学位論文
Synechocystis
sp. PCC6803 の
環 境スト レス 耐性に 関与 する細 胞外 多糖
合 成関連 遺伝 子群の 解析
平成
29 年 3 月
松橋 歩
目次
要約 第1 章 研究背景 P.1-8 第2 章 材料と方法 P.9-34 第3 章 結果 P.35-109 第4 章 考察 P.110-115 参考文献 P.116-128 謝辞 P.129要約
この研究はシアノバクテリアSynechocystis sp. PCC6803 の酸性ストレス耐性に関与するス トレス応答タンパク質をコードする遺伝子の転写解析および細胞外多糖合成に関与する遺 伝子の解析に関わるものである。 生物は常に周囲の環境からのストレスに曝されており、ストレスに抵抗するために細胞 内で様々な適応機構を発達させてきた。環境ストレスには、温度、乾燥、塩、などが代表的 なものとして挙げられるが、近年においては、海洋の酸性化が問題視されている。 生態ピラミッドの生産者に位置するシアノバクテリアは好アルカリ性であることから、 海洋の酸性化が進むにつれて生態系に影響を及ぼす可能性が考えられる。そのため、本研究 はシアノバクテリアの酸耐性機構を明らかにすることを目的とした。 以前に当研究室で行ったDNA マイクロアレイ解析により、酸性ストレス下で約 30 個の 遺伝子が発現誘導されることが明らかになった(Ohta et al., 2005)。これらのうち、劇的に転 写が誘導されたsll1558 に着目した。sll1558 は UDP-glucose pyrophosphorylase をコードしており、gulucose-1-phosphate に uridine triphosphate を付加することによって UDP-glucose を合
成する酵素である(Maeda et al., 2014)。
始めに、Sll1558 をコードする遺伝子欠損株の構築を試みたが、完全欠損株を構築するこ
測定したところ、酸性ストレス下において野生株よりも有意に転写量が減少した。この変異 株を酸性ストレス下で生育させると、野生株よりも抑制されることが明らかになった。また、 細胞外多糖のリポ多糖(LPS)、莢膜多糖(CPS)、遊離多糖(RPS)の構造を調べると野生株と異 なる構造であることが判明した。さらに、sll1558 株は高塩や高浸透圧、温度ストレスでも 生育が抑制され、さらにUV-B ストレス下での生存率も低下したことから、細胞外多糖がこ れらのストレス耐性に関与することが示された。 これまでの結果では、LPS、CPS、RPS すべての構造が野生株と異なることから、酸性ス トレス耐性に重要な因子を特定できなかった。そこで、LPS が膜にアンカーされる lipid A を持つことを利用してLPS 欠損株を構築し、LPS がストレス耐性に重要であるかを確認し た。 大腸菌のMsbA は lipid A を輸送することが明らかにされており、LPS の構築に必要不可
欠なタンパク質である(Karow and Georgopoulos, 1993)。Synechocystis においても似た機能を
持つタンパク質の存在が予想されることから、BLAST 検索を行った。その結果、Synechocystis にはMsbA と相同性の高いタンパク質が 4 つ(Slr2019、Sll1276、Sll1725、Slr1149)見つかっ た。これらの遺伝子変異株を構築したが、sll1276 のみ完全欠損株を得ることができなかっ た。4 つの変異株から LPS を抽出して野生株と構造を比較すると、slr2019、sll1276 株のみ が野生株と異なる構造を持つことが明らかになった。また、4 つの変異株を酸性ストレス下 で生育すると、slr2019、sll1276 株の 2 つが生育抑制された。さらに、slr2019、sll1276、sll1725、
