2018 年 4 月 25 日発行
記録 ・ 調査報告 Note
1〒 986–2242 宮城県牡鹿郡女川町小乗浜字向 3–1 東北大学女川フィールドセンター 2〒 041–8611 北海道函館市港町 3–1–1 北海道大学水産学部 3〒 041–8611 北海道函館市港町 3–1–1 北海道大学大学院水産科学院 4〒 041–8611 北海道函館市港町 3–1–1 北海道大学大学院水産科学研究科・水産科学研究院 (2017 年 7 月 11 日受付;2017 年 10 月 22 日改訂;2017 年 10 月 24 日受理;2018 年 3 月 9 日 J–STAGE 早期公開) キーワード:アカガレイ , 噴火湾 , 遺伝的多様性 , ミトコンドリア DNA(mtDNA) Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2018Yuki Yamamoto, Minoru Ikeda, Mitsuhiro Nakaya*, Kota Suzuki and Tetsuya Takatsu. 2018. Genetic diversity of the flathead flounder Hippoglossoides dubius population in Funka Bay inferred from mitochondrial DNA analysis. Japan. J. Ichthyol., 65(1): 75–80. DOI: 10.11369/jji.17-034
Abstract The flathead flounder Hippoglossoides dubius population in Funka Bay,
characterized by unique ecological features, has been thought to be demographically independent of other conspecific populations. To clarify such, sequence data of the hyper variable domain in the mitochondrial DNA control region was collected from the Funka Bay population and the genetic diversity compared with previously published data for other populations (Sea of Japan and northwestern Pacific off Tohoku–Hokkaido). Because the level of diversity of the former population differed little from that of other populations, with no significant genetic differentiation detected, it is likely that contemporary or recent historical gene flow has occurred extensively in Japanese coastal waters. The likelihood of gene flow and connectivity among the populations, all characterized by long pelagic egg and larval stages, being associated with the Tsugaru Warm Current is considered.
*Corresponding author: Faculty of Fisheries, Hokkaido University, 3‒1‒1 Minato-cho, Hakodate, Hokkaido 041‒8611, Japan (e-mail: [email protected])
ア
カ ガ レ イ Hippoglossoides dubius は 日 本 で は 山口県以北の日本海,宮城県以北の太平洋, および北海道全域の水深 500 m 以浅の砂泥底に分 布する漁業対象種である(尼岡,2016).遺伝的 手法を用いて区分した系群ではないが,主な漁場 である島根県から青森県沖合の日本海で漁獲され るものを本州沖日本海系群,太平洋に開口した北 海道噴火湾で漁獲されるものを噴火湾系群として 資源評価が行われている[1991‒2015 年の年間漁 獲 量 は そ れ ぞ れ 2,281‒6,158 ト ン( 藤 原・ 上 田, 2016),382‒2,607 トン(北海道立総合研究機構水 産研究本部,2017)].一般に魚類資源の加入量を 左右するとされる生活史初期の生態について,本 種 は 分 離 浮 性 卵 を 産 出 し, ふ 化 後 長 い 浮 遊 期 (6‒15˚C の飼育環境下で 90‒120 日)を経て,底生 生 活 に 移 行 す る こ と が 知 ら れ て い る( 松 田, 2017).日本海では 2‒4 月に産卵がみられ,西部 における主産卵場は,若狭湾内,経ヶ崎および赤 碕沖を中心とする隠岐諸島周辺(永澤,1993), 北部では新潟県粟島北方の海域と考えられている (藤原・上田,2016).Kojima et al.(2014)は日本 海南部(島根県太田沖),北部(北海道留萌沖) および太平洋東北・北海道(宮古沖,釧路沖)で 採集されたアカガレイのミトコンドリア DNA(以 下,mtDNA と記す)分析を行い,海域間の遺伝 的差異は認められず,広範な遺伝子流動が生じて いることを示唆した. 一方,噴火湾では主に 1‒3 月に湾奥部で産卵がミトコンドリア DNA によって検討した噴火湾に生息する
