TheJapaneseJeurnalofPsychononli ‘Science 2013
,
VoL32
,
No
,
1,
95−
100講 演論 文
網月
莫
か
ら
の マ ルチ
ニ ュー
ロ ン記録
と
行 動実 験
に よ る
視 覚 系
の
解 明
石
金
浩 史
専 修 大 学Functional
analysis ofthe visual systembased
on multi−
neuronal retinal signalsand
behavioral
responsesHiroshi
IsHIKANE
Senshu University
Neuroscientists
have
studied the visual systemby
accumulatingknowledge
about the structural andfunctional
properties
of single neurons.
However,
recent studiesperforming
multi−
unit recordings have revealed that a signifi−
cantfraction
of neurons exhibit temporally correlated activit孟es.
It has been suggested that correlated activities among neurons may have afunctional
role in visual information processing,
though there is a possibility that such activities may be obtained as the byproduct.
The present article reviews recent advances in retinal neuroscience us−
ing multi
−
unit recordings,
mQlecular techne1ogies,
and pharmacological manipulations.
The analysis Qf visuallyguided
behaviors revea 工ed the functional signi 丘cance of neural codes suggested by electrophysiological data・
These approaches also enabled us to clarify the neural mechanisms ofthedetectien
ofimage motion,
whichis
fundamentalto Vision
.
Key words :retina
,
multi−
channe1 recording,
Visually guidedbehavior
は じ め に 英 国の生 理 学 者Adrian は
,
感 覚 強 度が感 覚ニ ュー
ロ ンに おける活動 電 位の生 起 頻 度 (ス パ イ ク頻 度 )と対 応 関係が ある こ と に基づ き,
感覚情 報がニ ュー
ロ ンの スパ イク頻 度に よっ て符 号 化 さ れて い る と考 え た (Adrian,
1947)。
以後,
研 究 者はニ ュー
ロ ン活 動の記 録 結 果にっ い て機 能 的意義を論ずる に あ た り,
ス パ イ ク頻 度を拠り 所とするこ と になっ た。 分子 生物 学レベ ル,
シナプス レ ベ ル,
局 所 神 経回路 網レ ベ ル か らシス テム神 経 科 学に い た る ま で,
神経科 学分野で は多か れ少なか れ ス パ イク頻 度と の関係性を意識して研究が行わ れ てい る。
ニ ュー
ロ ンか らスパ イクを 記録する研究者は,
複 数回の刺 激呈示 を行っ て記 録さ れ たス パイ ク列からラス ター
プロ ッ トを 作 成し,
そこ から一
定の時間幅 (time bin)で区切 ら れ たCorresponding address :Department of Psychelogy
,
School ofHuman Sciences
,
Senshu University,
2−
1−
1 Higashimita,
Tama
−ku,
Kawasaki,
Kanagawa 214−
8580,
Iapan.
E−
mail :ishikane@
psy・
senshu−
u・
ac・
jp
peri
−
stimulus timehistogram
(PSTH )を作 成し, 刺 激に依 存し た頻 度の変化や時間経 過を解 析す る。
こ の よ う に,
ス パ イ ク頻 度はニ ュー
ロン に よる最 も基 本的な情報 符号 化の様 式で ある が,
近 年の多 細 胞 同 時 記 録 法の進 歩によ り,
スパ イ ク頻 度 以 外の方 略でも情 報が符 号 化される可 能 性が 示 さ れ る よ うになっ た。
視 覚 系にお け る相 関 発 火 とその機 能 視覚系に お け る相関 発 火の研 究は, 同 時 記 録さ れ た網 膜神経 節細胞の 2 細胞間に お け る相互相関ヒ ス トグ ラム の解 析に は じまる。 ネコ お よ びキ ンギ ョ網 膜を用い た研 究に おい て,
受 容 野の特 性が同一
タ イプの ニ ュー
ロ ン問 で は同期 発火が観察さ れ る が,
異な る タ イプのニ ュー
ロ ン間では同 期 発 火が観察さ れず,一
方のニ ュー
ロ ン に お い てスパ イ クが生起 すると他 方のニ ュー
ロ ンのス パ イク 頻 度が低下 す る場 合が あ るこ と が報 告さ れた (Rodieck,
1967;Arnett,
1978;Mastronarde,
1983)。 これ らの研究は自 発 発 火を対象に して お り,
相互相関ヒ ス トグラ ム か ら そ の原 因とな る共 通 入 力の特 性やその神 経 基 盤につ い て推96 基礎心 理学 研 究 第32巻 第1咢 定が なさ れ た
。
た だ し,
当 時 報 告さ れ たの は受 容 野が互 い に重な る よ う な近接し たニ ュー
ロ ン 間の相関発火で あっ た。 そ し て,
受容野が 彑い に重ならない ような遠 く 離れ たニ ュー
ロ ン間で は, 発 火に相 関がない と考え られ た。
そ の後, ウ サ ギ や タ イ ガー
サ ラマ ンダー
を用い た研 究が な さ れ,
同様の結果が報 告さ れ て い る (Arnett・&Spraker
,
1981;Meister,
Lagnado,
&Baylor
,
1995;Brivanlou,
Warland
,
& Meister,
1998)。 受 容 野が近 接 してい るニ ュー
ロ ン 間の 同期 発 火の機 能 的 意 義につ い て は,
Meisterら が タ イ ガー
サ ラマ ンダー
の網 膜神経 節 細 胞 群か ら同 時 記 録ざ れ たス パ イ ク を解 析 し,
興 味 深い 結 果を 示 した (Meister et a工.
