ジベレリンの作用発現の分子機構に関する研究
中嶋 正敏
東京大学大学院農学生命科学研究科応用生命化学専攻
Studies on molecular mechanism for gibberellin
-triggered actions
Masatoshi NakajimaDept. Appl. Biol. Chem., Univ. of Tokyo
要旨 : Gibberellin (GA) regulates various developmental processes in plants. To study on some actions triggered by GA biochemically, we prepared antisera against GA. At first, we used the antisera for the detection of GA in plants. In developing seeds of morning glory and also of Arabidopsis, GA was immunologically detected in their integuments where starch granules were observed and then disappeared. Taking togeth-er with othtogeth-er result for related genes expression, we proposed that GA plays a role for seed maturation of dicotyledonous plants at least through the degradation of starch. Secondly, we established a biochemical method for a detection of water-soluble GA receptor candidates in crude
solu-tion, which led to the identification of a rice GA receptor, Gibberellin Insensitive Dwarf1 (GID1). Its GA-binding activity and ligand selectivity
were confirmed. The suppressor of GA signaling called DELLA boosted the GA-binding activity of GID1 in vitro, which means the formation
of a GID1-DELLA complex tightens the binding of GA to GID1. In Arabidopsis, we found three GA receptors. By using loss-of-function
mutants, we elucidated the functional redundancy and partial specificity of their receptors. After some approaches, we proposed that the pheno-type(s) appeared in the mutants depends on the abundance of the remaining receptor and on its preference to DELLAs.
1. ジベレリン抗体の調製と免疫組織化学的手法の構 築 植物ホルモンとしてジベレリン(GA)は,種子の発芽 過程や茎部の伸長を促進するが,現在においてはその分子 機構もかなり細部まで明瞭になりつつある.筆者が研究に 着手する時点では,オオムギやイネなど穀類の種子におけ る加水分解酵素の誘導系が,比較的単純な応答系として利 用し易い状況であったことから,他に知られる GA の応答 系と比較してより多くの情報が蓄積されていた.それでも なお,GA の受容部位に関する具体的な情報は乏しく,そ もそも GA が組織レベルにおいて空間的にどのように分布 しているかすら十分には分かっていない状況であった.そ こで,高濃度に存在することが知られている植物組織を対 象として,GA の組織内局在を明らかにすることを発端と する,GA の生理機能解析を目的に掲げて研究を開始した. 現在でこそ LC/MS の技術が発達し,それまで当然の如 く要求された多段階の精製操作が省略され,多種類の物質 を対象とする一斉の定量定性分析が可能な時代に到達して いるが,研究開始当時は植物ホルモンの定量分析と言えば GC/MSを用いる手法が主流であり,各々の標的物質にあ わせて多段階からなる分画操作を経ることが常道であっ た.他方,抗体を用いて行う免疫学的定量法(イムノアッ セイ)もその GC/MS 分析への接続作業として要求される 各精製画分の簡易チェック法として大変有用で,周囲から のニーズが存在した.そこで,イムノアッセイ用途に加え て,組織化学的な GA 検出にも応用することを視野に入れ, GAと特異的に結合する抗体の調製を行った(Nakajima et al. 1991a).各種の GA を特異的に認識するポリクローナ ルおよびモノクローナル抗体を調製し,検出限界 10 フェ ムトモル程度の GA を捉えることが可能となった.加えて, 抗体を樹脂と結合させた状態でカラムに詰め,GA の簡便 な精製法も確立した(Nakajima et al. 1991b). 酵素標識した抗体を用い,組織レベルでの GA 検出法を 詳細に検討した結果,最大の問題点が組織中のタンパク質 に GA を固定する過程で生じる「オリジナル位置からのド リフト」にあることを明らかにした.試料の凍結乾燥と気 体のカルボジイミド系固定剤を組みあわせた新たな固定方 式を導入して改善を図り,組織レベルでは十分な位置精度 を維持した免疫組織化学による GA 検出を初めて可能にし た.そして,イネ花粉や,次項で触れるアサガオ未熟種子 の中の GA 局在に関する解析が可能となった(Hasegawa et al. 1994 ; Hasegawa et al. 1995).
