The Japanese Society for Chemical Regulation of Plants (JSCRP)
NII-Electronic Library Service The Japanese Sooiety for Chemioal Regulation of Plants (JSCRP }
塵
フ
ラ ボ
ノ
イ ド生
合
成
遺
伝
子
と
花
色 調 節
*嶋
田
幸
久
(
理化
学 研究
所植物科
学 研 究セ ン ター
* * ) は じ め に 色 とりどりの美 しい花々 やテー
ブル をに ぎ わ す フ ルー
ツ や野菜, 秋の風情の代 名 詞とも言 え る 紅 葉 な ど 植 物の色は様々な形で 見る人の 日を楽 し ま せ て くれ る.
これ らの色は値 物が子孫を残すた めの戦 略と し て 用意し た も の であ り,
受 粉を媒 介し た り,
果実を食べ て種子を運ん で くれ た りする虫や鳥や その他の 動物を 惹 きつ け る た めの生 物 学 的 な 意 義 を持って い る.
これ らの植 物の色を担っ てい る色素 は大 きく分けて カロ チ ノ イ ドと フ ラボノ イ ド に わ か れ る.
中で もフ ラ ボ ノイ ドは青, 赤, オレ ンジ, 黄 色 など幅 広い色を担い, もっ と も多くの種で花 色を 受 け持っ てい る.
こ こ ではフ ラ ボノ イ ド の中で も 人 の目に見え る可視光を 吸収し花の 色素 と して中 心 的 な 役割を 担っ てい る アン トシ アニ ン の生合 成 と花色の関わ りにっ い て, 我々 が行 っ て き たFlavonoid−3/
,
5’
−hydroxylase
(F3 ’
5’
H
)を用い た花 色 の遺伝 子 操 作 を 中 心に紹介する.
F3 ’
5’
H
遺 伝子の クロー
ニ ン グや 同遺伝 子 を 用いた 花色の遺伝子操 作につ い て は, 日本のバ イ オ テ ク ノ ロ ジー
関連 企業であるサン トリー
社,キ リン ビー
ル社,そ して著 者らの所 属 して いた協和 醗 酵工業社の3
社の研 究グルー
プを中 心と し た激しい 研 究 開発競 争が行わ れ,
1993 年に 3社か ら特 許 として研 究 成 果 が 公 表さ れ た1−
3 〕.
こ の うち, サ ン ト リー
社の グルー
プ が多くの総 説を発 表して お り4−
S) ,ま たマ スコ ミが大きく取り上 げ た た め, 国 内では 専門分 野の研究者の間で さえ も 「こ の課題は すで に研 究が尽くさ れ た」との誤解が少なか ら ず 生 じた.
そこ で こ こ で は何が明 らか に さ れ, 何が 明 ら か に なっ てい ない の かを 再 検 証 しなが ら 紹 介 し,
これ から何が 必 要 なの かを展望し て み たい.
sFlowercolor eugineering with fiavonoid
−
biosyn−
thetic gens
,
By
Yukihisa
SH
【MADA (PSC ,
RIKEN ,
2
−
1,
Hirosawa,
Wako,
Saitama,
351−
019.
8)” 〒
351−0198
和光市 広沢Z−1
1
青い 花 色とF3 ’
5’
H
今日我々の身近で栽 培さ れて い る花は ほ と ん ど が育 種 家の乎による交配・
改 良を経て生み出さ れ た もの で あ り,
今日で も なお この品 種 改 良の試み は続 け られ て い る.
育 種の最も大き なター
ゲ ッ トと されて い るの が 青い花 を 欠 く種に青い 花の品種を作出 するこ と であ る.
切 り花 と して 最 も 多 く消 費 さ れてい るバ ラ , カー
ネー
シ ョ ン, チュー
リップ,
キ クに はい ずれ も青い 花 の 品種が存 在し ない.
その最 大の原 因と し て着 目さ れ たの がF3 ’
5
’H
で あっ た.
す な わ ち , 青い花の多くが3「
,5’
位に水 酸 基を持つ ア ン トシアニ ンを 主 要 な色素 として蓄 積 す るの に対 し, これ らの種は3’
,51
位に水 酸 基 を もつ アン トシ アニ ン を 合 成 す るこ と が で き な い.
そ こ で遺 伝子工学 的な手 法を用い て3f,5
’
位に水 酸 基 を もつ アン トシアニ ンを合 成させ るこ と が で き れ ば, これ らの種に青い花 を咲か せるこ とがで き る 可 能 性が あ る とい う産 業上 の 利用 価 値か ら各 社 がF3rsiH
遺 伝 子のクロー
ニ ング を日指した.
アン トシアニ ン を含む フラ ボ ノイドの生 合 成 経 路の 概 略を図1
に示 した.
ア ン ト シ アニ ン は配 糖 体も し く は アシ ル化 配 糖 体 として細 胞 内の液 胞に蓄 積す る.
ア ン トシアニ ン の ア グ リコ ン (色素 本 体 ) は アン トシア ニ ジン と呼 ばれ る.
ア ン ト シ アニ ジンに は オ レ ン ジの 花に主として蓄 積 す るpelargonidin, 赤 や ピン ク な ど の花に蓄 積するcyanidin とpeonidin,
青や紫の花に蓄 積 す る
delphinidin
とpetunidin と inalvodin が 存 在す る.
これ らア ン ト シ ア ニ ジン以 外に naringenin chalcone は黄色の花に蓄 積す る,
そ れ 以外の フ ラボノ イ ドは 淡 色か無 色で あ る.一
方,
アン トシ アニ ンの生 合 成経 路は網 目状で多くの酵 素が複 雑に絡み合っ て代 謝が進み, 産 物 が 合 成 さ れ る.
花 色の研 究によ く用い ら れ る ペ チュ ニ ア で はFlavonoid−3’
−hydroxylase
(F3SH
) はnaringenin をeriodictyol に,
dihydro
−
kaernpferol
をdihydroquercetin
に そ れ ぞ れ 変 換 す る.
