Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬科学) 報 告 番 号 甲第1768号 学 位 記 番 号 第365号 氏 名 山田 亜紀 授 与 年 月 日 令和 2 年 3 月 31 日 学位論文の題名 イチゴおよびモミジバダイオウにおける配糖化酵素の機能解析 論文審査担当者 主査: 中川 秀彦 副査: 牧野 利明, 星野 真一, 佐藤 匡史
名古屋市立大学学位論文
イチゴおよびモミジバダイオウにおける配糖化酵素の
機能解析
令和元年度
(2020 年 3 月)
名古屋市立大学大学院薬学研究科
生薬学分野
山田 亜紀
1. 本論文は2020 年 3 月、名古屋市立大学大学院薬学研究科において審査されたもの である。 主査 中川 秀彦 教授 副査 牧野 利明 教授 星野 真一 教授 佐藤 匡史 准教授 2. 本論文は学術情報雑誌に収載された次の報文を基礎とするものである。 基礎となる報文
① Aki Yamada, Kan’ichiro Ishiuchi, Toshiaki Makino, Hajime Mizukami, and Kazuyoshi Terasaka
A glucosyltransferase specific for 4-hydroxy-2,5-dimethyl-3(2H)-furanone in strawberry Bioscience Biotechnology and Biochemistry 83, 106–113 (2018)
② Aki Yamada, Miku Kaneko-Kondo, Kan’ichiro Ishiuchi, Toshiaki Makino, and Kazuyoshi Terasaka
Isolation and characterization of a novel glucosyltransferase involved in production of emodin-6-O-glucoside and rhaponticin in Rheum palmatum
Plant Biotechnology : in press (2020)
3. 本論文の基礎となる研究は、牧野 利明 教授の指導の下に名古屋市立大学大学院 薬学研究科において行われた。
【目次】 序論 1 本論 第一章 イチゴ香気成分の配糖体の生合成に関わる配糖化酵素の単離と機能解析 4 第一節 データベース検索と候補遺伝子のクローニング 6 第二節 組換えタンパク質を用いた配糖化活性の解析 9 第三節 遺伝子発現とHDMF 配糖体の蓄積 13 考察 14 第二章 モミジバダイオウのアントラキノン配糖化酵素の機能解析 17 第一節 モミジバダイオウからの配糖化酵素遺伝子の網羅的クローニング 18 第二節 組換えタンパク質を用いた配糖化活性の解析 21 第三節 遺伝子発現解析と代謝産物の蓄積 26 考察 28 結論 30 実験方法 31 引用文献 40 謝辞 44
本論文では以下のような略号を用いた (アルファベット順) 。
aa : amino acid bp : base pair
CBB : Coomassie Brilliant Blue
DMMF : 2,5-dimethyl-4-methoxy-3(2H)-furanone E1G :emodin 1-gluoside
E6G :emodin 6-gluoside E8G :emodin 8-gluoside
FaGT : Fragaria x ananassa glycosyltransferase
FaQR/FaEO : Fragaria × ananassa quinone oxidoreductase/enone oxidoreductase FaOMT : Fragaria × ananassa O-methyltransferase
GT : glycosyltransferase
HDMF : 4-hydroxy-2,5- dimethyl-3(2H)-furanone His : histidine
HMMF : 4-hydroxy-5-methyl-2-methylene-3(2H)-furanone HPLC : high performance liquid choromatography IPTG : isopropyl β-D-1-thiogalactopyranoside IR : immature receptacle
kcat : turnover number
Km : Michaelis constant
MeOH : methanol
MR : mature receptacle
NMR : nuclear magnetic resonance PCR : polymerase chain reaction
PSPG : plant secondary product glycosyltransferase RACE : rapid amplification of cDNA ends
Rp : Rheum palmatum
RT-PCR : reverse transcriptase- polymerase chain reaction
SDS-PAGE : sodium dodecyl sulfate polyacrylamide gel electrophoresis UDP : uridine diphosphate
1 【序論】
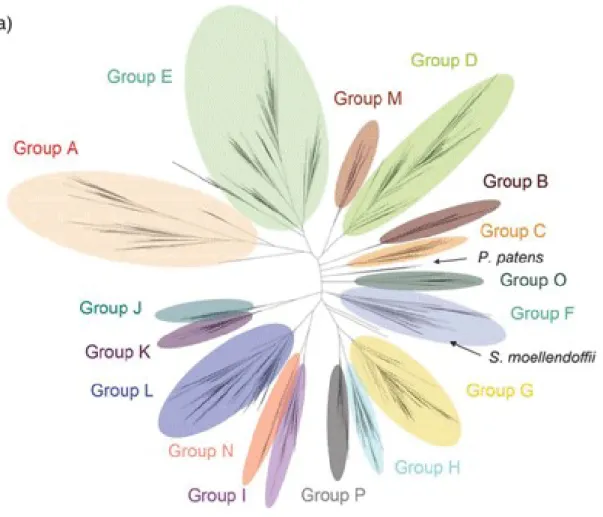
人類と植物の関係は多様であり、古来、食物連鎖上の消費者と生産者の関係にとどまらず 建材や繊維、エネルギー生産などの原料などとして様々な形で利用してきた。19 世紀初頭 のケシ (Papaver somniferum) からの morphine の単離によって植物が生み出す化学物質が生 理活性を示すという事実が発見されると、以降の医学、薬学などが急速に発展するようにな った。 植物が生み出す化合物のうちで重要なものの一つが、morphine のような二次代謝産物と 呼ばれる低分子有機化合物である。二次代謝産物は、植物種によって異なる進化を遂げた二 次代謝経路から生合成され、化学構造だけでなく、生理活性の面からも非常に多彩な化合物 群となっている。植物二次代謝産物は、植物自身の生存にとって必要であるばかりでなく、 医薬品、食品添加物、香料、工業原料など、私たちの生活の様々な場面で活用されている。 それらはその構造と生合成経路によって分類されているが、多くの化合物に共通した構造 として、水酸基などに付加された糖残基の存在が挙げられる。この糖残基は生合成経路の最 終段階で付加されることが多く、植物二次代謝産物に糖残基が付加されて配糖体となるこ とが、植物体内での二次代謝産物の蓄積に一定の役割を果たしている1。 植物が含有する配糖体の生成を担っているのが、植物配糖化酵素であり、特に植物におけ る二次代謝に関わる分子種は、植物二次代謝糖転移酵素 (PSPG, Plant Secondary Product Glycosyltransferase) と総称され、ヒト (Homo sapiens) の薬物代謝酵素の一種であるグルク ロン酸抱合酵素と同じくFamily 1 に属する糖転移酵素群として知られている2。PSPG は、 植物内で生合成される様々な低分子化合物をアクセプター基質とし、UDP-glucose などの nucleotide-activated sugar から糖を植物二次代謝産物へ転移する反応を触媒する2,3。PSPG の 多くは、二次代謝産物の生合成に関与するものとして単離、機能解析が行われてきた。それ 以外の機能として、PSPG は植物ホルモン濃度の制御など植物の生存に必須の化合物の細胞 内のホメオスタシスの維持に寄与している4。さらには、外来性の低分子有機化合物が植物 体内に取り込まれた際に、毒性を回避するシステムの一つとして、すなわち、PSPG が外来 低分子有機化合物を配糖化することで水溶性を増大させ、液胞へと輸送・蓄積するメカニズ ムの一部として機能している5–8。このように、PSPG は種々の内在性、外来性基質を配糖化 する機能があり、植物はそれぞれに対応した基質特異性を持つ数多くのPSPG をゲノム中に 有している 1–7。全ゲノム配列の解読が植物で最初に行われた、モデル植物シロイヌナズナ においては、約25,000 の遺伝子の中に 100 種類以上の PSPG が存在していることが報告さ れた1。シロイヌナズナ由来PSPG は、アミノ酸配列の相同性から分子系統樹上で Group A からGroup N までの 14 のグループに分けられているが、近年、他の植物種でもゲノム配列 が明らかになり、シロイヌナズナには含まれないGroup O と Group P が追加され、16 のグ ループに拡大された9。それぞれのグループで配列の相同性と基質特異性に相関があるとさ れ、例えば、グループA に属する配糖化酵素は配糖体の糖鎖伸長、グループ L に属する配 糖化酵素はカルボキシル基を配糖化することが知られている2 (Fig. 1)。
一方、天然で含量の少ない配糖体を生産する、非天然の配糖体を生産するといった有用物 質生産の観点から、PSPG を利用した様々な試みが行われている。その中には、有用化合物 の水溶性の増大や安定性の向上、毒性の軽減あるいは生理活性の高機能化を目的とした配 糖体の生産などが挙げられる8。 植物における二次代謝経路の解明の点からも、物質生産の点からも注目されているPSPG ではあるものの、天然に存在する多数の配糖体と植物ゲノム中に存在する多数のPSPG との 関係性は十分に明らかになっているとは言えず、種々の配糖体が生成される際のPSPG が同 定されていないケースが多い。そこで本研究では、植物に含まれる配糖体の生合成過程にお いて未知の配糖化酵素を同定することを目的とした。具体的には、イチゴの重要な香気成分 である4-hydroxy-2,5-dimethyl-3(2H)-furanone (HDMF) の配糖体と、モミジバダイオウの重要 な成分であるアントラキノンの配糖体の生合成に関わる配糖化酵素を単離した。さらに、こ れらの配糖化酵素の基質特異性や遺伝子発現について解析を行い、HDMF 配糖体およびア ントラキノン配糖体の生成への寄与について明らかにした。
3
【本論】 第一章 イチゴ香気成分の配糖体の生合成に関わる配糖化酵素の単離と機能解析 イチゴ (Fragaria × ananassa) はバラ科オランダイチゴ属に属する多年草で、ビタミン C やキシリトール、アントシアニン類を豊富に含み、主に花托の部分が食用とされている10,11。 イチゴの花托は食用とされるだけでなく、近年では、閉鎖型植物工場での栽培の容易さを利 用して、花托で組換えタンパク質 (インターフェロン) を作り、それを原薬として用いる製 薬技術が実用化されている12。 イチゴの花托には様々な香気成分が含まれているが、最もよく知られているものの 1 つ が4-hydroxy-2,5- dimethyl-3(2H)-furanone (HDMF) である (Fig. 2) 13,14。HDMF はパイナップ
ルやソバ、トマトなど多くの植物に含まれる芳香化合物であるが、揮発性が低い特徴から持 続性のある香りとして注目されている15–17。主に化学合成によって生産され、イチゴの香料
として食品や化粧品等に使用されている。