,
1995) 。 まず,
多数の ランダム ピ クセ ル を 継 時的に呈 示 し,
ニ ュー
ロ ン のスパ イク応 答か ら各 受 容 野 を 逆 相 関法 (Jones
& Palmer,
1987)を用いて算 出した。 彼ら は, 受 容野 が部分 的に重なっ てい る2つ のニ ュー
ロ ンを選 び,
そ れ らの同期 発火の みを抜き出し て同 期ス パ イ ク列を作 成し た。 こ の同 期ス パ イク列か ら再 び 逆 相 関 法に より受容野 を 算 出 したと ころ,
元 々の2つ の ニ ュー
ロ ンの受 容 野が重なっ た部 位が受 容野 と し て浮か び上 がっ た。 し た がっ て,
網膜の次の段階に おい て同期 発 火 を復号化で き れ ば,
同 期 発 火に よっ て 2っ の神 経 節 細 胞 の受 容 野の重なっ た領 域の光情 報を伝 達で き ること に な る。
網 膜に投 影され た像は,
限ら れ た数の神経節細胞の 軸 索を介 して脳に送 ら れて い る。 各 神 経 節細 胞が その受 容野 内の視 覚 情 報 をスパ イク頻 度の み によ り互い に独 立 して符 号化 して い るな ら,
神 経 節 細 胞の数が画 素 数とな り,
脳に送 ら れ る視覚像の空間分解 能が そ れ に依 存す る。
し か し,
Meisterらの結果は,
神経 節 細 胞 群が同 期 発 火と い う も う1つ の符 号 化の チャネル を加 えるこ と で,
よ り空 間 的 解 像 度の高い情報を脳に送っ てい ること を示 唆して お り,
大 変に注 日を浴び た。 だ が,
実際に同 期発 火 が 網 膜の次の段 階に お い て復 号 化されて い る か ど うかは示さ れて お らず,
同期 発 火の機 能 的 意 義は証 明さ れ なか っ た。
ま た,
情 報理論 (infermation theory)をマ ウス網膜 神経節細胞の ス パ イク列に対 して適 用 した 研 究 で は,
ス パイ ク頻 度に よ る情 報 符 号 化に加 えて相 関 発 火 を符 号 化に用い て も,
情報量がほ と ん ど増え ない こと か ら,
相 関発 火が視 覚 情 報を符 号化 して いると は考えに くいと報 告さ れ た (Nirenberg
,
Carcieri,
Jacobs
,
& Latham,
2001)。
ニ ュ
ー
ロ ン活 動に対 して情 報 理 論 を適 用 し た研 究で は,その段 階に お け る情 報 量の み を問題に す る。 しか し
,
機能 的 意 義を実証 す る に は
,
次の段階における復 号 化に関す る知 見が必 要で ある が
,
その詳 細は不 明な た め に結 論が 出に くい
。
例 えば,
次の段 階の すべ て のニ ュー
ロ ン がcoincidence detector(
Jeffress
,
1948;Kuba
,
2007)として の性 質を持っ てい ると,
収 斂 結 合す るニ ュー
ロ ン群が同期 発 火し ない 限り信 号が伝わ ら ない こと と な る。 し た がっ て,
情 報 理 論に より情 報 量が算 出さ れて も,
次の段 階で は有 効で は ない 場合が 出て く る。 な お,
ネコの外側膝 状 体に おい て も相関発火が観 察さ れ るこ とが知ら れており (Alonso,
Usrey,
& Reid,
1996),
情 報 理 論の適 用に より同期 発 火が視 覚 情 報を符 号 化 してい る こ と が 示唆さ れ てい
る (Dan
,
Alonso,
Usrey,
&Reid,
1998)。 情報理論の適 用はその段階にお け る神 経 表 現につ いて有 用な知 見を与えて くれ る が
,
受け手 側にお け る復 号 化に関 する特 性が不 明 な場 合に は解 釈に慎 重を要す る。
視覚皮質
にお ける離
れ た 細胞
間の相関
発 火 視 覚皮質に おい て も,