一
方,F3
’
5’
H
は青い花の主 たる色素 成 分である3’
138
植 物の 化 学 調 節 N工 工一
Eleotronio LibraryHO
OH
OH O
Naringe
職in
cha星cone
CHl
嬰
ず
P
・
Coumaronyl −CoA
HO HO OH ONar ;ngenin OHF3
’H
HO↓
F3H
ク 唖 OH OHF3
’ ,5
’H
−一■
Dレyo
置F3H
HO 〃T
hQH O OHf OH OHPen
吐ahydroxy 衄avanone0 HO
黛
OH グN
OHF3 ’ ,5
’H
→ HH3F
↓
DihydrokaempferolF3
’H
OH ODihydroquercetin
N h OH ODihyd グNOH 朋コ
romyrlcetinFSo
「FLS
fiavones
orflavonols
DFRANSUFGT
DFRANSUFGT
1 l l H i i i i I H I b b OH lPeonidin
−
derivati
b 〕 卩:
:
:
:
Anthocyanins
:
フラボ ノイ ド合 成 系 路の簡 略 図 OHCyanidin・
derivative
OHPeonidin鬯
deri
▼ative HODFRANSUFGT
丶ミ + h 40H OH クN
OH OH OR1
)elphin
卩
derivative
I H° OH
Petunidin
・
derivative
OCH3 H° N , ハ 。H M彡 〃 OCH3 0R OHMalvidin
・
derivati
▼e 図 1Flavonoid−
3ノ
ー
hydroxylase
,
F3’
H;日avomid
.
.
3’
,
5「
.
−
hydroxylase,
F3
写H
;flav
〔〕ne synthase,
FS ;f!avonolsynthase
,
FLS ;F3且,
flavanone
.
3hydroxylase ;UFGT ,
UDP−
glucose:3・
○.
.
flavonoid glucosyl trans−
ferase;DFR
,
dihydroflavonol−
4−
reductase ;CHS ,
chalcone synthase ;CHI ,
chalcone 正somerase ;ANS , anthocyanidin synthase ;AMT ,
anthocyanin methyltransferase.
R represents the sugar side−
chain.
と
5t
位 に水 酸 基 を 持 つ ア ン トシ ア ニ ジ ン (del−
phinidin
,
petunidin,
malvodin )生合成の 鍵 酵 素で あ り, ペ チュ ニ ア で は naringenin とeriodictyoi をpentahydroxyflavanone に変 換し
,
dihydrokaernpfer・
01 と
dihydroquercetin
をdihydromyricetin
に変 換すVol
,
35,
No.
2
(2000
)る
.
ペ チュ ニ ア で はdihydrokaempferol
とdihydro
−
quercetin
,
dihydromyricetin はflavonol
synthase(
FLS
)に よっ てflavonol
に変 換 さ れ る.
ま た, キク科な ど植 物 種 に よっ て は naringenin とeriQdictyo1
,
peIltahydroxyHavanone は
fiavone
synthase (FS
) にThe Japanese Society for Chemical Regulation of Plants (JSCRP)
NII-Electronic Library Service The Japanese Sooiety for Chemioal Regulation of Plants (JSCRP }
よっ て
flavone
に変 換 さ れる.
フラ ボノー
ルや フ ラ ボ ン はコ ピグメン トと呼ばれ, アン トシアニ ン と共 存し て花 色に影 響を及ぼす.
図1
で は一
般 的な生合 成 経路 を 単 純 な 矢 印に よって示 し たが,
各 酵 素の存 否 や 酵 素 の基 質 特 異 性 は 種によって異 な り,
種 内の品 種によっ て も異な る.
こ れ ら酵素の存否, 酵 素の基質 特異性と 酵素活性の発現部位, 発現 時 期 が絡み合っ て複 雑 な花 色,
花柄を生み出す第一
の要 因 となっ て い る.
IIF3
’
5t
」e
遺伝 子の クロー
ニ ングF3
’
srH
遺 伝 子の クロー
ニ ン グ は 特 許 と して1993
年に公 開さ れ,
学 術 論 文と し て はIlolton
らが酵 母に おける機 能 発 現な ど を 元に同年 報 告してい る9).
そ の 中で,彼 ら はF3 ’
5’
H
遺 伝 子の相 補 実 験につ い て述べ てい る.
し か しなが ら, 「色 素の組 成が相 補される」と い う肝心のデー
タ が省略 さ れて お り, 今日 まで発表 さ れ なか っ た.
す な わ ち,F3
’
5’
ff
を 欠 損 す る 植 物 にE315
∫∫候 補 遺 伝子 を導入 し た場 合に,
欠損 植 物の表 現型 (アン トシ アニ ンの組 成 )が相 補さ れ る こ とを厳 密に証 明 し た報告 はこれ まで存 在 し な かっ た と 言 え る.
この原 因 は 研究を リー
ド し て きた 研究者が企業に 所 属して お り, 詳 細な デー
タ 公表が 企業秘 密として控 え ら れ た こ と が最大の原 因で あ ると筆 者 は 考 えて い る.一
方で最 近,
フ ラボノイ ド水 酸 化 酵 素で は ない遺 伝子4
岬 が ア ン トシ アニ ンの3 ’
J「「
位の水 酸 化と花 色に影 響 を与 えるこ とが 報 告 さ た 軌 従っ て,F3
’
5’
H
と報告された遺 伝 子が アン トシ アニ ン組 成を相 補 する こ とを色 素組 成の観 点か ら厳 密に再 検 証する必 要性が 生 じてい た.
そこで我々 は 「姐 砂 型の植 物 にAK14
をセ ン ス の方 向で導入し た場 合にHfl 一
型の植 物と表 現型 が同じ に な るこ と」を ア ン トシ アニ ンの組成か ら 検証 し た 結 果 を公 表 し た11〕.
こ の相 補 実 験の結 果か ら 我々 はAK14
がE3
’5H
遺 伝 子の cDNA で ある と結 論し た1).
我々が特 許として こ の結 果を 公開し た時に は, 高 等植 物か ら機 能 が 同 定 さ れ たP450
遺伝 子は他 に存 在 し なかっ た.
つ ま り,F3
’5
’H
遺 伝子 はCin−
namate−4−hydroxylaseiz
}と並ん で高 等 植 物か ら最 も 早 期に機 能が同 定されたP450
遺 伝 子で ある.
以 下に, 我々が行っ たF3
’
5 /
H
遺伝子の クロー
ニ ン グの手 法 につ い て紹介 す る.
ペ チュ ニ ア ゲ ノ ム に はHfl
と1
ガ2
とい う2
っ のE
γ5四 構造 遺伝子が存 在する と予 想されて い た13).
こ の う ち,Hfl
は花 弁 全 体で よ り強い活 性を発 現す る の に対 し, Hf2 は花 弁の 縁の部分で の み活 性 を 示 す13 ).
そ こ で我々 はHfl
にター
ゲッ トを絞っ て クロー
140
ニ ン グ を試みた.F .
?ノ5’H
は 祖 精 製 さ れ た 酵 素のin
vitro にお ける挙 動か らP450
酸 化 酵 素で ある こ と が 示唆さ れ てい た14).
P450
遺 伝子は スー
パー
ファ ミリー
を 形 成 して お り,
多 種 多 様 な 代 謝 反 応に関与するこ と が知 られ てい る15,
16 ).