植物中においてHDMF は、アグリコンとしてだけでなく、様々な修飾を受けた形で存在 している。HDMF は、D-fructose-1,6-diphosphate を出発物質として前駆体である 4-hydroxy-5-methyl-2-methylene-3(2H)-furanone (HMMF) が生合成され、その後、FaQR/FaEO (Fragaria × ananassa quinone oxidoreductase/enone oxidoreductase) によって還元されることで生成する (Fig. 2) 18,19。生成したHDMF は、FaOMT (Fragaria × ananassa O-methyltransferase) によって
水酸基がメチル化された 2,5-dimethyl-4-methoxy-3(2H)-furanone (DMMF) や配糖体 (HDMF 4-O-glucoside)、配糖体がさらに修飾された malonyl 化体 (HDMF 4-O-glucoside malonate) と して存在している20,21。これらのうち、配糖体は成熟段階で必要に応じて糖が加水分解され
て放出されると考えられているが、HDMF を配糖化する過程の詳細は明らかになっていな い20。
本研究の開始時点では、Fragaria × ananassa glycosyltransferase (FaGT) として、イチゴよ りいくつかの配糖化酵素が単離され、これらは多くの天然・非天然の低分子有機化合物に対 する配糖化活性が報告されていた22–25。しかしながら、これらFaGT1~7 は、いずれも HDMF に対する配糖化活性がない、もしくは配糖化活性があっても、その活性が特異的ではなかっ た。また、本研究の途中で、HDMF 配糖化酵素として報告された UGT71K3 も、その基質特 異性は非常に低く、様々な化合物を配糖化することが明らかとなっていた26。 そこで本研究では、様々なデータベース情報を利用し、イチゴ花托中で発現している HDMF 特異的な配糖化酵素遺伝子を単離し、その配糖化活性と香気成分蓄積への寄与につ いて明らかにすることを目的とした。
第一節 データベース検索と候補遺伝子のクローニング
EST 情報とは材料となる植物において発現している mRNA 情報を集積したものであり、 目的の遺伝子配列を単離する上で非常に有用なリソースとなる。そこで、データベースとし て一般に公開されているイチゴの EST 情報から配糖化酵素遺伝子の配列情報を取得するこ ととした。最初に植物のEST データベースの 1 つである Plant GDB (http://www.plantgdb.org/) のデータから、Fragaria × ananassa EST (3035 クローン) を選択し、キーワード (UDP, glycosyl, glucosyl) を利用した配糖化酵素の候補遺伝子の網羅的探索を行った。その結果、157 クロー ンが見出され、それらの塩基配列を再度blastx 検索することで、既知の植物二次代謝糖転移 酵素と相同性のある28 クローンに絞り込んだ。この 28 クローンの機能を推定するため、 アミノ酸配列の相同性に基づいて構築されている既知の植物糖転移酵素の16 のグループの どこに属するかを決定した。その結果、10 クローンは低分子化合物のカルボキシル基を特 異的に配糖化することが知られているGroup L に属すると考えられた。そこで残った 18 ク ローンについて全長cDNA の単離を進めることとした。
イチゴの成熟花托からtotal RNA を抽出し、これをもとに cDNA を調製した。これを鋳型 としてEST データベースから得た配列情報をもとに作製した gene specific primer で、候補 遺伝子配列の増幅を行い、その配列を解析した。その結果、14 クローンについては花托で の発現が確認でき、その配列も EST 情報と同一のものであった。しかしながらそのうち 3 クローンは既知のFaGT3、FaGT4、FaGT6 の部分配列であった。また、残りの 4 クローンに ついては増幅が確認できなかったため、イチゴの花托では発現していないと考えられた。部 分配列をもとに5’-RACE、3’-RACE を進めていたところ、2013 年にイチゴの全ゲノム解析 が終了し、Fragaria × ananassa をはじめとしたイチゴのゲノムデータベース (Strawberry GARDEN (Genome And Resource Database ENtry)) が公開された27。また、ブドウのHDMF 配
糖化酵素としてUGT85K14 が報告されたため 28、データベースの検索システムを利用して
UGT85K14 と相同性の高いものを検索した。その結果、得られた配列情報も利用してこの 12 クローンの全長 cDNA 単離を進め、最終的に 6 クローンの全長 cDNA を単離することに 成功した。これらの 6 種のクローンを単離した順にそれぞれ FaGT8 (Fragaria × ananassa glycosyltransferase 8) ~13 とし、UGT ナンバーを取得した (Table 1) 。これらの得られた配列 についてアミノ酸配列を用いて比較を行った。その結果、FaGT1 および FaGT11 (UGT76A16)、 FaGT12 (UGT76A17) の間の相同性は 90%と高い値となった。また、その他のクローンは FaGT1~7 に対して、40~50%ほどの相同性を示した。
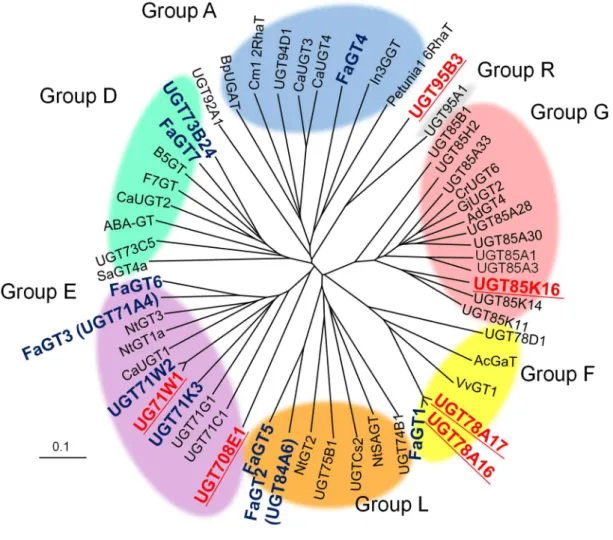
単離したFaGT8~13 の機能推定を行うことを目的として、既知の植物配糖化酵素のアミ ノ酸配列を用いて分子系統樹を作成した (Fig. 3) 。その結果、FaGT8 (UGT708E1) および FaGT10 (UGT71W1) は Group E、FaGT11/12 (UGT78A16/17) は Group F、そして FaGT13 (UGHT85K16) は Group G に属することがわかった。一方、FaGT9 (UGT95B3) は既知の配
7
糖化酵素グループにも分類することができなかった。
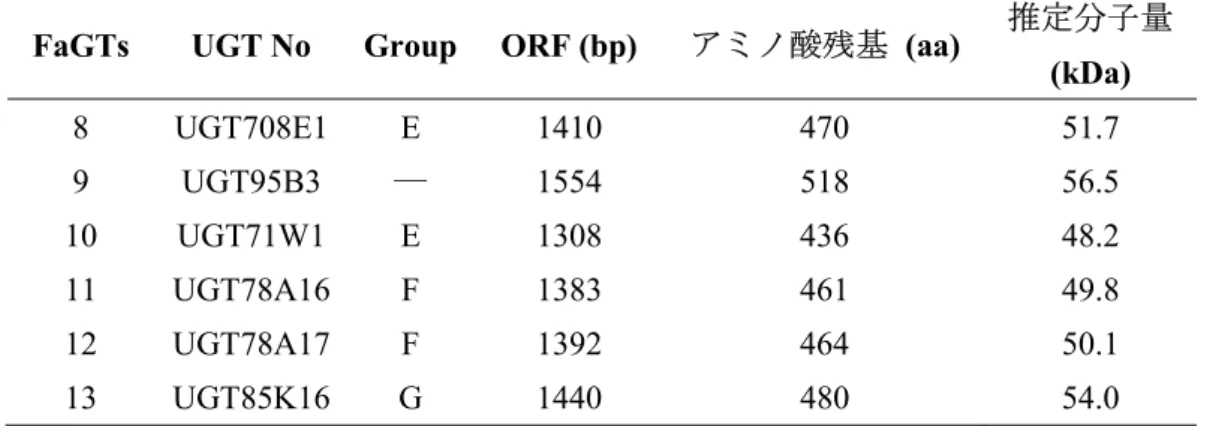
Table 1 イチゴ花托より得られた配糖化酵素遺伝子
FaGTs UGT No Group ORF (bp) アミノ酸残基 (aa) 推定分子量
(kDa) 8 UGT708E1 E 1410 470 51.7 9 UGT95B3 ― 1554 518 56.5 10 UGT71W1 E 1308 436 48.2 11 UGT78A16 F 1383 461 49.8 12 UGT78A17 F 1392 464 50.1 13 UGT85K16 G 1440 480 54.0
Fig. 3 イチゴ花托から単離した配糖化酵素および他の植物の PSPG による分子系統樹
Clustal W を用いて解析を行い、TreeViewで系統樹を作成した。各酵素の基原と accession number は実験の部に示した。本研究で単離した配糖化酵素は赤文字、過去にイチゴから単離された 配糖化酵素は青文字で示した。
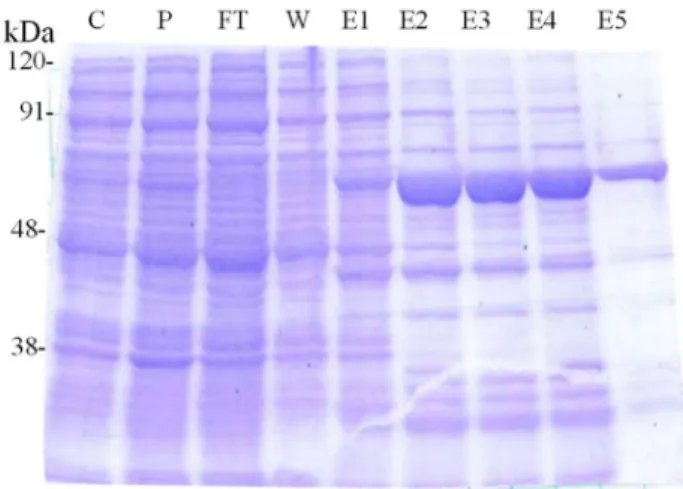
9 第二節 組換えタンパク質を用いた配糖化活性の解析 前節で得た6 分子種の ORF を大腸菌発現用ベクター pQE30 に組込み、N 末端での 6 × His-tag 融合組換えタンパク質として発現するようにした。これらを導入した組換え大腸菌 を培養し、IPTG 添加により組換えタンパク質の発現を誘導し、得られた菌体を超音波破砕 して得られた可溶性タンパク質とNi-NTA 担体で His タグによるアフィニティー精製をした タンパク質をSDS-PAGE し、CBB 染色によって組換えタンパク質の発現と精製を確認した (Fig. 4)。その結果、UGT708E1 および UGT78A16、UGT78A17、UGT85K16 のみ組換えタン パク質の発現を確認することができた。
組換えタンパク質の発現が確認できた4 分子種について、その精製酵素を用いて、UDP-glucose 存在下、HDMF に対する配糖化反応を行い、生成物を HPLC で分析した。その結果、 UGT85K16 のみ HDMF 配糖化活性を検出した (Fig. 5)。この生成反応は UDP-glucose 依存的 であり、配糖化反応の経時変化を解析したところ、経時的にピークが増大し、反応生成物の 構造解析をNMR および ESI-TOF MS で行った結果、HDMF 4-O-glucoside であることがわか った21。また、反応時間と面積値をプロットしたところ、初速は2 分までと考えられた。 HDMF 配糖化活性を示した UGT85K16 の基質特異性を検討するとともに、UGT708E1 お よびUGT78A16、UGT78A17 の配糖化活性についても検討することとした。FaGT1~7 によ る配糖化活性について報告のあった天然由来化合物のうち、cyanidin、p-coumaric acid、 cinnamic acid を選択し、さらに HDMF と類似した成分である EHMF と HMF、イチゴに含ま れる主要な香気成分であるvanillin をアクセプター基質として用い、配糖化活性を検討した。 その結果、UGT85K16 は HDMF 以外に EHMF および HMF、vanillin にも配糖化活性を示し たが、その他の基質には配糖化活性を示さなかった。また、UGT78A16 および UGT78A17 は、今回検討した基質の中では、cyanidin にのみ配糖化活性を示し、他の基質については配 糖化活性を示さなかった (Fig. 6)。また、UGT708E1 についてはいずれの基質に対しても配 糖化活性を示さなかった。
続いて、UGT85K16 の酵素反応速度論パラメーターを算出した結果、HDMF に対する Km
値、kcat値、kcat/Km値はそれぞれ0.32 mM、1.1 sec-1、3.7×103 M-1sec-1、vanillin に対する Km値、
kcat値、kcat/Km値はそれぞれ1.5 mM、0.11 sec-1、0.067×103 M-1sec-1だった (Table 2)。これら
の結果から、UGT85K16 の HDMF に対する触媒効率は vanillin に対する触媒効率よりも 55 倍高く、HDMF に対する基質特異性の方が高いことが示された。
Fig. 4 大腸菌に発現させた UGT85K16 の SDS-PAGE による精製 精製段階のタンパク質10 µg をアプライし CBB 染色を行った。 Elution 2 (E2) を酵素活性の検討に用いた。
Lane C: Crude enzyme Lane : Pellet Lane F: Flow through Lane W: Wash Lane E1-E5: Elution
13 第三節 遺伝子発現とHDMF 配糖体の蓄積
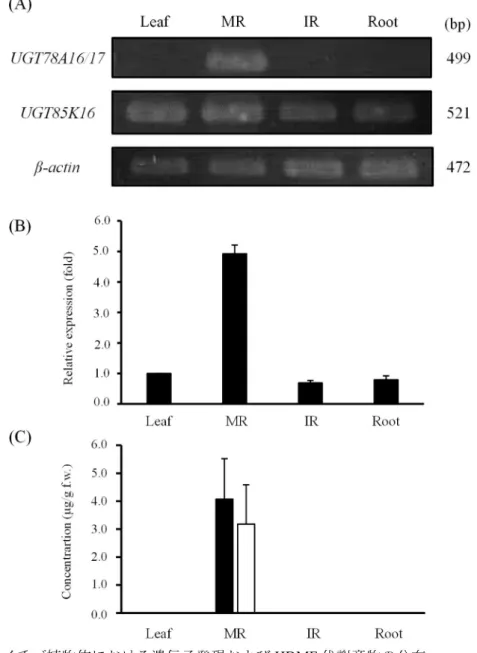