Vl野 を中心に多 細 胞 同時 記 録 法 が適用さ れ る ようにな り,
相 関 発 火が調べ られるよ うに な っ た。
Wiese1 らの研 究グルー
プは,
Vl 野のニ ュー
ロ ン 間に同 期 発 火が存 在す ること を発 見し た (Ts’
o,
Gilbert,
& Wiesel,
1986)。
彼 ら はネコ Vl 野の213層の異な る2つ の 部 位か ら2つ のニ ュー
ロ ン の スパ イ ク を同 時 記 録し た。 記 録 され たスパイ クにつ い て相 互 相 関ヒス トグラムを算 出し た結果,
方 位選択 性や 眼 優位 性とい っ たニ ュー
ロ ン の受容野特性が類 似してい ると,
二 y ロ ンが同 期 発 火 等の相関 発 火を示 すこ とが明 らか になっ た。
また,
類 似 の波 長に選 択 性を持つ 異な るプロ ッ プ (blob
)に位置す るニ ュー
ロ ン間や,
類似の方位選択性を持っ プロ ッ プ問 (interbl。b)に位置す るニ ュー
ロ ン間で は相関 発 火が見 ら れ る が,
プロ ップニ ュー
ロン とプロ ップ 間ニ ュー
ロ ン の 間で は相 関 発 火が 見 られ ない こ と もわか っ た (Ts’
o & Gilbert,
1988)。
これ らの 相関 発 火は,
皮 質 内の数 ミ リメ
ー
トル にわ たる水平 結 合 (Fisken,
Garey
,
&Powell
,
1975)を介して引 き起こ され ると推 定されてい る(Tsb et aL
,
1986; Ts’
o & Gilbert ,19SS)。
そのた め,
視野 上に おける受容野 の位 置が 互い に全く重な ら ないニ ュー
ロ ン間にも相 関 発 火が見 られ,
こ の水 平 結 合に よ る グロー
バ ル なネッ ト ワー
クを介 する こ と で.
各ニ ュー
ロ ンは そ の古 典的受 容 野よりも は る かに広い領域の視覚情報を統合 する と推 定 さ れて い る (Gilbert
,
1992
)。
さ らに, 複 数の ド イ ツ の研 究グルー
プは,
ネコの視 覚 皮 質に おい て ガンマ 帯域の周 期 性を持つ同期発火を数ミリメー
トル 離 れ たニ ュー
ロン問に観 察 し た (Edkhorn et al
、
,
1988;Gray,
K6nig,
Engel,
&Singer
,
1989;Engel,
K6nig,
Gray,
& Singer,
1990)。 こ の周期的 同 期 発 火が共通運命の要因や連 続性等に依 存 してい る
こと か ら
,
ゲシ ュ タル ト的 知 覚 統 合に関 与す る神経基 盤で あると考え ら れ た (Singer & Gray
,
1995)。 しか し なが石 金:網 膜か らのマ ル チニ ュ
ー
ロ ン記 録と行 動 実 験に よ る視覚系の解 明 97 れ た に過ぎず,
周期 的同期発 火が 「神経 活動の結果生じ た単な る副 産 物に過ぎ ない」と い う可 能 性 を排 除 するこ とはで きなか っ た た め,
そ の機 能 的 意義につ い て は結論 が出なか っ た。
カエ ル網膜
の周期的
同 期 発 火 皮質の発 達し ていない下等 脊椎動 物で は,
ほ乳類にお いて皮 質で実 現さ れて い る機 能が前 倒 しで末 梢 側に実 装 さ れ て い る こ と が 多い。
例えば,
カエ ル 網 膜では,
神 経 節 細 胞に大き く分け て4種 類のサブタ イプが あ り,
虫検 出 器などの よ うに抽 象 化さ れ た情報を脳に送っ て い る ことが知 られて い る (Lettvin
,
Maturana,
McCulloch,&Pitts, 1959)
。
Ishikaneらは,
ウ シ ガエ ルの剥 離網膜に多 点電極を 適用し
,
光 刺 激に対す る神 経 節 細 胞の スパ イク応 答を記 録 した (lshikane
,
Kawam,