例えば今日で はアラ ビ ドプシス のゲノム 中に は300
近い 数のP450
遺伝子 が存在す る と予 想 さ れてい る.
遺 伝 子 間の配 列 保 存 領 域 は非 常に 限 られて お り,
P450
で ある とい う情 報のみ を元 に 目的 の遺 伝子 をクロー
ニ ング する こ と は今日 で も非 常に困 難である.
当 時 我々 がF3 ’
5
・
ll
gene の クロー
ニ ング を試み てい た 時 期には 高 等 植 物から 単 離 さ れ たP450
遺 伝子は,
機 能が未 知のCYP71
ただ一
つ し か存 在し なかっ た17).
そこ で, 我々 はP450
ファ ミリー
で最も よ く保 存 さ れて い るヘ ム結 合 領 域に着目 し,
この領域を 増 幅するデジェ ネレー
トプライマー
を設 計し た.
この プ ライマー
を 用い てRT −PCR
を 行い , 青 花 品 種 のペ チュ ニ ア 花 弁で発 現 して い るP450
遺 伝 子の カタロ グ を作 製し た1B).
次に こ れ らP450
遺伝子の 中か ら,
青花 特 異 的に発 現する 僧’
5 ∫∫gene をクロー
ニ ング する た めにP450
遺伝 子 クロー
ニ ン グの新 手 法 を 開 発 し た1°).
まず,
花 弁で発 現 す る17
種のP450
遺伝 子 を特 異的に増 幅す るプライマー
を合成 し た.
一
方,del
−
phinidin を産 生 す る系 統に特異 的に発 現 し,Hfl
遺 伝 子座にコー
ド されてい る も のをス ク リー
ニ ング する目 的で, ペ チュ ニ ア のニ ア ア イ ソ ジェ ニ ック ラ イ ン (純 同 質 遺 伝 子 系 統 ) を 作 出 した.
す な わ ち,
delphinidin
を蓄 積す る青 色の 品種とcyanidin を蓄 積す る赤色の 品種を交雑し,これ を繰り返 して cyanidin を 蓄 積 す る 品 種に戻 し交 雑し た.
ニ ア ア イソ ジェ ニ ックライン の 花 弁か ら調 製し た cDNA を鋳 型と し て上記の17
種の プ ラ イマー
を 用い て single−
.
g pecific−
primerPCR20
}を 行い,F3
’
5
’
H
活 性の発 現 と遺 伝 子の発 現 が 連 鎖 す るP450
遺伝 子の 断 片を クロー
ニ ン グし た.
このP450
遺 伝子の断 片をプ ロー
ブ と してペ チュ ニ ア の cDNA ラ イ ブラリー
をス ク リー
ニ ン グす る こ とに より,F3
’
5’
H
遺 伝子の候 補と な る全 長 cDNA クロー
ンAK14
を 単 離 し た19〕.AK14
を 用い たペ チュ ニ ア相 補 実験の 結 果か ら我々 はAK14
がE3 厂
5 ’
∬ 遺 伝子の cDNA であ ると結 論 し た1D.
III
他の 植 物か らのF3 ’
5’
H
遺伝 子の クロー
ニ ング ペ チュ ニ アか らF .
?’5SH 遺伝 子が クロー
ニ ン グさ れ て以来, そ の遺伝子 を プV一
ブ と して多 くの植 物か らF3 ’
5’
H
遺 伝 子が ク ロー
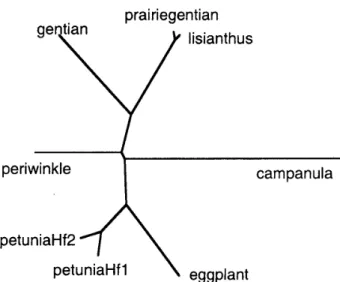
ニ ング さ れ て い る (図2
).
こ の うち, 我々 は ト ルコ ギキョウ (Eustoma
rZtssel一
植 物 の 化 学調節 N工 工一
Eleotronio Librarypo しunia 且fl P6tuniaU 重2 e 口9Pla 鳳t P【 己 iriegenti &n 1工8i 題n ヒhu8 98nt :魯n pe =↓wi 風k18 ca 題P邑【巳u1 匹
鑼
i
…
難
鑾
靉灘
韈
難
i
;
;
1
…
1
petuni &Hfl pe ヒunia9 重2 ●99P1 旦ロ匕 P【 己 iri 騒gen ヒ ユ己口 zi8 ⊥己n 七hu8 9●n七i驫 ロ pe=
ivinkle c 邑 ■Lpanu ↓a 33677188 55555655 potuni 邑 冨£ 1 po しu 鳥⊥眞HfZ e99Plant P「直工
=工
e 算entユ
“n li 已i轟nthu8 “ en ヒi臨 駄 po =iwi :L1【1e e 墨 甅P己nu1 己 1131t3116117117121 ユ13113 KT 肥 S謡
饑
鬻
一
: = = V一
胛
闇
一
一
陶
P P巴tuni 己亘窰1 POtU れi眞ヨ 虹2 0 ⊆ 百P1 贏n し P望airi8ge 血t 三& 皿 1i8i &顎しhu 屑 暫 o馳し工邑瓧 poriw ±nk10 e 魯 躍LP 邑nU1 轟 171171174175175179176178liiiiiiiii
覊
:TSKLPSSisisASV pe ヒU 駆i墨亘fl P6 ヒu 既i旦E藍2 09 臼Pユ魯n 七 pr 毳ir 二6qen し i邑
n li 隅工anthu8 gon ヒ↓轟ロ pa =工冒ink16 ¢ 魯 狙panu1 墨 22222a22s226225230227238 PO ヒUhi墨
Eft po ヒu 晶i驫 翼f2 099P1 農島ヒ P= 區 ⊥rio9 ◎ n しi驫恥 1i8 ↓ant’
hu 圃 9en 七i邑 n por ⊥”i見 1● c 昌■
LPB腿魁
1匹
工 1455968 888 巴 8889 22222322 potuni 墨月fl po ヒu 馳i購翼歪2 0督 πP1 驫島し pr 邑⊥蓄iogo 島ヒia口 1i 島i愚n ヒhu 醫 9en ヒi驫風 P6 コrilri 島 10 ¢ 邑 ■lp 臨晶u1 畠 11455968 ● 4444445 33333333 po セun 孟&琶重1 po ヒu口i邑区f2 07 官P1 邑 島 し pr 轟i】rioge 恥しian lis 工邑nthu8 gen しi醜 コ por 三1甲i塊kユo c 昌 囗 P轟:LU 1驫 400400403404404409405417 pa しuni 己ほ 重1 po しuni 驫口 £ 2 0 百9P1 昌 ロ し pr 己i【二■窟 ent ⊥an li5t 驫ロ亀hu8 9en ヒian perivinklo C 邑 ■ P邑【LUla 459 ‘59 ‘62 ●63 ‘63669 ‘64476 図2F3’
5’
H 遺伝子 にコー
ドさ れ る タ ンパ ク質の ア ミ ノ酸 配 列の比較lianum
)か ら遺伝子 を クロー
ニ ングし,
タバ コ発 現 系 を用い て機 能 を証 明 した21).