配糖化活性を検出した UGT85K16 および UGT78A16/17 の植物体における機能を推定す るため、遺伝子発現を半定量 RT-PCR によって解析した。その結果、葉および未成熟花托 (IR)、根と比較して、成熟花托 (MR) でより高い発現を示した (Fig. 7A, B)。同時に各器官 のMeOH 抽出液を調製し、HDMF と HDMF 配糖体の蓄積を HPLC 分析によって定量したと ころ、成熟花托にHDMF と HDMF 配糖体がより多く蓄積していた (Fig. 7C)。このことか ら、UGT85K16 の発現と HDMF 配糖体の蓄積には相関があると考えられた。
Fig. 7 イチゴ植物体における遺伝子発現および HDMF 代謝産物の分布
イチゴ植物体の各器官から total RNA を抽出し、semi-quantitative RT-PCR を行っ た。また、70%MeOH 抽出液を調製し、HPLC で分析した。(A) UGT85K16 および UGT78A16/17、β-actin の PCR 増幅産物の電気泳動写真。(B) UGT85K16 の相対発 現量。(C) HDMF (■) および HDMF 配糖体 (□) の蓄積。データは、平均±標準 誤差 (n = 3) で示した。MR, mature receptacle: IR, immature receptacle: f.w., fresh weight.
第一章 考察 本章では、イチゴのHDMF 配糖化酵素を同定するため、イチゴ花托中に発現する新規配 糖化酵素遺伝子の網羅的クローニングを試みた。データベース検索によるクローニング方 法は、homology-based PCR クローニングと比較して、発現量に依存せず、より多くのデータ が得られる上に、キーワードを利用した簡便な検索方法も利用できることから、効率よく、 目的遺伝子の部分または全体の配列情報を得られ、クローニングも容易となる。本研究では 当初Plant GDB の EST データベースから 28 種しか配糖化酵素遺伝子を見出せなかったが、 一般的にPSPG は一つの植物に 100 種程度はあると推定されている3。本研究の途中で公開 されたイチゴのゲノムデータベースStrawberry GARDEN を利用すれば、さらに多くの候補 遺伝子を見出せたと考えられる。しかしながら、最終的にHDMF 配糖化酵素として見出し たクローンはゲノムデータベース由来であり、EST データベース由来の配糖化酵素には HDMF 配糖化活性はなかった。このことから、EST データがどのような植物材料を利用し て構築されたかが重要であり、目的の生合成酵素遺伝子の効率の良いクローニングには、植 物材料の状態、可能であれば代謝産物のデータを考慮した遺伝子情報の利用が必要である。 植物は、様々な香気成分として知られる揮発性化合物を生合成し、その一部を配糖体とし て蓄積している。これら香気成分の配糖体は、水溶性が高くなり香りが低くなる性質を持つ ことが多く、香気成分の配糖化と脱配糖化の制御は、植物の香りの制御に重要な役割を担っ ていると考えられている。イチゴにおける重要な香気成分であるHDMF の配糖化について は、これまでも配糖化酵素の単離が試みられてきた。本研究より先にUGT73K3a/b が HDMF 配糖化酵素として報告されたが、UGT73K3a/b は基質特異性が低く、本研究で解析した UGT85K16 と比較して vanillin に対する活性が高い26。また、phloroglucinol をはじめとした
他の多くのフェノール性化合物も配糖化する29。さらに、イチゴの一過的発現系を利用した UGT73K3a/b の発現抑制でも HDMF 配糖体や malonyl 化配糖体含量の低下は大きくないと 報告されている。これらのことから、HDMF 配糖化に対する UGT71K3a/b の関与は部分的 で、他にHDMF 配糖化酵素が存在することが考えられ、本研究で解析した UGT85K16 は酵 素活性や発現部位の点から HDMF 配糖化酵素の有力な候補であると考えられる。今後、 UGT71K3a/b のように、一過的発現系を利用した解析を行うことで、UGT85K16 の HDMF 配 糖化とHDMF 放出制御との関係を明らかにできると考えられる。HDMF の放出制御機構に おける配糖化酵素の役割が明らかになれば、香気成分をターゲットとしたイチゴの育種の 可能性も広がると期待できる。 UGT85K16 のアミノ酸配列は、単離の際に配列情報を参考にしたブドウの HDMF 配糖化 酵素であるUGT85K14 のアミノ酸配列と 68.8%の相同性がある 28。UGT85K14 は、HDMF に対して特異性が高く、linalool のようなモノテルペンアルコールなどの揮発性化合物は配 糖化しない。また、ブドウにおいてUGT85K14 の遺伝子発現と HDMF 配糖体の蓄積と相関 があることが明らかとなっている。これらのことから、UGT85K14 と UGT85K16 はオーソ
15
ログの関係にあると考えられる。ブドウ以外の植物おけるUGT85K サブファミリーの機能 は、植物の香気成分の生合成および蓄積と関わりが深いと考えられている。チャノキの UGT85K11 は geraniol や eugenol、(Z)-3-hexenol を含む広い範囲の揮発性化合物を配糖化す るが、HDMF に対する配糖化活性は明らかになっていない30。UGT85K16 および UGT85K14、 UGT85K11 のアミノ酸配列を比較すると、UGT85K11 のみで違いがあるアミノ酸残基があ ることから、タンパク質の三次元構造のホモロジーモデリングを基にした点変異酵素など を作製し解析することで、HDMF に対する UGT85K16 や UGT85K14 の特異性を担うアミノ 酸残基が明らかになることが期待される (Fig. 8)。さらに、UGT85A サブファミリーではあ るが、geraniol や citronellol などのモノテルペンアルコールを配糖化するシロイヌナズナの UGT85A3 やブドウの UGT85A28、A30、A33 も配列比較の対象に加えることで、揮発性化 合物を認識する配糖化酵素の分子メカニズムを解明できると考えられる30。
Fig. 8 UGT85 サブファミリーのマルチプルアライメント
17 第二章 モミジバダイオウのアントラキノン配糖化酵素の機能解析 モミジバダイオウ (Rheum palmatum) は生薬ダイオウの基原植物であり、その根茎は瀉下 薬として大黄甘草湯に、また活血化瘀薬として桃核承気湯や大黄牡丹皮湯に配合されるな ど、ヒトにとって有用な植物である。同属植物であるカラダイオウ (Rheum rhabarbatum) は 食用ダイオウとしても知られ、葉柄部分が食用とされている。モミジバダイオウは、瀉下成 分としてsennnoside A をはじめとしたアントラキノン類を含有し、他にも rhatannin をはじ めとしたタンニン類が含まれている31。 アントラキノン類はキノン骨格を持つ天然物の中で大きなグループを占めており、高等 植物や菌類、地衣類などに含まれている32。アントラキノン類はアセチルCoA およびマロ ニルCoA を出発物質とし、酢酸マロン酸経路を経て基本骨格が形成され、その後、水酸化、 メチル化、酸化、配糖化などの修飾反応を経て生合成される (Fig. 9)。これらは植物体では アグリコンや配糖体として蓄積しているが、アントラキノン類を蓄積する植物において、そ れぞれの変換反応を触媒する詳細な酵素の単離報告はほとんどない。また、ダイオウ属植物 では多くのアントラキノン類は配糖体としても蓄積されていることが知られている 31が、 モミジバダイオウにおいてもアントラキノン生合成酵素遺伝子のほとんどが未知である。 これらのことをふまえ、本研究では、アントラキノン生合成に関与する特異的配糖化酵素を 同定することを目的とした。 Fig. 9 アントラキノン類の推定生合成経路
第一節 モミジバダイオウからの配糖化酵素遺伝子の網羅的クローニング モミジバダイオウは、第一章で研究対象としたイチゴとは異なり広く栽培されている植 物ではないため、配列情報のデータベースが整備されていなかった。そこで、アントラキノ ン配糖化酵素を探索するにあたり、本研究ではhomology-based PCR 法を用いて網羅的なク ローニングを試みた。 PSPG には高度に保存された 44 アミノ酸残基からなる PSPG-box と呼ばれる領域が存在 する3。この領域のアミノ酸配列をもとに縮重プライマーを設計し、モミジバダイオウの根
茎から調製したtotal RNA を逆転写して得た cDNA を鋳型として、homology-based PCR によ り3’末端の増幅を行った。得られた 3’末端側断片を元に、残りの 5’末端側を 5’RACE 法に より単離した。その結果、モミジバダイオウから配糖化酵素RpUGT (Rheum palmatum UDP-glycosyltransferase) 1~3 を単離した。これら 3 分子種の UGT ナンバーを取得し、分子系統 学的解析を行った結果、RpUGT1 が Group D、RpUGT2 および 3 は Group E に属しているこ とがわかった (Table 3, Fig. 10, 11) 。Group D と Group E はともに様々な基質を認識する配 糖化酵素が報告されていることから、RpUGT1~3 のどれがアントラキノン配糖化能を持つ か推定することは困難だった。
Table 3 モミジバダイオウより得られた配糖化酵素遺伝子
RpUGTs UGT No Group ORF (bp) アミノ酸残基 (aa) 推定分子量 (kDa)
1 UGT73BE4 D 1428 476 52.8
2 UGT72B49 E 1431 477 51.6
Fig. 11 単離したアントラキノン配糖化酵素および他の植物の分子系統樹
Clustal W を用いて解析を行い、TreeView で系統樹を作成した。各酵素の基原 とaccssion number は実験方法に示した。今回単離した配糖化酵素は赤文字示し た。
続いて、Rheum 属植物に含まれることが報告されているアントラキノン配糖体やスチル ベン配糖体のアグリコン31、また、RpUGT1 と相同性の高い UGT73 ファミリーの基質であ
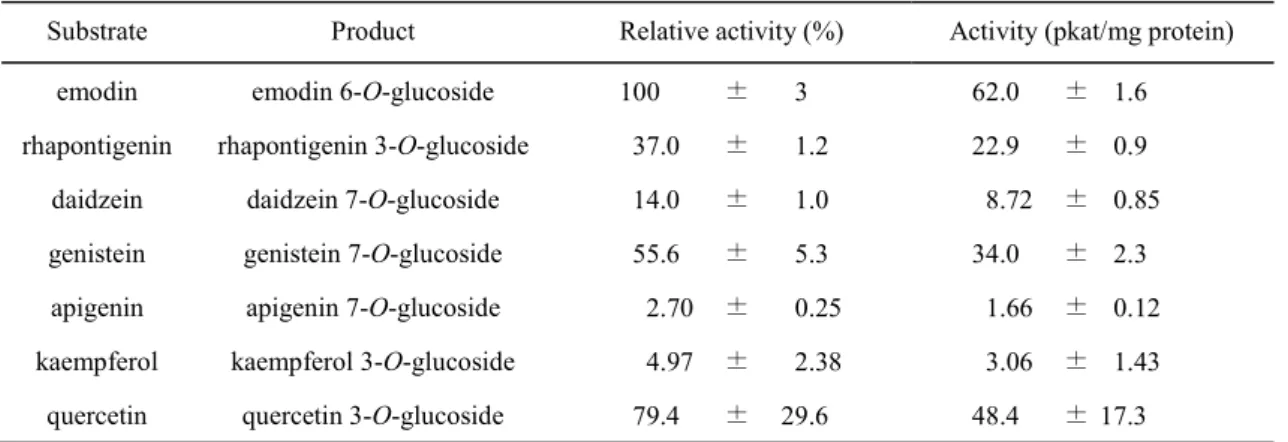
るフラボノイド類を用いて RpUGT1 の基質特異性について検討を行った。すべて配糖化反 応後にHPLC で分析し、配糖体の標品と比較した (Fig. 13)。その結果、RpUGT1 はアントラ キノン類ではemodin、スチルベン類では rhapontigenin、フラボノイド類では daidzein、genistein、 apigenin、kaempferol、quercetin に対して配糖化活性を示した (Fig. 14)。RpUGT1 は、 rhapontigenin に対しては 3 位のみを配糖化して rhaponticin を生成したが、フラボノイド類で は複数の水酸基に対して配糖化活性を示した。RpUGT1 が配糖化活性を示した基質に対す る活性を比較した結果、emodin に対する活性を 100%としたとき、rhapontigenin に対する配 糖化活性は約40%だった (Table 4)。またフラボノイド類では quercetin に対する配糖化活性 が最も高く、約 80%だったが、それ以外の基質については、60%以下の活性だった (Table 4)。 そこで、反応生成物が単一で比較的活性の高い化合物を選択して、RpUGT1 の酵素化学的 解析を行った。反応時間4 分で、基質濃度を 0.05 mM~1 mM として親和性を検討したとこ ろ、emodin に対する基質親和性は約 0.22 mM で、rhapontigenin に対する基質親和性とほぼ 同等だった (Table 5)。また、emodin に対する kcat値は、rhapontigenin に対する kcat値の約3