slr1149 の酸性ストレス下における転写量を測定すると、pH3.0 では sll1276 の転写量が増加 し、pH6.0 では slr2019 の転写量が増加した。これらの結果から、Synechocystis では lipid A flippase ホモログが 2 つ存在し、slr2019 は sll1276 の補助的な役割を果たしている可能性が 示唆された。 本研究は、細胞外多糖に着目してシアノバクテリアの酸性ストレス耐性を検討した初めて の 研 究 で あ る 。 細 胞 表 層 の 構 築 に 寄 与 す る sll1558 の遺伝子変異株の解析により、 Synechocystis の酸性ストレス耐性に細胞外多糖が重要であることを明らかにした。また、 MsbA と高い相同性を持つ遺伝子変異株を構築し、LPS 変異株を得た。これらの変異株によ る解析の結果、酸性ストレス耐性だけでなく、いくつかの環境ストレス耐性への LPS の関 与を明らかにした。
1
第
1 章 研究背景
生物は常に周囲の環境からストレスを受けており、生存するために細胞内で様々な適応 機構を発達させている。周囲の環境から受けるストレスを環境ストレスと呼ぶが、偏に環境 ストレスといっても、その様態は温度、乾燥、塩や光など様々である。植物や微生物におい ては、可動範囲が狭いために、環境ストレスに如何に抵抗するかが、生存のカギとなる。例 えばシアノバクテリアでは、低温ストレスに対して膜脂質を構成する脂肪酸に不飽和結合 を導入して、低温により低下する膜脂質の流動性を維持することや(Los et al., 1997)、塩また は浸透圧ストレスに対して適合溶質である Sucrose や Glucosylglycerol(GG)を合成することなどが明らかにされている(Reed and Stewart, 1985)。近年では古くから知られている環境ス
トレスだけでなく、海洋のpH の低下(Dore et al., 2009)が問題となっている。 生態系を構成する生物集団は、生産者、消費者、分解者に分類される。生産者に位置する シアノバクテリアは好アルカリ性であることから、海洋におけるpH 低下が進行すると、生 態系に影響を及ぼす可能性が考えられる。また、シアノバクテリアの酸性ストレスへの応答 機構は、現在までにほとんど解明されていない。そこで本研究では、シアノバクテリアの酸 性ストレス応答機構の解明を主題とした。 本研究で用いたシアノバクテリアSynechocystis sp. PCC6803 (Synechocystis) は水を分解し 酸素を発生させる高等植物型の光合成を行うことから、酸素発生型光合成の祖先生物と考
2 えられている。また、一般的に高等植物の世代期間は数カ月から数年であるのに対して、 Synechocystis は数日の培養で実験に用いることが可能であるため(吉川と堀, 2009)、光合成 やストレス応答の研究でモデル生物として広く用いられている。 タンパク質発現にはタンパク質をコードする遺伝子を転写(Transcription)し、mRNA をリ ボソームによって翻訳(Translation)する過程を辿る(Fig. 1-1)。生物はストレスを感知すると、 そのストレスに抵抗するためにストレス応答タンパク質を発現する。タンパク質の発現量 を増加するためには転写の促進、翻訳の促進、タンパク質分解の抑制の 3 通りが考えられ る。 ストレスに応答する遺伝子を網羅的に解析するためには、mRNA またはタンパク質の量 を比較することになるが、実験の性質上タンパク質の定量よりも転写の定量の方が遥かに 容易に行うことが可能である。DNA マイクロアレイ解析もその一つである。 Synechocystis の酸性ストレス耐性における知見は、他にもいくつか報告されている。DNA マイクロアレイ解析による先行研究において、酸性ストレス下で約30 個の遺伝子が転写誘
導されることが明らかになった(Ohta et al., 2005)。脂質を輸送するトランスポーターSltA1、
SltB2、SltC2 の欠損株は酸性ストレス下で生育が抑制されることが明らかにされている
(Tahara et al., 2012)。また、レスポンスレギュレーターの SphR は、酸性ストレス耐性に関与
するslr0967 の転写を制御しており、slr0967 により転写を制御されている sll0939 もまた、
3
al., 2015)。さらに、site-2-protease のホモログである Slr0643 が酸性ストレス耐性に関与する ことも報告されている(Zhang et al., 2012)。
本研究では、DNA マイクロアレイ解析の結果(Ohta et al., 2005)をもとに、σ 因子などの
