論文
琵琶湖周辺内湖の植物プランクトン増殖に
対する栄養評価
三田村 緒佐武
1、尾崎 正樹
2、後藤 直成
3Evaluation of Nutrient Deficiency for Phytoplankton Growth
in Lagoons connected to Lake Biwa
Osamu MITAMURA
1, Masaki OZAKI
2and Naoshige GOTO
31. Faculty of Education, Shiga University 2. Dear departed
3. School of Environmental Sciences, University of Shiga Prefecture
To elucidate deficiencies in the nutrients needed for phytoplankton growth, nutrient limits were examined in 24 lagoons, SATOMIZU "Naiko", connected to Lake Biwa. The evaluation parameters consisted of particulate bioelements (particulate carbon, nitrogen and phosphorus), chlorophyll-a, and nutrient (nitrogenous and phosphorus compound) concentrations and their ratios. The phytoplankton in almost all lagoons was not limited by either nitrogen or phosphorous compounds. However, phytoplankton growth in some shallow lagoons with low phytoplankton concentrations was affected by phosphorus nutrients. All lakes throughout the watershed were identified as eutrophic based on total phosphorus concentration as the trophic state index. Furthermore, six lagoons were identified as oligotrophic or mesotrophic lakes, and the other 18 lagoons were identified as eutrophic lakes, judging from chlorophyll-a concentration indicators.
Keywords: nitrogen and phosphorus deficiency, trophic state index, phytoplankton, SATOMIZU, lagoon “Naiko”, Lake Biwa 1滋賀大学教育学部 2故人 3滋賀県立大学環境科学部
1 はじめに
琵琶湖周辺には内湖と称される潟湖が 1940 には 37 内湖 (2,900 ha)あったが、食料不足解消の田畑に転用するた めに干拓され、現存は 23 内湖(530 ha)にすぎない。そ れまで内湖は、比較的自然潟湖の生態学的景観であったが、 高度成長期を経て人間生活の影響を多大に受けるようにな り、三田村ら(2014)が指摘する 2 次自然景観の「里水」 内湖に変貌した。内湖は、湖内で生息・生育する水生生物 のみならず内湖生態系を利用する生物が多様である(中西・ 関 野,1997; 山 田・ 野 崎,1997;Nagoshi, 1965;Hoyerand Canfield, 1994;Mehner et al., 2005;西野,2005;浜端, 2010)。倉田(1984, 1988)は、琵琶湖周辺内湖が、水質浄 化機能、生態機能、治水機能、利水機能、観光機能、景観 機能、水産機能、生活機能を有しているとした。湖水中の 生 元 素 化 合 物(Miyake et al., 2002; 三 田 村・ 後 藤, 2005)、懸濁有機物(大久保,1998)、および植物プランク トンによる基礎生産(三田村・後藤,2005;Mitamura et al., 2009)に関する研究成果から、琵琶湖内湖は生物地球 化学的循環特性を有することが報告されている。
おいて溶存無機全窒素とリン酸態リン現存量の比(DIN: DIP 比)を Redfield(1958)が報告した通称 Redfield 比と 比較することにより、植物プランクトン増殖を制限する要 因を判断する指標を提唱し、DIN:DIP 比(質量比)が 5 以 下 は 窒 素 律 速、12 以 上 は リ ン 律 速 で あ る と し た。 Tezuka(1984)は、湖水の窒素およびリン栄養塩化合物 の現存量比から、琵琶湖では植物プランクトン増殖にリン が制限因子として働いていると判断した。一方、Healey and Hendzel(1980)は、懸濁態炭素(PC)、懸濁態窒素(PN) および懸濁態リン(PP)の比から植物プランクトン増殖 に 対 す る 栄 養 制 限 要 因 の 指 標 を 提 唱 し た。 そ こ で、 Tezuka(1985) と Nakanishi et al.(1990) は、 湖 水 の PC:PN:PP 比から琵琶湖の植物プランクトンに対する 栄養制限因子を評価した。Aizaki and Otsuki(1987)は、 水深が浅い富栄養化した霞ヶ浦において、PC、PN、およ び PP 現存量にクロロフィルa(Chl.a)現存量を加えたこ れらの比を指標として植物プランクトンに対する栄養制限 因子を評価した。
Vollenweider and Kerekes(1980)は、植物プランクト ン現存量は湖水中の全窒素と全リン現存量との間に正の相 関があり、この関係は湖沼の富栄養化対策管理の基礎に使 用できると報告した。さらに OECD(1982)は、年平均 の全リン、全窒素および Chl.a 現存量から湖沼の栄養段階 の解析基準を提唱した。Sakamoto(1966)、Carlson(1977)、 Forsberg and Riding(1980)は、DIP 現存量レベルから 湖沼の栄養段階を評定したが、彼らが用いた栄養段階の判 定基準の現存量のレベルは異なる。 しかし、これまで琵琶湖周辺「里水」内湖のすべてを対 象として、これら懸濁態生元素および溶存栄養塩化合物の 現存量を詳細に解析し、内湖の植物プランクトン増殖に対 する栄養制限要因を研究した報告はない。本研究は、溶存 窒素栄養塩化合物とリン栄養塩化合物、懸濁態炭素、懸濁 態窒素、懸濁態リン、クロロフィルaの現存量、およびこ れらパラメーター間との関係の分布変動から、植物プラン クトンの基礎生産に対する制限栄養塩因子を探ることを目 的とした。さらに本研究は、湖水の全リンとクロロフィル a 現存量から内湖の栄養段階を評定し、琵琶湖周辺内湖の 富栄養化の現況を調べ、植物プランクトン増殖に対する栄 養制限との関わりを明らかにすることを試みた。
2 方 法
