九州大学学術情報リポジトリ
Kyushu University Institutional Repository
酸素電極法を用いたイネ光合成能力の種内変異と遺 伝に関する研究
長峰, 司
https://doi.org/10.11501/3070096
出版情報:Kyushu University, 1993, 博士(農学), 論文博士 バージョン:
権利関係:
第5章 酸素放出量の遺伝分析
第4章の実験でイネ葉身酸素放出量には在来品種および改良品種に大きな遺 伝的変異があること、 また、 突然、変異処理によっても酸素放出量に遺伝的変異 を誘発できることなどが明らかとなった。 すなわち、 イネ種内に存在する光合 成能力の変異の大きさを把握することができ、 人為的な改良も可能であること がわかった。 そこで、 品種の光合成能力の改良を具体化するため、 品種間交雑 の雑種後代を用いて酸素放出量の遺伝様式を調べた。
第1節 特定の生育時期における酸素放出量の遺伝分析
酸素放出量の遺伝様式を明らかにするため、 交雑により数組合せの雑種集団 を作出し、 ある特定の生育時期における酸素放出量の遺伝分析を行った。
材料および方法
第4章第1節の酸素放出量の種内変異に関する実験および予備実験の結果か ら高、 低酸素放出量系統をそれぞれ選んだCTable 16)。 すなわち、 材料は、
中国の改良品種窄葉青8号(高) x日本の在来品種信州金子(低)と日本の在 来品種十石(高) x中国の在来品種納西(低)の正逆F2雑種集団および窄葉青 8号×信州金子のF1に窄葉青8号を戻し交雑したB1F1集団、 十石×納西のF1、
納西×十石のF1に それぞれ十石を戻し交雑したB1 F 1集団であった。 Table 16'こ 両親の形質を示した。 F2集団およびB1F1集団を両親、 F 1とともに19 8 5年5月8
に播種し、 6月5日に圃場に1株1本植えした。 酸素放出量の測定数は、 両
親、 F1が5から10反復で、 F2集団は反復せずに約300個体、 B1F1集団は反復な しで約100個体とした。 F2集団およびB1F1集団の酸素放出量の測定にあたって は周辺効果を避けるため試験区周辺の個体を除いて行った。 集団を構成する各 個体の生育ステージは異なると予想されたが、 移植後の一時期に酸素放出量を
b�
Tab1e 16. Agronomic characteristics of parental lines used for genetic analysis
Indigeneous Heading Day s to Cul m Panic1e Number of Apicu1us Variet y name Origin or improved date heading length 1ength panic1es Awn co1or
(days) (cm) (cm) (/ p 1 a n t)
C〈T 、〉 Jukkoku J apan Indigenous 29/Aug. 113 98 21. 0 13 No Ye110w
Shinshu-kaneko do. Indigenous 15/Aug. 99 104 21. 7 15 No Ye 11 ow Tamanishiki do. Indigenous 28/Aug. 112 115 21. 6 12 No Y e 11 ow
Naxi China Indigenous 27/Aug. 111 159 3 O. 0 11 No Ye 11 ow
Zaiyeqin 8 do. Improved 18/Aug. 102 89 21. 5 11 No Y e 11 ow
測定して酸素放出量の遺伝様式を明らかにしようとした。 すなわち、 Table 17 に示す一時期にそれぞれの集団における酸素放出量の測定を行った。 酸素放出 量と葉身形質および農業形質との関係を明らかにするため、 酸素放出量の他に 酸素放出量を測定するのに用いた葉身の葉面積、 同乾物重、 稗長、 穂、長、 1株 穂数、 出穏期を個体ごとに調査した。
結果および考察
窄葉青8号×信州金子の組合せにおけるF 1植物の酸素放出量は537μmo1 02/
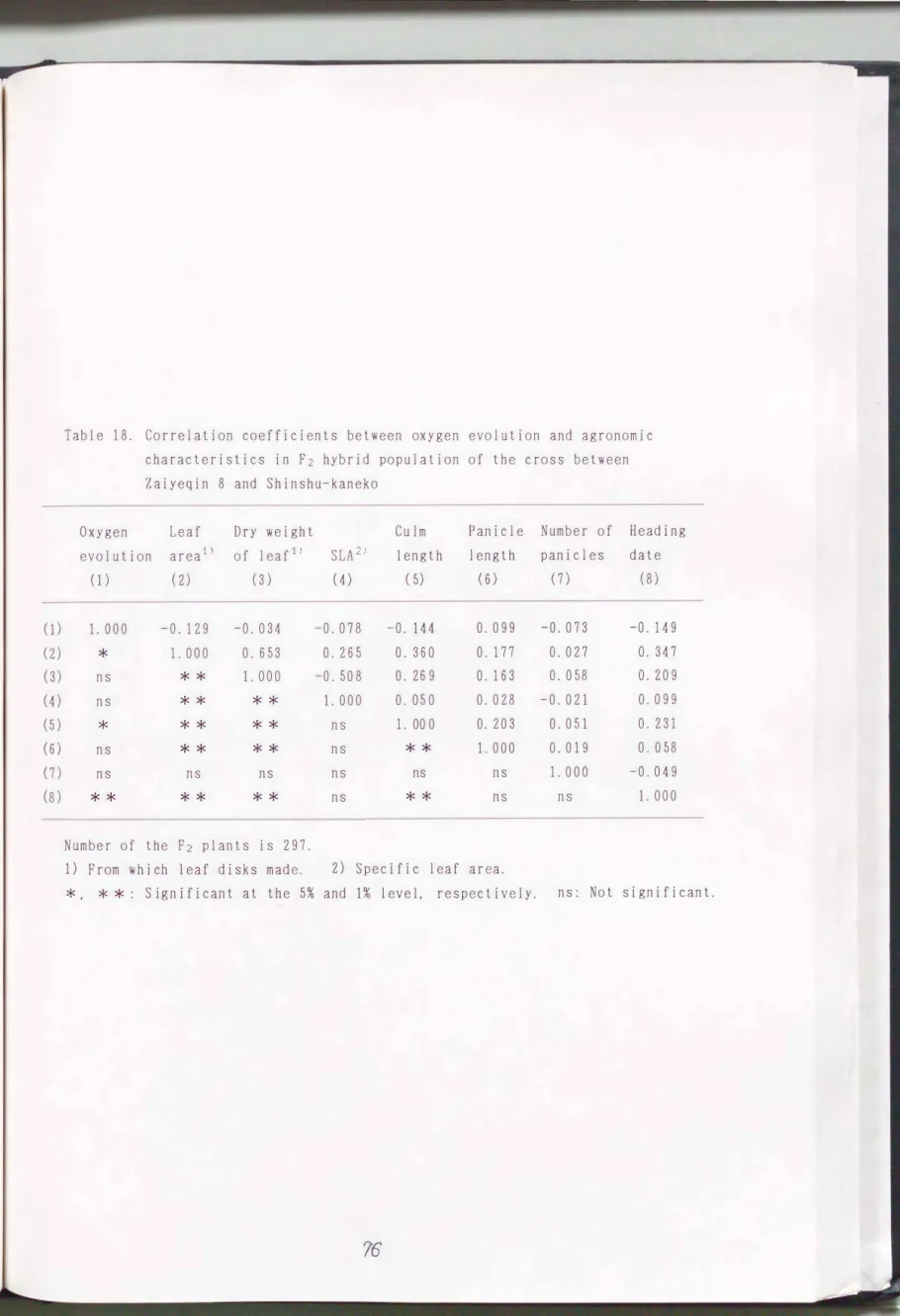
dm2/hrであり、 低放出量親である信州金子の値510μmol 02/dmL/hr'こ近かった (Table 17)。 つぎに、 F2雑種集団の放出量を7月19日から21日(移植後44'"'"' 46日)にかけて測定したところ、 最低値301μmol 02/dmL/hrから最高値 952μ mol 02/dm2/hrで変異の幅は広く、 両親の値より超越した値となった個体があ った(Fig. 16) 0 297個体からなるF2雑種集団の酸素放出量の分布型は正規分 布に似ていた。
逆組合せである信州金子×窄葉青8号のF 1植物の酸素放出量は479 μmo1 02 /dm2/hrであり、 低酸素放出量親である信州金子の値557μmol 02/dmL/hrに近 かった(Table 17) 0 7月23日から24日(移植後48'"'"' 49日)にかけて測定した 293 個体からなるF2集団の酸素放出量の変異は、 最低値が226μmol 02/dmι/hr、
最高値が973μmol 02/dm2/hrであり、 両親の値より大きく超越した分布となっ た(Fig.17)。 その分布の形は正規分布に近かった。
窄葉青8号×信州金子のF1に窄葉青8号を戻し交雑したB1F1雑種集団の酸素 放出量の変異は最低値368μmol 02/dm2/hrから最高値1, 040μmol 02/dm2/hrで あり、 二頂分布よりも正規分布に近い 分布型となった(Fig. 18)。
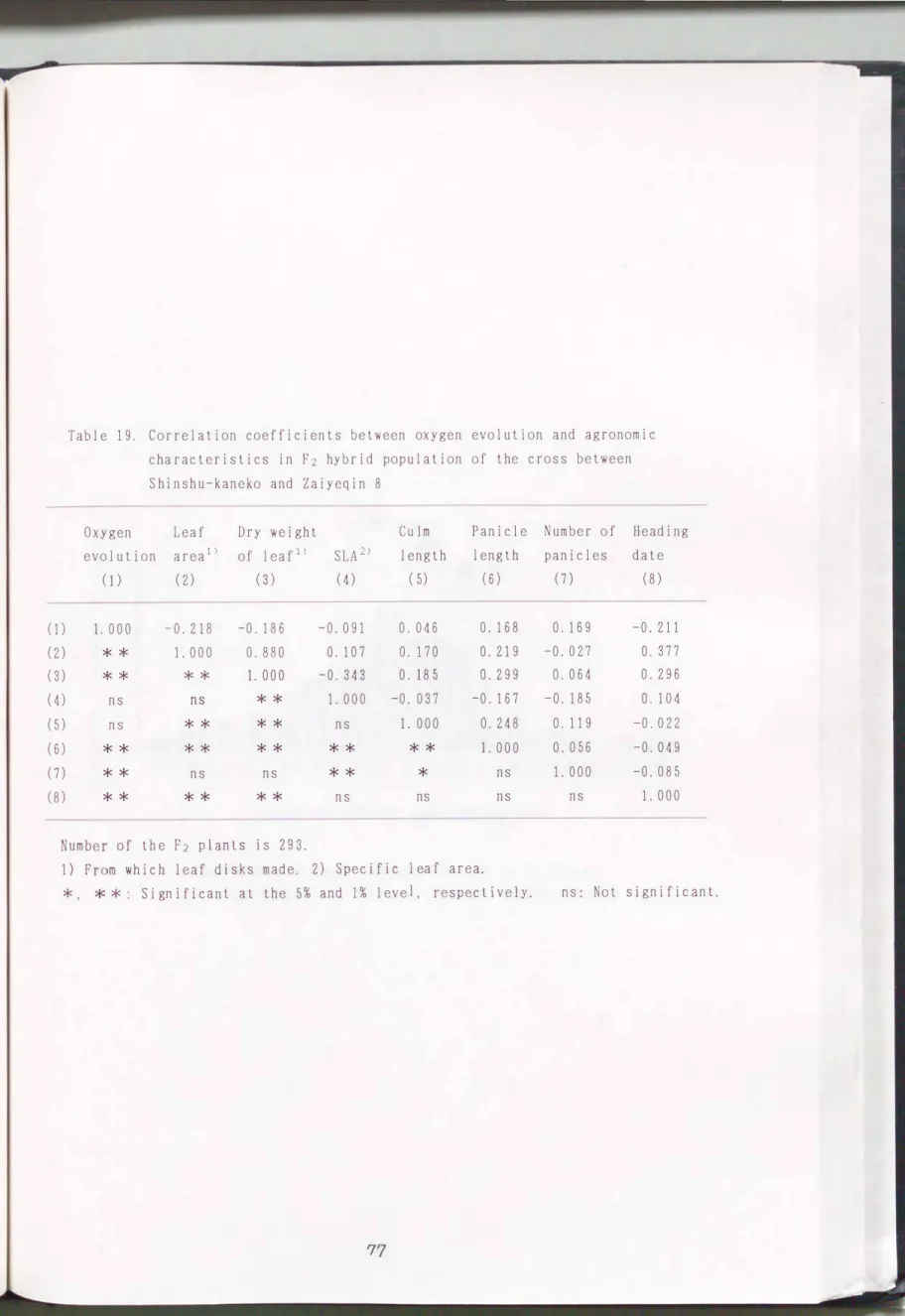
窄葉青8号×信州金子および信州金子×窄葉青8号の正逆F2個体聞には、 酸 素放出量の測定に用いた葉身の葉面積と葉身乾物重との聞に正の有意な相関 (r=0.653州、 r=0.880*勺
、
葉身乾物重とSLAとの聞に負の相関(r=一O. 508料、守0
