCN, MeOH, EtOH)
*1 ピリジンは水素化カルシウムにより蒸留を行い、モレキュラーシーブ 4 Å 共存下保存したものを用いた。
*2 DMF はモレキュラーシーブ 4 Å 共存下で保存したものを用いた。
〈NMR 用重溶媒〉
【和光純薬】 CD3OD, DMSO-d6
【Cambridge Isotope Laboratories】 CDCl3 (TMS 含有)
〈オリゴヌクレオチドの合成及び精製〉
【Aldrich】 1.0M TBAF/THF 【キシダ化学】 Trichloroacetic acid
【Glen research】 各 種 ア ミ ダ イ ト (rA,rG,rC,rU), Fluorescein, Chemical Phosphorylation reagent
【ナカライテスク】 28% Ammonia water, EtOH (99.5%), AcOH, N,N'-Methylene-bisacrylamide, TEA, Urea, acrylamide
【和光純薬】 1
H-Tetrazole, MeCN (核酸合成用、合成 用脱水溶媒), CH2Cl2 (ペプチド合成用), ホ ル ム ア ミ ド (脱 イオ ン 処 理 済 み ), APS, TEMED, EDTA-4Na, ホウ酸
〈生物系実験〉
【promega】 psi-CHECK™-2 vector, Dual-Glo™ Luciferase Assay System, TransFast™ Transfection Reagent
【Wako】 D-MEM (high glucose)
【Invitrogen】 Trypsin-EDTA、MitoTraker Red CMXRos, OPTI-MEM
実験方法
Scheme 1 Initial synthesis route of 6'-abasic glucosamine monomer unit. Reagents and conditions: (a) (i) MeONa, MeOH, rt; (ii) Phthalic anhydride, MeOH, rt; (iii) Acetic anhydride, pyridine, rt, 68%; (b) PhSH, Et2O·BF3, CH2Cl2, rt, 93%; (c) benzyloxyethanol, NIS, TfOH,
CH2Cl2, 4Å MS, rt, 60%; (d) MeONa, MeOH, rt, 85%; (e) TBDMSCl, pyridine, rt, 97%; (f)
BzCl, pyridine, rt, 95%; (g) Pd(OH)2/C, H2, THF, rt, 95%; (h) DMTrCl, pyridine, rt, 88 %; (i)
TBAF, THF, 0ºC, 35%; (j) (i) Succinic anhydride, DMAP, pyridine, rt; (ii) CPG, EDC, DMF, rt, 28 μmol/g.
1-O-(2-Benzyloxyethyl)-3,4,6-tri-O-acetyl-2-deoxy-2-N-phthalimido-β-D-gluco- pyranoside (4) 3 (100 mg, 0.190 μmol) をデシケーターで一晩乾燥後、CH2Cl2 (2 mL) に溶解し、 MS 4Å (100 mg)、 NIS (87 mg, 0.387 μmol)、TfOH (2 μL) を加えアルゴン雰囲 気下室温で 2 時間攪拌した。反応終了後、MS 4Å をセライトでろ過した。ろ 液を CHCl3 (10 mL) にて抽出し、有機層を飽和チオ硫酸ナトリウム水溶液 (10 mL) 、飽和炭酸水素ナトリウム水溶液 (10 mL) 、飽和食塩水 (10 mL) で洗浄 し、無水硫酸ナトリウムで乾燥後、溶媒を減圧留去した。残渣を中性シリカゲ ルカラムクロマトグラフィー (ヘキサン : 酢酸エチル = 3 : 1) で分離精製し、 白色固体 4 (65 mg, 60%) を得た。 1-O-(2-Benzyloxyethyl)-2-deoxy-2-N-phthalimido-β-D-glucopyranoside (5) 4 (1.00 g, 1.76 mmol) をデシケーターで一晩乾燥後、MeOH (30 mL) に溶解し、