& Tachibana,
1999)。
彼ら は , 神 経 節 細 胞のサブタ イプで ある ディ ミン グ検 出 器 群が,
光刺激に依 存して周期 的に同期発火す るこ とを見いだ し た。 こ の周期 的同期発火は,
細胞体が網 膜 上で 2mm 以 上離れ たニ ュー
ロ ン間で も観 察され,
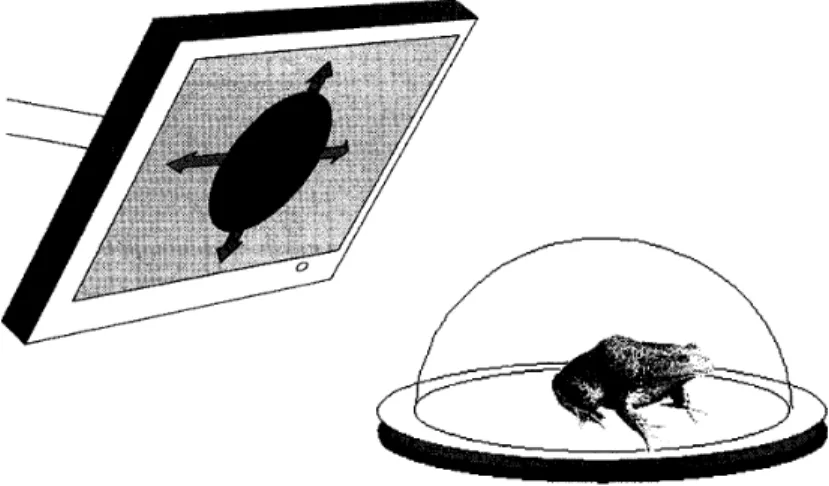
視 覚 皮 質で報 告さ れ た 周期 的同期 発 火と相同 な特 性を持つ こと が わ かっ た。 周 期 的 同 期 発 火による視 覚 情 報 符 号 化の実 証 カエ ル は大き な物 体が接 近す る と逃 げる。 ウシガエ ル を透 明なア ク リル ドー
ム の 中に座らせ,
PC に接 続さ れ たモニ タか ら拡 大 する黒い スポッ トを 旱示 すると,
刺 激 が ある程 度の 大 き さに なると逃 避 行 動が 誘 発される (Figure 1)。
こ の逃 避 行 動を誘 発す る刺 激を剥 離 網 膜に 呈示す る と,
ディ ミ ング検 出器群に周期 的同期発火が観 察さ れる。
そこ で,
Ishikaneらは周 期 的 同 期 発 火の機 能 的 意 義 を調べ る た め , 薬 理 学 的に周 期 的 同 期 発 火の強 度 を操 作し、
逃 避行 動に与え る影 響を調べ た (lshikane,
Gangi
,
Honda,
& Tachibana,
2005)。 周 期 的 同 期 発 火 はGABAA 受容体の阻 害 剤で あ る
bicuculline
の投 与に よ り抑 制さ れ (lshikane et al.
,
1999),
GABAc 受容体の阻害剤で あ る (1,
2,
5,
6−
tetrahydropyridin−
4−
yl)methylphosphinic add(TPMPA )の投 与に より促 進さ れる (Arai, Yamada , Asaka
,
&Tachibana ,2004;Ishikane et al
.
,20Q5)。
そこで, カエ ル の 眼 球 内にbicucullineを投与し たと ころ,
拡大する黒い ス ポッ ト刺 激 を呈 示 して も逃 避 行 動が 誘 発され なか っ た。
ま た,
同 様にTPMPA を 投 与し た ところ,
注 入 前 まで は 逃 避 行 動が誘 発さ れ なか っ た 小 さ な刺 激を 呈 示 して も,
逃 避 行動が誘 発さ れ る ように なっ た。 すな わち,
周 期 的 同期発 火を抑制 する と逃 避 行 動の誘 発が抑 制され,
周 期 的 同 期 発 火を促 進す る と逃 避 行 動の誘 発が促進さ れ た。 これらの薬物は双方ともGABA 受容体の阻害剤で あ るた め, 投 与 後は各ニ ュー
ロ ンのスパ イ ク頻度が増える傾 向 に あ る が, ディ ミン グ検 出器 や他のニ ュー
ロ ンのス パ イ ク頻 度で は逃 避 行 動の変化を説明で き なか っ た。 し た がっ て,
周 期 的 同 期 発 火がカエ ル の視 覚 誘 発 性 逃 避 行 動 に関連 する情 報 を符 号 化 し,
脳に伝 達 してい る こと が明 らか になっ た。
皮 質で は,
新し く発 見さ れ た情報 符号化様式が,
実 際 に視覚情報処 理に使わ れ て い るか ど うかを証 明す るこ と は容 易で はない。
神 経回路 網の複 雑さ や解 剖 学 的 構 造ゆ え に適 用 可能な実験手法が限られて い る た めで ある。
し か し,
比 較 的ア ク セ ス しやすい下 等 脊 椎 動 物の初 期 視 覚 系か ら= ユー
ロ ン活 動 を記 録し,
ニ ュー
ロ ン活 動の操 作 や行 動 実 験 を組み合 わせ るこ とで,
情 報 符 号 化 様 式の実 証が可能な場合も あ る。 運 動残
効 と運 動方
向 選 択 性ニ ュー
ロ ン 滝の錯 視と して太 古より知ら れ てい る運動残効は,