守
、、‘
Table 17. Oxygen evolution in Fl plants and their parents
Oxygen evolution (μmol 02/dmL/hr)
Low capacity High capacity Measurement
Cross combination parent parent Fl plant date
Zaiyeqin 8(H) X Shinshu-kaneko(L) 510 674 537 19/Jul. -21/Jul.
Shinshu-kaneko(L) X Za iyeqin 8 (H) 557 673 479 23/Jul. -24/Jul.
Jukkoku(H) X Naxi (L) 526 589 538 5/Aug. -7/Aug.
Naxi (L) X Jukkoku(H) 476 547 480 l/Aug. -3/Aug.
H or L in the parentheses shows a high or low capacity of oxygen evolution, respectively.
(N)
L
8 Shinshu-kaneko
aL__ f
→
t -
S司..
Zaiyeqin
-凡
L
15
10 ωμ口付叶門凶
比4
0 ヘハυロつごピ引い』
hHOいω』Eコロ
豆沼E555555555555555EE冨己55冨55 02/dm2/hr)
(ドmol Oxygen evolution
Dislribulion of oxygcn evolulion in 1'2 hybrid popu!alion of lhe cross belween Zaiyeqin 8 16
F i g.
and Shinshu-kaneko.
ワ乙
Shinshu-kaneko
『 a・・k I
' ‘・, I
ト.ト寸 Zaiyeqiτ1
8F ,
l
I 竺hωμロの,〔(円
(N) 同ohHω門戸Eコロ出ohυロωコσりい』
タ mol 02/dm2/hr) Oxygen evolution
Distribution of oxygen evolution in F2 hybrid population of the cross between Shinshu-kaneko and Zaiyeqin 8
F i g. 17
At
I .4111・h
司..
Zaiyeqin 8
(N) ωμ口付叶(円同ohHω』Eヨロ刷ohυロωコピωい』
<=コ 1.、 ε:::> o..n r工コ 1.1"、 C二:> l("ラ Cこコ lO Cコ 1("ヲ ζ二:> l.t"ラ C:::::> LC'> C::J l.l"") c::> L.rl Cコ Ln cニコ ,.("、 Cコ u"、 ζ亡:> o..n lf、E、- Cコ c、寸 lf、 ε、. ζ二コ c、.J LO c-- c:コ p、J lfヲ E、- cコ〈、J lf予 r::-- Cコ c、、, LCヲ官、- cコ C"'--.I LC'l C、- cコc、J r-、 ,...., 竜三ー ‘才 司オー 、.J- lO l.r) ‘r> lf、 uコ ζ工3 ζD c...n c-- c--ーr::-- r::-- 口コ ロコ cx) co 0'""") CTl CT") 0-' C:=> Cコ 一一
い01 02/dm2/hr)
Oxygen evo1ution
Distribution of oxygen evolution in B1Fl hybrid (Zaiyeqin 8/Shinshu-kaneko)//
population of Zaiyeqin 8.
F i g. 18
ク''1
r=-0.343京本)が観察された(Table 1 8 、Table 19)。 酸素放出量と葉面積、 手早 長、出穏期との聞の相関係数はそれぞれ個体数が多いので有意となったが、 値 は小さかっアこ。
つぎに、十石×納西の組合せにおいて高放出量親十石の酸素放出量は 589μ mol 02/dm2/hr 、低放出量親納西は52 6 μmol02/d ν/hrであった。 F1植物の値 は538μmol02/d ザ/hrであり、 納西の値に近かった(Table 17) 0 F2雑種集団 296個体の酸素放出量を8月5日から7日(移植後 61--..63日)にかけて測定し たところ、 最低値1 52μmol02/dm2/hr 、 最高値867μmol02/d ド/hrであり、
両親の値よりはるかに超越した分布であった(Fig. 19)。
正逆組合せの納西×十石において、 納西の酸素放出量は476μmol02/dmL/hr 、 十石は5 47μmol02/dm2/hrであった。 F1植物は4 80μmol02/dm2/hrであり、 低 放出量親の納西の値とほとんど同じであった(Table 17) 0 8月1日から3日 (移植後57--..59日)にかけて測定した、 299個体からなるF2集団の酸素放出量 は、十石×納西の組合せと同じように両親の値より大きく超越した分布となっ アこ(Fig. 20)
十石×納西のF 1に十石を戻し交雑したB1Fl集団、 納西×十石のF 1に十石を戻 し交雑したB1F1集団における酸素放出量は、 ともに正規分布に似た分布であっ
Tこ(Fig. 21 、Fig. 2 2)。
以上をまとめると、 二つの正逆組合せともにF2雑種集団における酸素放出量
の分布では、 両親の値より超越した個体が出現し、 酸素放出量の分布型は正規 分布に似ていた。 B1Fl雑種集団においても酸素放出量の分布型は正規分布に近 かった。 すなわち、ある特定の一測定時期の値からでは酸素放出量の遺伝様式 は簡単ではなかった。
酸素放出量の遺伝様式が簡単ではない理由として二つのことが考えられる。
一つは酸素放出量はもともと単一の主働遺伝子支配ではない、もう一つは用い た組合せが遠縁であり、 遠縁交雑の雑種集団において出穏期が両親より超越し
(1) (2) (3) (4) (5) (6 ) (7) (8)
Table
18.