ま た 我々 は カン パ ニ ュ ラ (CamPanula
medittm )か ら も同遺伝子 を単 離し,
タバ コ発 現 系を用い て機 能を同 定して い る.
これ につ い て は 論 文 は未発表であるが, 特 許 1〕お よ び デー
タベー
ス(
The
accession number of theEMBL ,
GenBank
andThe Japanese Society for Chemical Regulation of Plants (JSCRP)
NII-Electronic Library Service The Japanese Sooiety for Chemioal Regulation of Plants (JSCRP }
図
3
F,
3’
5,
H 遺伝子の分 子 系統 樹 NJ 法の計 算 結 果に基づ いて分子系統 樹を描画 した.
DDBJ
databases,
Dl4590
)でデー
タ を公開し てい る.
こ れ ら 以 外 にHolton
らは ペ チュ ニ ア か らHfl と 劫2
に関す る2
つ の 遺伝 子 を 単 離 して yeast で発 現 さ せ て機 能を証 明し て お り9 同じグルー
プの 田中らは リン ド ウ (
Gentiana
tnt70ra
)由来の遺伝子 をyeastに お い て機能発現さ せ
,
基 質 特異性を報告して い る22 },
ま た最近で はKaltenbach
ら がニ チニ チ ソ ウ か ら単離 した 遺 伝 子 を大 腸 薗で機能発現 させ, 基 質特異性を報 告 して い る23}.
こ れ ら 以 外 に,Toguri
ら は ナス (Solanum
melongena )か ら24} , ま たNielsen
らはトル コ ギキョ ウ (E .
grandtveontm )か らL5)オル ソログ と示 唆される遺 伝子の クロー
ニ ン グに つ い て報 告 してい る.
ま た, 田中ら は トレ ニ ア,
ラベ ンダー,
バー
ペ ナ,
チョ ウマ メ か ら もF3
’
5 ’
H
を単 離し たと総 説 中で紹 介 して い る が,配 列等は未 公 開で あ る f’
}.
これ まで に配 列が公開さ れ たE3 ’
5 ’
H
遺 伝子につ い て分子系 統 樹 を作 製し た結果を図3
に示 す.
これ らF3’
5/
H 遺伝子 はP450
遺伝 子の 命 名コ ミッティー
に よっ てP450
遺 伝子 と し て の名 前が与え ら れ て お り,
全て がCYP75
ファ ミリー
のCYP75A
サブファ ミリー
に分 類さ れ て い る.
これ らの 遺 伝 子に はP450
遣 伝 子に特 有の コ ン セ ン サ ス配 列が保 存さ れ て お り,N
末 端に は ミ ク ロ ソー
ム膜 に局 在 する た め と思わ れ る疎 水 性の領 域 を有 し てい る.
こ れ ら既知のP450
遺 伝子の 中で我々の単 離し たカン パニ ュ ラ由来の クロー
ンが最も特 異な 配 列 を 有 してお り,
他の酵 素 とは 異 な る生 化 学 的 な 特 性 を 有 してい るこ と が 期待さ れる.、
142
IV
F3 ’
5’
H
遺伝 子 を 用い た花 色と アン トシ アニ ン 組 成の遺 伝 子 操 作ペ チ ュニ アF3 ’
5 ’
JJ
遺 伝子を用い た花 色の遺 伝子操 作は これ ま で たびたび総 説 等で そ の可能 性が論じ ら れ て き た の に対 し,
実際に組 み 換 え 植 物 を作 出 した 例 は, 最 初にHolton
ら が遺伝 子の ク u一
ニ ン グを報 告 し た際の1
例に とど まっ ていた9>.
我々 は,
遺 伝子工学 的な手 法を 用い てF3
’5iH 遺 伝 子の導 入 を行い ,ア ン トシアニ ン の組 成と花 色を実際に操 作で き る こ と を示 すた めに,
ペ チ ュ ニ ア由来の E3’
5 ’
遅 構造 遺伝子 をセ ン ス ま た は ア ンチセ ン ス の方向でCaMV35S
プロ モー
ター
の 下流につ ない で栽 培品種の ペ チュ ニ アへ 組 み込 ん だ11)。
まず 最 初に , 赤ま た は ピ ン ク色の品 種 (Falcon
Rose
,Falcon
Red ,
Falcon
Pink
Vein
>にセ ン ス 方 向の
E3 ’
5
’H
遺伝 子 を 導 入 し た.一
般 に赤,
ロー
ズ, ピ ン ク色 の花のペ チュ ニ ア 品種はE3 ’
5/
H
を欠 損 して お り,Flavonoid−
3
’
−
hydroxylase
(F3
,
H
)活 性を持っ てい る.
ペ チュ ニ アのdihydroflavonol
−4−
reductase (DFR
)はdihydrokaempferol
を基 質 と し な い た め,F3’
H
の産 物で あるdihydroquercetin が DFR に変 換 さ れ,
最 終 産 物 と し て cyanidin または peonidin タ イ プの アン ト シ アンが花 弁ア ン トシアニ ン の 主成 分を占める
.
・
遺伝子導入前のFa
正conPink
Vein
はう すい ピン ク地に赤紫の線の入っ た 花色で あっ たの に対 し
,
多 くの ト ラン ス ジェ ニ ッ ク個 体がNo .13
の ように,
赤紫 色の バ ッ クに濃い紫の線の入っ た花色に変化 し た (図4A
).
遺 伝子導入に よる花 色の差 はつ ぼみの段 階で最 も顕 著であっ た (図4A
).
その意 義に つ い て は後ほ ど 議 論 する.
次にFalcon
Rose
に導入 した例を図 4B に 示す.
遺 伝子 導 入前は明るい ピン ク色で あっ た が,
ト ランス ジェ ニ ックNo .102
は花全 体が明るい 赤 紫に変 化 し た.
また,
トラン ス ジェ ニ ッ クNo .106
は星 形に色 の変 化し た部分が現れ,
新しい模 様が出現 し た.
こ の 個 体で は赤 紫色の部 分の割 合は枝ごと に多 少変 化し た が, こ の模 様の花が安定して咲き続け た.