ストレス応答に関与することが報告されている遺伝子を除いて、先行研究ですでに機能が
明らかにされている、変化の大きかった遺伝子としてsll1558 に着目した。Kyoto Encyclopedia
of Genes and Genomes (KEGG:http://www.genome.jp/kegg/)では、sll1558 は Mannose-1-phosphate
guanyltransferase をコードする遺伝子として登録されていたが、近年、前田らによって
UDP-glucose pyrophosphorylase をコードすることが明らかになった(Maeda et al.,
2014)。UDP-glucose pyrophosphorylase によって合成される UDP-2014)。UDP-glucose (Fig.1-2)は糖タンパク質(Silbert et
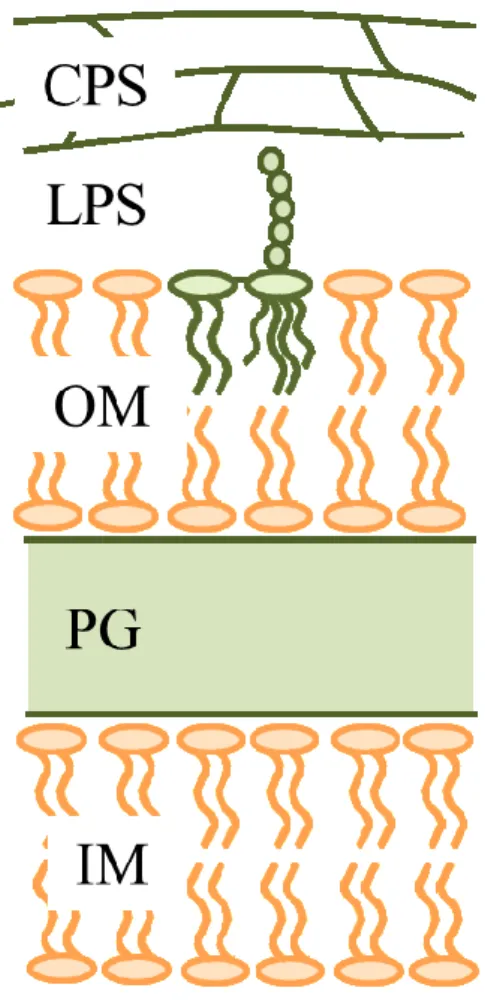
al., 1995; Maeda et al., 2014)、糖脂質(Sandhoff et al., 1992)合成に用いられているため、細胞表 層の構築に寄与している。Synechocystis を含むグラム陰性細菌の細胞表層は、細胞の内側か
らinner membrane (IM)、peptidoglycan (PG)、outer membrane (OM)、lipopolysaccharide (LPS)、
capsular polysaccharide (CPS)となっている(Fig.1-3)。大腸菌では LPS は抗生物質や環境スト
レスに抵抗するための手段の一つとされている(Raetz and Whitfield, 2002)。
LPS は内毒素活性を示す lipid A、core oligosaccharide、菌株特有の配列を持つ O-antigen か
ら構成されている。大腸菌ではLPS 自体が強い毒性を示すものもいる。さらに外側には、
細胞から遊離した released polysaccharide (RPS)や exopolysaccharide (EPS)が存在している。
4
1990)、細胞を保護している(Soule et al., 2016)。砂漠などの環境中においては、水や利用でき
る栄養素などが制限されているが、EPS はその環境中から不足している金属や栄養を捕獲
して、代謝を助ける(De Philippis et al., 2007)。
大腸菌の msbA は生存するために必須な遺伝子の一つであり、msbA 欠損株は単離できな
いことが報告されている(Karow and Georgopoulos, 1993)。MsbA は IM に存在し、lipidA を
inner leaflet から outer leaflet へ flip-flop する(Polissi and Georgopoulos, 1996) (Fig. 1-4)。外膜に
輸送されたLPS は lipid A 部を外膜にアンカーして、糖鎖部を膜の外側へ出す(Sperandeo et
al., 2008)。しかし、シアノバクテリアのLPS には強い毒性がないことや(Durai et al., 2015;
Fujii et al., 2012; Rapala et al., 2002; Stewart et al., 2006)、lipid A のアシル鎖が通常の 6 本では
なく4 本であることなどが報告されている(Durai et al., 2015)。 本研究では、細胞外多糖に着目してシアノバクテリアの酸性ストレス耐性を検討した。第 一に、細胞表層の構築に寄与するsll1558 株の解析により、Synechocystis の酸性ストレス耐 性への細胞外多糖の関与を明らかにした。第二に、MsbA と高い相同性を持つ Slr2019、Sl1276、 Sll1725、Slr1149 の遺伝子変異株を構築し、LPS 変異株を得た。これらの変異株による解析 の結果、酸性ストレス耐性だけでなく、いくつかの環境ストレス耐性への LPS の関与を明 らかにした。
5 Fig. 1-1 ストレスに応答する遺伝子の転写・翻訳 細胞はストレスを感知するとストレス応答タンパク質を発現する。青い矢印は通常時、赤 い矢印はストレス時の転写・翻訳のモデルである。ストレス条件下ではストレス耐性に関わ る遺伝子の転写量が増加してタンパク質合成を促進し、ストレスに対応する。 ストレス応答 タンパク質の増加