2. ジベレリンの生理機能の解明に向けて① ― ア サガオ未熟種子 ヒルガオ科やマメ科など一部の双子葉植物は,成熟過程 の種子中に他の器官・組織と比べて高濃度の GA を蓄積す るが,その役割は未解明であった.そこで,ヒルガオ科植 物のアサガオ(Ipomoea nil)を対象として,種子成熟過程
における GA の機能を明らかにすることを目的として,免 疫組織化学的な解析を開始した.当初は,圃場で開花させ た未熟種子を材料として用いたことから,半ば季節仕事で あったことと,得られた結果の再現性を確保することが非 常に困難であった.そこで,気象室内でアサガオの結実条 件を精査し,通年栽培による種子調製を可能にした後,得 られた種子の GA 内生量についてラジオイムノアッセイ法 で調べた.その結果,開花後 6 日目(6 DAA : days after anthesis)の段階では痕跡量しか活性型 GA が存在しない のに対して,その後急増し,12 DAA 付近で量的なピーク を迎え,以降減少することが判明した.そこで,先に確立 した凍結乾燥とカルボジイミド系試薬を組み合わせた免疫 組織化学的解析をアサガオの未熟種子に適用した結果,図 1に示すとおり 9 DAA 頃から珠皮(integument)に GA の 局在を検出した(Nakayama et al. 2002).珠皮には GA 以 外にデンプン粒も存在し,GA の局在に伴ってそれらが次 第に分解される様子が観察の結果,判明した.これにより, 穀類種子の発芽過程において見られる現象に似て,珠皮中 に存在するデンプン粒の分解に GA が関与する可能性を見 出した. そこで,成熟過程にあるアサガオ種子中で発現する α -アミラーゼ遺伝子 2 種(InAmy1 および 2)をクローニン グし,それらの発現状況を解析した.ノーザン解析の結果, 種子における発現は InAmy1 が支配的であった.種子内に おける InAmy1 の発現時期として,6 DAA でほとんど認め られず,9∼12 DAA で次第に増加し,15 DAA で定常域に 達した.そこで,発現量の漸増期にあたる 9 DAA の未熟 種子を用いて InAmy1 の発現に関する GA 応答性を調べた. その結果,0.1 µM の GA3を種子に投与した後,6 時間経 過した時点で顕著にその発現量の増加を認め,本遺伝子の GA応答能を確認した(Nakayama et al. 2002).大腸菌発現 系を用いて調製したリコンビナント InAmy1 を免疫原とし てポリクローナル抗体を得て,未熟種子に含まれる推定分 子質量 45 kDa の InAmy1 と特異的に反応性を示すことウェ スタン解析により明らかにした.その上でさらに,6∼18 DAAの種子切片に対する InAmy1 の免疫組織化学を行い, 12 DAA以降に明瞭となる GA 局在部位とほぼ重複する部 位に,InAmy1 も局在することを明らかにした(Nakayama et al. 2002 ; Nakajima et al.
2004).以上のことから,(i)In-Amy1遺伝子の発現に関する GA 応答性の付与を確認し, 加えて(ii)InAmy1 と GA が存在する時期および部位が重 複したことから,アサガオ未熟種子における GA の生理機 能として,α-アミラーゼ遺伝子の誘導を介したデンプンの 分解過程が含まれることを強く示唆する結果が得られた. ところで,GA や InAmy1 の局在を認めた珠皮は,処理 法として非常に穏やかに組織を固定した場合でも,構造的 には何も無いように見える空疎の領域があり,細胞の流失 が生じたと思われる切片画像を得ていた.このことは,珠 皮が本当に GA の生理機能の場であるか,換言すれば α -アミラーゼを生合成する場であるか疑問が生じた.採用し た組織の固定法は,操作中の GA のドリフト防止には有効 であるが,生合成されてその場で留まる GA 分子と,他か ら移動してその場に居た GA 分子とを区別できるものでは ない.従って,珠皮以外の場所から GA が移動してきた可 能性を検討するため,InAmy1 遺伝子と GA 生合成酵素遺 伝子の in situ ハイブリダイゼーションを計画した.アサガ オの GA 生合成酵素遺伝子については当時報告されたもの が存在しなかったことから,未熟種子由来の cDNA ライ ブラリーから計 4 種(InGA20ox1 および 2,InGA3ox1 およ び 2)をクローニングした.いずれの翻訳産物とも予想ど おりの酵素活性を示すことが GC/MS を用いて確認できた. そこで,本来の検討に戻り,in situ ハイブリダイゼーショ ンによる各遺伝子の空間的発現部位を特定した.その結果, 珠皮におけるデンプン粒が分解し始める時期において