アカガレイ個体群の遺伝的多様性
山本佑樹
1・池田 実
1・中屋光裕
2・鈴木孝太
3・髙津哲也
4みられ(鈴内,1983;横山ほか,1991),卵・仔 魚が湾内に多数分布することが知られており(宮 本ほか,1993),当湾周辺で行ったトロール調査 や標識放流実験の結果,未成魚・成魚は年間を通 して湾内を主生活域としていることが確認されて いる(横山ほか,1990).本種の漁獲量は卓越年 級群の発生により支えられているが(中谷ら, 1990;藤原・上田,2016),噴火湾系群は本州沖 日本海系群とは異なる漁獲量の推移を示している ため(本間,2009;藤原・上田,2016),卓越年 級群の発生する年が異なる可能性がある.また, 本州沖日本海系群の産卵場の 1 つである若狭湾で は,中・後期卵および仔魚は主に水深 100 m より も浅い層(冬季の鉛直混合が生じており,水深に よる温度差はほとんどない)で採集され,水温は 10˚C 前後である(永澤,1993;安達,2006;藤原・ 上田,2016)のに対し,噴火湾では卵・仔魚は水 深 15 m よりも浅い層で多く採集されており(鈴内, 1983),水温は高くても 6˚C,低い場合は 2˚C を 下回る時期に出現する(宮本ほか,1993).この ように成魚の季節移動,卓越年級群の発生状況お よび卵・仔魚の出現時期の水温が大きく異なるこ とから,噴火湾に生息するアカガレイ個体群は本 州沖日本海系群とは遺伝的に異なっている可能性 がある.しかし,当湾に生息するアカガレイにつ いて,これまでに遺伝学的解析は行われておらず, 他海域との遺伝子流動の有無については不明なま まである.本研究は,噴火湾で採集されたアカガ レ イ に つ い て Kojima et al.(2014) と 同 様 の mtDNA 分析を行い,標本集団内の遺伝的多様性 ならびに Kojima et al.(2014)によって報告され た他海域の標本集団との間の遺伝的分化の有無に ついて調べた. 材 料 と 方 法 2017 年 2 月に噴火湾において北海道大学水産学 部附属練習船うしお丸(179 t)を用いて行われた 幼魚用着底トロールで採集されたアカガレイ成魚 36 個体(全長範囲 : 202‒355 mm)を分析した.冷 凍保存された各個体の胸鰭を試料とし,QuickGene DNA tissue kit S と Mini80(いずれも KURABO 社) により DNA 抽出を行った.抽出された DNA をテ ンプレートとして mtDNA の調節領域前半部 475 bp を PCR 増 幅 し た.PCR 反 応 液 の 組 成 は, 鋳 型 DNA 溶 液 1.0 µL, 超 純 水 8.88 µL,10 × Buffer (TOYOBO 社)1.25 µL,dNTP Mixture 1.25 µL,プラ イマー(50 µM)各 0.25 µL,Blend Taq-Plus(TOYOBO 社)0.125 µL の計 13 µL とした.プライマーならび に PCR の温度サイクル条件は Kojima et al.(2014) に従った.増幅の有無を 1% アガロース電気泳動 により確かめた後,反応液の一部を AMPURE XP (Bechman Coulter 社)を用いて精製し,サイクルシー ケンス反応に供した.サイクルシーケンス反応は BigDye Terminator Sequencing Standard Kit ver. 3.1 (Life Technology 社)を用い,反応液の組成と温度 サイクル条件ならびに反応後の精製は添付のマニュ ア ル に 従 っ た.3500 XL Genetic Analyzer(Life Technology 社)によりキャピラリー電気泳動を行い, 塩基配列を解読した.解読した配列中から Kojima et al.(2014)と同じ 475 サイトを解析対象とした. また,DNA データバンクに登録されている Kojima et al.(2014)による日本海南部(島根県太田沖, Acc. No. AB859317, AB859323‒AB859339),北部(北 海道留萌沖,Acc. No. AB859317, AB859320, AB859331, AB859334, AB859337, AB859339, AB859340, AB859351, AB859354, AB859360, AB859366‒ AB859382)および北東太平洋(宮古沖,Acc. No. AB859318, AB859321, AB859322, AB859335, AB859337, AB859340‒AB859347; 釧 路 沖,Acc. No. AB859317, AB859319, AB859327, AB859331, AB859334, AB859337, AB859338, AB859349‒ AB859365)のアカガレイ標本集団の配列データを 加えて,噴火湾におけるアカガレイ標本集団の遺 伝的多様性ならびに他の標本集団との間の遺伝的
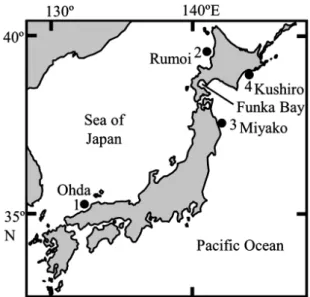
Fig. 1. Sampling sites of Hippoglossoides dubius.