運 動 情報処理の メ カニ ズム を解明 する上で人変重要な手が か りで ある と考え られ,
盛ん に研 究 対 象と さ れ て きた98 基 礎 心 理 学 研究 第32巻 第1号
(Anstis
,
Verstraten,
&Mather,
1998)。 視覚 系には運 動検出器となるニ ュ
ー
ロ ンが存 在し,
受 容 野が特 定 方 向の運 動刺 激に より走 査さ れ た場 合に高い頻 度でスパイ クを発 生
し
,
その逆方向の運動 刺 激の場合には ほとん どスパ イ クを発生 し ない。 ニ ュ
ー
ロ ン の ス パイ クの頻 度が最も高くな る刺 激の運動 方向 をpreferred directionと呼 び
,
最 も低下する運動 方 向をnull directionと呼ぶ。 Barl。w らは
,
ウサ ギ網 膜の運動検 出器で あ る運動方向選択性 神経節細胞
(direction
−
selectiveganglion
cell; DSGC )を発 見した (Barlow &Hill
,
1963a)。 さ ら に,
DSGC に おい て長 時 間の運 動 刺 激 呈示後に観察さ れる スパ イク頻度の変化に基づい て,
運 動 方 向 検 出器 群の出 力バ ラン ス の変 化に よっ て運 動 残 効 が説 明で きると考 え た (Barlow &Hil1
,
1963b)。
しか しな が ら,
こ の よ う な 運動 方 向選択 性ニ ュー
ロ ンは,
視 覚 系 の複数の段階において存 在してお り,
ヒ トや サル の視覚 的意識 上の運 動 検 出と高い相 関 を持つ の はMT 野 等の皮 質 領 野の活動で あ ることが知られ ている(Stm MurasugLBritten
,
& Newsome,
王992;Zeki,
Watson,
&Frackowiak,
1993;Born & Bradley
,
2005)。
た だ し,
視 覚 的 意 識 上の運 動 検出とMT 野 のニ ュ
ー
ロ ン活 動が対 応 し ない場 合も あ る上に (Maj aj
,
Carandini,
&Movshon,
20e7; Hedges et al.
,
2011),
運動情報を処理 する皮 質 領 野は数 多く存 在するた め
,
運 動 情 報 処 理の神 経 基 盤 や 視 覚 的 意 識と の関 連につ い て全 貌を解 明す るこ とは容 易では ない。
ま た,
皮 質ニ ュー
ロ ンに おい て は,
運動 方 向選択性を形成す る局所神経回路 網の解析も進んで いない。
網 膜の運 動 方 向 選 択 性 神 経 節 細 胞と視 機 性反応 Barlowらに よるDSGC の発 見 後,
ウサギの DSGC の活 動は視 機 性の反 応の制 御に関 与 する こ とが示 唆さ れ た(Oyster
,
Takahashi,
&Collewijn,
1972)。
さ ら に,
マ ウス に分子生 物 学 的 手 法を適用 す るこ と で DSGC の機 能が詳
細に明ら か に さ れ た。 分子 生物 学的 手法の
一
種で ある
immunotoxin
−
mediated cell targeting(IMCT )法 をマウス に適 用し
,
網 膜 内の特定の アマ ク リン細 胞 を 選択的 に 死滅 させ ると,
DSGC の運 動 方 向 選 択 性が 消 失 する と と も に,
視 機 性の眼球運動が消 失した (Yoshida et a1.
,
2001)。
これ らの結果に よ り,
運動 方 向選択 性の形成に介 在 ニ ュー
m ンが必 要で あるこ とが証明さ れ る と と も に,
DSGC の 活 動が視 機 性の眼 球 運 動に 関 与 する こ と が証 明さ れ た。 ま た,
現在で はpreferreddirection
選 択 的 に DSGC に分 子マー
カー
を発 現さ せ る こ と が 口∫能になっ て い る(Yenehara et al.