2-1 研究対象内湖 調査を行なった琵琶湖周辺内湖を図 1 に示す。調査研究 対象とした内湖は、現存すべての 24 内湖である。 図 1 1940 年 に 確 認 さ れ た 琵 琶 湖 周 辺 内 湖( 倉 田, 1984). 現存の堅田内湖(1)、近江舞子内湖(2)、乙女ヶ 池(3)、松ノ木内湖(4)、五反田沼(6)、十ヶ坪沼(7)、 菅沼(8)、浜分沼(10)、貫川内湖:南(11)、貫川内湖: 北(11)、野田沼:湖北(14)、南浦内湖(16)、細江内湖 (17)、蓮池(18)、野田沼:彦根(21)、曽根沼(22)、神 上沼(24)、伊庭内湖(25)、西の湖:西(28)、西の湖: 東(28)、北の庄沢(29)、北沢沼(31)、志那中湖(35)、 柳平湖(36)、および平湖(37)を調査対象とした.黒く 塗りつぶして記したものは干拓により消失した内湖. 図 1 に白抜きで記した堅田内湖(1)、近江舞子内湖(2)、 乙女ヶ池(3)、松ノ木内湖(4)、五反田沼(6)、十ヶ坪沼 (7)、菅沼(8)、浜分沼(10)、貫川内湖:南(11)、貫川 内湖:北(11)、野田沼:湖北(14)、南浦内湖(16)、細 江内湖(17)、蓮池(18)、野田沼:彦根(21)、曽根沼(22)、 神上沼(24)、伊庭内湖(25)、西の湖:西(28)、西の湖: 東(28)、北の庄沢(29)、北沢沼(31)、志那中湖(35)、柳平湖(36)、および平湖(37)が現存内湖である(カッ コ内の数字は図中に記載した番号と対応する)。なお、図 中に黒く塗りつぶして記したものは、現在は消失した内湖 である。かつて大中の湖(図 1 の 26)は広大な面積を誇っ ていたが、現在は干拓されて消失した。現存する琵琶湖内 湖は、狭い面積の五反田沼(1.2 km2)から比較的広い面 積を有する西の湖(221.9 km2)までさまざまである(Miyake et al., 2002)。なお、本研究対象とした 24 内湖の中には貫 川内湖のように小水路で分割された内湖もある。この両内 湖を同じ内湖として名称を用いることがあるが、本研究で は両内湖を貫川内湖(北)と貫川内湖(南)とした。した がって、一般に現存内湖数は 23 と称されるが(倉田、 1984)、本研究の琵琶湖周辺の現存内湖数は 24 になる。さ らに、最大内湖の西の湖は、内湖の東部定点と西部定点(こ の定点は、琵琶湖への流出河川の長命寺川に近いため、西 の湖水質の全平均値を示すと考えられる)を設定した。内 湖景観は、周辺の土地利用や内湖利用の影響を受け、さま ざまな特徴がある。例えば、野田沼(彦根)、曽根沼、伊 庭内湖のように一部分が干拓された内湖、五反田沼、十ヶ 坪沼、菅沼のように緑地公園整備が進行した内湖、そして 堅田内湖のように水産物の養殖に利用されている内湖など である。 2-2 湖水の採取と測定方法 調査観測は、2004 年 6 月、8 月、10 月、そして 12 月の 4 回行なった。内湖の開水面水域を見渡せる湖岸から、ロー プ付きプラスチックペールを用いて数 m 沖の表面水を採 取 し た。 採 取 し た 試 水 の 水 温(WT) は 電 気 温 度 計 (Tohodentan type ET-5)、透視度(Tr)は 1 m の透視度計、
濁度(Turb)は濁度計(HACH 2100N)、電気伝導度(EC) はポータブル電気伝導度計(Yokogawa model SC-51)、そ して pH はガラス電極法(Yokogawa PH-82)を用いてそ れぞれ測定した。 現場から持ち帰った試水は、420℃で燃焼処理したガラ ス繊維ろ紙(Whatman GF/F)でただちにろ過した。ろ 紙は懸濁態生元素(懸濁態炭素;PC、懸濁態窒素;PN、 懸濁態リン;PP)、およびクロロフィルa(Chl.a)の分析 に、ろ液は栄養塩化合物(アンモニア態窒素;NH4+-N、 亜硝酸態窒素;NO2--N、硝酸態窒素;NO3--N、リン酸態 リ ン;PO43--P)、 お よ び 溶 存 有 機 物( 溶 存 有 機 炭 素; DOC、溶存有機リン;DOP)の分析に供した。なお、ろ 紙およびろ液は、実験室における化学分析まで-20℃で凍 結保存した。ケイ酸態ケイ素(DSi)測定のための試水は、 紙ろ紙(Advantec No.5C)でろ過してろ液を分析まで冷 蔵庫で冷却保存した。NH4+-N はインドフェノール法(Sagi, 1966)、NO2--N は BR 法(Bendschneider and Robinson, 1952)、NO3--N は 硫 酸 ヒ ド ラ ジ ニ ウ ム 法(Mitamura, 1997)で測定した。PO43--P(DIP)はアスコルビン酸還 元によるモリブデン青法(Murphy and Riley, 1962)、DSi はモリブデン黄法(Mullin and Riley, 1955)で測定した。 DOC は全有機炭素計(Shimadzu TOC-5000A)を用い、 DOP は Menzel and Corwin(1965)によるペルオキソ二 硫酸カリウム分解法で測定した。PC と PN は CHN コー ダ ー(Yanaco MT-5 type) を 用 い て 測 定 し た。PP は Menzel and Corwin(1965)に従った。Chl.a 現存量はろ 紙上の色素を 90%アセトンで抽出し、蛍光光度計(Turner 10-AU)で測定した。
3 結果と考察
3-1 内湖における生元素化合物の分布と懸濁物の特徴 琵琶湖周辺 24 内湖 25 定点において測定した水質パラ メーター(WT、Tr、Turb、EC、pH、NH4+-N、NO2--N、 NO3--N、DIP、DSi、DOC、PC、PN、PP、お よ び Chl.a 現存量)の分布変動を表 1 に示す。なお表は、それぞれの パラメーターの 6 月、8 月、10 月、12 月に得られた値か ら算出された平均値、標準偏差、および変動計数を示して いる。水温は、12 月に低いが他の季節は比較的高かった。 Tr ならびに Turb から判断すると、内湖水は比較的濁水 に近いといえる。そしてそれぞれの内湖ごとに、また季節 ごとに水中浮遊懸濁量は変動していた。EC は、琵琶湖北 湖の西岸に近い内湖で低く、東岸に位置する内湖は高い値 であった。この分布特徴は、Miyake et al.(2002)が指摘 したように、内湖集水域の地質が反映した結果であると考 えた。琵琶湖南湖東岸に位置する柳平湖と平湖は植物生産 活性が高いために pH が高く、北湖西岸域の内湖は低い傾 向があるため活性が低いと考えられた。なお、北沢沼の pH は変動が激しかった。内湖における現存量の多さから 判断すると富栄養化関連栄養塩化合物として機能しないと 考えられる DSi は、EC の分布変動と異なり、内湖ごとに また季節ごとに比較的変動は小さく安定したパラメーター であると考えられた。なお、現存量が低い浜分沼や五反田 沼などにおいては、DSi は珪藻の栄養源として考慮する必 要がある。DOC と DOP 現存量は、東岸に位置する内湖 で高く集水域などからの汚濁負荷の影響を受けていると考表 1 琵琶湖周辺 24 内湖 25 定点における水温(WT : ℃) 、透視度(Tr : cm) 、濁度(Turb : NTU) 、25℃における電気伝導度(EC : mS m -1 )、pH、溶存無機全窒素(DIN : µM )、アンモニア態窒素 ( N H4 -N:µM) 、 亜 硝 酸 態 窒 素(NO 2 -N:µM) 、 硝 酸 態 窒 素(NO 3 -N:µM) 、 リ ン 酸 態 リ ン(DIP:µM) 、 ケ イ 酸 態 ケ イ 素(DSi:µM) 、 溶 存 有 機 炭 素(DOC:µM) 、 懸 濁 態 炭 素(PC:µM) 、 懸 濁 態 窒 素(PN:µM) 、 懸 濁 態 リ ン(PP:µM) 、 お よ び ク ロ ロ フ ィ ル a(Chl.a:mg chl.a m -3 )現存量 .平均値 ( A V )、 標準偏差(SD) 、変動計数(CV:%)は 4 試料から算出. 