こ れ ら の ト ラ ン ス ジェ ニ ッ ク植 物か ら,
アン トシ ア ニ ン を抽 出し て加 水 分 解 処理 に よっ て アン トシアニ ジ ン に変換し,
その組 成をHPLC
で定量 した結果 を 表1
に示す.
赤や ピンク色の品種は前述の よ う にF3 ’
5 !
H
を 欠 損 ま た は 部 分 的に欠 損 す る た めに3
’ 位に水 酸 基 を持つ アン トシ アニ ジン (特に peonidin )を 主成 分と して 蓄 積 して い るこ と がわか る.
これに対 し,F3 ’
S’
H
遺伝子 を導入する と,
peonidin がほ と ん ど合 成さ れ な く な り, 代わっ て 3’
s「
位 両 方に水 酸 基 を持つ アン トシ 植 物の 化 学調 節 N工 工一
Eleotronio LibraryA
No
。13
B
control4
) ) 図 帆 償No
.
102
No
.106
ピンク色系 統の品種に対す るEヲ厂
5万 遺 伝子導入の効果Falcon Pink Vein 品 種の花とつ ぼみ
Falcon Rose 品 種の花
表
1
ピンク色系統品種とその組換え個 体の ア ン ト シ アニ ジン組 成3
’
−
hydroxylated 3’
,
5’
−
hydroxylatedcyanidin peonidindelphinidin petunidin malvidin
total
(μ9/gfw >
control Falcon RDse
transgenic No
.
102control Falcon Pink Vein
transgenic No
,
13 13= ln.
d.
8エ 3n,
d.
552±1 4±3144 ±933 ±4 n,
d.
2±1n.
d.
9
±2
1⊥179 土5n.
d.
19
±7
16±6459
±3270 ±13234 ±2
工 583±19544 ±30223 エ20295 ±27
トランス ジェ ニ ッ ク個体,
Falcon
Rose
No.
102,
Falcon Pink Vein No、
13 と遺伝 子 導 入前の同 品種の アン ト シ アニ ジン組 成を
HPLC
で分 析し た.
値は3っの花の平 均値土標 準 誤差.
n.
d,
は検出 限 界 以 下,
gfwは gram fresh weight を表す
.
アニ ジン (特に malvidin ) を主 成 分 と して蓄 積 す るこ と が わ か っ た
.
この こ と は,F3 ’
Jr’
H の存 在下 で はE3
’H
の 産 物が蓄 積 し な くな る,
すなわち見か け 上F3 ’
H
活 性 が 見 え な くなるこ と を意 味 す る.
こ の メ カ ニ ズム に つ い て も後ほ ど考 察す る,
この よ う な遺 伝子 導 入に よ る花 色と色 素 組 成の 変 化は, 遺 伝子導入 を 行っ たほ と ん どの植 物で観 察された.
つ まりペ チュ ニ ア で は,CaMV35S
プロ モー
ター
に よ る発 現 系で十 分 なF3 ’
5 ’
H
活性が発 現で き るこ と が わ かっ た.
次に,
紫や青の系統のペ チ ュ ニ ア品種(Falcon
Blue,
Cascade
Roya1 )にセ ン ス方 向のF3 ’
5’
H
遺 伝 子 を 導 入し た.
遺 伝子導入前のCascade
Roya1
は深み のある 丶ノol.
35,
No .
2 (2DOO)143
The Japanese Society for Chemical Regulation of Plants (JSCRP)
NII-Electronic Library Service The Japanese Soolety for Chemloal Regulatlon of Plants (JSCRP }
A
B
control5
) ) 図 値 旧No
.96
No
.60
紫 色系 統の品種に対する F.
?’
5/
H 遺 伝 子導入 の効果Cascade
Royal 品 種の花Falcon
Blue 品種の花 表2 紫色系 統 品 種 と その組 換え個 体の アン トシ アニ ジン組 成 3’
−
hydroxylated 3’
,5’
−hydroxylated
cyanidin peonidindelphinidin petunidin malvidin
total (μg/gfw )
control Cascade
Roya
ヱtransgenic
No .
33
control Falcon Blue
transgenic
No .
110 4t111 ±210 ±314 ! 16
±2478
±34
3.
L2577
.
L 43 30こ 53 ±222 ±3n.
d.
817= 75 1,
159:ヒ46 2,
16±114134
:ヒ17 275土13902
:t59 292±14 1,
593±92 1,
921±98
n,
d.
25
± 工615
±44
トランスジェ ニ ッ ク個 体,
CascadeRoyal
No.
33, Falcon BlueNo .
110と遺 伝子 導 入 前の同 品 種の ア
ン トシアニ ジン組 成を
HPLC
で分 析し た.
値は3
つ の花の 平均 値±標 準 誤差.
n.
d は検 出限 界 以 下,
gfw は gram fresh weight を表す.
青紫 色であるの に対 し, 多く の ト ラン ス ジェ ニ ック個 体が
No .
33
のよ うな薄 紫 色に変化 した (図5A
).
ま た,
トラ ン ス ジェ ニ ッ クNo .
60 は花弁の縁の 部 分 を 残 し て薄紫 色に変 化 した.
さらに,Falcon
Blue
に導 入し た ところ,
多 くの個 体はCascade
Royal
由来のNo ,33
の よ うな表現 型 を 示 し た が (結果省 略), 中に はNo
,
96
144
の よう な 斑 入 り状に色が変 化 す る ものが低 頻 度で現 れ た (図5B
).
これ らの 青 色 品 種 由来の ト ラン ス ジェ ニ ッ ク植 物か ら
,
アン トシアニ ジン色素の組 成 を定 量し た結 果を表 2 に示 す.
紫や青の系 統のペ チュ ニ ア品 種で あるCas
−
cadeRoyal
もFalcon
Blue
も遺伝子導入前の個体は植 物 の 化 学 調 節
peonidin とmalvidin がアン ト シ アニ ジン組 成の大半 を占め て い る
.
一
方 トラ ン ス ジェ ニ ック個 体で はpeonidin
, malvidin の蓄 積が阻害され, アン トシ アニ ジン蓄積量 全体が減 少し てい る.
この こ と か ら, ト ラ ン ス ジェ = ッ ク植 物では 導 入 し たF3
’5
,H
遺 伝 子 が 内 在性のF3 ’
5’
H
遺伝 子 の 発 現を押 さ え る コ サ プ レ ッショ ン と呼ぼ れ る現 象が起 こ り, 内 在 性 のF3 ’
J「’
ff
活性が 阻 害され たこ とが 予 想できる.一
方で 注 目 すべ き 点 は, トラン ス ジェ ニ ック個体で peonidin の蓄 積が大 幅に増 加し てい る点で あ る.