Numerals (1‒4) indicate sampling sites in Kojima et al. (2014).
分化の有無について検討した. 噴火湾ならびに Kojima et al.(2014)が調べた 標本集団の地理的位置を Fig. 1 に示す.標本集団 内の遺伝的多様性指数としてハプロタイプ多様度 (h;Nei, 1987)と塩基多様度(π;Nei, 1987)を求 めた.標本集団全体の構造化の有無ならびに標本 集団間の遺伝的分化の有無について検討するため, AMOVA(Excoffier et al., 1992) と ペ ア ワ イ ズ FST 分析(Weir and Cockerham, 1984)を行った(いず れも 104 permutation).これらの計算には Arlequin ver. 3.5.2(Excoffier and Lischer, 2010)を使用した. また,ハプロタイプ間の系統関係について検討す るため , Median Joining 法(Bandelt et al., 1999)に よりネットワーク系統樹を作成した.系統樹の作 成には Network 5.0.0.1(http://www.fluxus-technology. com)を用いた. 結 果 と 考 察 対象とした噴火湾のアカガレイ 36 個体につい て遺伝的多様性指数を算出した結果,ハプロタイ プ 数 は 29 個(Acc. No. LC310823‒LC310851), ハ プロタイプ多様度(h)は 0.984,塩基多様度(π) は 0.00982 となり,Kojima et al.(2014)による他 の標本集団の値と比較してもほぼ同レベルの値を 示した(Table 1).標本集団間の遺伝的分化の有 無を明らかにするため AMOVA 分析を行った結果, 標本集団間の差異によって説明できる遺伝的変異 性は全体の遺伝的変異性の 0.86%(P = 0.116)と なり,構造化は生じていないと考えられた.さら に標本集団間のペアワイズ FST 値を求め,シー ケンシャルボンフェローニ補正(Rice, 1989)の 下で FST 値が 0 より有意に大きい組み合わせが あるか否かを調べた結果,そのような組み合わせ はみられなかった(Table 2).今回の調査で検出 された噴火湾の 29 個のハプロタイプと Kojima et al.(2014)で検出された 83 個のハプロタイプのネッ トワーク系統樹を Fig. 2 に示す.ネットワークの 中心部にある主要なハプロタイプはほとんどの標 本集団間で共有されており,いずれかの標本集団 で派生的なハプロタイプが特異的に保有されてい るという傾向は認められなかった.これらの結果 は,噴火湾も含めた日本列島沿岸におけるアカガ レイには海域間の遺伝的な違いはなく,Kojima et al.(2014)が示唆したように近い過去も含めて広 範囲な遺伝子流動が生じていることを意味してい るのかもしれない. 一般に,魚類の遺伝子流動は,未成魚・成魚の 移動あるいは卵・仔魚の分散によってもたらされ ている(西田ら,1998;秋元ら,2003).沈性付 着卵を産出するハタハタでは,卵・仔魚期の輸送 による分布の拡大よりも親魚の広範囲な回遊に よって高い遺伝子流動がもたらされていることが 示唆されている(柳本,2004).また,スケトウ
Table 1. Genetic diversity of Hippoglossoides dubius populations
Table 2. Pariwise FST estimates among populations of Hippoglossoides dubius (lower) and corresponding p-values (upper).