堅田内湖 近江舞子 乙女ヶ池 松ノ木 五反田沼 十ヶ坪沼 菅沼 浜分沼 貫川内湖 貫川内湖 野田沼 南浦内湖 細江内湖 蓮池 野田沼 曽根沼 神上沼 伊庭内湖 西の湖 西の湖 北の庄沢 北沢沼 志那中湖 柳平湖 平湖 内湖 内湖 (南) (北) (湖北) (彦根) (西) (東) WT AV 20.9 20.4 20.1 18.6 21.6 19.8 20.7 19.9 21.1 21.2 21.8 21.6 20.0 22.0 20.1 21.4 19.8 20.8 20.6 20.5 20.3 20.7 21.6 22.1 21.8 SD 7.9 7.3 7.0 7.5 8.7 6.6 7.3 6.7 7.8 8.3 7.6 9.0 7.1 8.5 6.8 8.2 5.8 7.1 7.8 7.2 6.7 8.6 7.5 8.3 8.5 CV 38 36 35 41 40 33 35 34 37 39 35 42 35 39 34 38 29 34 38 35 33 42 35 38 39 Tr AV 38 65 42 74 53 62 78 82 62 43 37 51 84 48 59 70 40 64 58 61 36 50 35 37 20 SD 8 20 15 31 16 30 6 21 19 4 11 16 20 23 29 18 14 25 15 31 28 14 6 13 11 CV 21 31 37 42 31 48 8 26 31 10 31 31 24 48 48 26 35 39 27 51 78 28 17 36 54 Turb AV 12.1 4.1 6.4 5.3 5.5 11.1 5.1 2.0 7.9 7.4 7.8 8.3 4.4 7.7 6.1 3.7 13.0 5.6 5.6 5.1 10.2 9.5 11.4 9.8 25.7 SD 5.4 0.6 3.2 2.0 1.4 4.1 1.5 0.6 5.7 2.8 2.5 1.1 1.6 1.5 3.4 1.1 6.9 2.7 1.4 2.3 4.4 6.2 1.4 2.4 17.6 CV 45 15 51 38 25 37 30 32 72 37 32 14 37 20 56 30 53 49 26 44 43 65 12 24 68 EC AV 15.8 8.5 10.6 9.9 9.8 7.7 9.6 9.8 9.8 11.4 14.9 20.1 18.4 24.5 25.8 20.7 25.3 15.6 21.1 20.7 23.3 13.8 24.6 22.1 18.9 SD 1.8 1.0 0.6 0.2 1.4 0.5 1.0 0.4 0.3 0.4 0.9 4.5 0.7 4.3 2.5 1.3 2.0 2.5 2.5 4.2 2.1 1.2 3.4 3.7 2.7 CV 12 12 5 2 14 6 11 4 3 3 6 22 4 18 10 6 8 16 12 20 9 9 14 17 14 pH AV 7.7 7.9 7.6 7.1 7.2 7.1 7.2 7.3 7.2 7.2 8.4 7.4 7.6 7.3 7.7 7.7 7.5 7.9 7.7 7.9 7.2 6.7 7.3 8.6 8.7 SD 0.5 1.0 0.5 0.2 0.5 0.2 0.1 0.2 0.1 0.1 0.9 0.4 0.1 0.2 0.3 0.3 0.4 0.5 0.3 1.1 0.0 1.7 0.2 0.8 0.8 CV 7 13 6 3 7 3 2 3 2 1 11 6 2 3 4 4 6 6 4 14 1 25 2 10 9 DIN AV 36 16 3 42 5 20 32 40 72 51 25 13 46 54 91 13 59 61 91 146 125 57 62 11 14 SD 23 24 2 7 5 11 16 15 34 34 27 16 15 24 40 12 46 47 38 67 97 67 55 13 16 CV 64 156 75 16 100 56 52 39 46 67 110 123 34 44 43 94 78 77 42 46 78 117 89 122 119 NH 4 AV 6.8 5.0 0.6 7.5 0.7 2.4 5.4 2.4 2.7 1.6 0.9 4.7 7.0 6.6 4.1 5.5 17.8 6.4 4.5 4.4 11.8 6.2 4.6 2.8 5.0 SD 7.6 8.5 0.3 2.2 0.2 1.5 4.0 2.1 2.4 1.7 0.9 6.0 2.7 2.2 2.0 7.0 10.9 3.9 3.1 3.1 8.8 6.7 4.0 2.3 5.7 CV 112 170 50 30 33 63 73 86 88 106 99 127 38 34 49 126 61 61 69 72 74 109 87 83 116 NO 2 AV 1.2 0.4 0.1 0.7 0.3 0.3 0.5 0.3 0.5 0.5 0.5 1.0 0.8 1.4 3.0 0.5 3.0 1.1 1.1 1.8 2.5 1.3 1.2 0.5 0.4 SD 0.5 0.4 0.1 0.1 0.3 0.1 0.1 0.0 0.2 0.1 0.5 1.7 0.2 0.5 0.9 0.4 2.9 0.2 0.5 1.0 1.6 1.4 1.0 0.8 0.4 CV 44 97 94 22 103 48 26 14 32 23 84 171 30 36 31 88 97 23 46 53 66 108 79 145 102 NO 3 AV 27 10 2 34 4 17 26 37 69 49 23 7 38 46 84 7 38 53 85 140 111 50 56 7 8 SD 21 16 2 8 5 10 12 16 31 33 26 8 17 25 40 7 33 46 39 70 91 61 53 10 10 CV 76 151 88 24 113 61 48 45 45 68 114 115 45 56 47 104 88 86 46 50 82 122 95 146 122 DIP AV 0.37 0.68 0.56 0.98 0.75 0.42 0.88 0.46 0.28 0.30 1.37 0.32 0.67 0.55 0.62 0.36 1.51 0.72 0.33 0.45 0.49 0.70 1.63 0.33 3.44 SD 0.26 0.29 0.25 0.25 0.65 0.25 0.22 0.15 0.08 0.12 0.61 0.10 0.35 0.16 0.19 0.16 1.33 0.21 0.25 0.29 0.17 0.71 0.93 0.20 4.67 CV 72 42 44 26 87 61 25 32 29 41 45 30 52 28 30 45 88 30 75 63 34 102 57 61 136 DSi AV 200 251 245 167 116 157 153 91 162 164 229 159 201 165 206 193 399 234 215 296 245 153 309 160 124 SD 33 44 193 14 22 14 18 11 16 12 50 45 25 72 43 13 46 11 47 27 171 130 118 44 68 CV 16 17 79 8 19 9 12 13 10 7 22 29 13 44 21 7 12 5 22 9 70 85 38 28 55 DOC AV 229 253 178 104 261 135 119 126 112 151 182 255 121 200 154 392 159 148 216 227 224 234 273 368 479 SD 53 83 57 38 110 84 49 60 60 61 96 56 72 97 68 93 59 70 59 46 90 80 57 70 