Correlation coefficients between oxygen evolution and agronomic characteristics in F2 hybrid population of the cross betweenZaiyeqin
8
and Shinshu-kanekoOxygen Leaf Dry weight evolution area 1) of leaf1)
(1) (2) (3)
1. 000 -0.129 -0. 034
*
1. 000
O.653
ns **
1. 000
ns ** **
〉ド ** **
ns ** **
ns ns ns
** ** **
Number of the F2 plants is
297.
1)
From which leaf disks made.SLA 2)
(4)
-0. 078
O.
265 -0. 508 1. 000
ns ns ns ns
Culm length
( 5)
-0. 144
O.
360
O.
269
O.
050 1. 000
**
ns
**
Panicle length
(6)
O.
099
O.
177
O.
163
O.
028
O.
203 1. 000
ns ns
2)
Specific leaf area.*, * *: Significant at the 5児and
1%
level, respectively.76
Number of Heading panicles date
(7) (8)
-0.073 -0. 149
O.
027
O.347
O.
058
O.209 -0.021
O.099
O.
051
O.231
O.
019 0.058 1. 000 -0. 049
ns
1. 000
ns: Not significant.
Table 19. Correlation coefficients between oxygen evolution and agronomic characteristics in F2 hybrid population of the cross between Shinshu-kaneko and Zaiyeqin 8
Oxygen Leaf PLU HU 2EEE-- m Dry weight
evolution areal) of leaf1i SLA2.i
(1) (2) (3) (4)
Panicle Number of Heading length length panicles date
( 5) (6) (7) (8)
(1) (2) (3) (4) (5) ( 6 ) (7) (8)
1. 000
**
**
一0.218 -0.186 1. 000 0.880
* * 1. 000
-0.091 0.046 0.107 0.170 -0.343 0.185 1. 000 -0. 037
ns 1. 000
** **
* * *
ns *
*
*
*
*
*
s
*
*
n
*
* ns
*
*
*
*
*
*
ns ns
* * * * ns ns
O. 168 O. 219 O. 299
-0. 167
O. 248 1. 000
O. 169
-0.027
O. 064
-0.185
O. 119 O. 056
1. 000
-0. 211
O. 377 O. 296 O. 104
-0. 022 -0.049 -0. 085 1. 000
ιEEU nu nd PL ・唱EAri --ムn 円KO--A nb 4'EM O MN
cu nH
VJ 1sA 凸UVv ・-AaTEしv
nd nu 凸U ρu
rA ny
a
凸U s
rIA TA
nd ρu' 11A、11Aρu
nu vv
・-A凸U 21ふ Vla
-唱目ふ
C UA 凸U 1i
ny n\u
nH ,パU 11J 内d
nJ臼UA qu-Ed
nHd 凸U 円ノ臼
,パu
ρu
nd Lu cu m川 GL・-A nb ふL S LA
a
ふEL Fb
nu-
-A ゐL a
,Hu
n
--
nd nμ・
nすA
FUa・l
Aソ』
凸U
ρTA
nドE・
1lA
・1Anu
e -n
円6
2nH FU
--4 4L・
l pゐ 'hμ 内十A山w--om* ro= ρUTA叫司
'hu nドa
m
'
uO*
ns
ns ns
10
(N) ωμロの什円円山相ohHω門戸日ロロhHohυロωロげωhH』 、、,ノr 咽n,,,,,, つ』m AU ,f''' qL ハU1ムo m μJ ,,E、
Oxygen evolution
Distribution of oxygen evolution in F2 hybrid population of the cross between lukkoku and Naxi.
F i g. 19.
ケS
Naxi
l a・It. I
t ‘. I
t ..F1
t 司. I
lS
10
(N) ωμロωHn円刷Oいω門戸巨コロhHohucωコσωUH』
い mol 02/dm2/hr) Oxygen evolution
Dislríbulion of oxygen evolution in F2 hybrid populalion of the cross belween Naxi and Jukkoku 20.
F i g.
Fl
同・4
1 4・ 1
‘・, I
Jukkoku 15
10
(un) ωμロ刑WH門凶川相ohHω』Eコロ刷ohυロωヨげωhH』
Lr> c:;:) 色r> c::l Lr> ε:=r U"'") c;:, u""で. c:;:) u"、 cニコ u"、 C) Lf") c:::l Lr、 C:I Lr> Cコ Lr> ζ=コ u'・)C:;:)Lr>Cコ u"、 定:::0 ‘r、
ぐ、4 LCI C"- c::::l C、、I lrヲc-司C二:l C、、a‘r> c‘- = C、-4 Lr> C‘- c:コe、. Lr> C、- c:コc、." l..C) r- C::JI C""叫Lr> C、- Cコc、叶ーーー・ ,ー... - - c'、I c:、I C、I C:、I (""") C守") (〒) (""") 句CT 咽t- ...-:t. _ u"で, Lr) lI) LC') 巴.D<.D(..C)<.D��
い
mol 02/dm2/hr)hybrid Oxygen evolution
Distribution of oxygen evolution in B1F1 population of Jukkoku/Naxi//Jukkoku.
F i g. 21.
80
f旬、
訟�
、、J
∞
H 牛
吋 ロJ
。4 斗� 15
。 ω い
23
ロ 10斗イ
。
td ω ua コ h
J j
-
J 5h H
Fl
1 4・ 1
2 司. I
Jukkoku -
巴ニコ c:::> c二�c:=:Jt=)c::::Jc:lC::コ E二) = C:::> C:::> Cコ ε::::> cコ c::コ � c:::> c=> c:> ε=コ
= c、サ司- uコロコE二) c'司司すt..OCOC=>C司.q- uコCD c::::> c'叶唱す<..D co C:::>
Fヲc--,
...
ヲ c--, c"->司才、す『す司才司才Ln LLラLCで. 1.0 LCラ(..c)(.Q (..c) (.Q (.Q r-ーOxygen evolution (;;rnol 02/ drn2 /hr)
Fig. 22. Distribution of oxygen evolution in B1F1 hybrid population of Naxi/Jukkoku//Jukkoku.
て遅れる個体があり出穏期の変異が大きくなり、 同一の生育ステージにおける 酸素放出量の正確な測定が困難になったためである。 そこで、 まず二番目の原 因を明らかにするため、 F2雑種集団を構成する個体の生育時期を揃えて酸素放 出量を測定して、 詳細な分析を行おうとした。
第2節 個体の生青時期を考慮した酸素放出量の遺伝分析
前節では F2集団やB 1 F 1集団を用いて酸素放出量を特定の一時期のみ測定して 遺伝分析を試みたが、 その遺伝機式は簡単ではなかった。 そこで、 酸素放出量 の遺伝分析をさらに正確に行うため、 F2雑種集団を構成する個体の生育時期を 知る必要があった。 F2個体の生育時期を追跡する方法としてもっとも確実な方 法は個体ごとに出穏期を調査することである。 各個体の出穏期から逆算して個 体の生育時期を揃えることができると考えた。 そこで、 F2雑種集団において 生 育時期を揃えて酸素放出量を比較する実験を行なった。
材料および方法
窄葉膏8号(高放出量) x日本の改良品種玉錦(低)のF 1個体およびF2雑種 集団を用いた。 これらを両親とともに1986年5月8日に播種し、 6月10日に圃 場に1株1本植えした。 酸素放出量を第1期:7月7日,..._ 10日(移植後27,..._ 30 日)、 第2期:7月26日,..._29日(同46,..._ 49日)、 第3期: 8月2 5 日,..._27日(同 76,..._ 7 8日〉の3期に測定した。 測定数は両親、 F 1が12個体(反復)、 F2集団が 反復なしで295個体とした。 各測定時期で用いた葉身は主稗の最上位完全展開 葉であった。 あわせて F2集団の各個体の生育時期を把握するため個体別に出穂 期を観察
・
調査した。82
結果および考察
第1期の測定では、 両親の窄葉青8号、 玉錦の酸素放出量はそれぞれ 76 1、
592μmol 02/dmL/hrであり、 F
1の値は低放出量親の値とほとんど同じ591μmol
02/dm2/hrであった。 F2雑種集団の放出量の変異は、 最低値が269μmol02/dm2 /hr 、 最高値が1. 387μmol 02/dm2/hrであり、 両親の値より超越した個体がみ られ、 その分布型は正規分布に近かった(Fig. 23a)。第2期の測定では、 両親、 F 1、 F2集団ともに第1期の値より低下したが、 F1
の値は低放出量親の値に近く、 F2集団における酸素放出量の変異の分布型は第 1期と同じように正規分布に似ていた(Fig.23b)。 第3期の測定においても F2集団の放出量の変異は正規分布に近かった(Fig. 23c)。
したがって、 いずれの測定時期においても酸素放出量の遺伝様式は単純では なく、 これは第1節で得られた結論と同じである。
しかし、 F2雑種集団には約50日の出穏期の差があった(Fig. 24)。 すなわ ち、 幅広い生育時期の変異があったと判断してよい。 すでに、 第3章第5節で 述べたように葉令が進むにつれて酸素放出量が大きく低下し、 安定した測定に は生育時期の特定が不可欠であった。 供試するF2雑種集団の酸素放出量の遺伝 様式を解析するには、 まず、 生育時期を特定することが先決である。 しかし、
約 300個体のF2雑種集団のすべての個体について主稗の葉令を追跡することは 困難であるし、 長期間にわたって連続的にF2集団の酸素放出量を測定すること も現実的でないので、 以下に述べる方法で個体の生育時期を揃えて変異の解析 を試みた。 F2集団のそれぞれの個体の出穂期から出穂前35日を算出し、 その日 にもっとも近い測定時期における放出量をF2各個体の修正値とした。 すなわち、
出穂の早い個体は第1期の測定時期の値を、 出穂の遅い個体は第3期の値を修
正値とした。 また、 両親、 F1も出穂期からそれぞれ修正値を求めた。
その結果、 Fig. 23dに示したように、 F2集団の修正した酸素放出量の分布型 は明らかに二頂分布であった。 この分布型からみて単遺伝子分離の可能性があ
a
•
!)hl1se J.