In-GA20ox1と InGA3ox2,加えて InAmy1 のいずれもがほぼ重
複して発現する様子を確認したが,その部位は珠皮ではな く,それと隣接する種皮(seed coat)であった.これにより, 活性型 GA は本来種皮で生合成され,その場で InAmy1 の 生合成までを誘導するが,両分子とも隣接する珠皮へ移動 して蓄積するとともに,InAmy1 はその移動した場でデン プン分解機能を果たしていることが判明した(Nakajima et 図 1 アサガオ種子成熟過程におけるジベレリンの局在 (a)アサガオ未熟種子の断面.矢印で示す珠皮において GA およびデンプン粒の局在が認められた.bar = 1 mm.(b) -(f)免疫組織化学的解析による GA 局在部位.矢印で示す 茶色の染色した部位は GA の局在を意味する.写真は順に 開花後 6 日目(b),9 日目(c),12 日目(d),15 日目(e), 18日目(f).bar = 10 µm.(Nakayama et al. 2002)
図 2 アサガオ種子成熟過程における GA 関連遺伝子の発現
(a1)(a3)GA 生合成酵素遺伝子 InGA20ox1 用アンチセンスプローブを用いた in situ ハイブリダイゼーション.- (b1)(b3)
-α-アミラーゼ遺伝子 InAmy1 用アンチセンスプローブを用いた in situ ハイブリダイゼーション.上段から順に開花後
9日目,12 日目,15 日目.矢印で示す箇所から各遺伝子の発現が始まっている.(c)開花後 12 日目の果実縦断面を
用いて,InGA20ox1 用アンチセンスプローブを用いた in situ ハイブリダイゼーションを実施した様子.2 個の種子を 比較する限り,発現状況はほぼ同調的と言える(矢印).bar = 1 mm.(Nakayama et al. 2005)
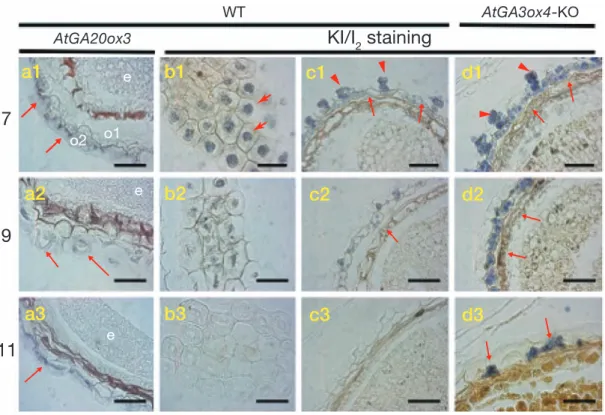
図 3 シロイヌナズナ種子成熟過程における GA の生理機能
(a1)(a3)野生型株由来の未熟種子を対象とする GA 生合成酵素遺伝子 AtGA20ox3 の in situ ハイブリダイゼーション.
-o1は外珠皮内層,o2 は外珠皮外層と呼ばれる.e は胚.未熟種子表層(b1)(b3)および断面(c1)- (c3)に関するヨ
-ウ素染色結果.矢尻は外珠皮外層に認められるカラム状構造の中のデンプン粒.矢印は外珠皮内層に認められるデン
プン粒.(d1)(d3)AtGA3ox4- -KO株由来の未熟種子断面のヨウ素染色結果.GA 生合成過程進行の支障はデンプンの
al. 2004 ; Nakayama et al. 2005).GA および InAmy1 のいず れも,種皮における局在状況を免疫組織化学的に検出でき なかったことについて,シンク器官として種皮から珠皮へ のフローがあり,その流れに乗じて生合成された GA も InAmy1も速やかに移動するためか,加えて,そもそも検 出感度を下回る量しか種皮に存在しないことなどが原因と 考えられる. なお,この解析から GA や α-アミラーゼの生合成は種 皮の全域で一斉に生じるのではなく,種子が胎座と接する 部分,すなわちシンク器官として唯一外部と接する部位か ら始まることが明らかとなり(図 2),外部から胎座を経 由して種子内に運ばれる何らかの制御物質の関与が示唆さ れる(Nakayama et al. 2005).アサガオの芽生えでは,活 性型 GA1の直前に位置する生合成前駆体 GA20が子葉から 茎頂へ移行する際の移動型分子形態であるとの知見が報告 されていたこともあり,上記制御物質が GA20である可能 性を検証したがこれを否定する結果を得ており,物質の正 体は明らかにはなっていない. 3. ジベレリンの生理機能の解明に向けて②―シロイ ヌナズナ未熟種子 種子成熟過程における GA の生理機能として,アサガオ 種子を用いた解析から判明した「α-アミラーゼの誘導によ りデンプンを分解して得られるエネルギー」は,おそらく その近傍に存在する子葉の発達あたりに使われるものと予 想した.