All FST values were not significantly greater than 0 following sequential Bonferroni correction (Rice, 1989)
Sampling site Number of samples Number of haplotypes Haplotype diversity (h ± SD) Nucleotide diversity (π ± SD)
Funka Bay 36 29 0.984 ± 0.012 0.00982 ± 0.00548
Off Ohda* 20 18 0.990 ± 0.019 0.00767 ± 0.00453
Off Rumoi* 33 27 0.983 ± 0.014 0.00779 ± 0.00449
Off Miyako* 14 13 0.989 ± 0.031 0.00960 ± 0.00563
Off Kushiro* 31 24 0.976 ± 0.016 0.00859 ± 0.00490
*Data were referred from Kojima et al. (2014).
Sampling site Funka Bay Off Ohda* Off Rumoi* Off Miyako* Off Kushiro*
Funka Bay – 0.418 0.047 0.037 0.688
Off Ohda* 0.0009 – 0.291 0.279 0.247
Off Rumoi* 0.0189 0.0046 – 0.830 0.186
Off Miyako* 0.0349 0.0075 -0.0187 – 0.081
Off Kushiro* -0.0058 0.0078 0.0086 0.0295 –
ダラは北太平洋,アラスカ湾および日本海に広く 分布しており,噴火湾内でも冬季に浮遊卵・仔魚 が多数採集されているが,未成魚や成魚の能動的 な移動や浮遊卵・仔魚の受動的輸送のため,日本 海,北太平洋からアラスカ湾にかけて遺伝的分化 が顕著ではないことが確認されている(Yanagimoto et al., 2004, 2012).噴火湾に生息するアカガレイ の生態的特徴として,成魚の湾外への移動はほぼ ないこと(横山ほか,1990),主産卵場が湾内に 形成されること(鈴内,1983;横山ほか,1991), 卵・仔魚は主に湾内に分布すること(宮本ほか, 1993),分離浮性卵で卵・仔魚期が長いこと(松田, 2017;鈴木ほか,2017)が知られている.これら の知見から判断すると,湾内に一度着底した個体 が大規模回遊を行い,他海域に分布する個体群と 遺伝的に交流する可能性は低く,むしろ卵・仔魚 期に海流により輸送されることで遺伝子流動が生 じる可能性が高いと考えられる.津軽海峡の東側, 北海道沿岸の表層域では,沿岸親潮が海峡中央部 にあたる函館山付近まで西向きに流れ,海峡中央 部の最深部ではごく弱い西向きの流れが観測され ることがある(杉本,1985).一方,海峡西口は 全水深層が津軽暖流で覆われ,周年日本海から太 平洋に向かって流れており,太平洋側の水塊が日 本海側まで達することは異常低水温年の春季を除 いてない(大谷,1987;西田,2003).海峡東口 を太平洋に出た津軽暖流の挙動は年により異なり, 南下して八戸・三陸方面へ向かう沿岸モードと, 東進して襟裳岬方面へ向かう渦モードの 2 つに分 け る こ と が で き る(Conlon, 1982; 杉 本・ 川 崎, 1984;菱田,1987).また,一部は北上して噴火 湾へ流入することが知られている(Conlon, 1982; 菱田,1987).本種の日本海個体群は沿岸モード で八戸・三陸方面,渦モードで襟裳岬から道東沿 岸に生息する個体群との間で遺伝子流動が生じて いる可能性がある.これらと同様,噴火湾内に生 息するアカガレイについても,日本海で発生した 卵・仔魚が津軽暖流により噴火湾内に輸送される ことで,日本海個体群との間で不定期に遺伝子流 動が生じていることが予想される(Fig. 3). 本研究から,噴火湾と他地域のアカガレイで遺 伝的分化が起きていないことが明らかとなった. しかし,これは噴火湾と他地域が同一の個体群で あることを必ずしも意味するものではない.遺伝 子分化は少しの遺伝子流動があれば抑制されるが (Waples and Gaggiotti, 2006),少数の個体が移動し ても個体群デモグラフィーには影響しない.実際, 遺伝的分化が起きていない地域間でも個体群動態 や性比やサイズ構造が異なることが知られている (Koizumi et al., 2008).また,実際に遺伝子流動が なくても,有意な遺伝的分化が認められない場合 もある.遺伝子流動が途絶えたのが最近であり,