倍だった。これらの結果から触媒効率を示すkcat/Km値で比較すると、emodin に対する触媒
23 Fig. 13 RpUGT1 とによる反応生成物の代表的なクロマトグラム 代表的な生成物のピークを黒、基質のピークを白でマークした。 反応条件、HPLC 条件は、実験の部に示したとおりである。 -0.001 0.000 0.001 0.002 0.003 0.004 0.005 5 10 15 20 25 -0.0020.000 0.002 0.004 0.006 0.008 0.010 0.012 0.014 5 10 15 20 25 rhapontigenin 3-glucoside rhapontigenin rhapontigenin -0.001 0.000 0.001 0.002 0.003 0.004 0.005 5 10 15 20 25 daidzein 7-glucoside daidzein -0.001 0.000 0.001 0.002 0.003 0.004 0.005 5 10 15 20 25 genistein 7-glucoside genistein -0.0020.000 0.002 0.004 0.006 0.008 0.010 0.012 0.014 5 10 15 20 25 apigenin 7-glucoside apigenin -0.001 0.000 0.001 0.002 0.003 0.004 0.005 5 10 15 20 25 kaempferol 3-glucoside kaempferol quercetin 3-glucoside quercetin
Retention time (min)
A bs or ba nc e at 3 00 n m (A U ) A bs or ba nc e at 3 40 n m ( A U ) daidzein genistein apigenin kaempferol quercetin
Fig. 14 RpUGT1 の基質特異性の検討で使用した化合物の構造式 RpUGT1 は、実線で囲んで示した化合物を配糖化した。
25 Table 4 RpUGT1 の基質特異性
データは、平均±標準誤差 (n = 3) で示した。
Table 5 RpUGT1 の酵素化学的パラメーター
Substrate Km (mM) kcat (×10-3 s-1) kcat/Km (M-1 s-1)
Acceptor substrate (vs. UDP-glucose)
emodin 0.22 ± 0.01 1.9 ± 0.1 8.5 ± 0.7 rhapontigenin 0.32 ± 0.01 0.6 ± 0.04 1.9 ± 0.04
Donor substrate (vs. emodin)
UDP-glucose 0.63 ± 0.17 3.1 ± 0.3 4.9 ± 0.6
データは、平均±標準誤差 (n = 3) で示した。
Substrate Product Relative activity (%) Activity (pkat/mg protein)
emodin emodin 6-O-glucoside 100 ± 3 62.0 ± 1.6
rhapontigenin rhapontigenin 3-O-glucoside 37.0 ± 1.2 22.9 ± 0.9
daidzein daidzein 7-O-glucoside 14.0 ± 1.0 8.72 ± 0.85
genistein genistein 7-O-glucoside 55.6 ± 5.3 34.0 ± 2.3
apigenin apigenin 7-O-glucoside 2.70 ± 0.25 1.66 ± 0.12
kaempferol kaempferol 3-O-glucoside 4.97 ± 2.38 3.06 ± 1.43
第三節 遺伝子発現解析と代謝産物の蓄積
モミジバダイオウ植物体でのRpUGT1 の機能を推定するために、根、根茎、葉柄、葉、実 生を用いて、qRT-PCR 法により RpUGT1 の遺伝子発現の解析を行った。その結果、RpUGT1 は根や根茎と比較して、葉柄や葉、実生など地上部で高い発現量を示した (Fig. 15A) 。同 時に、各器官の70% MeOH 抽出液を調製し、RpUGT1 の基質や生成物である emodin 類や rhapontigenin、rhaponticin の蓄積を HPLC 分析により定量した。その結果、emodin および emodin 8-glucoside は植物体全体に分布しており、emodin 1-O-glucoside および emodin 6-O-glucoside はほとんどが地下部に蓄積していた (Fig. 15B)。また、rhapontigenin および rhaponticin も植物体全体に分布していたが、地下部においてより多く蓄積していた (Fig. 15C) 。これらの結果から、RpUGT1 の発現と emodin 6-O-glucoside の蓄積には直接の相関関 係は見いだされなかった。
第二章 考察 本章では、アントラキノン配糖化酵素を同定するため、モミジバダイオウからの網羅的な 配糖化酵素遺伝子の単離を試みた。第一章と異なりデータベース情報もなかったことから、 homology-based PCR クローニングを行った。その結果、今回は 3 分子種の単離に留まった。 PSPG は一つの植物に 100 種程度存在することが推定されていることから、根茎という植物 一般に存在する器官ではない部分を材料にしたとはいえ、単離できた数としては少ないと 考えられた。今後、RNA-seq などを活用すれば、より多くの配列情報を得ることができると 考えている。 単離した3 分子種の組換え酵素を用いた解析から、RpUGT1 が emodin にのみ配糖化活性 を示し、emodin の 6 位を配糖化した。ただ、アントラキノン配糖体の単離や含量の報告か ら、emodin 型のアントラキノンの多くは 8 位の水酸基が配糖化された配糖体として蓄積し ている 31。したがって、RpUGT1 とは別の 8 位の水酸基を配糖化する配糖化酵素が存在し ていると考えられる。これは、植物体における遺伝子発現と代謝産物の解析の結果でも、 RpUGT1 の発現が比較的高い葉柄において、emodin および emodin 8-O-glucoside の蓄積があ り、他の配糖体があまり蓄積していなかったことからも十分推定される。Emodin 8-O-glucoside を生成する別の配糖化酵素が emodin に対して RpUGT1 のより高い emodin への親 和性を持つために最終的にemodin 8-O-glucoside が蓄積することになっていると想定される。 ただ、emodin や emodin 配糖体をはじめとして、アントラキノン配糖体が生合成される器官 と蓄積される器官の関係は全くわかっておらず、アントラキノンやアントラキノン配糖体 の器官間での転流も考慮する必要はある。そのためにも、まずin vitro で emodin 8-O-glucoside を生成する特異性の高い配糖化酵素を単離し、その遺伝子発現を明らかにする必要があり、 クローニングを行う際の遺伝子源としてモミジバダイオウの葉柄は適切な材料となる可能 性がある。また、モミジバダイオウ植物体におけるemodin 配糖体の蓄積に関して、その分 布はほとんど解析されておらず、本研究でemodin 1-O-glucoside や emodin 6-O-gluocside が 含有されていることを初めて明らかにした。これは、emodin 1-O-glucoside を生成する配糖 化酵素を単離する際の貴重な代謝物情報となると考えられる。一方、rhapontigenin の配糖化 についても、RpUGT1 の発現と相関していないことから、こちらも別の配糖化酵素の可能性 を考える必要がある。
本研究で単離したRpUGT1 について分子系統学的解析を行うと、RpUGT1 は Group D に 分類された (Fig. 11)。また、UGT ナンバーで見ても、RpUGT1 が属する UGT73 ファミリー はフラボノイド類などを配糖化する PSPG が数多く報告されている 34。このことから、
RpUGT1 が様々なフラボノイドに対して配糖化活性を示したと考えられる。またフラボノ イド類の部分構造が、emodin と似通っていることから、RpUGT1 の配糖化活性の emodin の 6 位への位置選択性は、分子系統学的に比較的近い分子種である F7GT のようなフラボノイ ドの7 位への配糖化活性から機能分化したものと考えられる35。ただし、イソフラボノイド
29 の7 位に特異的な配糖化酵素は別に報告されており、UGT88 サブファミリーに属している 36。このことから、emodin 配糖化活性への機能分化の分子メカニズムを明らかにするには、 点変異酵素などを利用した酵素の構造活性相関に着目した解析が必要である。酵素化学的 な視点からは、RpUGT1 がスチルベン類の中で、rhapontigenin に配糖化活性を示したにも関 わらず、resveratrol や piceatannol に対して配糖化活性を示さなかったのは非常に興味深く、 スチルベン類が構造活性相関の解析の対象として有用であると考えられる。
【結論】 本研究では、イチゴのHDMF 配糖化酵素とモミジバダイオウのアントラキノン配糖化酵 素の単離と機能解析を試みた。本研究でイチゴから単離、機能解析したHDMF 配糖化酵素 UGT85K16 については、これまで報告されていた HDMF 配糖化酵素と比較して HDMF 配糖 体の生合成への関与が高いと考えられる。今後、イチゴにおける香気成分の人為的な改変の ツールとして利用できることが期待できる。 一方、モミジバダイオウからはアントラキノン配糖化酵素としてemodin 6-O-glucoside を 生成する配糖化酵素を単離した。今まで、アントラキノンに対する配糖化活性を検討した報 告はないことから、今後より特異的なアントラキノン配糖化酵素を単離する上で重要な情 報となる。 植物の二次代謝産物の配糖体の生合成やそれに関わるPSPG の単離については、近年多く の報告があるが、PSPG の分子進化や基質認識のメカニズムは未解明な課題となっている。 本研究が、それらの解明につながる基礎的な知見の一つとなることを期待したい。