それはつまり,穀類種子の発芽過程における GA の生理機能のアナロジー的発想とも言える.この予想され る GA の生理機能が双子葉植物に普遍的に認められるもの であるか検討するため,今度はシロイヌナズナを対象に据 えた. アサガオの解析例に倣い,未熟種子を用いた免疫組織化 学の実施を端緒として,組織レベルでの GA の局在部位の 把握を試みたが,どこにもシグナルを確認することができ なかった.そこで,モデル植物の利点を活かし,各関連遺 伝子をクローニングする手間もなく GA 生合成酵素遺伝子 の発現状況を調べ,発現するものについては時期・部位特 異性を把握して,最終的に in situ ハイブリダイゼーション による空間的な遺伝子の発現状況を調べた.その結果, AtGA20ox3 は未熟種子中の珠皮内において,また,AtGA-20ox2と AtGA3ox4 は珠皮と胚表層の 2 カ所で発現してい ることを明らかにした(Kim et al. 2005).このうち,珠皮 にはアサガオと同様にデンプン粒の存在と,GA 生合成遺 伝子の発現後のデンプン粒の分解・消失を認めたことから, シロイヌナズナにおいても再び GA のデンプン消化への関 与が示唆された(図 3).そこで,以降の対象を珠皮に据 えた.続いて,種子成熟過程で珠皮のデンプン粒分解に機 能する α-アミラーゼを探すため,ゲノムに存在する全 α -アミラーゼ遺伝子の発現状況を調べ,うち 3 種の未熟種子 中における発現を確認した.中でも AtAmy3 の発現レベル が未熟種子中で支配的であると判明したことから,AtA-my3の in situ ハイブリダイゼーションを実施した.その結 果,AtGA20ox2 や AtGA3ox4 と同様に珠皮と胚表層にその 発現を認めた.加えて,AtAmy3 の発現に GA 応答性が付 与されていることも確認できた.よってこれらは,シロイ ヌナズナにおいてもアサガオで確認した現象と同じく, GAが α-アミラーゼ遺伝子の発現誘導を介してデンプンの 分解を促すと考えて矛盾のない結果が得られた(Kim et al. 2005). より確定的な情報を得るために,アサガオでは実施の難 しい各種変異株を用いた検討を計画した.すなわち,At-GA3ox4はその発現が種子成熟過程に高い特異性が認めら れることから,T-DNA挿入ラインの中から遺伝子発現が 低下した機能欠失型変異株(AtGA3ox4-KO)を選抜した. AtGA3ox4-KO株の未熟種子を対象として,成熟に伴う珠 皮中のデンプン粒の分解状況を調べた結果,野生型株の種 子と比較して明瞭な過程進行の遅れが認められた(図 3). また,α-アミラーゼ遺伝子の発現状況も,AtAmy3 をはじ めとして全て発現が抑制された状態にあることを確認した (Kim et al. 2005). デンプン粒の存在部位であり,GA 生合成酵素遺伝子や α-アミラーゼ遺伝子の発現部位でもある珠皮は,完熟する につれて種皮と接合する.このことから,デンプン粒の分 解過程が遅れることは種皮表層にも何らかの影響を及ぼす と想定した.そこで,AtGA3ox4-KO株の完熟種子を対象 として,走査型電子顕微鏡による種子表面の観察を行った. その結果,野生型株で認められる六角模様の配列が AtGA-3ox4-KO株では大きく乱れていることが判明した(図 4). この傾向は,GA 生合成能の欠損変異株 ga1 を用いても同 内容が生ずることを確認した.すなわち,ga1 は GA に関 する内生量が低下しており,正常な発芽や稔実のためには 適度な GA 処理を要求する.そこで,投与する GA 量を調 節して中程度の GA 欠乏状態をつくり,その状況において 形成される種子を電子顕微鏡を用いて観察した.その結果, これら GA 欠乏状態の完熟種子もその表層模様の配列に乱 れが生じていた.加えて,十分量の GA を投与した ga1 変 異体の種子ではこの乱れが回復することも確認した(Kim et al. 2005).これらの結果を統合して,珠皮における GA は, デンプン粒の分解過程に関与して規則的な種子表面構造の 形成に影響を及ぼすとの結論を得た.この構造的な乱れは, 吸水時に保湿目的でほぼ均一に種子の外周部に膨潤する 「粘液質(mucilage)とよばれる多糖類」を不均一にする. そしてこれは,発芽の効率に影響を及ぼす.従って,互い の過程こそ異なるものの,本質的には単子葉植物の発芽過 程における GA の生理機能と同様に,双子葉植物であるシ ロイヌナズナに関しても未熟種子中の GA の生理機能が結