(.JlIly 7th-10th)
J>ltasc 11 ( .J 1I 1)' 26 t h -29 t h ) 15
nu l
5
。 15
nu
のJFC伺}丘一一OいωDEコ己」ohυcωコσω」L(日)
b
C
d Iコhl1SC II[
(八lIgust 25th-幻th)
• P1 StandarJizcJ
vílllICS
x�
1=3.18nu l
0 15 5
5
0 10 5
11Hoo
oco--ωド∞
的ド∞ltH凶∞
O凶∞}tωN∞
旧N∞1IHO∞
00∞lt@ドド 旧ドドllHmド omドll@Nト 的NドitHOド1l
och-rωk
mhω!日由γ
02lωN匂m
mNωlHS
A ooui-ωド的2
rm・-【目的
o
ommllCNm
J山
mNmlHom
叩
oom--ωドザ
'hvmドザlHmマnOのすltω刊す・MmNすlHO守川0072{匂ド門m
mド門lH旧門
eO的門11ωN門n
mN門1HO門
PCO門luトNり
川トNl-mN
OomN
11市UN
N
旧引引11-ON OO引}{ωh-
mド-ltHm-
Oの-l己目
的n-lf-c-
oo--tCド
。
the cross Distributioll of oxygen cvolution in the f2 populaLion of
between ZaiycQin 8(Pl) and Tamanishiki(P2) f i g. 23
8/1-
50
40
30
ι」ω
F吋ロ4 0..
4-;
20
。
トωィ iコ Z
510
。 守 ιo
開
∞522252竺5353誌記FSR F 門 的h 小 二n lf)ヒE日R;qRR 凶
Heading date
Fig. 24. Dislribulion of heading date in the F2 populalion of the cross between Zaiyeqin 8 and Tamanishiki.
ったので、 551μmol 02/dm2/hrを境に2群にわけ、 単遺伝子分離の期待比で検 定を行なった。 χ三値は 3. 18であり、 単遺伝子分離によると考えてもよい結果 であった。 すなわち、 低放出量が優性、 高放出量が劣性といえる。 また、 修正 したF 1の値は修正した低放出量親の値に近かったことを あわせて考えると、 酸 素放出量は低放出量が優性の単遺伝子によって支配されていると推定できる。
光合成能力の遺伝的支配に関する報告は実験の困難さのためか非常に少ない。
すでに、 Hayashi et al. (1977)はイネ雑種集団を用いて、 炭酸ガス交換速度 からみた光合成能力は低い能力が優性の単遺伝子分離をすると報告し、 光合成 能力の遺伝的改良が可能であると示唆した。 鮫島(1985)はイネ雑種集団の炭 素同位体分別能を測定し、 弱い分別能が優性の単遺伝子分離にみえる結果を得 た。 測定方法は異な ってもHayashi et al. (1977)や鮫島(1985)の報告は、
本実験において酸素放出量を測定して得た低放出量が優性の単遺伝子によって 支配されているという結果を裏付けるものである。
以上述べたような光合成能力が単遺伝子で支配されるという結論は他の作物 でも報告がある。 トマトで、 Augustine et al. (1976)は炭酸ガス固定能力が 単一の遺伝子に支配され、 しかも低い能力が優性であることを明らかにした。
方、 光合成能力は主働遺伝子支配ではなくて、 微動遺伝子で支配されると いう報告がある。 Izhar and Wallace (1967)は、 インゲンマメで光合成能力 の遺伝は量的であるが、 関与する遺伝子数は比較的少なく、 低光合成能力はあ る程度優性であると結論した。 小島(1971)はダイズの農林1号とHarosoyお よび満州とHarosoyの2組合せでF
1にはヘテロ シ スがなく、
低光合成能力が優 性であると述べ、 光合成能力が単純な遺伝をしないと報告した。 Harrison豆1 al. (1981)はダイズの群落光合成能力の遺伝力は41 %であるとした。 また、Wiebold et al. (1981) もダイズの炭酸ガス交換速度の広義の遺伝力は36完ま たは56%であったと報告した。 Ellison et al. (1983)はコムギで止葉の光合 成能力には相加的な遺伝子の効果がとくに重要と結論した。 Heichel and Mus-
86
grave (1969)、 Monrna and Tsunoda (1979)はトウモロ コシで光合成能力の 著しいヘテロシスが遠縁品種閣の単交雑で認められると報告した。 Crosbie et al. (1978)はトウモロ コシで炭酸ガ、ス交換速度に有意なヘテロシスを観察し、
炭酸ガス交換速度は核内遺伝子の相加的作用に大部分支配されるとした。 ペレ ニアルライグラスでWilson and Cooper (1969)は光飽和状態の光合成能力で は相加的な遺伝分散が重要な要因であるが、 光制限状態では非相加的遺伝分散 と相加的遺伝分散の効果は同じであるとした。
本実験ではF 1の酸素放出量にヘテロシスを認めなかったが、 村山ら(1982) はイネにおいて7組合せ中6組合せでF 1の光合成能力が中間親より高いことを 報告した。 また、 坂(1985)はイネのクロリナ×正常種のFl個体で光合成暗反 応の炭酸固定初発反応を触媒する酵素RuBPC(リブロース二リン酸カルボキシ ラーゼ)活性は生育期間をとおして両親より高い傾向があり、 RuBPO (リブロ ース二リン酸オキシゲナーゼ)活性は逆に 両親より顕著に低く、 酵素面からみ た光合成、 光呼吸のヘテロシス的現象があることを報告した。 玄ら(1989)は 雄性不稔系統を利用したF 1雑種で若い止葉で得られる光合成速度にはヘテロシ スがみられないが、 登熟期の止葉にはヘテロシスがみられるとした。 しか し、
Yarnauchi and Yoshida (1985)は35組合せのイネF
1雑種の光合成能力、
葉面 積、 分けっ力などを調べて、 F 1には光合成能力のヘテロシスは表れず、 旺盛な 分けっと早い葉面積の展開がF 1強勢をもたらすと報告した。 イネのF 1雑種にお いて光合成能力のヘテロシスが表れるかは不明で あり、 今後の研究が必要で あ る。本実験結果はF2雑種集団において高放出量個体を選抜することによって、 後 代で高放出量系統を作出できる可能性があることを示す。 とくに高放出量は劣 性 遺伝子に支配されると考えられるので、 比較的短期間に高放出量固定系統を
確立できると考える。
出穂期で個体の生育時期が揃うように修正を行ってもF2集団の中には両親の
値から超越した個体が若干出現したが、 これは測定時期数が少なすぎてその個 体の生育時期を他の個体と揃えること ができなかったためと考える。 一連の実 験ではいずれも類縁関係の遠い品種を用いて酸素放出量の遺伝分析を試みたが、
両親の遺伝的背景は大きく異なっている。 これも光合成能力の遺伝分析を困難 にしている理由のーっといえる。 遺伝的背景 がほとんど同じで酸素放出量のみ 大きく異なる遺伝系統を用いて測定することにより、 詳細な遺伝分析 が可能と なると考える。
第3節 F"系統における酸素放出量の遺伝分析
前節ではF2世代における酸素放出量の遺伝様式を調べ、 生育期間が大きく拡 散するF2雑種集団では個体の生育時期を考慮すれば遺伝分析が容易になり、 単 純な遺伝様式となる場合があることを述べた。 本節ではわ世代とF3世代聞の酸 素放出量の遺伝相関を調べ、 さらに、 光合成能力の遺伝的機作を詳細に知ろう とした。
材料および方法
窄葉青8号×信州金子および十石×納西の二つのF2雑種集団から酸素放出量 に関してそれぞれ1 0個体の高放出量と低放出量個体を選抜した。 1個体のF2 から由来する50個体のF3系統を198 6年5月8日に播種し、 6月10日に園場に1 株1本植えした。 これらの高放出量と低放出量F3系統についてそれぞれ10個体 を無作為に選び、 7月30日から8月5日にかけて酸素放出量を測定した。
結果および考察
窄葉青8号×信州金子のF2個体の酸素放出量とその個体に由来するF3系統の 系統内平均酸素放出量との関係を示した(Fig. 25) 0 F3系統には系統内放出
�
ωω ロ J判 叶
**
γ=0.646 600
500
400
300
(主\NEガ\NO叶05ミゾ O叫ん 3
内』dJ門口O叶υコHO〉ω
ω〉ω∞伺いH〈ロω∞A×o
1,000 800 900
Oxygen evo1ution of F? individua1s
ヤ
mo1 02/dm2/h�) 600 700400 500
ReJalionship belwecn oxygen evolutioll in the P:2 and P3 generalion of lhe cross belween Zaiyeqin 8 and
Shinshu-kaneko.