31 実験方法 【植物材料】 イチゴ(Fragaria ananassa)の未成熟花托及び葉は、本学薬用植物園で栽培した個体を 用いた。成熟花托は名古屋市内のスーパーマーケットで 2013 年に購入したものを用いた。 モミジバダイオウ (Rheum palmatum) の植物体は、医薬基盤研究所薬用植物資源研究セン ター (名寄) から分与していただいた種子を用いて育種した。種子を次亜塩素酸ナトリウム 水溶液で表面殺菌した後、LS (Murashige and Skoog Basal Salts with minimal organics, Sigma Aldrich, St. Louis, MO, USA) ゲランガム培地に播種した。2 日間 4℃で春化処理した後、25℃ で静置した。発芽後、1 ヶ月程度の実生を、ポッド内の土 (コンパル, 住友林業緑化, 東京) に植え替え、20〜25℃、14 時間明期、10 時間暗期の光周期で、育苗器内 (植物育成ラック プラントマスターP100, バイオメディカルサイエンス, 東京) で生育維持した。
【試薬】
HDMF、EHMF、HMF、p-coumaric acid、emodin は Sigma-Aldrich (St. Louis, MO, USA) から 購入した。UDP-glucose、vanillin、resveratrol、daidzein は富士フイルム和光純薬 (大阪) から 購入した。Cyanidin chloride は EXTRASYNTHESE (Lyon, France) から購入した。Vanillin 4-O-β-D-glucoside、emodin 1-O-glucoside、emodin 8-O-glucoside は Toronto Research Chemicals, Inc. (Downsview, Canada) から購入した。Chrysophanol、piceatannol、rhapontigenin、genistein、 apigenin は東京化成工業 (東京) から購入した。Kaempferol は Santa Cruz Biotechnology, Inc. (Dallas, TX, USA) から購入した。 Quercetin dihydrate は AdipoGen Life Sciences (San Diego, CA, USA) から購入した。
【イチゴからの配糖化酵素遺伝子のPCR cloning】
Fruit-mait for RNA purification (タカラバイオ, 草津) および Plant total RNA Mini (VIOGENE, Taipei, Taiwan) を用いて、イチゴ花托から抽出した total RNA を鋳型として、Cap FishingTM
Full-length cDNA Premix Kit(Seegene, Rockville, MD, USA)または、SMARTer® RACE 5’/3’ Kit
(タカラバイオ) により、添付のプロトコールに従って cDNA を合成した。
イチゴ花托由来のcDNA を鋳型として、全 ORF を含むように設計した primer (Table 6) を 用いてPCR 反応を行い、目的サイズの PCR 産物のゲルを切り出した後、pMD20 T-Vector (タ カ ラ バ イ オ) に サ ブ ク ロ ー ニ ン グ し た 。 挿 入 し た PCR 産 物 の 配 列 の 確 認 に は BigDye®Terminator v3.1 Cycle Sequencing kit (Thermo Fisher Scientific, Waltham, MA, USA) と
vector に対応する primer (Table 6) を用い、添付のプロトコールに従ってサイクルシークエ ンス反応を行った後、ABI 3130 Genetic Analyzer (Thermo Fisher Scientific) でプロトコールに 従って解析し、塩基配列を決定した。
Table 6 イチゴからのクローニングに用いたプライマー
Primer (name) Sequence (5' to 3') 備考
UGT708E1 fw-full ATGTCAAACTCTCATGCACAGCC 全長ORF
rv-full CTAGTTCCTCTTCTGCATCAAAGAC 全長ORF
Fw-KpnI GCGGTACCATGTCAAACTCTCATGCACAGCC pQE30 vector への導入用
Rv-PstI GCCTGCAGCTAGTTCCTCTTCTGCATCAAAGAC pQE30 vector への導入用
UGT95B3 fw-full ATGGCAAATGAGATATGGATCGTTCCG 全長ORF
rv-full TTAGTTTTTATGTCTAAGGAAGTCTCCAAAGGC 全長ORF
Fw-PstI GCGCATGCATGGCAAATGAGATATGGATCGTTCCG pQE30 vector への導入用 Rv-HindIII GCCTGCAGTTAGTTTTTATGTCTAAGGAAGTCTCCAAAGGC pQE30 vector への導入用
UGT71W1 fw-full ATGAAATCACCATTTGGGGTCGC 全長ORF
rv-full TCAGTTATTCTCAAGGTTTTGTGACAG 全長ORF
fw- KpnI GCGGTACCATGAAATCACCATTTGGGGTCGC pQE30 vector への導入用
rv- SphI GCGCATGCTCAGTTATTCTCAAGGTTTTGTGACAG pQE30 vector への導入用 Table6 続き
UGT78A16 fw-full ATGGCACCAGTAGGTCATGT 全長ORF
rv-full TTAATTGGTTGTAGTCATTTCCAACAATG 全長ORF
fw-BamHI GCGGATCCATGGCACCAGTAGGTCATGT pQE30 vector への導入用
rv-PstI GCAAGCTTTTAATTGGTTGTAGTCATTTCCAACAATG pQE30 vector への導入用
UGT78A17 fw-full ATGGCATCAAACCAGGCGGG 全長ORF
rv-full TTAATTGGTTGTAGCCATTTCCAACAATG 全長ORF
fw-BamHI GCGGATCCATGGCATCAAACCAGGCGGG pQE30 vector への導入用
rv-PstI GCAAGCTTTTAATTGGTTGTAGCCATTTCCAACAATG pQE30 vector への導入用
UGT85K16 fw-full ATGGGTTCAGCACAATTAGGAGCC 全長ORF
rv-full TTATTGCAGACACTGCTCAATCAATC 全長ORF
Fw-KpnI GCATGCATGGGTTCAGCACAATTAGGAGCC pQE30 vector への導入用
Rv-BamHI CCCGGGTTATTGCAGACACTGCTCAATCAATC pQE30 vector への導入用
【モミジバダイオウからの配糖化酵素遺伝子のPCR cloning】
Plant Total RNA Extraction System (VIOGENE) または innu PREP plant RNA kit (Analytik Jena, Thuringia, Germany)を用いて、モミジバダイオウの根茎から抽出した total RNA を鋳型とし て、SuperScriptTM III Reverse Transcriptase (Thermo Fisher Scientific) または SMARTer® RACE
5’/3’ Kit (タカラバイオ) により cDNA を合成した。
33
(Promega, Madison, WI, USA) を用いて、homology-based PCR により 3’末端側断片を得た37。
このPCR 断片をサブクローニングした後、塩基配列を決定した。決定した塩基配列を元に primer (Table 7) を設計し、GoTaq® Green Master Mix (Promega) または KOD FX-Neo (東洋紡, 大阪) を用いて、5’末端側配列を 5’ RACE 法により得て、同様に塩基配列を決定した。得ら れた5’および 3’末端断片の塩基配列から primer (Table 7) を設計し、KOD -Plus- Neo (東洋 紡) を用いて全長 cDNA を増幅し、塩基配列を決定した。
Table 7 モミジバダイオウからのクローニングに用いたプライマー
【大腸菌発現系を用いた配糖化酵素遺伝子の発現と組換えタンパク質の精製】
全長cDNA が組み込まれたプラスミドを鋳型にして、ORF の両端に制限酵素サイトを付 加した primer (Table 6, 7) を設計し、PCR を行った後、pQE30 ベクター (Qiagen, Hilden,
Primer (name) Sequence (5' to 3') 備考
UGT2mFw TTYBTIWSICAYTGYGGITGGAA Degenerate primer for 3'-RACE
PSPG2Fw TGYGGITGGAAYTCIRYIYTIGA
3'-RACE primer CTGTGAATGCTGCGACTACGAT
10×Universal Primer Mix TAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT
RpUGT1 No 6Rv1 TGCATCCAACATTTGAGTCAACCCA 5'-RACE (1st PCR)
No 6Rv2 CACCAACCAATCGCAGTTCTGATCC 5'-RACE (nested PCR)
Table 7 続き
RpUGT1Fw ATGGCTTCTCATAACCATCTCCACATG 全長ORF
RpUGT1Rv ACAGCCATGCATGACCACATTTAGCAC 全長ORF
pESUMO_Bam_Fw CAGCAGACGGGAGGTGGATCCATGGCTTCTCATAACCATCTCCACA pE SUMO pro3 vector への導入用 pESUMO_Rv ATTCGGATCCTCTAGATTAGCACTTTATAGAAGTCAATGCATCCAAC pE SUMO pro4 vector への導入用
RpUGT2 No 2Rv1 AGCCTTAGTAGAGGAGCCGTCTTCACT 5'-RACE (1st PCR)
No 2Rv2 ATTCCTCCATCCGAGTCCTGATCCGCTT 5'-RACE (nested PCR)
RpUGT2Fw ATGGAATCTTCACCCGCCGCCG 全長ORF
RpUGT2Rv ACTAAACCTTGCTCTCCCAT 全長ORF
pESUMO_Bam_Fw CAGCAGACGGGAGGTGGATCCATGGAATCTTCACCCGCCG pE SUMO pro3 vector への導入用 pESUMO_Rv ATTCGGATCCTCTAGACTAAACCTTGCTCTCCCATTTTCTTGC pE SUMO pro4 vector への導入用
RpUGT3 No 4Rv1 GAAGAACCACCAACTTCCAAAGCCTTG 5'-RACE (1st PCR)
No 4Rv2 CACGTCGTACCTATAATCCATCCTCAGC 5'-RACE (nested PCR)
RpUGT3Fw TATGGAAGACGGAGAGCTGGTGTTCATC 全長ORF