146 CV 23 33 32 36 42 63 41 48 54 40 53 22 59 48 44 24 37 48 27 20 40 34 21 19 30 PC AV 126 142 150 71 159 212 50 94 55 124 155 179 27 62 107 114 122 97 47 91 123 113 47 147 241 SD 57 114 102 72 61 215 20 78 23 33 46 116 11 24 65 48 39 68 15 59 70 36 6 68 27 CV 45 81 68 101 38 101 40 83 42 26 29 65 41 39 61 42 32 70 31 65 57 32 14 46 11 PN AV 21.2 23.0 21.2 12.4 23.7 32.0 8.1 14.2 8.5 21.7 20.9 22.1 4.8 9.2 17.5 16.4 21.2 18.9 8.8 13.7 19.0 16.9 7.4 20.1 34.1 SD 11.8 17.8 10.4 11.6 8.7 32.0 3.6 11.3 4.6 3.1 5.4 9.3 2.8 3.7 10.8 4.8 7.2 16.2 3.6 6.9 11.3 6.7 1.2 9.2 3.1 CV 56 77 49 93 37 100 45 80 54 14 26 42 58 40 62 29 34 86 41 50 59 39 16 46 9 PP AV 1.66 1.39 1.97 2.20 2.61 2.56 1.66 0.96 0.95 2.15 1.80 2.00 1.72 1.98 1.98 0.98 3.40 1.19 0.72 1.16 2.37 1.84 2.78 1.62 2.18 SD 0.63 0.69 1.04 1.18 1.25 1.37 0.46 0.62 0.24 0.79 0.46 0.53 0.39 0.66 0.89 0.35 0.94 0.36 0.28 0.40 0.80 1.28 0.84 0.67 0.30 CV 38 50 53 54 48 53 28 64 26 37 26 27 23 33 45 36 28 31 38 34 34 70 30 41 14 Chl.a AV 23.1 56.9 28.6 13.3 23.2 96.6 5.2 25.4 4.0 21.6 30.8 29.2 1.9 6.1 9.3 11.7 16.2 18.7 7.2 13.8 14.8 15.4 2.1 17.8 31.0 SD 16.0 89.8 33.5 20.3 20.2 134.9 4.1 35.9 3.9 12.8 25.6 11.8 1.3 6.3 4.6 5.2 12.1 23.4 4.3 12.9 16.3 8.3 1.4 10.7 14.3 CV 69 158 117 152 87 140 78 141 98 59 83 40 68 104 49 45 74 126 59 93 110 54 65 60 46
えられた。それぞれの内湖ごと季節ごとの変動は比較的小 さかった。
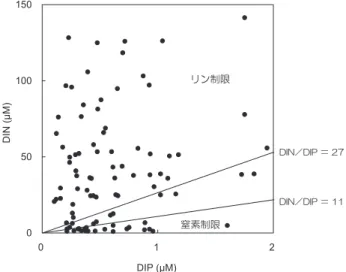
溶存無機全窒素(DIN:NH4+-N、NO2--N、NO3--N の 合計)、NH4+-N、NO2--N、NO3--N および DIP 現存量は、 琵琶湖北湖東岸と南湖岸に位置する内湖で高く、北湖西岸 内湖で低い傾向があった。人間生活による内湖集水域から の流入負荷による影響を受けた結果であると考えた。今後、 集水域の土地利用や内湖流入水の汚濁処理などの実態を把 握する必要がある。これら窒素栄養塩化合物の現存量は夏 季に低く冬季に高くなる傾向を示したが、内湖ごとに大き く異なり、また季節変動も激しかった。一方、DIP 現存量 は、各内湖において明らかな季節変化の傾向を示さなかっ た。高い現存量の PC、PN が、十ヶ坪沼と平湖で観測され、 細江内湖の約 10 倍現存していた。そして、夏季に高く冬 季に低くなる季節変化を示した。一方、PP 現存量は PC と PN の分布と異なった変動パターンを示した。これら懸 濁態生元素は、窒素栄養塩化合物やリン栄養塩化合物のよ うに激しく変動しないが、それぞれの内湖で、そして季節 によってもかなり異なっていた。Chl.a 現存量は、懸濁態 生元素の現存量と同様に夏季に高く、冬季に低い季節変化 傾向を示したが、内湖ごとに大きく変動していた。とくに 十ヶ坪沼の Chl.a 現存量が他の内湖で PC と PN から想定 されるより高いことから理解されるように(表 1)、植物 プランクトン現存量は、それぞれの内湖ごとに、そして季 節ごとに大きく異なっていたことを示す。 本研究水域とした水深が浅い琵琶湖周辺の「里水」内湖 が、いかなる水環境にあるのかを水中懸濁物の特徴から明 らかにしようと試みた。水域に生息する植物プランクトン 細胞の PC、PN、PP 量と Chl.a 量の比は、植物プランク トンの生理状態や種によって異なることがよく知られてお り、一般に PC:Chl.a が 30 ~ 100、PN:Chl.a が 5 ~ 15、 PP:Chl.a が 2 ~ 4 の値を得ることが報告されている(例; 西條・三田村,1995)。琵琶湖周辺内湖において PC:Chl.a、 PN:Chl.a、PP:Chl.a 比を計算した結果、一般に得られ る植物プランクトンの比より高い値が半数近くの内湖で得 られた。これは、琵琶湖内湖水は、植物プランクトン以外 のデトリタス状の有機物を多量に含んでいたことを示唆し ている。一方、比較的高い Chl.a 現存量が測定された内湖 では、これらは低い比の値が計算され、この内湖水の懸濁 物のほとんどは、植物プランクトンで構成されていたと考 えた。このことは、夏季と秋季に十ヶ坪沼と秋季の近江舞 子沼できわめて高い Chl.a 現存量が得られ、それに対応し たきわめて低い PC:Chl.a 比が後述で示す表 2 からも理解 される。 これらのことは、一般に水深が浅い琵琶湖周辺内湖の水 中懸濁有機物は、植物プランクトン以外の物質を多量に含 むことを示唆している。この原因として、内湖で生産され た沈積堆積物が波風により湖底から再懸濁したことなどが 考えられる。また、自然あるいは人間活動による内湖集水 域からの有機懸濁物の流入負荷も考慮しなければならな い。なお、本研究では、水中 Chl.a 現存量を植物プランク トン現存量の指標としたが、大型水生植物の破片あるいは 付着藻類の剥離色素量の一部が測定されていたことは言う までもない。 3-2 溶存無機窒素と溶存無機リンの比および懸濁物質の生 元素比から判定した植物プランクトン増殖の栄養制限因子 本観測期間に得られた DIN:DIP 比は、冬季に高くな る傾向が認められた。これは、DIP 現存量が明らかな季節 変化を示さなかったが、DIN 現存量が冬季に高くなった ことによる。これらのことから、上述したように内湖の栄 養塩化合物現存量は内湖ごとに、また季節によっても大き く異なり、水中の構成栄養塩は変動していると考えられた。 したがって、内湖水の栄養塩化合物は、湖内で生産された 自生性有機物の分解・無機化に伴った生成と、内湖外から の他生性の栄養塩種の供給が複雑に絡み合い、内湖の内部 生産、すなわち植物プランクトン、付着藻類、ならびに大 型水生植物などの光合成基礎生産を支えていると想定され た。 そ こ で、Forsberg and Ryding(1980) が 提 唱 し た
図 2 DIN:DIP 比値(●)から判定した植物プランクト ンの栄養制限関係図. 図中の直線は Forsberg and Ryding (1980)による判定基準線を示す.