6 Fig. 1-2 Sll1558 が触媒する反応機構
UDP-glucose pyrophosphorylase は gulucose-1-phosphate に uridine triphosphate を付加すること
7 Fig. 1-3 グラム陰性菌の細胞表層構造
IM; inner membrane、PG; peptidoglycan、OM; outer membrane、LPS; lipopolysaccharide、CPS;
8 Fig. 1-4 LPS の合成・輸送モデル
Lipid A と polysaccharide 部分がそれぞれ合成されたものが、peptidoglycan を含む periplasm
9
第
2 章 材料と方法
2-1.培養
2-1-1 シアノバクテリアの培養
シアノバクテリアSynechocystis sp. PCC6803 は、10 mM TES-NaOH(pH 8.0) で緩衝した
BG-11 (Table 1) 液体培地に空気を通気させ、蛍光灯による光照射(光強度 25-40 μ mol photon
m-2 s-1)、30 °C の環境下で培養した。また、BG-11 寒天培地では終濃度 1.5%になるようにア ガロースを添加し、Na2S2O3を終濃度0.3%になるように加えた(Stanier et al., 1971)。形質転 換体は 20 μg/mL のカナマイシン(Km)またはクロラムフェニコール(Cm)を含む培地で培養 した。 2-1-2 各ストレス下での培養 酸性ストレス処理は10 mM MES-NaOH (pH 6.0) または 10 mM Glycine (pH 3.0)で緩衝した BG-11 で行った。高塩ストレスは 0.5 M NaCl を pH 8.0 BG-11 に、高浸透圧ストレスでは 0.25 M または 0.5 M Sorbitol を pH 8.0 BG-11 に添加した。温度ストレスは低温が 20 °C また は25 °C、高温ストレスは 40 °C で培養した。金属イオン欠乏または過剰ストレスには、培 地に含まれている II 価カチオンのマンガン, マグネシウム, カルシウム, 銅, 亜鉛, コバル
10
トについて10 倍過剰及び欠乏培地で行った。各金属イオン濃度は Table 2 に示した。
吸光度を測定後、サンプルの細胞量を一定に揃え1,1/10,1/100 に希釈、それぞれの条件の
寒天プレートにスポットし、1 週間培養した。
UV-B ストレスでは、吸光度を測定後サンプルの細胞量を一定に揃え、UV-B に 10 min ま
たは20 min 曝し、pH 8.0 BG-11 培地にスポットした。
2-1-3 大腸菌の培養
大腸菌の培養は、LB (10 mg/mL Tryptone, 5 mg/mL yeast extract, 10 mg/mL NaCl, 15 mg/mL
agar)液体培地で 37 °C の環境下で行った。また LB 寒天培地では終濃度 1.5%になるように
アガロースを添加した。形質転換体は100 μg/mL のアンピシリン、50 μg/mL の Km、または
11 2-2 欠損株の構築
各株の構築については、第3 章の各項にて記述した。欠損株の構築に必要な工程での操作
を以下に記述する。
12 Fig. 2-2 欠損株の構築方法 2
13 2-2-1 ゲノム DNA 抽出
以下の操作はバイオ実験イラストレイテッドに記載されているDNA 抽出法を一部改変し
て行った(中山ら, 1995)。培養した Synechocystis を 8000 rpm, 5 min, r.t (25 °C)で遠心して回収
し、上清を完全に取り除き、リゾチームを含むSTE Buffer (150 mM NaCl, 10 mM Tris-HCl, 10
mM EDTA)に懸濁した。1 h, r.t でインキュベートしたのち、懸濁液と等容の STE proteinase
solution(0.1%SDS、100 μg/mL proteinase K)を加えよく混和した。55 °C で 1 h インキュベート
し、その後37 °C で 1 h インキュベートした。PCI を等量加えて混和し、8000 rpm, 10 min, r.t