果的に発芽過程に影響を与えることを明らかにした. 4. ジベレリン受容体の候補探索と同定 GAからの信号を細胞内に伝え,GA 特有の遺伝子 発現の発端になるものとして動物・微生物の例にならい受 容体タンパク質が想定されてきた.今でこそこの分野にお ける遺伝学的な解析の有効性は衆目の一致するところだ が,研究を開始した時点ではまだ分子生物学的な手法など 「誰でもできる」などと言える状況になく,もっぱら生化 学的な手法を用いて約半世紀も前からその候補探索が試み られてきた.他の植物ホルモンに関する受容体候補の研究 例でも大半がそうであるように,長年に及ぶ追究にもかか わらず GA 受容体も有力な候補が特定されない状況が続い た.90 年代に入り,穀類種子の発芽過程で広く知られる GAの加水分解酵素誘導応答系を対象として,生化学的な 方法により「種子糊粉層において α-アミラーゼなどデン プン分解酵素の誘導に関わる GA 受容体が,少なくとも細 胞膜外側に存在する」ことを強く示唆する報告がなされ, 候補タンパク質の一次配列情報に注目が集まった(Hooley et al. 1991 ; Gilroy and Jones 1994).存在量の乏しさが原因 か,あるいは,膜結合型タンパク質としての操作上の難し さゆえか,最終的な単離報告は現在に至ってもまだない. 筆者らも,その注目情報がもたらされる頃には,調製し た GA 抗体を受容体に見立て,候補タンパク質を生化学的 に検出するための系を整備していた.幸いなことに,各種 のモノクローナルな GA 抗体は,活性型 GA に対する親和 性として,最も高い値を示すクローンと最も低い値を示す クローンとではおよそ 100 倍程度のひらきがあり,活性型 GAに対する受容体の親和性など全く手がかりの無い中 で,想定される親和性に応じた候補タンパク質検出法の整 備に活用することができた.また,世間的には「GA 受容 体は膜結合型で,細胞膜表層に存在する」とする見解を誰 も否定しない情勢の中で,GA 分子が示すある程度の脂溶 性を根拠として,細胞質にも GA 受容体が存在する可能性 について調べていく立場をとった.可溶性の候補タンパク 質として,マングビーンやダイコンなど育成が比較的簡単 な植物を中心に探索を進めた(Nakajima et al. 1993).アズ キからは結合に関する GA の構造要求性が各々の生理活性 とほぼ一致する有力な候補を見出し,その精製を展開した (Nakajima et al. 1997).最も精製を進めた画分中に含まれ るタンパク質群の N 末端および内部のアミノ酸配列情報 を明らかにし,これらタンパク質をコードする遺伝子のク ローニングまで行ったが,そのリコンビナントタンパク質 からは GA 結合活性を確認することができず,いずれが活 性本体であるか確認するには至っていない(Park et al. 2005).こうした状況の中,共同研究の形でイネ GA 受容 体遺伝子の特定に関わることになった(Ueguchi-Tanaka et al. 2005).それは,既知情報の延長線上になく(筆者から すれば想定どおりの)可溶性の分子形態で,動物など他生 物までを視野に入れてもこれまでに知られていない仕組み により GA の受容が行われることを意味するものであっ た. GA受容体の機能に異常をきたせばその信号がうまく伝 わらなくなるため,矮性の形質が現れたり,GA に対する 感受性が低下するとの見通しから,シロイヌナズナをはじ めとしていくつかの植物を用いて GA 受容体に関連する変 異体の捜索が行われてきた.そして,イネについても変異 体プールの中からこうした発想にもとづく探索が行われた 結果,GA に対して非感受性で,極矮性を示す変異体 gid1 が見出された.イネ gid1 変異体は成長しても 5 cm 程度で, 花をつけず,GA 投与による葉鞘の伸長応答が認められな い.これに加えて,種子発芽における加水分解酵素の分泌 誘導も認められない.これは,細胞膜外側にその存在が想 定されてきた GA 受容体と,細胞内に存在する GID1 が機 能的に一致するものでありながら空間的にも分子形態とし ても異なるものと考えられるため,両者の関係について過 去に報告された結果を含め検証が必要な状況となってい る.大腸菌発現系を用いて調製した GID1 タンパク質は活 性型 GA 選択的な結合能を有し(図 5),極矮性となる gid1変異体において認められたものと同じ変異を導入した 翻訳産物は,完全に GA 結合能を失っていた.さらに,酵