有効集団サイズが大きい場合,たとえ移動が制限 されていても遺伝的分化は検出されない(例えば, Teixeira et al., 2011;Garvin et al., 2013).海産魚類 は一般的に有効集団サイズが非常に大きい.もし アカガレイが噴火湾に隔離されたのが最終氷河期 以降など比較的最近であれば,遺伝的分化が認め られない可能性も十分にある.以上を総合すると, 現時点では,これまで通り噴火湾のアカガレイは ひとつの系群として資源評価を行うのが予防原則 的にも安全である.今後,マイクロサテライト DNA や SNP な ど 解 像 度 の よ り 高 い 遺 伝 子 マ ー カーを用いた解析や生態的特性を考慮した総合的 な研究が必要である. 謝 辞 本研究の分析を行うに当たり,ご助言・ご協力 頂いた東京大学大気海洋研究所小島茂明教授に感 謝致します.また,標本採集等にあたりご協力を 頂いた北海道大学水産学部附属練習船うしお丸亀 井佳彦船長はじめ乗組員各位に御礼申し上げま す.本研究の一部は,科学研究費補助金基盤研究 (C)(課題番号 20580189, 25450269) および公益 財団法人北水協会水産研究助成金によって行われ た. 引 用 文 献 安 達 辰 典.2006. 若 狭 湾 に お け る ア カ ガ レ イ Hippoglossoides dubius の卵および稚仔魚の鉛直分 布.日本海ブロック資源研究会報告(平成 15・ 16 年度),日本海区水産研究所,新潟.27. 秋 元 清 治・ 瀬 崎 啓 次 郎・ 三 谷 勇・ 渡 部 終 吾. 2003.ミトコンドリア制御領域の塩基配列分析 による日本周辺漁場におけるキンメダイの集団 遺伝構造の解析.神奈川県水産研究所研究報告, 8: 89‒97. 尼岡邦夫.2016.日本産ヒラメ・カレイ類.東海 大学出版部,東京,229 pp.
Bandelt, H. J., P. Forster and A. Röhl. 1999. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol., 16: 37‒48.
Conlon, D. M. 1982. On the outflow modes of the Tsugaru Warm Current. La mer, 20: 60‒64.
Excoffier, L and H. E. L. Lischer. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour., 10: 564‒565.
Excoffier, L., P. E. Smouse and J. M. Quattro. 1992. Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics, 131: 479‒491. 藤原邦浩・上田祐司.2016.平成 27(2015)年度 アカガレイ日本海系群の資源評価.平成 27 年度 我が国周辺水域の漁業資源評価(魚種別系群別 資源評価・TAC 種以外)第 3 分冊,水産庁増殖 推 進 部, 東 京.1674‒1692. http://abchan.fra.go.jp/ digests27/details/2763.pdf(参照 2017-4-7).
Garvin, M. R., C. M. Kondzela, P. C. Martin, B. Finney, J. Guyon, W. D. Templin, N. DeCovich, S. Gilk-Baumer and A. J. Gharrett. 2013. Recent physical connections
Fig. 3 Image of current pattern near shore of Japan and
around Tsugaru Strait. Data from The Sasakawa Peace Foundation Ocean Policy Research Institute
(https://www.spf.org/opri-j/projects/information/ newsletter/backnumber/2003/80_3.html; 28 May 2017), Conlon (1982) and Hishida (1987).
may explain weak genetic structure in western Alaskan chum salmon (Oncorhynchus keta) populations. Ecol. and Evol., 3: 2362‒2377. 菱田昌孝.1987.津軽暖流の南下と季節変動につ いて.水路部研究報告,22: 1‒22. 北海道立総合研究機構水産研究本部.2017.データ ベ ー ス 検 索 . http://www.fishexp.hro.or.jp/list/fisheries/ marine/h3mfcd0000000ge0.html(参照 2017-4-7). 本間隆之.2009.噴火湾のアカガレイ調査~小さ なアカガレイの採集を対象としたソリネット調 査~.北水試験だより,79: 11‒14.
Koizumi, I., S. Yamamoto, K. Nomoto and K. Maekawa. 2008. Synchrony in local population dynamics of stream-dwelling Dolly Varden: do genetically similar groups show similar demography? Popul. Ecol., 50: 367‒377.