こ の こ と は,青 色 品種がR
γ∫f
酵 素を持 っ てい る にもか かわ らず,外 来性のE315 戸
H
遣伝 子 を導入 する前に は,3
’
位に水酸 基を持っ アン ト シアニ ン を合 成し ない こ とを示 し てい る.
す な わ ち,E3 〆
5
烈 活 性が存 在する状態で は, 見 かけ一
lt
F3rH
活性が 無 く な る よ うに見 え る わ けであ る.
一
般に野 生 型のペ チュ ニ ア で はF3
厂
5
野 とF3
’
u
の両 者が共 存す る場 合に は3
’
St
位 両方 に水 酸 基 を持 つ ア ン トシ ア ニ ンが蓄 積 す る こ とが知 られて い た が13),
今回の表2
の結果は,F3
’5 ’
ff
遺 伝子が内在性 プロ モー
ター
の制 御 下で発 現して もCaMV35S
プロ モー
ター
の制御 下で発 現してもこの現 象 が 起 きるこ と 示して い る.
現時 点で はこ の現象はペ チ ュ ニ アの フ ラ ボノ イ ド生合成 酵 素の基質特 異 性と酵 素の競 合から以 下のよ うに説 明でき る.
ペ チュ ニ アのDFR
はB
環の 水 酸 基が よ り多いdihydroflavono
工を 基質と し て好む 性 質が ある.
す なわ ち,dihydromyricetin
を最も効率 よ く基質として変 換するの に対し,dihydrokaempfer−
ol は 基 質 と な ら ない.一
方,
ペ チュ ニ アflav
()nol synthase (FLS
)の基 質 特 異性は逆で,B
環の水酸基 が よ り少 ないdihydroflavonol
を 基 質 と して好 む性質 がある.
す な わ ちdihydrokaempferol
を もっ も効率よ く変 換 し,dihydromyricetin
はあま り変換 しない.
従っ て,E .
9’
5fH の 存 在 下で はdihydromyricetin
が 生合 成 中 間体 と し て多く存 在する と想 定で き る の で, この場合に は3’
5’
位に水 酸 基 を 持つ アン トシ ア ニ ン が主 成 分 と して蓄 積 し,一
方E9
〆5 野 活 性 を 欠 損 す る 品種で,FLS
が存 在す る条 件で はFLS
の働 きに よっ てfiavonol
が蓄 積し, 同時に アンb
シアニ ン として は 3’
位に水酸 基 を持つ もの が蓄 積する こ とに なる.一
方,FLS
が存 在しない 品 種で は もっ ぱ ら3’
位に水 酸 基 を 持つ ア ン トシ アニ ンが蓄 積 するこ とに な る.
しか し な が ら他 方で, 転 写レ ベ ルや酵 素 活性の調 節レ ベ ル でF3
’5
’H
がF3
’H
の活 性 を押 さ え る何らか の メ カ ニ ズ ム が存 在する可能 性も現時 点で は否定で き ない.
一
方で, ピン ク 系 統のペ チュ ニ アにF3
’5’H
を 導 入 するとつ ぼ み の段 階で最も顕 著な色の違いが観 察され るこ と を既に紹介 し たが, この メ カニ ズム は発達 に伴 う生 合成 遺 伝子の発 現の特 徴に よっ て次の ように説明 で き る.
花の形成過 程 に おい てflavonol
の生合成は anthocyanin の生 合 成 よ り も早 く起 こるこ と がペ チュ ニ アで は知 ら れて い る.
つ まり非組換え体の植 物の つ ぼ み は, ア ン ト シ アニ ン よ りもフ ラボノー
ル が 主とし て蓄 積 してい る.一
方, セ ン ス 方 向のF3 ’
5 ’
H
遺伝 子 をCaMV35S
プロ モー
ター
の制御下で発 現 す る トラ ン ス ジェ ニ ック個体のつ ぼ み で は, 非 組 換え体よ り もdihydromyricetin
が多量 に生 合 成中間 体 と して蓄積 して い る こ と が予想で きる,
こ のdihydromyricetin
は 前 述の ようにFLS
に よっ て変 換さ れ に くいた め,組 換 え個体で は フ ラボノー
ル の蓄 積が 抑制さ れ, 代わっ て アン トシ アニ ンの合 成 量が増 加 す るこ と が予 想で き る.
次に,紫や青の系統のペ チュ ニ ア品種(Falcon
Blue,
Cascade
Royal
>に アン チセ ン ス方 向のF3 ’
S’
H
遺 伝 子 を導入実 験を実 施し た.
この時 得ら れ た トラ ン ス ジェ ニ ック個 体はセ ンス方向の遺伝子導入 と同様の花 色 変 化 を示 し た が, その出 現 頻 度 はセ ン ス方 向の遺 伝 子導入 と比 較し て極め て低 頻 度で あっ た.
従っ て,
F3 ’
5 ’
H
遺伝子の発現 を押さえる に はセ ン ス 方向の 遺伝 子 導 入がアンチセン ス方向より もより効 果 的であ る と考えてい る.一
方,一
連の トランス ジェ ニ ッ クペ チュ ニ ア作出 実 験の中で, 矮 化 し た 植 物 や帯化 し た 植 物が低頻度で得られた が, それら は導 入 し たコ ン ス ト ラ ク トの方 向や導入親と なっ た品種 と無 関係に得ら れ た た め, 導入遺 伝 子に よ る効 果と は無 関係である と判 断 した.
V
F3 ’
5’
H
遺 伝子の異 種 植 物での 発現一
トラン ス ジ ェ ニ ッ クタバコの例 「EgtsfH
遺 伝子 を欠損する植 物にF3
’S
’H
遺 伝子 を導入 する と花色 が青くなる の は当然であろう」とい う趣 旨の コメ ン トを 国 内の専 門 家か ら頂 戴 した こ とが あ る.
し か し な が ら, その よ う な事実は こ れ ま で一
度 も証明さ れ た こ とが ない ばかりでな く, クロー
ニ ン グ されたF3 ’
5’
H
遺伝 子 を 異 種植 物で 発 現 するこ とに 成 功し た とい う報 告も存 在し な か っ た.