MeONa (28% in MeOH) (10.0 mL, 48.9 mmol) を加え室温で 30 分間攪拌した。 反応終了後、AcOH を加え反応を中和し、溶媒を減圧留去した。残渣を中性シ リカゲルカラムクロマトグラフィー (クロロホルム : メタノール= 15 : 1 → 8 : 1) で分離精製し、白色固体 5 (662 mg, 85%) を得た。 1-O-(2-Benzyloxyethyl)-6-O-tert-butyldimethylsilyl-2-deoxy-2-N-phthalimido-β-D-glucopyranoside (6) 5 (400 mg, 902 μmol) をデシケーターで一晩乾燥後、ピリジン (10 mL) に溶解 し、TBDMSCl (273 mg, 1.81 mmol) を 0 °C で滴下しアルゴン雰囲気下室温で 5 時間攪拌した。反応終了後、0 °C で MeOH (25 mL) を加え反応を終結させ、 さらに 1 時間室温で攪拌した。溶媒を減圧留去した後、残渣を中性シリカゲル カラムクロマトグラフィー (クロロホルム : メタノール= 15 : 1) で分離精製 し、無色油状 6 (492 mg, 97%) を得た。
1-O-(2-Benzyloxyethyl)-3,4-di-O-benzoyl-6-O-tert-butyldimethylsilyl-2-deoxy- 2-N-phthalimido-β-D-glucopyranoside (7) 6 (380 mg, 681 μmol) をデシケーターで一晩乾燥後、ピリジン (10 mL) に溶解 し、BzCl (4.62 mL, 39.8 mmol) を 0 °C で滴下しアルゴン雰囲気下室温で 6 時 間攪拌した。反応終了後、0 °C で MeOH (25 mL) を加え反応を終結させた。 溶媒を減圧留去した後、残渣を中性シリカゲルカラムクロマトグラフィー (ヘ キサン : 酢酸エチル = 3 : 1 → 1 : 1) で分離精製し、無色油状 7 (495 mg, 95%) を得た。 3,4-Di-O-benzoyl-6-O-tert-butyldimethylsilyl-2-deoxy-1-O-(2-hydroxyethyl)- 2-N-phthalimido-β-D-glucopyranoside (8) 7 (506 mg, 661 μmol) をデシケーターで一晩乾燥後、THF (20 mL) に溶解し、 20% Pd(OH)2/C (152 mg, 30 wt %)を加え H2雰囲気下室温で 20 分間攪拌した。 反応終了後、Pd(OH)2/C をセライトでろ過した。ろ液をを減圧留去した後、残 渣を中性シリカゲルカラムクロマトグラフィー (クロロホルム : メタノール = 30 : 1 → 25 : 1) で分離精製し、無色油状 8 (425 mg, 95%) を得た。 3,4-Di-O-benzoyl-6-O-tert-butyldimethylsilyl-2-deoxy- 1-O-[2-(4,4'-dimethoxytrityloxy)ethyl]-2-N-phthalimido-β-D-glucopyranose (9) 8 (251 mg, 371 μmol) をデシケーターで一晩乾燥後、ピリジン (10 mL) に溶解 し、DMTrCl (160 mg, 475 μmol) を加えアルゴン雰囲気下室温で 6 時間攪拌し た。反応終了後、MeOH (10 mL) を加え反応を終結させ、溶媒を減圧留去した。 残渣を中性シリカゲルカラムクロマトグラフィー (数滴ピリジンを有するヘ キサン : 酢酸エチル = 3 : 1 → 1 : 1) で分離精製し、淡黄色油状 9 (318 mg, 88%) を得た。
3,4-Di-O-benzoyl-2-deoxy-1-O-[2-(4,4'-dimethoxytrityloxy)ethyl]- 2-N-phthalimido-β-D-glucopyranose (10) 9 (476 mg, 487 μmol) をデシケーターで一晩乾燥後、THF (25 mL) に溶解し、 TBAF (625 μg, 625 μmol) を加えアルゴン雰囲気下 0 °C で 3 時間攪拌した。反 応終了後、溶媒を減圧留去した。残渣を中性シリカゲルカラムクロマトグラフ ィー (数滴ピリジンを有するヘキサン : 酢酸エチル = 3 : 1 → 1 : 1) で分離精 製し、淡黄色油状 10 (145 mg, 35%) を得た。 Solid support (11) 10 (213 mg, 247 μmol) をデシケーターで一晩乾燥後、ピリジン (4 mL) に溶解 し、無水コハク酸 (128 mg, 1.28 mmol)、DMAP (3 mg, 25 μmol) を加えアルゴ ン雰囲気下室温で 36 時間攪拌した。反応終了後、反応液を酢酸エチルと水に て抽出し、有機層を減圧除去した後、精製せず、一晩乾燥した。その後、アル ゴン雰囲気下粗生成物を DMF (6.2 mL) に溶解し、CPG 樹脂 (522 mg, 62 μmol) を加え樹脂がなじむように軽くナスフラスコを振とうさせ、EDC (48 mg, 247 μmol) を加え、室温で 3 日間振とうした。カップリング反応終了後、ガラスフ ィルターで溶媒を減圧除去し、ピリジンで樹脂を洗浄した。