,2009)。
こ の よ う に, 網 膜のDSGC は 視 機性反応に おける機能や,
方 向選 択性の形 成に関す る 知 見が蓄積 して い る。 し た が っ て,
運 動 情 報 処理 や 運動 残 効の神 経 基 盤 を解 明 する た めの アプロー
チの 1つ とし て,
運動 刺 激の呈 示 に対す る視 機 性の反 応と網 膜の DSGC の活動特性を調べ るこ とが挙 げら れ る。 ゼ ブラ フィ ッ シュ の視 運 動 反 応と 運 動 刺 激に対 する順 応 代 表 的な実 験 動 物であるゼ ブ ラフ ィッシ ュ は,
マ ウス と並ん で分子生 物 学 的 手 法の適用 が盛ん に行わ れ てい る。 ま た,
網 膜に始まる視覚系が非常に早い成熟を 示 す と ともに,
その機 能を調べ る た めの視 覚 誘 発 性の反 応が 豊富で ある ため,
視 覚 研 究に よ く用い られる。 特に,
視運 動 反 応 (Qptomotor response ;OMR )は
,
明 瞭に観 察され る視覚誘発性の反応であ り
,
運動刺激を追 うように泳 ぐ行 動と して観察するこ とがで きる。
lshikaneらは,
ゼ ブラフ ィ ッ シュ の長 時 間の運 動 刺 激の呈 示に対す る OMR と網 膜 神経 節 細 胞の応答 特 性を調べ た (lshikane,
2011)。
ゼ ブ ラフ ィッ シュ をドー
ナツ型の水 槽に入れ,
4 つ の液 晶モニ タを用いて水 平 方 向に運 動 する矩 形 波 縞 を 呈 示し,
ビデ オ カメ ラ でOMR を記 録し た (Figure 2)。
刺 激の時間 周波 数お よ び空間 周波 数を操作 し,
OMR の刺 激 依 存 性 を 調べ たと ころ,
先 行 研 究 (Maaswinkel& Li,
石 金 :網 膜か らのマ ル チニ
ュー
ロ ン記 録 と行 動 実 験 に よ る視 覚 系の解 明 99 2003)と一
致 した。
さらに,
最 も長 時 間OMR を誘 発 す る時 間 周 波 数お よび空 間 周 波 数を選 び, 運 動す る矩 形 波 縞を120秒間 呈 示 し た ところ,
刺激呈 示終了後に そ れ ま で呈 示さ れ て い た刺 激の運 動 方 向と は反 対 方 向に反 転 し て泳ぐ ゼブラフィ ッ シュ の行 動が観 察さ れ た。
この行 動 が運動 刺激呈 示終了後か ら5秒以内に生起し た確 率は,
より短い 60秒 間の刺 激 呈 示 終 了 後より有 意に高か っ た。
運動残効の強度は,
順 応 時 間が長 くな ればな る ほど増 大 す る た め,
こ の反 転 行 動 が 運 動 残 効 に 由 来 す る可 能性が 示 唆さ れ た。 た だ し,
観 察さ れ た反 転 行 動が運 動 残 効に 由来す ると結 論す るた めに は,
こ の行 動が他の刺 激 属 性 に対す る順 応に由来す る可能性や,
順 応 刺 激と は 反対 方 向の運動刺 激に対す る感度の上昇 な どを確認 する必要が ある。 ゼ ブ ラフィ ッシュ 網膜
の運動方向
選択性
神 経 節 細 胞 と運 動 刺 激に対 す る 順 応 特性
ゼブラ フ ィ ッシ ュ の視 機 性 反 応の 1つ で あるQMR に 運 動 残 効 様の反 転 行動が観 察さ れ た た め,
網 膜にお け るDSGC
の存 在 を調べ た (lshikane、
2011)。 剥 離 したゼブ ラ フ ィッ シュ 網 膜 に多 点電極を適用 し,
運動 刺 激の 呈 示 に 対す る神経節細 胞の スパ イ ク応 答を 記録し た と ころ, ゼ ブラフ ィ ッシュ 網膜に もDSGC が存在す ること が明ら か になっ た。
さらに,
OMR の誘 発で用いた運 動する矩 形 波縞 刺 激を様々 な運 動 方 向に おいて60秒 間 または120秒 間呈示し た。 その結 果,
null directionの運 動 刺 激を呈 示 し た場 合,
120秒 間の 旱示が終 了 した後か ら5秒 以 内の ス パイ ク数は,
他の方 向の運 動 刺 激 を呈 示し た後と比べ て増 大し た。 ま た,
運動刺 激を60秒間 呈 示 し た後の ス パ イク数は,
呈 示 さ れ た運動刺 激の方 向が どの方 向で あっ て もほぼ 同一
で あっ た。
DSGC は,
ス パ イ ク の頻度 が上昇 す るこ と で,
そのpreferred directionの運 動が受 容 野内に存 在す る こ と を脳に伝 える と考 え られ て い る (Barlow&Hill,
1963b)。 言い 換え れば,
脳 に とっ て その ニ ュー
ロ ン の活 動は特 定 方 向の運 動情 報の存在 を意 味す る。 ゼブラフ ィッシ ュ 網 膜の DSGC は,
nulldirectien
の 運 動 刺 激に対 して長 時 間順 応 した後にス パ イ ク頻度が 上 昇 する ことで,preferred
directionの運 動 情 報の存 在 を 脳 に送るこ とが推 定さ れ る。
した が っ て,
OMR に おい て 長 時 間の運動 刺 激の 呈示 後に観 察さ れ る,
あた かも順 応 刺激の運動方 向とは反対 方向の運 動 刺 激が 呈示さ れ たか の ような運 動 残 効 様の行 動は,
ゼ ブラ フィ ッ シュ
網 膜の DSGC の活 動に より説 明 可 能であるこ と が わか っ た。 ま と め 本 論文で は,
皮質で実証が難し かっ た周期的同期発火 の機 能 的 意 義につ い て, カエ ル網 膜のニ ュー
ロ ン の活 動 を薬 理 学 的に操 作し, 行 動と の対 応 関係を調べ ること で 明らか に し た研究と,
運動残効様の行動が ゼ ブラ フィ ッ シュ の視 覚 誘 発 性の行 動で観 察さ れ,
網 膜のDSGC の活 動に よっ て説 明 可 能で あることを 示し た研 究 を 紹 介 し た。
あ る動 物 種に おいて 証 明 さ れ たことが他の動 物 種に おい て当ては ま るとは限ら ない し,
高次機能は高等ほ乳 類を用い ない と解 明で き ない 場 合も多い。