さ ら に,P450
で あるF3 ’
5’
H
遺 伝 子 を植 物 に導 入 する際には, その 活 性を十分に発 現さ せ るこ とすら難しい であろ うとい うこ と が以前か ら予 想でき た25}.一
般に,P450
遺伝 子 を発 現さ せ て酵 素 活 性 を 示 す こ と は今 旧で も難 しく,
多 くのP450
遺 伝 子の酵素 活性が, 生 化学的には証 明 Vol,
35,
No,
2 (2000)145
The Japanese Society for Chemical Regulation of Plants (JSCRP)
NII-Electronic Library Service The Japanese Sooiety for Chemioal Regulation of Plants (JSCRP }
図6 タバ コ に対す る1r3
’
5’
H
遺伝子導入の効 果 左か ら そ れ ぞ れ pBll21,
pTI201 (トル コギキョ ウ 由来F3 ’
5’H
),
pB853 (ペ チュ ニ ア由 来F3 ’
5’
H
) を導入 し た組 換え個体の タバ コ の花 され ない まま と なっ てい る.F3
’
5’
H
遺 伝子 を欠損す る植 物に クロー
ニ ング した同遺 伝子を導入し て 目的の 産 物を蓄 積 させ る た めに は, 種 間組 換え で組み込ん だ F3’
5 ’
H
遺 伝子が効 率 よ く働 く必要が ある.
そ こ で 我々は その一
例 と して,
ペ チュ ニ ア お よ び トル コ ギ キョ ウ か ら クロー
ニ ング し たF3 ’
5 ’
H
遺 伝子 を タバ コ (八Jicotianatabacum
cv,
Petit
Havana
SR1
)へ 導入 し
,
色 素 生合成の変 化を検 証 し た21).
我々 が用い た タバ コ の
Petit
Havana
SRI
品 種 はF3
’5’H
me
伝子 を 欠損し て お り,
cyanidin を 主た る ア ン ト シ アニ ン と して 蓄積してい る.
こ こへ ペ チュ ニ ア 由 来のF3 ’
5 ’
H
を発 現さ せ るコ ン ス トラ ク トpB853 と トル コ ギ キ ョウ由 来のF3 ’
5’
H を発 現 さ せ るコ ン ス トラ ク トpTG210 を導入 し た.
ペ チュ ニ ア 由 来 遣 伝 子の コ ン ス ト ラ ク ト は,
前述のよ うに タバ コ の近縁種 ペ チュ ニ アでは 十 分機能 する こ と を確認 し てい る.
ま ず, コ ン トロー
ル と してGUS
遺 伝子を発現さ せ た ト ラ ンス ジェ ニ ッ ク個 体は花 色が変 化せず,
薄ピンク 色 の花 を 咲か せ た.
一
方F3 ’
5 ’
H
遺伝 子 を 発 現さ せ た ト ラ ンス ジェ ニ ッ ク個 体は, わずか に花 色が濃 くなっ た (図6
).
花 色 が 変 化す る ト ラン ス ジェ ニ ック個体はペ チュ ニ ア と同等の高頻 度で 得 られ た が , 変化の度 合い はペ チュ ニ ア で観 察さ れ た も の と比べ て非 常に限 られ た もの であっ た.
ま た,
遺伝子の 由 来,
つ ま りペ チュ ニ ア と トル コギ キョ ウ の遺伝子間の違い は認 め ら れ な か っ た.
ペ チュ ニ ア お よび トル コ ギキョ ウ由 来のE3 ’
5
’
H
を発 現 する トラ ン ス ジェ ニ ック 個 体の う ち , もっ とも 顕 著に花色が変 化し た個 体の花 弁か ら ア ン トシアニ ン を抽 出し,
アン トシアニ ジン色 素の組 成を 分 析 し た (表3
).
コ ン トロー
ル と してOUS
遺伝 子 を 導入 し た個体 で は3’
位の みに水 酸 基 を もつ cyanidin の み が検出 さ れた の に対 し,
F3
’
5 ’
H
遺伝子を導入 した 個 体 で は cyanidin に加 え てF3rstH
の産 物で あるdelphinidin
が観 察さ れ た.
しかし, ト ランスジェ ニ ッ クペ チュ ニ146
表3
トランスジェ ニ ッ ク タバ コ のアン トシアニ ン 含 量Total
Cyanidin
Delphinidin (μg/gfw ) pBll21 242 (100) pTG201 ユ91 (77) pB853 152 (65) n,
d.
57
(23
>82
(35
) 242248234 値は5
つの花の平均 値,
カ ッコ 内の数 値は 個々 のアン ト シ アニ ジン の含量 を 百 分 率で表 す.
n.
d.
は検 出 限 界 以 下,
gfw はgramfresh
weight を表 す.
アで 得られた結 果と比較す る と その量は非 常に 限 られ て お り, 最大で
35
% であっ た.
我々 は100
個 体以 上の ト ラン ス ジェ ニ ッ クタバ コ個体を作 出し た が,
これ 以 上 デル フ ィニ ジン含量の多い個 体 を 作 出 する事は で き なかっ た.
一
方,
トラン ス ジェ ニ ッ ク植 物が デル フ ィ ニ ジン を蓄 積す る形質は,
次 世 代に安 定 して遺 伝す る こ とを確認で き た.
ト ラン ス ジェ ニ ック タバ コが な ぜ限 られ た 量のデル フ ィニ ジン し か蓄積で き ないの か とい う理 由につ い て は多くの原 因が考え ら れ, 現時点で は判 断 を下 すこ と は難しい。
例え ば, 前述のペ チュ ニ ア の項で述べ たよ う に,F
ヨ’
5 〆
H
と競 合 する内在 性 酵 素の 基 質 特 異 性に よっ て説 明す るこ とも可能で ある21〕.
しか し,筆 者ら はP450
と し ての 活 性 発 現 系に原因が ある可能 性も考 え て いる.P450
が 活性を発 現す るに はP450
に電子を供 給す る ミ クロ ソー
ム電 子 伝 達シ ス テ ム が機 能する必 要 がある.
この電子伝達経路の生理 的 な役 割は ほ と ん ど 解 明さ れ てい ない.P450
に電 子 を 伝 達す るNADPH
−
P450−
reductase は動 物 や 微 生物の場 合と異 な り,
高等 植 物で はア ラ ビドプ シス です ら2
つ の遺 伝 子に よっ て コー
ド さ れ てい る こ と が明 らか になっ て お り25・
2 η,
こ の2
つ の reductase の存 在 意義は現 在 不 明で ある.
ま た,
最 近de
Vetten
ら は 砌四 とい うcytochromeb5
をコー
ドす る遺 伝子がE3 ’
5’
H 活性の 十 分な発 現 に 植 物 の 化 学.
調 節 N工 工一
Eleotronio Library必 要で あ る こ と を 示 し た1G }
.
し た が っ て, こ れ ら電子伝 達 シ ス テム の因子 が十 分にそろわ な け れ ばF3
’
5 ’
ff
が十分 な 活性を発揮で き ない可能性があ る.