25 F i g.
lines show slandard deviation of oxygen evolution
nH
nH nu 0 .,i ・ 1 aIL 十し 内d 内d ri
rA
ρu nu nH nH PU
凸U 円る
円る
A1ι
'JιゎドanドAnH
nH JU ・ 1A ・ 1A
,パU 凸U nu nH nH --BA - TA n--ι nfi pu ρU JU
Au
pnv u
Hu nu
o r r 円る
円る
vd
vd 4L
a,Blv ・ 唱Ea・ -
1i pL C 内d 3u nν
na 円d nd pu
FL 、a
wn 円6
0 ・ l luHH
今、-ゎドA- M 0 ・
J i nes Verlical
量の変異の大きさを表すため標準偏差を示した。 F3系統の平均放出量はF2個体 の値より低かった。 この理由はF3世代における酸素放出量の測定時期がF2世代 における測定時期より遅く、 第3章第5節の実験で明らかになった生育時期の 進行にと もなう放出量の低下現象によると考える。 九世代で高放出量と判定し た10のF3系統の系統内平均酸素放出量は、 最低値392μmol 02/dド/hr 、 最高 値501μmol 02/dm.è/hrであり、 系統内平均値の系統聞における差は小さかった。
また、 2系統を除くと系統内の放出量の変異は小さかった。 すなわち、 高放出 であったF2個体に由来するF3系統の系統内平均酸素放出量は高かった。
一方、 F2世代で低放出量と判定した10のF3系統の系統内平均酸素放出量の変 異は、 最低値275μmol 02/dド/hr 、 最高値532μmol 02/dmι/hrであり、 高放出 群とくらべて系統閣の変異の幅が大きかった。 さらに、 F2世代で低放出量と 判定した個体から由来するF3系統の中には高放出量群と同程度の高い値をもっ 系統が出現した。 これは、 F3世代で低放出量と高放出量が分離したと考えるこ とができる。
十石×納西のF2個体の酸素放出量とその個体に由来するF3系統の系統内平均 酸素放出量との関係をFig. 26'こ示した。 この集団においては両者聞に有意な 正の相関関係(r=0.845料)が観察でき、 酸素放出量は遺伝力が高い形質であ ると推定した。 さらに、 窄葉青8号×信州金子の組合せにおいて、 F汁国体の酸 素放出量とその個体から由来するF 3系統の系統内平均酸素放出量との聞にはr=
O.
646料の正の有意な相関があった(Fig. 25)
0F2集団から選抜した高 ・ 低放出量個体とそれに由来するF3系統の酸素放出量 の関係をみると、 光合成能力は遺伝力が高い形質であると考えられ、 酸素放出
に関して高放出量および低放出量方向への選抜が可能であると結論した。
守0
ωωロ 什』[
•
. a・ - -
• 800700
♂ 汚
•。 600
500
**
r=0.845 400
(しHZ\
N 臼ガ\
NO 門』口刊ロOJ円以コ.HO〉ω ロω∞h×oω∞吋hH ω〉 〈 叶OEふ
。 300
OLヂ
100 700 800 900individua1s
600 500
Oxygen evo1ution in F2 いmo1 02/dm2/hr)
400 300
200
in F司 lhat Relationship between oxygen evolution in lhe F2 and generation of lhe cross between Jukkoku and Naxi.
26 Fig
in F2 generation.
group defined in F2 generation.
Low capacity group defined lIigh capacity
0
•
第6章 高
・
低酸素放出量系統の選抜前節まで に酸素放出量でみた光合成能力は遺伝的に支配されており、 その晶 低は測定値により判定が可能であるとの結論に達したので、 突然変異系統聞の 交雑分離後代を用いて高
・
低放出量系統の選抜の可能性を検討した。材料および方法
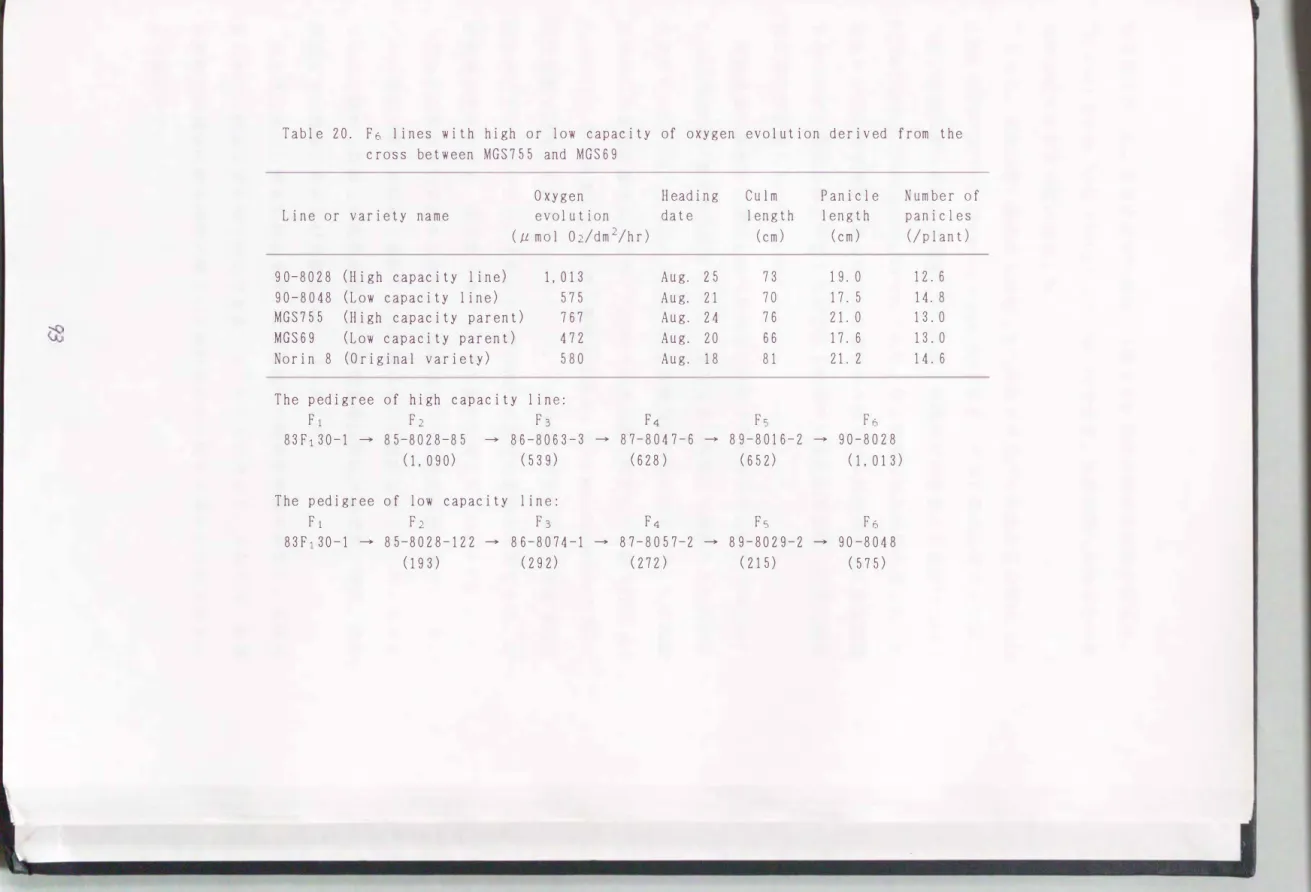
第4章第4節で酸素放出量を調査した農林8号の放射線人為突然変異系統13 5系統の中からMGS755 (高放出量、 Appendix 4の系統番号8291、 1983年測定値 569μrnol02/d ド/hr)およびMGS69 (低放出量、 Appendix 4の系統番号8226、
1 983年測定値155μrnol02/drn2/hr)の2系統を選んだ。 両親の酸素放出量と農 業形質をTable 2 0に示した。 MGS755とMGS69との交雑を行い、 F1個体、 F2雑種 集団を作出した。 F2雑種集団の酸素放出量を両親およびF 1とともに測定したo
F2集団の調査個体数は2 88であった。 酸素放出量はF2個体では1反復、 両親と F1は少なくとも5反復で測定した。 F2集団から酸素放出量に関して高放出量個 体を10個体、 低放出量を9個体を選んでF3系統を作成した。 F3世代以降は個体 を選抜の対象とし、 F6系統まで系統の確立を継続した。 各世代は一系統あたり 50個体を移植し、 そのうち無作為に10個体を選んで酸素放出量を測定した。
結果および考察
Table 2 1にF2から九世代までの酸素放出量の変化を示した。 F2世代は個体の 値を、 F3世代以降は系統内平均値を示した。
F2世代で高放出量と判定した群の酸素放出量の変異は、 924μrnol02/drnL/hr から1, 231μrnol 02/drn2/hrで、 一方、 低放出量群は172μrnol02/drnL/hrから 234μrnol 02/drn2/hrであった。
つぎに、 高放出量群のF2から由来するF3系統の酸素放出量の変異は326μrnol 02/drn2/hrから539μrnol02/drnL/hrであり、 すべての高放出量群F3系統は高い
?之
Table 20. F6 lines with high or low capacity of oxygen evolution derived from the cross between MGS755 and MGS69
Oxygen Heading Culm Panicle Number of Line or variety na me evolution date length length panicles
(μmol 02/dm2/hr) (cm) (cm) (/plant)
90-8028 (High capacity line) 1, 013 Aug. 25 73 19. 0 12. 6 90-8048 (Low capacity line) 575 Aug. 21 70 17. 5 14. 8 MGS755 (High capacity parent) 767 Aug. 24 76 21. 0 13. 0
\、JA、J MGS69 (Low capaci ty parent) 472 Aug. 20 66 17. 6 13. 0
Norin 8 (Original variety) 580 Aug. 18 81 21. 2 14. 6
The pedigree of high capacity line:
F'l F2 F3 F4 Fち F6
83F130-1→85-8028-85 →86-8063-3→87-8047-6→89-8016-2→90-8028
(1,090) (539) (628 ) (652) (1, 013)
The pedigree of low capacity line:
F1 F2 F3 F4 Fら F6
83F130-1→85-8028-122→86-8074-1→87-8057-2→89-8029-2→90 -8048
(193) (292) (272) (215 ) ( 575)
放出量であった。 低放出量群F2個体に由来するF3系統の酸素放出量の変異は、
273μmol 02/dm2/hrから391μmol 02/dm2/hrであり、 低放出量F3系統の中には 扇放出量を有する系統が出現した。
さらに、 高放出量F3系統から選抜したF4系統の系統内平均酸素放出量は 470 μmol 02/dm之/hrから 628μmol 02/dmL/hrであり、 すべて高放出量であった。
一方、 低放出量F3系統から選抜したF4系統の系統内平均酸素放出量は211μmol 02/dm2/hrから 537μmol 02/dm2/hrであり、 中に高放出量系統が出現した。 す なわち、 F2世代で高放出量と判定した個体から得られた後代はすべて高放出量 であったが、 低放出量と判定した個体からは後代に高放出量系統と低放出量系 統の両者が分離した(Table 21)
高放出量F4系統から選抜したFs系統は2系統ともにそれぞれ652、 536μm0 1
02/dm2/hrで高い酸素放出量であった。 低放出量F4系統から選抜したFs系統は それぞれ215、 329、 337μmol 02/dm2/hrと低酸素放出量に固定した。 F2世代選 抜時点では低放出量であったが、 F4世代で高放出量と判定した系統(537μmol