し か し なが ら,
ニ ュー
ロ ン のスパ イ クパ ター
ンに よ る情 報 符 号 化と い っ た一
般 原理に か か わ る問題を解 明す る にあた り,
実 際に動 物の中 枢 神 経 系に お い てその符 号 化 様 式が情 報を 伝え てい る か どう か を確 認す る こ と の意 義は大きい。
そ の よ う な研究で は,
下等脊椎 動 物の比 較 的 単 純な視 覚 誘 発 性 行 動の特 性 を調べ た上で,
実 験 的 操 作の容 易な網 膜 のニ ュー
ロ ン活 動との対 応 関 係 を 検 討 するこ と が大 変 有 用であ ると考え られ る。
謝 辞 本 研 究の一
部は,科 学 研 究 費 補 助 金 (課 題 番 号22730586,
研 究 代 表 者 石 金 浩 史 )お よび 平 成23〜
27年 度 文 部 科 学 省 私立大 学 戦 略 的 研究基盤形成事業 (SllO1013)の補 助 を 受けた。引
用文 献
Adr正an
,
E,
D,
(1947).
The physi⊂al background ofpercept 孟on.
,
Oxf{)rd:Clarendon Press
.
Alonso
,
J.
M.
,
Usrey,
W M.
,
& Reid,
R C.
(1996).
Preciselycorrelated
firing
in ce且ls
of thelateral
geniculate nudeus.
Nature
,
383,
815−
819,
Anstis
,
S.
,
Verstraten F A.
,
& Mather G.
(1998).
The motionaftereffe⊂t
・
]「
Yends in Co9π討 れ丿
e Scien‘es,
2,
111−
117・
Arai
,
1.
,
Yamada,
Y,
Asaka,
T.
,
&Tachibana,
M.
(2004).
Light−
evoked Qs¢illatory
discharges
in retinalganglion
cells are
generated
by rh }塗hmic synaptic inputs.
Journat
(ゾNeuro−
phy
∫iology,
92,
715−
725.
Arnett
,
D、
W (1978).
Statistical dependence between neigh−
boring retinal ganglion cells in goldfish
.
Experimental BrainResea厂‘乃
,
32,
49−
53.
Amett
,
D,
,
&Spraker T.
E,
(1981),
Cross−
correlation analysisof th巳maintained discharge Qf rabbit retinal ganglion cells
,
Journal
〔,fPhys
ゴolog γ,
317,
29−
47.
Barlow, H
.
B.
,&Hill
,R,
M .
(1963a).
Selective
sensitivity to di−
rection of movement 貢n ganglion cells of the rabbit retina.
∫cience
,
139,
412−
414.
Barlow
,
H.
B.
,
&H丑1,
R,
M.
(1963b).
Evidence for a physiolog−
100
gmp,L)i-\lillfti
1347.Born
R.Tl,
&
BradleyD.C.
(2005),
Structure
andfunction
ofvisual area MZ Annual Review
of
Neuroscience, 28,189.Brivanlou,
I.H.,wrrland,D.K.,& Meister,M.
(1998).
anisms ef cencerted firingamong retinal ganglioncells,Nburon,20,527-S39.
Dan,g Alonso, I,M,,UsreM ML M., & Reid R.C.
(1998).
ing ofvisualinformationby
Precisely
correlated spikes inthelateral
geniculate
nucleus. Mture NIiuroscience,1, 507.Eckhorn,R.,Bauer, R.,
Jerdan,
NNC,Brosch, M,,Kruse,W,Munk,M.,&Reitboeck,H.
J.
(1988).
Coherent oscillations:A mechanism of featurelinkinginthevisual cortex?
pleelectrode and correlation analyses in thecat.Biolagical
dybernetics,
60,121-130,Engel,A.K.,K6nig,R,Gra" C.M,,& Singer,SM
(1990).
ulus-dependent neuronal oscillations jncat visual cortex:inter-columnarinteractionas determinedby
tionanalysis,
European
Jburnal
ofNeuroscience,
2,S88-606,Fisken,R. A..GareF L.
J.,
& Powell, [1RS.(1975).
Thesic,association and cemmissural connections ofarea 17 of
the
visual certex, Philosophical7ittnsactionsof
the RcyalSocietyB,272,487-536,
Gilbert,C.D.
(1992),
Horizental integrationand cortical narnics. Nburon, 9,1-13.Gray C,M., K6nig,R,Engel,A.K.,& Singer,W
(1989).