お わ り に こ こ で はい くつ かの 具体 例を挙 げなが らF3
’5
’U
遺 伝子 を用いた フラボノ イ ド組 成と花 色の遺 伝子工学 的操 作につ い て紹介 し, 考察し た,
今日の技 術で は試 行 錯誤的にF3f51H
遺 伝子 を用い て花 色をコ ン ト ロー
ル するこ と は可 能である.
実 際にこの技 術は実用 化 され, 紫色のカー
ネー
シ ョ ンが 産み出 さ れて実際に 商 品 化 さ れて い る6〕.
し か し,
こ こで示 し た よ うに,F3JstH
遺 伝子 をや み く もに異種の 植 物に 導入 し た と し て も, 花色が青 くな ら ない ばか りで な く, アン ト シアニ ンの組成 を変 化 させることす ら 容 易で は ない こ とが わか る.一
方,
青い花の花色の発 現 機 構 は 徐々に 解 明さ れつ つ あるが, 多くの花で未だ謎であ り, フ ラ ボノ イ ド色素の平 面構造, 立体 構造 と花 色の関係 が 今 後 も解明されてい く必要があ る。
また,
遺 伝子工学 を 用い て望み の色 素を蓄積させ る に は まだまだ解 決し な け れ ば な ら ない 技術的 な 側 面 が 多い.
特に,P450
の活 性 を 安 定して発 現さ せ るには同 酵 素の活 性 発 現の メカ ニ ズム解 明が必 須である.
ま た,E3
!
5 ’
H
は アン ト シ アニ ン合成 経路で最 大5
種類の酵 素と基質を取り合っ て競 合 す る た め (図1
), 希 望 通 りの色素 組 成 を実現 す る た めに は, そ れ ぞ れ の競合酵素の基質特異性と その 種 間 差, 発 現 部位,発 現時期が解 明さ れる必 要 が ある.
特に, 青 色の花 を 作 るた めに必 要 と され るフ ラ ボノイ ドや フ ラボノー
ル な ど の コ ピグメン ト を効 率よく蓄積 させ るためには,
これ らの合 成 酵 素 を 単 純に発 現させ た と して も酵 素に よ る 基 質の取 り合いが生 じて,
アン トシ アニ ン の蓄 積が減 少し て花 色が う す く な る こ と が 我々 の研 究 結果か ら も予 想できる.
この場合にはフ ラ ボノ イ ドの 生 合成を上 流か ら太 くす る た めに生 合成の 制 御因 子を発 現さ せ る な ど の 工夫が必 要と なっ て くる であろう.
望み の色の花 を遺 伝子工 学 的に作 り出す た めには,
設 計 通 りの構 造の色素 を 設 計 通 りの組 成に導 くた めの技 術が必 要であ り, その た めには花の どの発 生 段 階にどの場 所で どのよ う な 基 質特異[生の酵 素 を発 現さ せ るの か とい う5
次元的な酵 素 活性の発 現制 御が 必 要 となる.
フ ラ ボノ イ ドの生合成は植 物の 2 次代謝 の中で も最も複 雑で多様な構 成を持っ.
そ の人為 的な 制 御に は まだ解泱すべ き課題が山積さ れ てい る.
謝 辞 本総説で紹介した 研 究 成 果 は 協 和 醗 酵工業 株 式 会 社 筑 波 研 究 所 植 物 研 究グルー
プに おい て菊池 泰 弘 博士, 清 川繁人 博 士, 大林正 也 氏, 冲中泰博士, 嶋田律子氏 と の共 同 研究として 得 ら れ た もの で あ り, 共 同 研 究 者 の 方々に お礼 申し上 げます.
筑 波 研 究 所 植 物 研 究グ ルー
プ は企業の方 針に よ り1994
年を もv て解 散い た し ま し た が,
当 時 研 究 所に在 籍され 本 研 究に ご協 力 賜 りま し た協 和醸酵 社員 な ら びに元社員の皆様に この場 を 借 りて厚 くお礼 申 し上 げ ま す.
本総 説執 筆にあ た り ご助 力い た だ き ま し た 吉 田 茂男 博士 (理 化学研究 所)に 感 謝 申し上げま す.
) 1 )2
) 3 ) 4 5ハ
7 ) 8 ) 9 10) 11) 12)13
)14
) 15) 文 献YKikuchi ,
S.
Kiyokawa ,
Y、
Shimada
, R.
Shimada ,
M .
Ohbayashi
and Y.
Okinaka,
Intemaa−
tional Patent
No .
WO9318155
(1993
)T .
A 、
Holton
andY ,
Tanaka ,
Jntemaational
Patent
No .
WO9301290
(1993>T .
Ohtani,
T .
Toguri
, N
.
Umenloto
andO 、
Kobayashi ,ノ
itPanese
Pa tent No.
JP5
/84370 (1993)Y
.
Tanaka,
S.
Tsuda and T.
Kusumi,
PlantCel9
1
〕hysioi
39,1119
(1998
>田中 良 和
,
久 住高 章,
植 物の化 学 調 節33,
55 (1998)田中 良 和
,
榊 原圭 子,
実 験 医 学18,
767 (2000>T
.
Holton and E.
Cornish,
PlantCelZ
7,
IO71
(
1995
>T .
A .
Holton,
Drug
Metabogism
andP
Interac−
tions
12,359
(1995)T
,
A .
Hol亡on,
F.
Brugliera,
D.
R.
Lester,
Y.
Tana・
ka
,
C.
D,
Hyland,
」.
G .
T .
Menting,
C,
−Y ,
Lu,
E.
Farcy
,
T .
W .
Stevenson
andEC .
Cornish,
ハ/ature366,276
(1993
)N .
de
Vetten,
J
.
terHorst
, H
.
−P .
van Schaik, A.
deBoer
,
J.
Mol and R.
Koes,
Proc.
Natl.
Acad.
Sci.
〔JSA 96
,778 (1999>
Y
.
Shimada,
M.
Ohbayashi,
R、
Nakano−Shimada,
Y .
Okinaka,
S.
Kiyokawa
andY .
Kikuchi
,
Pgant
Cell
RePort,
in
press(2000)H
.
Teutsch,
M,
Hasenfratz,
A、
Lesot,
C.
Stoltz
,
J.
−
M,
Garnier,
J.
−
M.
Je
!tsch,
F.
Durst andD .
Werch
.
・
Reichhart,
Proc.
翫 鼠Acad.
Sci.
USA
90,
4
工02
(1993
)H .
Wiering
andP .
D .
Vlaming,
in
Petunia
(ed.
Sink,
K .
C.
)49
(Springer−Verlag,
Berlin
,
1984)W .Heller
andG .
Forkmann ,
in
The
fiavonoids
(ed
.
Harborne,
J.
B.
) 399 (Chapman and Hall,London,
1993)D