で遠心して上層を回収した。回収した上層に1/10 容の 3M 酢酸ナトリウムと 1 容の 2-プロ
パノールを加え、15000 rpm, 10 min, r.t で遠心した。上清を完全に取り除き、70%EtOH を 600
µL 加え、15000 rpm, 5 min, r.t で遠心した。上清を完全に取り除いて、乾燥させたのち、100
μl の TE Buffer (10 mM Tris-HCl, 1 mM EDTA)で沈殿を溶解させた。溶液に 1 µL の 1 µg/µL
RNase を加え、37 ºC で 30 min 処理した。PCI を 100 μl 加え、15000 rpm, 10 min, 4 °C で遠心
して上層を回収し、1/10 容の 3M 酢酸ナトリウムと 2.5 容の 100% EtOH を加えた。-20 °C
でインキュベートした後、15000 rpm, 15 min, 4 °C で遠心して沈殿物を回収した。上清を完
全に取り除いたのち、70%EtOH を 600 µL 加え、15000 rpm, 5 min, 4 °C で遠心した。上清を
14 2-2-2 PCR
抽出したゲノムDNA を Takara Gflex DNA Polymerase (Takara Bio Inc., Shiga, Japan)のプロ
トコルに従って溶液調製した後、 95 °C 10 sec, 60 °C 15 sec, 68 °C 1 min, 30cycle で PCR 反応
を行った。使用したプライマーはTable 3 に示した。
2-2-3 制限酵素処理
PCR 産物は目的の制限酵素及び適した Buffer を用い、付属のプロトコルに従って一晩イ
ンキュベートして反応させた。
2-2-4 アガロースゲル電気泳動
アガロースを1%になるように入れた TAE (40 mM Tris, 20 mM CH3COOH, 1 mM EDTA)を
電子レンジで完全に溶解させ、しばらく冷ました後、5 mg/mL エチジウムブロマイドを少量
加えて固化させたゲルにサンプルを注入し、100V で 30 min 泳動した。泳動バッファーには
TAE を用いた。泳動終了後は、LAS-4000 mini (Fuji film, Tokyo, Japan)を用いてバンドを確認
した。
2-2-5 DNA 精製
15
PCR Clean-Up System(Promega corporation, WI, USA)を用いて精製した。
2-2-6 Ligation
制限酵素で処理し、ゲル精製したサンプルをDNA Ligation Kit Ver.2 (Takara Bio Inc., Shiga,
Japan)を用いて、付属のプロトコルに従って結合させた。反応は 25 °C、5 min で行った。
2-2-7 コンピテントセル調製
大腸菌 JM109(endA1 hsdR17 gyrA96 Δ(lac proA) recA1 relA supE44 thi F’ (lacIq lacZΔM15
proAB+ traD36) (Yanisch-Perron et al., 1985)を一晩培養した。500 mL の LB 培地に前培養した
大腸菌を植菌し、初期の吸光度(OD600)を測定後、吸光度が 0.4-0.7 になるまで培養した。氷
冷した後、4500 rpm, 4 °C, 10 min 遠心して集菌した。10% Glycerol で 3 回 Wash し培地を完
全に除いた。上清を完全に取り除いたのち、3 mL の 10% Glycerol を加え 50μl ずつ分注し
た。液体窒素につけ急冷させた後、-80 °C で保存した。
2-2-8 形質転換
エレクトロポレーション法により大腸菌JM109 を形質転換した。装置は MicroPulser エレ
クトロポーレータ(BIO-RAD, CA, US)を使用し、プログラムされている Ec1(1.80 kV, 1 pulse)
16 2-2-9 プラスミドの精製
プラスミドの精製はZhou らのアルカリ-SDS 法を一部改変して行った(Zhou et al., 1990)。
50 mL コニカルチューブに抗生物質を含む LB 液体培地で、形質転換した大腸菌を 1 コロニ