DIN:DIP モル比と Redfield モル比との比較から、内湖の 植物プランクトン増殖因子を判定することを試みた。本研 究結果の DIN:DIP 比から内湖の栄養状態を判定するた めに、DIN:DIP 比のおよそ 11 以下を窒素律速、そして およそ 27 以上をリン律速とした(図 2)。図に見られるよ うに、一部の琵琶湖周辺内湖では、植物プランクトン増殖 に対して窒素栄養塩化合物が栄養制限因子として働いてい たが、ほとんどの内湖は、植物プランクトンの増殖に対し てリンが制限因子として機能していたと考えられた。 本調査で得られた PC、PN および PP 現存量の値から、 それぞれの内湖における PC:PN:PP モル比を計算し、 その季節変動を表 2 に示した。PC:PN 比の測定期間中の 平均値が最大となった内湖は南浦内湖で、最小値は伊庭内 湖で得られた。一方、PC:PP 比の最大平均値は曽根沼で、 最小平均値は細江内湖で得られた。これらの懸濁態生元素 モル比と Redfield(1958)が報告した Redfield モル比の 表 2 各内湖における懸濁態炭素(PC)、窒素(PN)、リン(PP)のモル比、および懸濁態炭素(PC)とクロロフィル a(Chl.a)の重量比の季節変動. PC:PN:PP PC:Chl.a
June August October December Jun. Aug. Oct. Dec.
堅田内湖 86 : 14.4 : 1 77 : 14.2 : 1 51 : 6.3 : 1 89 : 15.0 : 1 50 65 144 78 近江舞子沼 80 : 14.1 : 1 86 : 11.5 : 1 130 : 20.6 : 1 76 : 15.5 : 1 81 82 20 97 乙女ヶ池 83 : 11.8 : 1 71 : 13.4 : 1 86 : 10.3 : 1 53 : 8.5 : 1 146 65 46 123 松ノ木内湖 46 : 7.6 : 1 18 : 2.5 : 1 21 : 2.8 : 1 26 : 8.2 : 1 49 215 51 417 五反田沼 47 : 7.7 : 1 73 : 10.5 : 1 49 : 7.5 : 1 84 : 11.9 : 1 127 136 49 114 十ヶ坪沼 38 : 7.8 : 1 153 : 22.9 : 1 47 : 6.9 : 1 55 : 7.6 : 1 224 21 46 33 菅沼 33 : 5.0 : 1 43 : 8.1 : 1 17 : 2.6 : 1 29 : 4.4 : 1 94 89 139 516 浜分沼 120 : 17.6 : 1 68 : 11.9 : 1 100 : 13.5 : 1 86 : 13.9 : 1 32 173 49 135 貫川内湖(南) 89 : 15.2 : 1 76 : 12.2 : 1 37 : 5.1 : 1 39 : 4.8 : 1 133 97 505 1080 貫川内湖(北) 54 : 7.8 : 1 75 : 13.5 : 1 54 : 9.7 : 1 58 : 12.1 : 1 109 65 41 121 野田沼(湖北) 115 : 12.5 : 1 85 : 11.3 : 1 74 : 11.4 : 1 73 : 11.2 : 1 122 109 31 63 南浦内湖 128 : 13.1 : 1 45 : 7.1 : 1 69 : 11.2 : 1 105 : 12.6 : 1 133 78 30 72 細江内湖 20 : 4.3 : 1 16 : 2.3 : 1 12 : 2.3 : 1 14 : 1.7 : 1 168 1050 80 147 蓮池 30 : 4.4 : 1 31 : 5.1 : 1 40 : 6.0 : 1 25 : 3.3 : 1 268 464 65 130 野田沼(彦根) 63 : 10.2 : 1 69 : 12.0 : 1 35 : 5.4 : 1 42 : 6.7 : 1 196 125 79 195 曽根沼 117 : 16.3 : 1 131 : 16.4 : 1 67 : 14.4 : 1 165 : 22.5 : 1 156 99 139 94 神上沼 48 : 8.2 : 1 48 : 9.3 : 1 22 : 3.3 : 1 31 : 5.6 : 1 83 58 280 125 伊庭内湖 129 : 28.4 : 1 70 : 12.8 : 1 56 : 8.6 : 1 48 : 8.1 : 1 43 108 112 126 西の湖(西) 81 : 15.3 : 1 53 : 11.0 : 1 50 : 6.9 : 1 88 : 17.2 : 1 66 64 90 152 西の湖(東) 63 : 12.0 : 1 53 : 10.0 : 1 46 : 7.5 : 1 146 : 16.8 : 1 106 75 128 61 北の庄沢 56 : 9.3 : 1 92 : 13.2 : 1 25 : 3.0 : 1 39 : 7.2 : 1 60 127 406 234 北沢沼 83 : 11.8 : 1 40 : 6.4 : 1 70 : 9.3 : 1 94 : 14.1 : 1 140 67 84 98 志那中湖 20 : 3.4 : 1 25 : 4.3 : 1 16 : 1.9 : 1 11 : 2.1 : 1 320 128 390 591 柳平湖 102 : 14.0 : 1 114 : 13.5 : 1 98 : 15.7 : 1 59 : 8.2 : 1 88 152 92 93 平湖 96 : 14.9 : 1 112 : 16.5 : 1 130 : 18.7 : 1 109 : 13.3 : 1 54 231 71 124 106:16:1 とを比較すると、琵琶湖周辺内湖の平均 PC: PN 比は 6.5 と計算され、Redfield 比の 6.6 と同程度の値で あった。しかし、内湖水の平均 PC:PP 比、および平均 PN:PP 比はそれぞれ 62 および 9.5 と計算され、Redfield 比の 106 および 16 よりかなり低かった。