02/dm2/h け から選抜したFs系統の酸素放出量は、 414μmol 02/dm乙/hrであり、
高放出量で固定していることがわかった。 この系統の草型は低放出量親MGS69 型のそれとよく似ていた。 すなわち、 雑種後代に低放出量親の草型をもち、 高 酸素放出量を有する、 組換型と考えられる系統が出現した(Table 21)
二つの高放出量Fs系統から選抜したF6系統はすべて高放出量であり、 一方、
一つの低放出量Fs系統から選抜したF6系統はすべて低放出量であった。 このよ
うにして選抜を継続した高放出量および低放出量九系統の出穂期、 得長、 穂、長、
穂数などの農業形質および来歴をTable 20に示した。
以上の結果は、 酸素電極法を用いて雑種後代の酸素放出量を測定し、 高放出 あるいは低放出量系統を作出できることを示すものである。 すなわち、 育種 の過程で酸素放出量を選抜形質として光合成能力に関する選抜が可能であるこ とを意味する。
守午
Table 21. Changcs of oxygcn cvolution during F2 to F6 gcnerations of the cross bctwccn MGS755 x MGS69
No
8.
10.
o.
11 12.
13.
1�
15
16
17
18.
19.
MGS755 MGS69
_Bigh evo�Jlon group in F2 generation
Valuc in ^vcragc valuc ^vcragc valuc ^vcraged value ^vcragcd valuc ドi plants inド" 1 i ncs in FI\ 1 incs i nドs 1 i ncs in F6 lines
( 1985) (1986) (1987) (1989) (1990)
1, 231 一一一326
1, 100 一一511
「
517(O)605 (0) 588 (0)525 (0) 1. 013(0)古
o -_-- 539
[_�
551 (0) 622 (0) 628 621 (0) (0) ー一一 652 (0) 536 (0)E
1,005(0) 1,026(0) 997 (0) m5{O)F
m(O)1. 0 17 �93 1.024(0)
1. 021 3 � 1 1.042(0)
988 112 959 (0)
982 一一一 503 -
I
-- 528(O) 978 (0)505 (0)
959 �98
949 539
E
470(O) 477(0) 560 (0)924 440
Low evolu上辺旦group_lB F2 generation
Value in ^veragc value ^verage value ^veraged value ^vcraged value F〆plants in F3 1 ines in F4 lines in F5 1 ines in F6 lines
(1985) (1986) (1987) (1989) (1990)
234 一一一一318
234
叩 E 269 (.) 534(.) l川
346(.) 234一一一一- 391
m m
E
310 (.) m- -414(.)266 (.) 211(.)
川 - 277
一仁
251 (・)川)306 (.) 278(.)
E
4M(.)1こ
211働218(.)r--
5558(.) 明 会231 (・)
r--
533(.)3 ---292
l 三
272{.)I 三
215(.)228 (・) I___ 337(・)
r--
506(.)244(.) ロ9(.)
「
438(.)193 一一一一 361
f---
529(.)172 一一一 323
r--
510(.)539(.)
786 579 497 767
375 - 250 215 472
Unit of oxygen evolution is μmol 02/dmL/hr
(0) Plant type of MGS755. (・) Plant typc of MGS69.
Underline the recombinant lines having high evolution and planl lype of MGS755.
カ Eslablished 九1ines with high or low capacity of oxygen evolution
作物の育穫において光合成能力が実際に選抜の対象とされたことはなかったo Zelitch and Day (1973)はタバコを用いて光呼吸速度に関して高、 低能力系 統の選抜を継続したところ光呼吸を約半分に減らした系統は光合成能力が38%
増加したとして、 光呼吸速度による高光合成能力系統の選抜が可能であること を示した。 小島(1971)は2組合せのダイズのわとF3世代の炭酸ガス交換速度 においてr=0.45とr=0.66の相関を得て、 選抜が比較的効果があると報告した。
また、 Mahon and Hobbs (1981)はエンドウで高、 低光合成能力品種のダイア レル交雑を行い、 O.70の遺伝力を得て選抜は比較的効果があるとした。 栄養繁 殖期間における光合成能力の選抜はダイズやトウモロ コ シで効果があるという 報告がある( Secor弘主上198 2、 Moss and Musgrave 1971)。 一方、 Wiebold et al. (1981)はダイズで両親、 F 1、 F2の光合成能力の遺伝分散から遺伝力を 算出し選抜の効果はないとした。
光合成能力に関する選抜実験はイネでは行われていない。 本実験結果は雑種 後代の酸素放出量を測定することにより高、 低放出量後代系統を確実に選抜で きることを示したものであり、 高能率光合成能力系統の選抜
・
作出が酸素電極 法で可能であることを明らかにしたものである。テ6
第7章 総合考察
1 . 光合成多点測定装置の重要性
植物の光合成能力に関する種問、 種内変異の探索や遺伝分析は高能率光合成 系統作出のためのもっとも基礎的な研究である。 このためには、 多数の試料の 光合成能力を簡便、 迅速、 安定して測定できる方法を確立する必要がある。
赤外線ガス分析装置はさまざまな環境条件下におけるインタクトな同化器官 の光合成能力の変化を解析するためには有効な機器であるが、 短時間に多点数 を扱うような測定には不向きである。 一方、 酸素電極法は小型で構造が簡単で あり、 Jones and Osmond (1973)が初めて酸素電極法をワタの光合成能力の測 定に用いて以来、 オオムギ、 イネなどの高等植物でこの方法による光合成能力 の測定が試みられている(Ishii et al. 1977、 Yamashita立主豆1. 1978、 山末
・
植木1978、 石原ら1979、 坂・
千坂1981a、 1981b、 玄ら1981、 Hyeon旦 a1. 1982、 伊藤1982、 Delieu and Walker 1983)。 この利点、に加えて本測定 方法は液相中で測定を行うので、 光合成能力の大きな律速要因と考えられる葉 肉細胞内の炭酸ガス拡散に関する気孔抵抗や気孔開度の影響を除いた真の炭酸 ガス固定能力を推定できるとされている(Pitman立主主1. 1975、 石原ら1979)。そこで、 酸素電極法は 多点数の光合成能力の測定に最適であると考え、 6点 の酸素放出量を同時に測定できる装置を新しく2台開発し、 装置の実用化と育 種への応用の可能性を図った。 1回の測定は約40分で終了し、 2台の装置を用 いると1日に 150点の酸素放出量を測定できるようになった。 こ れにより、 酸
素放出量の遺伝変異の探索や遺伝分析が大きく前進したと考える。
2. 葉片前処理法の開発と測定条件の決定
多点数の材料を扱うという前提条件を満たすために測定前日に葉身を園場か ら採取し、 葉片を作成した後に一定の環境条件下で1日間保存するという前処
理方法を考えた。 すなわち、 葉身を採取後ただちに葉片を打ちぬいてシ ャーレ 内の蒸留水に浮かべ、 光照度4万ル yクス、 照明が午前6時から午後8時、 思 温が280Cで夜温が220Cの人工気象室で24時間インキュベートする葉片前処理方 法は、 午前9時から午後5時までの長い時聞にわたり安定して高い酸素放出量 をもたらすことがわかった。 また、 この前処理を行ったほうが当日葉身を採取 し葉片を作成するよりも高く安定した放出量になることを明らかにした。 1日 間の測定値の変動係数の変異は12.6 %から23.7 %であり、 この値は他の農業形 質の変動係数の大きさ( 1穂籾数: 15.4% ; 8.5%、 穏数: 23.8% ; 14.4%、 穂重
:
15. 0 % など)と比べるとほぼ同程度の大きさであり、 酸素電極法による酸素 放出量は光合成能力の遺伝分析に十分に適用できる精度であるといえる。 これ までの酸素電極法によるイネ光合成能力の測定では本論文のような葉片前処理 方法は行われていない。 この前処理方法の開発こそ実験精度を向上させ、 多点 数測定を可能とする重要な点であった。 これにより以下の遺伝変異の探索および遺伝様式の解明の実験に進むことができた。
3
.