母 two-hybrid系を用いて,GA からの信号を負に制御する
因子 DELLA と GID1 との GA 依存的な結合が確認される に至り,GID1 が DELLA 因子の機能制御を担うことから これを GA 受容体と結論するに足る証拠が得られた(Uegu-chi-Tanaka et al. 2005).
5.受容体が担うジベレリンの信号伝達機能の制御 GID1のオルソログとして,シロイヌナズナでは 3 種存在することを明らかにした(Nakajima et al. 2006).こ の 3 種の生化学的な性状は互いに似ており,GA の構造要 求性も似通っていた.そこで,機能的に何らかの差異があ るかを明らかにするため,機能を欠失させた変異体を作り 調べた(Iuchi et al. 2007).その結果,3 種それぞれの単独 変異ではいずれも明瞭な形質が現れなかったことから,互 いの機能を相補しあうと考えられた.しかし,3 種のうち 2種の機能が同時に異常となった変異体では,3 とおりの 組合せのうち 1 種のみに矮化傾向が認められ,また,それ とは別の 1 種のみに雄しべの伸長不良と稔実率の低下が認 められた(図 6).そこで,各受容体遺伝子のプロモーター によりその発現が制御される GUS 融合遺伝子発現形質転 換体(pGID1::GID1-GUS)を作り,それらの解析を行っ たところ,異常形質の原因として,残り 1 種の正常な受容 体が量的に欠乏していることが原因となる場合と,それだ けではうまく説明できない言うなれば「質的原因」が存在 す る 場 合 の 2 通 り あ る こ と が 判 明 し た(Suzuki et al.
2009). GAの信号伝達は,GA の結合により受容体の性状が変 化し,新たに,信号伝達の抑制因子 DELLA に対する親和 性が生じることによって受容体-DELLA間で複合体を形成 することが端緒となる.複合体の形成は DELLA 因子の持 つ信号伝達抑制機能を弱めることとリンクする.これを起 点として,2 つのアプローチを開始して現在に至っている. すなわち 1 つは,酵母の評価系を用いて受容体-DELLA間 の親和性を比較しており,そこからは雄しべの伸長不良が 現れた上記二重変異体において唯一正常に機能するはずの GA受容体と,雄しべにおいて支配的に機能すると考えら れる DELLA 因子との間の親和性が総じて低い傾向にある ことを明らかにし,受容体・DELLA 因子間の低親和性を 本質とする受容体の機能的劣性が異常形質を生じさせる要 因と解釈できる結果を得ている(Suzuki et al. 2009).もう 1つは,これら GA の信号伝達過程を制御する化合物を探 索することである.分解して考えてみると,受容体が関わ る GA の信号伝達過程は 3 つに分けられる(図 7).すな わち,受容体が GA と結合する過程,GA と結合した受容 体が性状を変えて DELLA 因子とも結合する過程,DELLA 因子がプロテアソームの関与するタンパク質分解過程に接 続する過程である.これらの各過程を対象としてその制御 に利用できる低分子化合物を幾つか探り当てており,以後 の研究に役立てたい(Otani et al. 2010 ; Yoon et al. 2013). ここに述べた研究手法は,ともすれば常道と呼ばれるもの から少し外れているものもあるかもしれないが,どんなア プローチも非常に有益な情報をもたらし得ることを身を 持って体験できたことは研究者としての幸せを大いに感じ 図 5 分子スイッチ機能に適した受容体の構造要求性 高等植物における一般的な GA 生合成経路.各 GA の分子構造とその下段にイネ GA 受容体に対する親和性について GA4を基準として相対的に示す.生合成前駆体および不活性化代謝物のいずれも非常に低い親和性しか示さない.