RpUGT3Rv CTACCAAATATAGTAAACTAGTTCATGG 全長ORF
pESUMO_Bam_Fw CAGCAGACGGGAGGTGGATCCATGGGTCATCTGGTGGCG pE SUMO pro3 vector への導入用 pESUMO_Rv ATTCGGATCCTCTAGACTAGTTCATGGATAAATTCTTAATAAGATCTTGAGT pE SUMO pro4 vector への導入用
Germany)または pE-SUMO pro3 (LifeSensors, Malvern, PA, USA) ベクターにサブクローニン グした。完成した発現用プラスミドを発現用大腸菌NiCo21 株 (New England Biolabs, Bevery, MA, USA) に導入した。これを 37℃で液体 LB 培地中で培養し、OD600が0.4~0.8 に達した
ところで終濃度1 mM または 5 mM となるように IPTG を添加し、18℃で 20~22 hr または 30℃で 6 hr 培養してタンパク質の発現を誘導した。
組換えタンパク質の精製は、QIA expression Type Ⅳ kit (Qiagen) を用いて、添付のプロト コールに従って精製を行った。精製後は SDS-PAGE によってタンパクの発現の確認し、濃 度はBradford 法により定量した38。
35 【組換え酵素を用いた配糖化反応】 配糖化活性の測定は、50 mM Tris-HCl (pH 7.5) または 50 mM glycylglycine (pH 8.4) に、1 mM のアクセプター基質、5 mM の UDP-glucose、15 µg 精製酵素を加えて行った。酵素反応 は30°C で一定時間インキュベートした後、50 µl の酵素反応液に 100 µl の MeOH を加えて 反応を止めた後、13,800 × g で 10 分間遠心分離し、上清を以下の HPLC 条件で分析した。 <共通するHPLC condition>
固定相:Cosmosil 5C18-ARⅡ column 4.6 × 1.5 mm (ナカライテスク, 京都)。移動相:(A) 0.1%
formic acid, (B) CH3CN。流速:1.0 ml/min。カラム温度:25°C。
<for HDMF enzyme assay>
移動相:0–10 min, 5–10% (B); 10–12 min, 10–100% (B); 12–16 min, 100% (B); 16–18.5 min, 100– 5% (B); 18.5–25 min, 5% (B)。測定波長:290 nm。溶出時間:HDMF, 7.8 min; HDMF-glc, 5.9 min。
<for cyanidin>
移動相:0–10 min, 10% (B); 10–20 min, 10–50% (B); 20–25 min, 50–100% (B); 25–28 min, 。100% (B); 28–30 min, 100–10% (B); 30–35 min, 10% (B)。測定波長:530 nm。Cyanidin, 16.3 min; cyanidin-glc, 2.8 min。
<for p-coumaric acid>
移動相:0–20 min, 10–20% (B); 20–25 min, 20–100% (B); 25–27.5 min, 100% (B); 27.5–30 min, 100–10% (B); 30–35 min, 10% (B) 。測定波長:530 nm。溶出時間:p-coumaric acid, 14.1 min。 <for cinnamic acid>
移動相:0–10 min, 10% (B); 10–20 min, 10-50% (B); 20–25 min, 50–100% (B); 25–27 min, 100% (B); 27–30 min, 100–10% (B); 30–35 min, 10% (B)。測定波長:530 nm。溶出時間:p-coumaric acid, 21.4 min。
<for anthraquinone glucoside>
移動相:0–20 min, 20–40% (B); 20–25 min, 40-100% (B); 25–27.5 min, 100% (B); 27.5–-30 min, 100–20% (B) ; 30–35 min, 20% (B)。測定波長:440 nm。溶出時間:emodin, 26.7 min; E1G, 8.9 min E6G, 17.4 min; E8G, 13.7 min。
<for anthraquinone aglycone>
移動相:0–22 min, 40–100% (B); 22–27 min, 100% (B)。測定波長:440 nm。溶出時間:emodin, 11.9 min。
<for flavonoids>
移動相:0-17 min, 10–50% (B); 17–22 min, 50–100% (B); 22–27 min, 100% (B); 27–30 min, 100– 10% (B); 30–35 min, 10% (B)。測定波長:340 nm。溶出時間:daidzein, 13.2 min; daidzein-7-glcoside, 7.8 min; genitein, 16.5 min; genitein-7-daidzein-7-glcoside, 10.2 min; quercetin, 14.3 min; quercetin-3-glcoside, 9.6 min;kaemperol, 16.9 min;kaemperol-7-quercetin-3-glcoside, 10.7 min;apigenin, 16.1
min;apigenin-7-glcoside, 11.3 min。 <for stilbene glucoside>
移動相:0–5 min, 5% (B); 5–20 min, 5–35% (B); 20–22.5 min, 35–100% (B); 22.5–27.5 min, 100% (B); 27.5–32.5 min, 100–5% (B); 32.5–37.5, 5% (B)。測定波長:300 nm。溶出時間:rhapontigenin, 23.1 min rhaponticin, 19.1 min。
<for stilbene aglycone>
移動相:0–22 min, 35–100% (B); 22–27 min, 100% (B)。測定波長:300 nm。溶出時間: rhapontigenin, 4.4 min。
【酵素反応速度論解析】
酵素反応速度論解析のための酵素反応は精製酵素15 μg を用いて 30℃で 2 min もしくは 4 min インキュベートした。アクセプター基質の解析には 10 mM UDP-glucose に対して 0.1 mM ~4 mM (HDMF, vanillin) または 50 μM~1 mM (emodin, rhapontigenin) アクセプター基質を 用い、ドナー基質の解析には5 mM HDMF または 2.5 mM emodin に対してそれぞれ 0.1 mM ~4 mM または 50 μM~1 mM UDP-glucose を用いた。パラメータは Hanes–Woolf plot より 算出した。
【遺伝子発現解析】
イチゴの各器官から抽出した total RNA を鋳型として、High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific) を用いて cDNA を合成した。それぞれの特異的 primer (Table 8) および GoTaq® Green Master Mix を用いて DNA 断片を増幅した。β-actin は 内在性コントロールとして用いた。PCR 産物は 1.5%アガロースゲルで電気泳動を行い、 SAFELOOK Post-Green Nucleic Acid Stain (富士フイルム和光純薬) で染色を行った。バンド のシグナル強度の解析はImage J (National Institutes of Health, Bethesda, MD, USA) を用いて 行った。
モミジバダイオウの各器官から抽出したtotal RNA を鋳型として、High-Capacity RNA-to-cDNA™ Kit (Thermo Fisher Scientific) を用いて cDNA を合成した。それぞれの特異的 primer (Table 9) および、SYBR™ Select Master Mix (Thermo Fisher Scientific)を用いて StepOne™リア ルタイムシステム (Thermo Fisher Scientific) で解析を行い、ΔΔCt 法を用いて発現量を定量 した。β-actin を内在性コントロールとして用いた。
Table 8 Semi-quantitative RT-PCR に用いた primer Primer (name) Sequence (5' to 3')
UGT78A16/17 fw-semi CCACTTGCAACCGCAGTTTTCA rv-semi AATCAAAGGCACTCCACCTGCTAC
37
UGT85K16 fw-semi GGCTCAAAGACATCCCTAGCTTCA rv-semi TGCTAGCACTTGCTCTTGTGGA
Table 9 qRT-PCR に用いた primer
Primer (name) Sequence (5' to 3')
RpUGT1 fw-qRT TCCGAGGAAGATGTTGGAGTCTTG rv-qRT GTAGATGACGGAGTTTGGAGGCTT RpUGT2 fw-qRT AACGATGCCTACAAGTGGGTCCTA rv-qRT CCCCTTCTACAGCCTCAAAGCTAT RpUGT3 fw-qRT CAATTCTTGAGCCACGCAACCA rv-qRT AGGAATCCGTTCGTCACTGGACAA 【イチゴおよびモミジバダイオウの代謝物解析】 植物体各器官各0.1 g に対し、1 ml の 70% MeOH を加えて 30 分の超音波抽出を 3 回行い、 抽出液を遠心分離し上清を濃縮乾固した。得られたエキスを300 µl の MeOH に再懸濁し 10 倍濃縮とし、前述のHPLC 条件で分析した。 【配糖化酵素のアライメントおよび分子系統学的解析】 配糖化酵素のアライメントおよび分子系統学的解析には、Clustal W を用いた。近隣接合法 (NJ 法) によってアミノ酸配列のマルチプルアライメントと TreeView を用いた分子系統樹 を 作 成 し た 。 パ ラ メ ー タ ー は 、Protein Weight Matrix; BLOSUM, GAP OPEN; 10, GAP EXTENSION; 0.20, GAP DISTANCE; 5, No end gaps; no, ITERATION; none, NUMITER; 1, CLUSTERING; NJ に設定して計算した。各酵素の起源と accession number を Table 10 に示し た。
Table 10 分子系統学的解析に用いた植物配糖化酵素