樹脂を集めたナス フラスコに 0.1 M DMAP 溶液 (ピリジン 13.5 mL、 無水酢酸 1.5 mL、 DMAP 183 mg) を加え、アルゴン雰囲気下、さらに室温で 2 日間振とうした。 キャッピング反応終了後、ガラスフィルターで溶媒を減圧除去しピリジン、 EtOH、MeCN で洗浄しデシケーターで乾燥し、CPG 樹脂固層担体 11 (455 mg) を得た。11 (6 mg) を測り取り、過塩素酸溶液 (HClO4 : EtOH = 3 : 2)で脱保護 し、その溶液が示す波長 498 nm でのトリチル基の吸光度から、以下の式を用 いて算出した。その結果、CPG 樹脂固層担体 11 の活性は 28.4 μmol/g であった。
Scheme 2. Synthesis route of 6'-abasic glucosamine monomer unit. Reagents and conditions: (a) (i) MeONa, MeOH, rt; (ii) Phthalic anhydride, MeOH, rt; (iii) Acetic anhydride, pyridine, rt, 68%; (b) PhSH, Et2O·BF3, CH2Cl2, rt, 93%; (c) Ethylenediamine, MeCN, reflux, 60%; (d)
TIPSCl, pyridine, rt, 86%; (e) (CF3CO)2O, pyridine, rt, 79%; (f) BzCl, pyridine, rt, 83%; (g)
benzyloxyethanol, NIS, TfOH, CH2Cl2, 4Å MS, rt, 78%; (h) Pd(OH)2/C, H2, THF, rt, 89%; (i)
DMTrCl, pyridine, rt, 98%; (j) TBAF, THF, rt, 68%; (k) (i) Succinic anhydride, DMAP, pyridine, rt; (ii) CPG, EDC, DMF, rt, 19 μmol/g.
Absorbance (498 nm) × Volume (HClO4) × 14.3 (ε value of DMTr)
Weight (CPG resin) CPG activity =
2-Deoxy-2-phthalimino-1,3,4,6-tetra-O-acetyl-β-D-glucopyranoside76) (2)
1 (10.0 g, 46.4 mmol) をデシケーターで一晩乾燥後、MeOH (500 mL) に溶解し、
MeONa (28% in MeOH) (11.4 mL, 55.7 mmol) を加え室温で 3 時間攪拌した。そ の後、無水フタル酸 (13.7 g, 92.8 mmol) を加え、さらに室温で 3 時間攪拌した。 反応終了後、溶媒を減圧留去し、残留物を一晩乾燥した。翌朝粗生成物をピリ ジン (400 mL) に溶解し、無水酢酸 (65.8 mL, 696 mmol) を加えアルゴン雰囲 気下室温で 12 時間攪拌した。反応終了後、0 °C で MeOH (200 mL) をゆっく り加え溶媒を減圧留去した。残渣を中性シリカゲルカラムクロマトグラフィー (ヘキサン : 酢酸エチル = 5 : 2) で分離精製し、無色固体 2 (15.1 g, 68%) を得 た。
Phenyl 2-deoxy-2-phthalimino-3,4,6-tri-O-acetyl-1-thio-β-D-glucopyranoside77)
(3)
2 (6.01 g, 12.6 mmol) をデシケーターで一晩乾燥後、CH2Cl2 (60 mL) に溶解し、
thiophenol (1.32 mL 12.6 mmol) 、BF3·OEt2 (10.0 mL, 37.8 mmol) を加えアルゴン 雰囲気下室温で 20 時間攪拌した。反応終了後、TEA を加え反応を中和し、CHCl3 (50 mL) と飽和食塩水 (50 mL) にて抽出した。その後、有機層を無水硫酸ナト リウムで乾燥後、溶媒を減圧留去した。残渣を中性シリカゲルカラムクロマト グラフィー (ヘキサン : 酢酸エチル = 5 : 2 → 3 : 1) で分離精製し、無色固体 3 (6.20 g, 93%) を得た。
Phenyl 2-amino-2-deoxy-1-thio-β-D-glucopyranoside78) (12)
3 (1.16 g, 2.20 mmol) をデシケーターで一晩乾燥後、MeCN (10 mL) に溶解し、
エチレンジアミン (8.90 mL, 132 mmol) を加えアルゴン雰囲気下 90 °C で 22 時 間還流した。反応終了後、反応温度を室温まで戻し、溶媒を減圧留去した。残 渣を中性シリカゲルカラムクロマトグラフィー (酢酸エチル: メタノール :