Healey and Hendzel(1979, 1980) は、PC:PN、PC: PP、PN:PP 比から、植物プランクトン増殖に対する栄 養制限因子を評価することを提唱した。彼らがその判定基 準とした関係をモル比で表現すると、植物プランクトン増 殖に対して、PC:PN 比が約 10 以下であれば窒素制限は 無く、10 ~ 15 であれば中程度の窒素制限があり、15 以上 であれば厳しい窒素制限がある。一方、PC:PP 比が約 130 以下であればリン制限は無く、130 ~ 260 であれば中 程度のリン制限があり、260 以上であれば厳しいリン制限 を植物プランクトンは増殖の際に受けることになる。 それぞれの琵琶湖内湖水の PC と PN あるいは PC と PP
の関係を図 3 に示した。PC:PN 関係図に見られるように、 6 月に湖北野田沼と南浦内湖でわずかに中程度の窒素制限 を受けていたが、琵琶湖周辺内湖のほとんどは窒素制限を 受けていないと考えられた。一方、PC:PP 関係図に見ら れるように、調査季節にもよるが伊庭内湖、曽根沼、十ヶ 坪沼、平湖、近江舞子沼、曽根沼、西の湖は中程度のリン 制限を受ける図に示した判定範囲にプロットされたが、そ の他の内湖は植物プランクトン増殖に対してリン制限を受 けていないと判断された。なお、PN:PP 比からも、一部 の内湖(伊庭内湖、十ヶ坪沼、曽根沼)では、中程度のリ ン制限を受けていたと判定されたが、琵琶湖内湖の多くは リン制限を受けていなかったと判断された。このように、 Healey and Hendzel(1979, 1980)が提唱した懸濁態生元 素比からの判定によると、琵琶湖周辺内湖水のほとんどは、 植物プランクトン増殖に対して栄養制限を受けていなかっ たと考えられた。 これは、内湖の窒素・リン・ケイ素栄養塩現存量に見ら れるように(表 1)、これら内湖が富栄養湖であることが 大きな要因であると考えられる。 3-3 植物プランクトン増殖に対する栄養制限判定の検討 PC:PN:PP 比から植物プランクトン増殖に対する栄 養制限要因を判定すると、上述したように琵琶湖内湖の植 物プランクトンは窒素あるいはリンによる栄養制限をほと んど受けていなかった(図 3)。一方、DIN:DIP 比から 内湖の植物プランクトン増殖に対する律速を判定すると、 多くの内湖の植物プランクトンはリン律速を受けていた判 定結果が得られた(図 2)。そこで、内湖の栄養制限を評 価する指標は、いずれの判定方法がより適しているかを検 討した。水中の DIN と DIP 現存量が植物プランクトンが 要求する栄養源に比して充分現存する場合は、DIN:DIP 比は基本的に変動しないと考えられる。したがって、水中 の DIN:DIP 比 を 植 物 プ ラ ン ク ト ン 細 胞 構 成 元 素 の Redfield 比と比較して、DIN と DIP のいずれが植物プラ ンクトンの増殖制限因子になりうるかの議論は、山本 (1999)が指摘するように、適切でないと考えられる。こ のことは、とくに富栄養潟湖の琵琶湖内湖の植物プランク トン増殖に対する栄養制限因子を探る方法は、水中栄養塩 の DIN:DIP 比から判断するよりも、水深が浅い内湖の 特性(濁水環境などの水環境)による誤判定を除いて、植 物プランクトン細胞の構成生元素比(PC:PN:PP 比) から判定することが上述の判定方法の中ではより妥当性が あると判断せざるを得ない。 したがって、本研究では、PC:PN:PP 比を内湖の植 物プランクトンの栄養制限の主たる判定指標に用いた。し かし、上述したように、琵琶湖内湖においては植物プラン クトン以外の懸濁物質が PC:PN:PP 比に影響を与える ことが充分考えられるため、懸濁態生元素比を指標として 植物プランクトンの栄養制限を判定するとき、内湖水の Chl.a 現存量のレベルを考慮に入れて判定することとした。 すなわち Chl.a が 15 µg chl.a L-1以上の試料に対しては、 懸濁態生元素比(PC:PN:PP)から判定した。この理由 は、これ以上の Chl.a 現存量の試水においては、PC:Chl.a、 PN:Chl.a および PP:Chl.a 比の値が一般に測定される植 物プランクトン細胞のこれらの構成比に近いと考えられる からである。なお、Chl.a がこの現存量以下である内湖は、 基礎生産が比較的低い内湖であり、湖水の栄養塩レベルが 低いために内湖の植物プランクトンの光合成生産を含めた 富栄養化が促進されなかったと判断し、DIN:DIP 比も植 物プランクトン増殖に対する律速因子の指標になりうると 考えた。 本研究で調査した琵琶湖内湖の植物プランクトン増殖に 図 3 PC:PN:PP 比値(●または×)から判定した植 物プランクトンの栄養制限関係図.上図は PC:PN 判定、 下図は PC:PP 判定.×:窒素またはリンが若干制限さ れた内湖.図中の直線:Healey and Hendzel(1980)の 判定基準線.