光合成能力のイネ種内変異について光合成能力を改良するためには、 イネ種内における光合成能力の遺伝変異を 明らかにすることがまず重要である。
すでに炭酸ガス交換速度でみたイネの光合成能力の最高値/最低値比として
1. 61 (村田1957)、 1.70 (秋田1980)、 1. 80 (Anonymous 1968)が報告され ているが、 今回、 酸素放出量で光合成能力の種内変異を調べたところ、 アジア 栽培イネには最高値/最低値比が1.7 9という大きな変異があることが明らかに なった。 この変異の大きさは、 本実験で供試した材料がアジア各地から広く収 集された品種であり、 供試数も多く、 アジア栽培イネの遺伝的多様性を網羅し て選ばれたためであろう。 この最高値/最低値比はアジア栽培イネにおける光 合成能力の遺伝的変異の程度を示すと考えてよいであろう。 この変異の大きさ
98
から考えてイネ種内の高放出量品種を利用して高能率光合成系統を作出できる 可能性があると判断した。 また、 突然、変異処理によって原品種の酸素放出量よ り150%も増加した放出量を持つ系統が得られており、 在来品種のみでなく突然、
変異系統を高能率光合成系統作出のための遺伝資源として利用できると考えら れる。
4. イネ光合成能力の遺伝的分化につい て
栽培イネの大 多数を構成するアジア栽培イネ(立工正三豆s a t i v a L.) ,こはイン ディ力、 ジャポニカという亜種レベルでの分化があり、 各地にさ まざまな品種 が存在する。 これまで、 光合成能力のイネ品種群における変異を研究した例は 少ない。 鮫島(1985)は炭素同位体分別能を測定し、 ジャポニ力、 ジャパニカ 品種群がインディ力、 シニカ品種群より分別能が弱いことを報告した。 また、
Cook and Evans (1983)はジャパニカ品種群が炭酸ガス交換速度が他の品種群 より低いことを報告した。 異なる光合成測定装置を用い たものの、 本実験結果 は、 ジャパニカ品種群の光合成能力が4品種群の中でもっとも低い点で、 Cook and Evans (1983)の結果と一致した。 鮫島(1985)の実験でも光合成能力の 品種群間差異は観察されてい る。
以上から、 アジア栽培イネの光合成能力には品種群でも差があることが明ら かになり、 今後、 高能率光合成能力イネ系統の作出にあたってその遺伝資源と して有用な品種群を特定することができた。
5. 多収性イネ育種の方向
イネの多収性品種の開発は育種目標の大きな柱であり、 これまで日本、 1RR 1、
韓国などでは草型育種 (角田1964)といわれる群落構造の改善により多収化 が追及されてきた。 その改善にあたって半倭性遺伝子をもっ日本の在来品種十
石や中国の在来品種低脚烏尖が果たした役割は大きいとされる(菊池ら 1985)。
十石の半媛性遺伝子を導入して育成されたホウ ヨ夕 、 シラヌイ、 コクマサリが 短稗で耐倒伏性をもち九州の収量水準をひき上げた例 (岡田ら1967)、 低脚 烏尖のもつ半綾性遺伝子を導入して育成された1RR 1の品種 1R 8が東南アジア各 国の収量を飛躍的に向上させた例(吉田1976)、 韓国での統一系品種による 収量性の顕著な増大(金1979、 Chung and Heu 1980)は有名である。
わが国のイネ育種では穂数型品種の育成によってシンク容量を増やしたが、
これを一層増加させるため現在わが国ではインディカ品種から大きなシンク容 量(総籾数x 1籾重〉を日本品種にとり入れようとしている。 穂重型で半媛性 の品種が育成された場合lこはソース量(葉面積×葉の光合成能力)の不足が問
題となり、 質的な改良、 すなわち、 単位葉面積あたり光合成能力を遺伝的に向 上させる必要がでてくると考える。 今後さらに多収性育種を推進するためには、
群落構造の改善による群落光合成能力の改良のみならず、 光合成能力の向上に も重点がおかれるべきである。 その理由は、 韓国で育成された多収品種密陽23 守、 水原258号などは極短稗で幅の広い直立葉が空間的に効率よく配置され、
多収のための理想、草型を有しており(武田ら1984)、 草型改良の点では限界 に来ていると考えるからである。
6. 光合成能力の遺伝子解析について
光合成能力の改善を進めるためには、 その遺伝様式を明らかにする必要があ
る。 品種聞の組合せのF2雑種集団、 81F1集団における酸素放出量の変異の分布 型は一時期の測定値を用いた場合、 いずれも正規分布に近い分布型となり、 単 純な遺伝様式を推定することは困難であった。 これは集団内の個体の生育時期 に大きな変異があり、 放出量の正確な変異を捉えることができなかったためと 考える。 そこで、 各個体の出穂期をもとに生育時期を揃え放出量を修正した。
その結果、 F2個体の修正した値は二頂分布となり、 また、 F1植物の修正値は低 放出量親の修正値に近かったことから低放出量が優性の単遺伝子分離をすると
100
結論した。 さらに、 F2個体の酸素放出量とその個体に由来するF3系統の系統内 平均酸素放出量との聞には正の相関があり、 遺伝力が高かった。 このことはF2
雑種集団において高放出量個体を選抜して後代に高放出量系統を作出できるこ とを示す。 また、 高放出量は劣性遺伝子で支配されるので短期間に固定系統を
確立できると考える。
以上の結果は品種閣の組合せを用いて得たものであるが、 遠縁な両親から養 成した雑種集団はそれを構成する個体の生育時期や諸形質の分散が大きく光合 成能力の遺伝解析は容易でない。 そこで本実験で行ったような個体の生育時期 を揃えるなどの処理が必要になる。 高放出量は劣性と考えられるので、 実際の 育種場面では酸素放出量の高い個体をF2世代で選抜し、 以降の系統で高放出量 を繰り返し選抜すれば比較的容易に高放出量後代系統を作出できょう。
光合成能力の遺伝分析を今後詳細に行うためには遺伝的背景が同一で酸素放 出量や炭酸ガス交換速度が異なる遺伝的変異系統を作出する必要がある。 第6
章における選抜実験は光合成能力lこ関する遺伝解析の材料を作出できる可能性 を示したものであり、 酸素電極法による葉身酸素放出量の測定で高能率光合成 能力をもっ系統の選抜
・
育成が可能なことを示すものである。摘要
イネの多収性品種育成を目的として、 単位葉面積あたりの光合成能力の種内 変異、 遺伝様式および高能率系統選抜の可能性を酸素電極法を用いて研究した。
以下にその概要を述べる。
1 )イネの個葉光合成能力の種内変異と遺伝分析を行うため、 酸素電極法を用 いた多点式光合成測定装置を新しく設計
・
開発した。2 )酸素放出量は葉身の採取時刻の経過にしたがって減少した。 そのため、 採 取時刻に関わりなく品種の遺伝的能力を反映した酸素放出量となる試料の前 処理方法が必要であった。
3 )葉身採取後ただちに葉片を打ち抜き、 蒸留水を入れたシ ャーレに浮かべ、
光照度4万ル yクス、 照明が午前6時から午後8時、 昼温が280Cで夜温が2 2 OCの人工気象室で 24時間インキュベー卜する葉片前処理方法を考案した。
4 )この前処理法は測定材料を栽培する圃場の気象条件にかかわらず安定した 値をもたらし、 測定日ごとの酸素放出量の変動係数は、 他の農業形質とほぼ
同じ程度であった。
5 )イネ葉身酸素放出量の最適測定条件は照射光照度7万ル ックス、 温度目。C、
HEPES緩衝液のpH 7. 2であった。 葉身中央部の葉片の表側を測定lこ用いること にした。 また、 主稗の最上位完全展開葉を用いることにした。
6 )酸素放出量は葉令が増加するにつれて低下した。
7 )新しく開発した多点式酸素放出量測定装置2台と葉片の前処理方法とを用 いて少なくとも1日 150点の酸素放出量の測定が可能となった。 従来の測定 方法では困難であった短時間における多点数測定を達成し、 光合成能力の遺 伝的変異や雑種集団を用いる遺伝分析を容易にした。
8 )アジア栽培イネの酸素放出量の変異の大きさを最高値/最低値比で表すと
1. 79となり、 光合成能力に大きな種内変異が存在した。
9 )アジア栽培イネの4品種群間の中で、 ジャポニカ品種群の酸素放出量はも
102
っとも高く
、
一方、 ジャパニカ品種群の平均値はもっとも低かった。 光合成 能力に関する品種群間変異が存在した。10)中国雲南省およびラオスの在来品種において、 ジャポニカ品種群に多くみ られるエステラーゼ同位酵素の遺伝子型6型( E st-11、 E st-20、 E st-31 )の 品種は酸素放出量がもっとも高く、 一方、 ジャパニカ品種群に多くみられる 遺伝子型8型( E st-10、 E st-21、 E st-31 )、 12型( E st-10、 E st-20、 E st-31 ) の品種は酸素放出量が低かった。 すなわち、 中国雲南省やラオスという限ら れた地域に存在する品種に焦点をあてた場合も光合成能力についてアジア栽 培イネの分化に一致する遺伝的分化を認めた。
11)日本の改良品種は在来品種より酸素放出量が高かった。 コシヒカリの系譜 の酸素放出量から、 育種の過程で光合成能力がおおむね向上したことがわか った。
12)人為突然変異処理によって酸素放出量に大きな変異を作出できた。 最高値 /最低値比は8. 41であり、 この値は在来品種を用いて得られた値(1.79)に 比べではるかに大きかった。 光合成能力lこ関する高能率系統の給源を人為突 然変異処理にも期待できると結論した。