Name Species Fuction Accession No. Aralia AcGat Aralia corbata Anthocyanin
3-O-galactosyltransferase
AB103471 B5GT Dorotheanthus Betanidin 5-O-glucosyltransferase Y18871 Zea bx8 Zea mays Benzoxazinoid
2-O-glucosyltransferase
AAL57037 BpUGAT Bellis perennis Anthocyanidin 3-O-glucoside
1,2-O-glucosyltransferase
BAD77944 CaUGT1 Catharanthus
roseus
UDP-glucose glucosyltransferase BAD29721 CaUGT2 Curcumin glucosyltransferase AB159213 CaUGT3 flavonoid glucoside
1,6-O-glucosyltransferase
BAH80312 CaUGT4 flavonoid glucoside
1,6-O-glucosyltransferase
BAH80313 CaUGT5 UDP-glucose glucosyltransferase BAH80314 CrUGT6
(UGT85A23)
Iridoid 1-O-glucosyltransferase Ab591741 CrUGT7
(UGT76A2)
Glucosyltransferase with braod substrate specificity
CrUGT8 (UGT709C2)
Iridoid 1-O-glucosyltransferase Cm1 2RhaT Citrus maxima Flavanone 7-O-glucoside
1,2-O-rhamnosyltransferase AAL06646 GjUGT2 (UGT85A24) Gardenia jasminoides
Ilidoid 1-O- glucosyltransferase AB555732 F7GT Scutellaria
baicalensis
Flavonol 7-O- glucosyltransferase BAA83484 In3GGT Ipomoea nil Anthocyanidin 3-O-glucoside
2”-O-glucosyltransferase
AB192314 NtGT1a Nicotina tabacum Glucosyltransferase with broad
substrate specificity
AB052558 NtGT2 Flavonol 7-O-coumarin AB072919 NtGT3 Glucosyltransferase with broad AB072918
substrate specificity
NtSAGT Salicylic acid glucosyltransferase AAF61647 Petunia 6RhaT Petunia hybrid Anthocyanidin 3-glucoside
1,6-O-rhamnosyltransferase
CAA50376 SaGT4 Solanum Seroidal sapogenin 3-O-
glucosyltransferase
BAD89042 SjUGT4
(UGT709C4)
Swertia japonica
UGT94D1 Sesamum indicum Sesaminol 2-O-glucoside 1,6-O- glucosyltransferase
39
UGT85B1 Sorghum bicolor p-Hydroxymandelonitrile O- glucosyltransferase
AAF17077 UGT76G1 Stevia rebaudiana Stevioside glucosyltransferase AAR06912 UGTCs2 Crocus sativus Crocetin glucosyltransferase AAP94878 UGT71G1 Medicago truncatula Flavonoid 3’-O- glucosyltransferase AAW56092 UGT71C1 Arabidopsis thaliana Hydroxycinamate, flavonoid3’-O- glucosyltransferase AC005496 UGT72E2 Coniferylalcohol glucosyltransferase AB018119 UGT72B1
UGT73C5 Zeatin O- glucosyltransferase AAD20156 UGT74B1 Thiohydroximate S-
glucosyltransferase
AV002396 UGT75B1 Indole 3-acetic acid,sinapic acid
glucosyltransferase
AAF79370 UGT76C1 Trans-zeatin N-glucosyltransferase BAB10792 UGT78D1 Flavonoid 3-O-glucosyltransferase AC009917 UGT85A1 Trans-zeatin O-glucosyltransferase AAF18537 UGT85H2 Medicago
truncatula
Flavonoid glycosyltransferase 2PQ6_A UGT92A1 Mucuna pruriens UDP-glycosyltransferase RDX95609.1 UGT95A1 Pilosella
officinarum
glycosyltransferase ACB56927.1 UGT71K3 Fragaria× ananssa anthocyanidin
3-O-glucosyltransferase
XP_004294260.1 UGT73B13 Linum
usitatissimum
UDP-glycosyltransferase AFJ52930 UGT73B24 Fragaria vesca
subsp. vesca
scopoletin glucosyltransferase XP_004304714.1 UGT71W1 UDP-glucose glucosyltransferase BBD34309.1 UGT85K14 Vitis vinifera UDP-glucose:furaneol
glucosyltransferase
BAS68504.1 UGT85K11 Camellia sinensis UDP-glycosyltransferase BAO51834.1 UGT85A28 Vitis vinifera 7-deoxyloganetin
glucosyltransferase
XP_002285770 UGT85A30 UDP-glycosyltransferase NP_001277170.1 UGT85A33 UDP-glycosyltransferase BAO51843.1 UGT71A16 Pyrus communis glycosyltransferase ACZ44836 UGT73B23 Fragaria vesca
subsp. vesca
scopoletin glucosyltransferase XP_004304022.1 UGT71A1 Manihot esculenta UTP-glucose glucosyltransferase CAA54611 UGT73B1 Arabidopsis
thaliana
AAN31894 UGT73C1 glucosyl transferase AAD20151
UGT73C6 glucosyl transferase AAD20155 UGT73D1 glucosyltransferase CAB64218 VvGT1 Vitis sinifera Flavonoid 3-O-glucosyltransferase AAB81682 FaGT1 Fragaria× ananssa UDP-glucose glucosyltransferase AAU09442 FaGT2 UDP-glucose glucosyltransferase AAU09443 FaGT3 UDP-glucose glucosyltransferase AAU09444 FaGT4 Putative UDP-rhamnose:
rhamnosyltransferase
AAU09445 FaGT5 UDP-glucose glucosyltransferase ABB92747 FaGT6 UDP-glucose glucosyltransferase ABB92748 FaGT7 UDP-glucose glucosyltransferase ABB92749 AdGT4 Actinidia deliciosa glycosyltransferase AIL51400.1 FaGT8
(UGT708E1)
Fragaria× ananssa UDP-glucose glucosyltransferase BBD34307.1 FaGT9 (UGT95B3) UDP-glucose glucosyltransferase BBD34308.1 FaGT10 (UGT71W1) UDP-glucose glucosyltransferase BBD34309.1 FaGT11 (UGT78A16) UDP-glucose glucosyltransferase BBD34310.1 FaGT12 (UGT78A17) UDP-glucose glucosyltransferase BBD34311.1 FaGT13 (UGT85K16) UDP-glucose glucosyltransferase BBD34312.1 RpUGT1 (UGT73BE4)
Rheum palmatum UDP-glucose glucosyltransferase LC528215 RpUGT2 (UGT72B49) UDP-glucose glucosyltransferase LC528216 RpUGT3 (UGT71AQ1) UDP-glucose glucosyltransferase LC528217 (Table 10 続き)
41 【引用文献】
1. Lim, E. K. Plant glycosyltransferases: Their potential as novel biocatalysts. Chem. - A Eur. J. 11, 5486–5494 (2005).
2. Caputi, L., Malnoy, M., Goremykin, V., Nikiforova, S. & Martens, S. A genome-wide
phylogenetic reconstruction of family 1 UDP- glycosyltransferases revealed the expansion of the family during the adaptation of plants to life on land. Plant J. 69, 1030–1042 (2012).