Oscillatoryresponses incat visual cortex exhibitcolumnar synchronization which reflects globalstimulus
properties.Nbture, 338,334-337.
Hedges,
J.
H.,Gartshteyn,YZ,Kohn,A.,Rust,N.C.,Shadlen,M.N,,Newsome, W T.,& Movshon,
J.
A.(2011),
tionof neuronal and psychophysicalrespenses tolocalandglobalmotion. CurrentBioiogy,21,2023-2028.
Ishikane,H.
(2011),
The metion after-effect inthe optomotorrespense ofzebrafish.
Jburnal
of
Vision,11,761-761.Ishikane,
H.,
Gangi,
M.,Honda,S,,& lhchibana,M,(2005).
Synchronizedretinal oscillations encode essentialtion forescape behayiorin fregs.IVbtureNeuroscience,8,
1087-1095.
Ishikane,H.,Kawana,A,,& Thchibana,M.
(1999).
Short-andlong-rangesynchronous activities indimming detectorsof
thefrogretina. Vl'sualNeuroscience,16,IOO1-1014.
Jeffress,
L.A.(1948).
Aplace
theory of soundlocalization,
Jburnal
of
Compatutive& Plysiolqgi'calPsp,chology,41,39.Jones,
J,
R,& Palmer,L.A.(1987).
Thetwo-dimensionalspa-tialstructure ofsimple receptive fieldsin catstriate cortex.
journalofNleuropPrysiology,58,1187-1211,
Kuba,H.
(2007).
Cellular and molecular mechanisms of avian auditery coincidence detection.NeuroscienceResearch,59,ag32ts
eelg
370-376.
Lettvin,
J.
Y,Maturana,H.R.,McCulloch,"LS.,
& Pitts,W H.(1959),
What thefreg's
eye tellsthefreg's
brain.
ingsof
thelnstituteqfRadioEngineers,49,1940-I95I.MaaswinkeL H,,& Li,L,
(2003).
SPatio-temporalfrequency characteristics of the optomotor response in zebrafish.VisionResearch,43,21-30.
Majaj,N.
J,
Carandini,M.,& Movshen,J,
A.(2007),
Motion integrationbyneurons inmacaque MT islocaLnot globaL
Journal
ofNeuroscience,
27,366-370,
Mastronarde,D. N.
(1983),
Correlated
firingof cat retinalganglioncells. I.Spontaneously active inputstoX- and
cells,JburnalofNeuroplrysiolqgy,49,303-324,
Meister,M.,Lagnado,L.,& Baylor,D. A.
C1995)
Cencertedsignaling
by
retinal ganglion cells.Science,270,1207-121O.Nirenberg,S,Carcieri,S,M.,
Jacobs,
A.L.,& Latham P.E.(2001).
Retinalganglioncells act 1argelyas independentencoders.Nbture,411,698-701.
Oyster,C.W, Thkahashi,E.,& Collesvijn,H.
(1972),
tien-selectiyeretinal ganglioncells and control ofnetic nystagmus intherabbit. VisionResearch,12,183-193.
Redieck,R.W
(1967).
Maintained activity of cat retinalganglioncells.Jburnal
ofIVeurophysiolqgy,
30,1043-le71.Salzman,
C
D,,Murasugi,C,M.,Britten,K,H.,& Newsome, W r(1992).
Microstimulationinyisual area MT: effectsondirectiondiscrimination
performance,
Journal
of
science, 12,2331-2355.
Singer,VLL,& GraF
C.
M,(199S).
Visualfeature
integrationand thetemporal correlation hypothesis.Ann"alRevieiv
of
Nleuroscience,18,555-586,
TS'o,D.YL,&
Gilbert,
C,
D.
(1988).
The
organization ofmatic and spatial interactionsinthe primate striate cortex.
journal
ofNburoscience,
8,1712-1727.1ls'o.
D.Y.,Gilbert,C.D.,& WieseLT.N.(1986).
shipsbetween
horizontalinteractions and functionaltectureincat striate cortex as revealed bycross-correlation
analysis. Jburnal
ofIVeuroscience,
6,1160-1170.Ybnehara,
K.,Ishikane,H.,Sakuta,
H,,Shintani,T, Ybnehara,K.,Karnlji,N.L.,...
Noda,M,(2009).
tionof retinalganglion
cellsand theirprojections
involvedin centraltransrnissionof informationabout upward and
downward
image motion. PLoSONE, 4, e4320,Ybshida,K,,Watanabe,D,,Ishikane,H.,1[hchibana,M., tan,I.,& Nakanishi,S,
(2001),
A key role of starburstcrine cells inoriginating retinal directionalselectivity and
optokinetic eye movement. IVburon,30,771-78e.
Zeki,S.,Watson,
J,
D,,& Frackowiak,R.S.(1993).
Goingbe-yond theinformationgiven:the relation of illuseryvisual motion to brainactivity