13)酸素放出量の高い品種と低い品種の2組合せの正逆F2雑種集団および2組 合せのB1Fl集団を用いて特定の一時期における酸素放出量を測定して遺伝分 析を行なった。 F1の酸素放出量は低放出量親の値に近かった。 F2雑種集団の 中には両親の値より超越した個体があり、 その分布型は正規分布に似ていたo B1F1雑種集団においても酸素放出量の分布型は正規分布に近かった。 すなわ ち、 F2集団やB1F1集団を用いである特定のー測定時期の値から酸素放出量の 遺伝様式をとらえることは困難であった。
14 )窄葉青8号(高放出量) x玉錦(低放出量)のF2雑種集団の酸素放出量を 3時期測定し、 出穂期をもとにF2個体の生育時期を揃えて酸素放出量の変異 の解析を行なった。 修正した酸素放出量は明らかな二頂分布となり、 低放出
量が優性、 高放出量が劣性の単遺伝子分離であった。
15)窄葉青8号×信州金子(低放出量)の組合せにおいて、 F2世代で高放出量 であった個体に由来するF3系統の系統内平均酸素放出量は高かった。 F2世代 で低放出量と判定した個体から由来するF3系統の中には、 高放出量群と同程 度の高い値となる系統が出現したので、 F3世代で低放出量と高放出量が分離 したと結論した。 F2個体の酸素放出量とその個体に由来するF3系統の系統内 平均酸素放出量との聞には r=0.646**の正の相関があり、 酸素放出量は遺伝 力が高い形質であると結論した。
1 6 )農林8号に放射線を照射して得られた突然、変異系統聞の交雑組合せから酸 素放出量で438μmol 02/dm2/hrの差があるが、 農業形質がよく類似した高酸 素放出量F6系統と低放出量F6系統が得られた。 この結果は酸素電極法による 葉身酸素放出量の測定で高能率光合成能力をもっ系統の選抜が可能なことを
示すものである。
以上の結果にもとづき、 酸素電極法による単位葉面積あたりの光合成能力を 改良する立場から、 多収性イネ品種育成の可能性を論じた。
/0斗
謝辞
本論文をまとめるにあたり、 原稿校閲の労をとっていただきました九州大学
・
農学部教授岩田伸夫博士、 同学部教授豚和一博士ならびに同学部教授奥達雄 博士の諸先生がたに対しまして厚く感謝いたします。この研究を行うにあたり、 終始あらゆる面でご指導くださいました農業生物 資源研究所
・
遺伝資源第一部・
前植物探索導入研究チーム長、 現遺伝資源第一 部長中川原捷洋博士に心から感謝の意を表します。 酸素電極法についてご指導 くださいました元農業技術研究所遺伝第七研究室長川上潤一郎博士、 同研究室 元主任研究官、 現植物探索導入研究チーム長奥野員敏博士ならびに同研究室元 任研究官、 現宮崎県総合農業試験場育種科長滝田 正博士に厚くお礼申しあげます。 また、 植物探索導入研究チームの同僚、 河瀬真琴博士、 江川宜伸博士、
勝田真澄主任研究官ならびに非常勤職員、 浅野文子さん、 片倉和枝さん、 真中 明美さんのかたがたには実験の遂行にあたり多大な協力と助言をいただきまし た。 ここに記して深く感謝いたします。
引用文献
Anonymous 1968. IRRI Annual Report. Plant physiology: 17-42.
秋田重誠1980. 作物の光合成、 光呼吸の種間差. 第I報 光合成、 光呼吸およ び生育の酸素濃度に対する反応の種間差. 農技研報 D3 1: 1 -58.
Augustine. J. J. and M. A. Stevens 1976. Genotypic variation in car
boxylation of tomatoes. Plant Physiol. 57: 3 25-333.
Augustine. J. J.. M. A. Stevens and R. W. Breidenbach 1976. lnheri- tance of carboxylation efficiency in the tomato. J. Amer. Soc.
Hort. Sci. 101: 456-460.
Bjork man. 0.. M. A. Nobs and J. A. Berry 1971. Further studies on hybrids between C3 and C4 species of A triple x. Carnegie lnst. Wash.
Year Book 70: 507-511.
Brugnol i. E.. K. T. Hubick. S. von Caemmerer. S. C. Wong and G. D . Farquhar 1988. Correlation between the carbon isotope discrimina- tion in leaf starch and sugars of C3 plants and the ratio of in
tercellular and atmospheric partial pressures of carbon dioxide. Plant Physiol. 88: 1418-1424.
Chung. G. S. and M. H. Heu 1980. Status of japonica-indica hybridiza- tion in Korea ln "lnnovate approaches to rice breeding". IRRI. Los Banos. Philippines: 135-152.
Cook. M. G. and L. T. Evans 1983. Some physiological aspects of the domestication and improvement of rice (ORYZA spp.). Field Crops Research. 6: 219-23 8.
Criswell. J. G. and R. M. Shibles 1971. Physiological basis for geno- typic variation in net photosynthesis of oat leaves. Crop Sci. 11:
106
550-553 .
Crosbie, T. M., J. J. Mock and R. B. Pearce 1978. Inheri tance of pho tosynthesis in a diallel among eight maize inbred lines from
lowa stiff stalk synthetic. Euphytica 27: 657-664.
Curtis, P. E., W. L. Ogren and R. H. Hageman 1969. Varietal effects in soybean photosynthesis and photorespiration. Crop Sci. 9: 323-326.
Del ieu, T. and D. A. Walker 1972. An improved cathode for the measure
ment of photosynthetic oxygen evolution by isolated chloroplasts.
New Physiol. 71: 201-225.
Del ieu, T. 1. and D. A. Wal ker 1983. Simul taneous measurement of oxy
gen evolution and chlorophyll fluorescence from leaf pieces. Plant Physiol. 73: 534-541.
Dornhoff, G. M. and R. Shibles 1976. Leaf morpholo gy and anatomy in relation to C02-exchange rate of soybean leaves. Crop Sci. 16:
377-381.
Ell ison, F., N. F. Derera and D. G. Pederson 1983. Inheri tance of physiological characters associated with yield variation in bread wheat. Euphytica 32: 24 1-255.
Harrison, S. A., H. R. Boerma and D. A. Ashley 1981. Heritability of canopy-app arent photosynthesis and its relationship to seed yield in soybeans. Crop Sci. 21: 222-226.
Hayashi, K., T. Yamamoto and M. Nak agahra 1977. Genetic control for leaf photosynthesis in rice, Ory互互主主tiヱ豆L. J apan. J. Breed. 27:
4 9-56.
Hei chel, G. H. and R. B. Musgrave 1969. Varietal differences in net photosynthesis of �旦豆旦旦主� L. Crop. Sci. 9: 483-486.