3. Gachon, C. M. M., Langlois-Meurinne, M. & Saindrenan, P. Plant secondary metabolism glycosyltransferases: The emerging functional analysis. Trends Plant Sci. 10, 542–549 (2005). 4. Kleczkowski, K., Schell, J. & Bandur, R. Phytohormone conjugates: Nature and function. CRC.
Crit. Rev. Plant Sci. 14, 283–298 (1995).
5. Rea, P. MRP subfamily ABC transporters from plants and yeast. J. Exp. Bot. 50, 895–913 (1999). 6. Messner, B., Thulke, O. & Schäffner, A. R. Arabidopsis glucosyltransferases with activities
toward both endogenous and xenobiotic substrates. Planta 217, 138–146 (2003).
7. Taguchi, G., Yazawa, T., Hayashida, N. & Okazaki, M. Molecular cloning and heterologous expression of novel glucosyltransferases from tobacco cultured cells that have broad substrate specificity and are induced by salicylic acid and auxin. 4094, 4086–4094 (2001).
8. Kaminaga, Y., Sahin, F. P. & Mizukami, H. Molecular cloning and characterization of a glucosyltransferase catalyzing glucosylation of curcumin in cultured Catharanthus roseus cells. FEBS Lett. 567, 197–202 (2004).
9. Cui, L. et al. Identification of UDP-glycosyltransferases involved in the biosynthesis of astringent taste compounds in tea (Camellia sinensis). J. Exp. Bot. 67, 2285–2297 (2016).
10. K.K Makinen, E. S. A quantitative study of mannitol, sorbitol, xylitol, and xylose in wild berries and commercial fruits. J. Food Sci. 45, 367–371 (1980).
11. Basu, A., Nguyen, A., Betts, N. M. & Lyons, T. J. Strawberry as a functional food: An evidence-based review. Crit. Rev. Food Sci. Nutr. (2014). doi:10.1080/10408398.2011.608174
12. DSファーマアニマルヘルス プレスリリース 2014年3月24日.
13. Mayerl, F., Näf, R. & Thomas, A. F. 2,5-Dimethyl-4-hydroxy-3(2H)-furanone glucoside: Isolation from strawberries and synthesis. Phytochemistry 28, 631–633 (1989).
14. Ubeda, C. et al. Glycosidically bound aroma compounds and impact odorants of four strawberry varieties. J. Agric. Food Chem. 60, 6095–6102 (2012).
15. Tokitomo, Y., Steinhaus, M., Büttner, A. & Schieberle, P. Odor-active constituents in fresh pineapple (Ananas comosus [L.] Merr.) by quantitative and sensory evaluation. Biosci. Biotechnol. Biochem. 69, 1323–30 (2005).
16. Janeš, D., Kantar, D., Kreft, S. & Prosen, H. Identification of buckwheat (Fagopyrum esculentum Moench) aroma compounds with GC-MS. Food Chem. 112, 120–124 (2009).
tomato using dynamic headspace sampling with sodium sulfate. J. Agric. Food Chem. 49, 4349– 4351 (2001).
18. Raab, T. et al. FaQR, required for the biosynthesis of the strawberry flavor compound 4-hydroxy-2,5-dimethyl-3(2H)-furanone, encodes an enone oxidoreductase. Plant Cell 18, 1023–1037 (2006).
19. Klein, D., Fink, B., Arold, B., Eisenreich, W. & Schwab, W. Functional characterization of enone oxidoreductases from strawberry and tomato fruit. J. Agric. Food Chem. 55, 6705–6711 (2007). 20. Schwab, W. Natural 4-hydroxy-2,5-dimethyl-3(2H)-furanone (Furaneol®). Molecules 18, 6936–
51 (2013).
21. Roscher, R., Herderich, M., Steffen, J. P., Schreier, P. & Schwab, W. 2,5-Dimethyl-4-hydroxy-3[2H]-furanone 6’ O -malonyl-β-D-glucopyranoside in strawberry fruits. Phytochemistry 43, 155–
9 (1996).
22. Lunkenbein, S. et al. Cinnamate metabolism in ripening fruit. Characterization of a UDP-glucose: Cinnamate glucosyltransferase from strawberry. Plant Physiol. 140, 1047–1058 (2006).
23. Griesser, M. et al. Redirection of flavonoid biosynthesis through the down-regulation of an anthocyanidin glucosyltransferase in ripening strawberry fruit. Plant Physiol. 146, 1528–1539 (2008).
24. Landmann, C., Fink, B. & Schwab, W. FaGT2: A multifunctional enzyme from strawberry (Fragaria x ananassa) fruits involved in the metabolism of natural and xenobiotic compounds. Planta 226, 417–428 (2007).
25. Griesser, M. et al. Multi-substrate flavonol O-glucosyltransferases from strawberry (Fragaria x ananassa) achene and receptacle. J. Exp. Bot. 59, 2611–25 (2008).
26. Song, C. et al. Glucosylation of 4-hydroxy-2,5-dimethyl-3(2H)-furanone, the key strawberry flavor compound in strawberry fruit. Plant Physiol. 171, 139–151 (2016).
27. Hirakawa, H. et al. Dissection of the octoploid strawberry genome by deep sequencing of the genomes of fragaria species. DNA Res. 21, 169–181 (2014).
28. Sasaki, K., Takase, H., Kobayashi, H., Matsuo, H. & Takata, R. Molecular cloning and characterization of UDP-glucose: furaneol glucosyltransferase gene from grapevine cultivar Muscat Bailey A (Vitis labrusca × V. vinifera). J. Exp. Bot. 66, 6167–74 (2015).
29. Song, C. et al. A UDP-glucosyltransferase functions in both acylphloroglucinol glucoside and anthocyanin biosynthesis in strawberry (Fragaria × ananassa). Plant J. 85, 730–742 (2016). 30. Ohgami, S. et al. Volatile glycosylation in tea plants: Sequential glycosylations for the
biosynthesis of aroma β -primeverosides are catalyzed by two Camellia sinensis glycosyltransferases. Plant Physiol. 168, 464–477 (2015).
31. Zheng, Q. et al. Review of rhubarbs: Chemistry and pharmacology. Chinese Herb. Med. 5, 9–32 (2013).
43
32. Caro, Y. et al. Natural hydroxyanthraquinoid pigments as potent food grade colorants: an overview. Nat. Products Bioprospect. 2, 174–193 (2012).
33. Demirezer, L. O., Kuruüzüm-Uz, A., Bergere, I., Schiewe, H. J. & Zeeck, A. The structures of antioxidant and cytotoxic agents from natural source: anthraquinones and tannins from roots of Rumex patientia. Phytochemistry 58, 1213–7 (2001).
34. Yonekura-sakakibara, K., Fukushima, A. & Saito, K. Transcriptome data modeling for targeted plant metabolic engineering. Curr. Opin. Biotechnol. 24, 285–290 (2013).
35. Hirotani, M., Kuroda, R., Suzuki, H. & Yoshikawa, T. Cloning and expression of UDP-glucose: Flavonoid 7-O-glucosyltransferase from hairy root cultures of Scutellaria baicalensis. Planta 210, 1006 (2000).
36. Funaki, A. et al. Identification of a highly specific isoflavone 7-O-glucosyltransferase in the soybean (Glycine max (L.) Merr.). Plant Cell Physiol. 56, 1512–1520 (2015).
37. Nagatoshi, M., Terasaka, K., Nagatsu, A. & Mizukami, H. Iridoid-specific glucosyltransferase from Gardenia jasminoides. J. Biol. Chem. 286, 32866–32874 (2011).
38. Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
【謝辞】 本研究を遂行するに当たり、終始懇篤なご指導、ご鞭撻を賜りました名古屋市立大学大学院 薬学研究科 牧野 利明 教授に心からの御礼申し上げます。 本研究を遂行するに当たり、終始懇篤なご指導、ご鞭撻を賜りました名古屋市立大学大学院 薬学研究科 石内 勘一郎 講師に心からの御礼申し上げます。 本研究を遂行するに当たり、終始懇篤なご指導、ご鞭撻を賜りました高知県立牧野植物園 水上 元 園長 (名古屋市立大学名誉教授) に心からの御礼申し上げます。 本研究を遂行するに当たり、イチゴ植物体を御分与いただきました名古屋市立大学薬用植 物園の皆様に深甚なる感謝の意を表します。 本研究を遂行するに当たり、様々な御助言、御討論、御協力を頂きました名古屋市立大学大 学院薬学研究科生薬学研究室の皆様に深甚なる感謝の意を表します。 最後に、本研究を遂行するに当たり、終始懇篤なご指導、ご鞭撻を賜りました名古屋市立大 学大学院薬学研究科 寺坂 和祥 講師に甚大なる感謝の意を表します。