(Ueguchi-Tanaka et al. 2005)
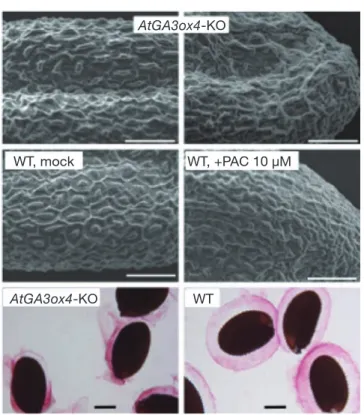
図 4 GA 欠損がシロイヌナズナ種子表面構造に与える影 響 (上段)AtGA3ox4-KO株由来の種子表面の電子顕微鏡写真. 独立した 2 ラインとも表面模様が乱れていた.bar = 50 µm. (中段)野生型株に終濃度 10 µM の GA 生合成酵素 阻害剤パクロブトラゾールを定期的に噴霧して調製した種 子表面の電子顕微鏡写真.対照区と比較して,表面模様に 乱れが認められる. (下段)AtGA3ox4-KO株由来の種子を 吸水処理後,ルテニウムレッド染色した様子.野生型株由 来の種子と比較して,粘液質の膨潤が不均一であった. bar = 200 µm.(Kim et al. 2005)
図 6 GA 受容体の機能欠損と表現形質
3種あるシロイヌナズナ GA 受容体の機能をすべて失った変異体(3KO)は極矮性で(a),GA 非感受性となる(b).
3種のうち 2 種のみ機能を失った変異体のうち,AtGID1a と AtGID1c の機能が同時に失われた変異体(ac)のみ矮性
となり(c),別の変異体(ab-2KO, AtGID1aと AtGID1b の機能が同時に失われた変異体)のみ雄しべが短いことが原
因で,稔実率が低下する(d).
図 7 GA 信号伝達過程における制御ポイント
受容体が関与する GA の信号伝達過程において,受容体と GA の結合に対する制御(X1),受容体と DELLA 因子の
結合に対する制御(X2),DELLA 因子が受容体から離れる際の制御(X3)が考えられる.これに加えて,受容体へ
ている.今後も,多勢に左右されず,よく考慮した上で自 分の信ずるところを進めていきたい. 謝辞 ここに述べたすべての研究を遂行するにあたり,積 極的に携わってくれた数多くの学部学生・大学院生の方々 や,そして,共同研究をご提案くださった研究者の方々, あるいは,こちらからの提案を快く受けてくださった研究 者の方々のご協力なくしては,いずれの成果も得られな かったと思っております.皆様方にこの場を借りて感謝申 し上げます. 文 献
Gilroy, S and Jones, RL (1994) Perception of gibberellin and abscisic acid at the external face of the plasma membrane of barley (Hordeum
vulgare L.) aleurone protoplasts. Plant Physiol 104 : 1185-1192.
Hasegawa, M, Hashimoto, N, Zhang, J, Nakajima, M, Takeda, K, Yama-guchi, I and Murofushi, N (1995) Immunohistochemistry of gib-berellin in rice anthers. Biosci Biotech Biochem 59 : 1925-1929.
Hasegawa, M, Nakajima, M, Takeda, K, Yamaguchi, I and Murofushi, N (1994) A novel gibberellin, 16α,17-d i h y d r o x y-1 6 , 1 7 -dihydrogibberellin A4-17-O-β-D-glucopyranoside, from rice anthers. Phytochemistry 37 : 629-634.
Hooley, R, Beale, MH and Smith, SJ (1991) Gibberellin perception at the plasma membrane of Avena fatua aleurone protoplasts. Planta 183 : 274-280.
Iuchi, S, Suzuki, H, Kim, Y-C, Iuchi, A, Kuromori, T, Ueguchi-Tanaka, M, Asami, T, Yamaguchi, I, Matsuoka, M, Kobayashi, M and Naka-jima, M (2007) Multiple loss-of-function of Arabidopsis gibberellin receptor AtGID1s completely shuts down a gibberellin signal. Plant J 50 : 958-966.