対する律速要因を、これらの基準から判定した結果は次の ごとくにまとめることができる。Chl.a 現存量が 15 µg chl.a L-1以上の内湖の一部では、図 3 に示したように、夏季に 中程度の窒素制限、秋季から冬季に中程度のリン制限を受 けたが、他のほとんどの内湖の植物プランクトンはその増 殖に対して栄養制限を受けていなかった。一方、Chl.a が 15 µg chl.a L-1以下の植物プランクトン現存量が比較的低 いレベルの内湖では、Forsberg and Ryding(1980)の指 標に従うと、植物プランクトン増殖に対して窒素あるいは リンの制限を受けていた(図 2)。ただし、琵琶湖内湖の 栄養塩現存量のレベルから判断すると(表 1)、この判定 結果はごく一部の内湖にのみ適用できると考えられた。 3-4 内湖の栄養段階 OECD(1982)は、湖沼水の全リン現存量の年平均レベ ルから、湖沼を貧栄養湖(全リン現存量が 0.3 µM 以下)、 中栄養湖(0.3 ~ 1.1 µM)、そして富栄養湖(1.1 µM 以上) に分類した。本研究で得た全リン(DIP と DOP と PP の 合計)現存量(ただし、表 1 には DOP 値を掲載していない) の年平均現存量(4 回測定した平均値を年平均現存量とし た)を OECD(1982)が提唱した分類基準と比較検討す ることにより、琵琶湖内湖の栄養段階を評価した。その判 定結果、琵琶湖周辺内湖のすべてが、全リンの平均現存量 が 1.1 µM 以上存在する富栄養湖であると考えられた。さ らに、OECD(1982)は、クロロフィルa現存量の基準か らも湖沼の栄養段階を分類した。本研究で得た Chl.a 現存 量(表 1)と OECD(1982)が提唱した Chl.a 現存量の基 準値を比較することにより、琵琶湖内湖の栄養段階を評価 すると、OECD 基準の 2.5 µg chl.a L-1以下の志那中湖と 細江内湖が貧栄養湖、西の湖(東)、菅沼、貫川内湖(南)、 蓮池が中栄養湖(2.5 ~ 8 µg chl.a L-1)、そして他の 18 内 湖が富栄養湖(8 µg chl.a L-1以上)に分類された(表 3)。 このように、内湖の栄養段階の評定は、全リン現存量か らの判定結果と、クロロフィルa現存量からの判定結果が 異なった。すなわち、OECD(1982)の基準は、栄養段階 に対する目安の評価であるといえる。とくに、内湖は小潟 湖のため、面積が小さく、水深が浅く、水と物質の流入・ 流出の速度・量が内湖容積の全量に比較して大きく、内湖 の湖内での滞留時間が短い水系であるため、高い栄養塩現 存量湖水であっても植物プランクトンが充分に増殖・成長 するまでの時間に湖水は内湖から流出することが考えられ る。これらのことが、クロロフィルa現存量による栄養段 階の評価が、リン現存量によるそれよりも貧栄養的に評価 されやすい可能性があると考えた。
4 おわりに
本研究対象とした琵琶湖周辺内湖の植物プランクトン は、懸濁態生元素比による判定基準(Healey and Hendzel, 1980)を適用すると、その増殖に対して栄養制限を受ける ことは少ないと判断された。ただし、植物プランクトン現 存量が低い内湖に対しては、懸濁態生元素比のみを適用し て栄養制限を判定することは困難であり、貧栄養湖あるい は中栄養湖と判定された 6 内湖では、栄養塩比による判定 基準(Forsberg and Ryding, 1980)を使用すると、植物 プランクトン増殖にとってリン制限を受けていた可能性が 示唆された。栄養段階判定(OECD, 1982)の結果、一部 の内湖は貧栄養あるいは中栄養湖と考えられたが、他のほ 表 3 TP 現存量および Chl.a 現存量から判定した内湖の 栄養段階.富栄養湖:TP > 1.1 µM または Chl.a > 8 µg L- 1、中栄養湖:Chl.a 2.5 ~ 8 µg L- 1、貧栄養湖:Chl.a < 2.5 µg L- 1. 栄養段階 TP 判定 Chl.a 判定 堅田内湖 富栄養湖 富栄養湖 近江舞子沼 富栄養湖 富栄養湖 乙女ヶ池 富栄養湖 富栄養湖 松ノ木内湖 富栄養湖 富栄養湖 五反田沼 富栄養湖 富栄養湖 十ヶ坪沼 富栄養湖 富栄養湖 菅沼 富栄養湖 中栄養湖 浜分沼 富栄養湖 富栄養湖 貫川内湖(南) 富栄養湖 中栄養湖 貫川内湖(北) 富栄養湖 富栄養湖 野田沼(湖北) 富栄養湖 富栄養湖 南浦内湖 富栄養湖 富栄養湖 細江内湖 富栄養湖 貧栄養湖 蓮池 富栄養湖 中栄養湖 野田沼(彦根) 富栄養湖 富栄養湖 曽根沼 富栄養湖 富栄養湖 神上沼 富栄養湖 富栄養湖 伊庭内湖 富栄養湖 富栄養湖 西の湖(西) 富栄養湖 中栄養湖 西の湖(東) 富栄養湖 富栄養湖 北の庄沢 富栄養湖 富栄養湖 北沢沼 富栄養湖 富栄養湖 志那中湖 富栄養湖 貧栄養湖 柳平湖 富栄養湖 富栄養湖 平湖 富栄養湖 富栄養湖とんどの内湖は富栄養湖と判定した。この富栄養内湖の植 物プランクトンは窒素あるいはリン栄養塩化合物による栄 養制限を受けることは少ないと判断された。このように、 琵琶湖周辺内湖の多くは、植物プランクトンあるいは付着 藻類増殖の制限因子になる栄養塩化合物現存量は豊富に存 在する富栄養湖であり、懸濁態生元素の比から判定して栄 養制限を示さなかった内湖が大半を占めていたことと一致 する。指摘されているように、富栄養湖沼においては栄養 塩比から栄養制限を評価することは困難であると考えられ る。とくに浅く小潟湖で富栄養化した琵琶湖内湖では、こ の栄養塩現存量比からの栄養制限の判定結果は信頼性を失 う。しばしばあらゆる水域で用いられているこの基準比は、 誤った判定を下すことになり、あくまで目安としてこの比 による栄養制限を判断するべきである。富栄養小湖におい て植物プランクトン増殖に対する栄養制限を評価する際に これらの問題点の解決には、本研究測定パラメーターとし た懸濁態生元素と栄養塩化合物現存量の多少とそれらの関 係に加え、さらに植物プランクトンの光合成活性、植物プ ランクトン増殖に働く微量成分、さらに物理・地学・生物 的要因などから総合的に判断して、植物プランクトンの増 殖の因子を探り解析していく必要がある。 上述したように、現存の琵琶湖周辺「里水」内湖の多く は富栄養湖であり、内湖水中の植物プランクトンは窒素あ るいはリン栄養塩化合物による光合成増殖の制限を大きく 受ける可能性は低いと考えられた。しかし、一部の内湖の ある季節では、貧栄養あるいは中栄養的であり、植物プラ ンクトンの光合成増殖は、リンによる制限により抑制され ていた可能性がある。このことは、琵琶湖内湖の富栄養化 防止対策のために、「里水」内湖の水環境復元のための湖 沼管理を今後とも充分に行っていくことが重要である。内 湖は琵琶湖と繋がっており琵琶湖の水環境に多大なる影響 を与える。琵琶湖内湖を琵琶湖流域の一部として思考視野 を拡大し里水保全のための水環境管理を真摯に考えていか なければならない。 本論文をまとめるにあたって追記したい。共著者の尾崎 正樹は本研究活動と成果を基に身近な水環境学習を学校教 育に生かすべく、中学校教育者として活躍していた。彼の 教育に対する高い志は万人が認めるところであり、高い評 価を得ていた。しかし、彼は志半ばで 2013 年 4 月 5 日永 眠し短い人生を終えた。彼の心中を察するに余りある。水 環境教育に対する彼の志を思い、ここに謹んで哀悼の意を 表するとともに故人の墓前に本論文を捧げたい。
謝 辞
本研究を進めるにあたり、野外調査と実験室における化 学分析測定に多大なる援助をいただいた滋賀県立大学環境 科学部湖沼環境実験施設の海外文一郎氏、ならびに同大学 環境科学研究科環境動態学専攻および環境科学部環境生態 学科の学生諸子に深謝を申し上げる。引用文献
Aikazi, M. and A. Otsuki (1987): Characteristics of variations of C:N:P:Chl ratios of seston in eutrophic shallow Lake Kasumigaura. Jpn. J. Limnol., 48: S99– S106.