Kojima, S., K. Sakuma and T. Yanagimoto. 2014. Contrasting genetic population structures between congeneric flounder species, Hippoglossoides dubius and H. pinetorum. Plankton and Benthos Res., 9: 99‒104. 松田泰平.2017.アカガレイの人工種苗生産につ い て ‒ 採 卵 ~ 仔 稚 魚 の 飼 育 ‒. 試 験 研 究 は 今, 826. http://www.hro.or.jp/list/fisheries/marine/att/ ima826.pdf(参照 2017-2-3). 宮本孝則・髙津哲也・中谷敏邦・前田辰昭・高橋 豊美.1993.噴火湾とその沖合におけるアカガレ イ 卵・ 稚 仔 の 分 布 と 食 性. 水 産 海 洋 研 究,57: 1‒14. 永澤 亨.1993.山陰沿岸におけるアカガレイの 産卵場.漁業資源研究会議北日本底魚部会報, 26: 19‒25. 中谷敏邦・小泉広明・横山信一・前田辰昭・高橋 豊美・松島寛治.1990.噴火湾産アカガレイの年 齢と成長.日本水産学会誌,56: 893‒901.
Nei, M. 1987. Molecular Evolutionary Genetics. Columbia University Press, New York, 512 pp.
西田 睦・大河俊之・磐田祐士.1998.ミトコン ドリア DNA 分析による集団構造解析法.水産育 種,26: 81‒100. 西田芳則.2003.春季における津軽海峡を挟んだ 水塊構造の変質.海の研究,12: 593‒602. 大谷清隆.1987.津軽海峡西部への沿岸親潮水の 流 入. 北 海 道 大 学 水 産 学 部 研 究 彙 報,38: 209‒220.
Rice, W. R. 1989. Analyzing tables of statistical tests.
Evolution, 43: 223‒225. 杉本隆成.1985.津軽海峡内の海洋構造.日本海 洋学会沿岸海洋研究部会「沿岸海洋誌」編集委 員会(編),pp.149–154. 日本全国沿岸海洋誌.東 海大学出版会,東京 . 杉本隆成・川崎康寛.1984.津軽暖流の季節・経 年変動とその力学的解釈.沿岸海洋研究ノート, 21: 1–11. 鈴内孝行.1983.幼稚仔,海況調査.昭和 57 年度 北海道立函館水産試験場事業報告,15‒22. 鈴木孝太・中屋光裕・柳 海均・松田泰平・髙津 哲也.2017.アカガレイ卵・仔稚魚の比重変化お よび卵発生速度と水温の関係.日本水産学会誌, 83: 580‒588.
Teixeira, S., M-A. Cambon-Bonavita, E. A. Serrão, D. Desbruyéres and S. Arnaud-Haond. 2011. Recent population expansion and connectivity in the hydrothermal shrimp Rimicaris exoculata along the Mid-Atlantic Ridge. J. Biogeogr., 38: 564‒574.
Waples, R. S and O. Gaggiotti. 2006. What is a population? An empirical evaluation of some genetic methods for identifying the number of gene pools and their degree of connectivity. Mol. Ecol., 15: 1419‒1439.
Weir, B. S. and C. C. Cockerham. 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358‒1370.
柳本 卓.2004.mtDNA の PCR-RELP 分析によって 明らかになったハタハタ集団の地理的分化.日 本水産学会誌,70: 583‒591.
Yanagimoto, T., T. Kitamura and T. Kobayashi. 2004. Complete nucleotide sequence and variation of mitochondrial DNA from 10 individuals of walleye Pollock, Theragra chalcogramma. Fish. Sci., 70: 885‒895.
Yanagimoto, T., T. Kitamura and T. Kobayashi. 2012. Genetic stock structure of walleye Pollock (Theragra
chalcogramma) inferred by PCR-RELP analysis of the
mitochondrial DNA and SNP analysis of nuclear DNA. Mar. Genomics, 7: 17‒25. 横山信一・前田辰昭・中谷敏邦・高橋豊美・松島 寛治.1990.噴火湾およびその沖合におけるアカ ガレイの分布・移動について. 水産海洋研究, 54: 373‒380. 横山信一・前田辰昭・高橋豊美・中谷敏邦・松島 寛治.1991.噴火湾におけるアカガレイ成魚の生 活年周期.日本水産学会誌,57: 1469‒1476.