Kim, Y-C, Nakajima, M, Nakayama, A and Yamaguchi, I (2005) Con-tribution of gibberellins to the formation of Arabidopsis seed coat through starch degradation. Plant Cell Physiol 46 : 1317-1325.
Nakajima, M, Nakayama, A, Xu, Z-J and Yamaguchi, I (2004) Gib-berellin induces alpha-amylase gene in seed coat of Ipomoea nil imma-ture seeds. Biosci Biotech Biochem 68 : 631-637.
Nakajima, M, Shimada, A, Takashi, Y, Kim, Y-C, Park, S-H, Ueguchi -Tanaka, M, Suzuki, H, Katoh, E, Iuchi, S, Kobayashi, M, Maeda, T, Matsuoka, M and Yamaguchi, I (2006) Identification and charac-terization of Arabidopsis gibberellin receptors. Plant J 46 : 880-889.
Nakajima, M, Takita, K, Wada, H, Mihara, K, Hasegawa, M, Yamagu-chi, I and Murofushi, N (1997) Partial purification and character-ization of a gibberellin-binding protein from seedling of Azukia angularis. Biochem Biophys Res Comm 241 : 782-786.
Nakajima, M, Yamaguchi, I, Kizawa, S, Murofushi, N and Takahashi, N (1991a) Preparation and validation of an antiserum specific for non
-derivatized gibberellins. Plant Cell Physiol 32 : 505-510.
Nakajima, M, Yamaguchi, I, Nagatani, A, Kizawa, S, Murofushi, N, Fu-ruya, M and Takahashi, N (1991b) Monoclonal antibodies specific for non-derivatized gibberellins. I. Preparation of monoclonal antibod-i e s a g a antibod-i n s t G A4 a n d t h e i r u s e i n i m m u n o a f f i n i t y c o l u m n
chromatography. Plant Cell Physiol 32 : 515-521.
Nakayama, A, Nakajima, M and Yamaguchi, I (2005) Distribution of gibberellins and expressional analysis of GA 20-oxidase genes of morning glory during fruit maturation. Biosci Biotech Biochem 69 : 334-342.
Nakayama, A, Park, S-J, Xu, Z-J, Nakajima, M and Yamaguchi, I (2002) Immunohistochemistry of active gibberellins and gibberellin
-inducible α-amylase in developing seeds of Morning glory. Plant Physiol 129 : 1045-1053.
Otani, M, Yoon, J-M, Park, S-H, Asami, T and Nakajima, M (2010) Screening and characterization of an inhibitory chemical specific to Arabidopsis gibberellin 2-oxidases. Bioorg Med Chem Lett
20 : 4259-4262.
Park, S-H, Nakajima, M, Hasegawa, M and Yamaguchi, I (2005) Sim-ilarity and difference between the characteristics of gibberellin-binding protein and gibberellin 2-oxidases in Adzuki bean (Vigna angularis) seedlings. Biosci Biotechnol Biochem 69 : 1508-1514.
Suzuki, H, Park, S-H, Okubo, K, Kitamura, J, Ueguchi-Tanaka, M, Iu-chi, S, Katoh, E, Kobayashi, M, YamaguIu-chi, I, Matsuoka, M, Asa-mi, T and Nakajima, M (2009) Differential expression and affini-ties of Arabidopsis gibberellin receptors can explain variation in phenotypes of multiple knock-out mutants. Plant J 60 : 48-55.
Ueguchi-Tanaka, M, Ashikari, M, Nakajima, M, Itoh, H, Katoh, E, Ko-bayashi, M, Chow, T-Y, Hsing, Y-C, Kitano, H, Yamaguchi, I and Matsuoka, M (2005) GIBBERELLIN INSENSITIVE DWARF1 en-codes a soluble receptor for gibberellin. Nature 437 : 693-698.
Yoon, J-M, Nakajima, M, Mashiguchi, K, Park, S-H, Otani, M and Asami, T (2013) Chemical screening of an inhibitor for gibberellin receptors based on a yeast two-hybrid system. Bioorg Med Chem Lett
23 : 1096-1098. 連絡先 : 〒 113-8657 東京都文京区弥生 1-1-1 東京大学大学院農学生命科学研究科応用生命化学専 攻 中嶋 正敏 TEL : 03-5841-5192 FAX : 03-5841-8025