Bendschneider, K. and R. J. Robinson (1952): A new spectrophotometric method for the determination of nitrite in sea water. J. Mar. Res., 11: 87–96.
Carlson, R. E. (1977): A trophic state index for lakes. Limnol. Oceanogr., 22: 361–369.
Forsberg, C. and S. O. Ryding (1980): Eutrophication parameters and trophic state indices in 30 Swedish wast-receiving lakes. Arch. Hydrobiol., 89: 189–207. 浜端悦治(2010):湖沼生態系の保全・回復に向けて.関
西自然保護機構会誌,32:47–51.
Healey, F. P. and L. L. Hendzel (1979): Indicators of phosphorus and nitrogen deficiency in five algae in culture. J. Fish. Res. Board Can., 36: 1364–1369. Healey, F. P. and L. L. Hendzel (1980): Physiological
i n d i c a t o r s o f n u t r i e n t d e f i c i e n c y i n l a k e phytoplankton. Can. J. Fish. Aquat. Sci., 37: 442–453. Hoyer, M. V. and D. E. Canfield (1994): Bird abundance
and species richness on Florida lakes: influence of trophic status, lake morphology, and aquatic macrophytes. Hydrobiologia, 279/280: 107–119. 倉田亮(1984):内湖 -その生態学的機能-.滋賀県琵琶 湖研究所所報,2:46–54. 倉田亮(1988):水草帯と内湖 -その現状と機能-.滋賀 県琵琶湖研究所 5 周年記念誌,琵琶湖研究集水域から 湖水まで,滋賀県琵琶湖研究所所,142–163.
Mehner, T., M. Dielmann, U. Bramick and R. Lemcke (2005): Composition of fish communities in German lakes as related to lake morphology, trophic state,
shore structure and human-use intensity. Freshwater Biol., 50: 70–85.
Menzel, D. W. and N. Corwin (1965): The measurement of total phosphorus in seawater based on the liberation of organically bound fractions by persulfate oxidation. Limnol. Oceanogr., 10: 280–283.
Mitamura, O. (1997): An improved method for the determination of nitrate in freshwaters based on hydrazinium reduction. Mem. Osaka Kyoiku Univ. Ser. Ⅲ, 45: 297–303. 三田村緒佐武・石川聡子・石田典子・後藤直成・橘淳治・ 丸尾雅啓(2014):びわ湖内湖・西の湖における水環 境教育.陸水研究,1:5–15. 三田村緒佐武・後藤直成(2005):琵琶湖周辺内湖の水質 特性 ―内湖の水質特性―.185–194,西野麻知子・浜 端悦治編「内湖からのメッセージ 琵琶湖周辺の湿地 再生と生物多様性保全」.サンライズ出版,pp.253. Mitamura, O., J. Tachibana, N. Ishida, Y. Seike and J. K.
Choi (2009): Photosynthetic activity of epiphytic algae in embayment reed zone in a lagoon connected with Lake Biwa. Kor. J. Limnol., 42: 48–57.
Miyake, H., K. Nozaki and O. Mitamura (2002): Chemical characteristics of small lagoons “Naiko” connected with Lake Biwa. Verh. Internat. Verein. Limnol., 28: 1727–1732.
Mullin, J. B. and J. P. Riley (1955): The colorimetric determination of silicate with special reference to sea and natural waters. Anal. Chim. Acta, 12: 162–176. Murphy, J and J. P. Riley (1962): A modified single
solution method for the determination of phosphate in natural waters. Anal. Chim. Acta, 27: 31–36. Nagoshi, M. (1965): Ecological studies on the population
of gengorobuna, Carassius cuvieri, in Lake Biwa. II. Mortality, size of spawning population and number of emigrants estimated with tagging method. Rep. Fac. Fish. Mie Pref. Univ., 5: 307–317.
中西正巳・関野樹(1997):琵琶湖水質の生物学的特徴. 環境技術,26:13–17.
Nakanishi, M., O. Mitamura and T. Matsubara (1990): Sestonic C:N:P ratios in the south basin of Lake Biwa with special attention to nutritional state of phytoplankton. Jpn. J. Limnol., 51: 185–189. 西野麻知子(2005):琵琶湖と内湖の関係.54–61,西野麻 知子・浜端悦治編「内湖からのメッセージ 琵琶湖周 辺の湿地再生と生物多様性保全」.サンライズ出版, pp.253. 大久保卓也(1998):ため池,内湖を利用した水質浄化 . 用水と排水 , 41:883–893.
OECD (1982): Eutrophication of waters: Monitoring, Assessment and Control. OECD, Paris.
Redfield, A. C. (1958): The biological control of chemical factors in the environment. Am. Sci., 46: 205–221. Sagi, T. (1966): Determination of ammonia in sea water
by the indophenol method and its application to the coastal and off-shore waters. Oceanogr. Mag., 18: 43– 51.
西條八束・三田村緒佐武(1995):新編湖沼調査法 . 講談社, pp.230.
Sakamoto, M. (1966): Primary production by the phytoplankton community in some Japanese lakes and its dependence on lake depth. Arch. Hydrobiol. 62: 1–28.
Tezuka, Y. (1984): Seasonal variations of dominant phytoplankton, chlorophyll a and nutrient levels in pelagic regions of Lake Biwa. Jpn. J. Limnol., 45: 26– 37.
Tezuka, Y. (1985): C:N:P ratios of seston in Lake Biwa as indicators of nutrient deficiency in phytoplankton and decomposition process of hypolimnetic particulate matter. Jpn. J. Limnol., 46: 239–246. 山田佳裕・野崎健太郎(1997):炭素・窒素安定同位体比 精密測定法を用いた琵琶湖生態系の解析.月刊海洋, 29:399–407. 山本民次・橋本俊也・松田治・多田邦尚(1999):瀬戸内 海における植物プランクトン態 N:P 比とその変動要 因 . 水産海洋研究 , 63:6–19.
Vollenweider, R. A. and J. J. Kerekes (1980): Synthesis Report, Cooperative Programme on Monitoring of Inland Waters (Eutrophication Control). Report prepared on behalf of Technical Bureau, Water Management Sector Group, Organization for Economic Cooperation and Development (OECD), Paris, pp.290.