ー植菌して37 °C, over night で大量培養した。培養液を 6000 rpm, 5 min, r.t で遠心し、集菌し
た。上清を完全に取り除き、TENS(10 mM Tris-HCl, 1 mM EDTA, 0.1N NaOH, 0.5% SDS)を 300
μl 加えて穏やかに混合した。r.t で 5 min インキュベートし、3M 酢酸ナトリウムを 150 μl 加
えて転倒混和した。15000 rpm, 5 min, r.t で遠心し、上清を回収した。1 容の 2-プロパノール
を加え混合し、15000 rpm, 5 min, r.t で遠心した。上清を除いて乾燥させ、100 μl の TE Buffer
を加えて沈殿を溶解させた。その後、1 µL の 1 µg/µL RNase で 30 min 処理した。PCI を 100
μl 加え、15000 rpm, 10 min, 4 °C で遠心した。上層を回収し、1/10 容の 3M 酢酸ナトリウム と2.5 容の 100% EtOH を加え、-20 °C でインキュベートした後、15000 rpm, 15 min, 4 °C で 遠心した。上清を完全に取り除き、70% EtOH を 600 µL 加え、15000 rpm, 5 min, 4 °C で遠心 した。上清を完全に取り除き、乾燥させた。50 μL の TE Buffer で溶解した。 2-2-10 相同組換え 相同組換えはWilliams のプロトコルに従って行った(Williams, 1988)。培養している野生株
のSynechocystis を 8000 rpm, 5 min, r.t で遠心し、集菌した。上清を取り除き、10 mM NaCl で
17
懸濁した。懸濁液を 1.5 mL エッペンチューブに移し、回収したプラスミドを加え、30 °C,
over night で rotate させた。その後、6500 rpm, 10 min, r.t で遠心し、上清を 700 μl 取り除い
た。残った液に沈殿を懸濁し、抗生物質を含む BG-11 プレートにまいた。出てきたコロニ
18 2-3 細胞外多糖の分析
19 2-3-1 RPS 抽出 液体培地で生育させたSynechocystis を、遠心分離(8000 rpm,5 min,25 °C)により集菌し、上 清400 µL を回収した。100%EtOH を 1600 µL 加え、-20 °C で一晩インキュベートし、(15000 rpm, 7 min, 0 °C)で遠心した。上清を完全に取り除いて乾燥し、dH2O 120µL で溶解した。 2-3-2 CPS 抽出 液体培地で生育させたSynechocystis を、遠心分離(8000 rpm,5 min,25 °C)により集菌し、上 清を除いた。沈殿した菌体に0.05% NaBH4 aq 120 を加えよく懸濁し、100 °C で 1 h インキ ュベートしたのち、14000 rpm, 5 min, 25 °C で遠心し、上清を回収した。 2-3-3 LPS 抽出 CPS 抽出で NaBH4処理した後の遠心後、沈殿を回収した。沈殿に対し、脱イオン水と90% Phenol を体積比 1:1 になるように加え 65 °C 放置したのち、遠心し(14000 rpm, 15 min, 25 °C)、 上清を回収した。これを生成するために、1/10 容の酢酸ナトリウムと 2 容の 100%EtOH を 加え-20 °C に 20 min 置いた後に遠心し、沈殿を回収した。沈殿に 70 %EtOH を加え、遠心 後、乾燥させた。得られた沈殿に対し脱イオン水を加えLPS 粗抽出画分とした。 LPS 租抽出画分に 20 mg/mL Proteinase K 1.0 µL と 10%SDS を加え、50 °C で一晩インキュベ ートした。インキュベートしたのちPCI を加えて激しく撹拌し、15000 rpm, 15 min, 0 °C で
20 遠心し上清を回収した。上清に1/10 容の酢酸ナトリウムと 1 容の 2-Propanol を加え-20 °C に20 min 置いた後に遠心し、沈殿を回収した。沈殿に 70%EtOH を加え、遠心後、乾燥させ た。得られた沈殿に対し脱イオン水を加えLPS 画分とした。 2-3-4 糖定量 糖定量には検量線法を用いた。100.00、50.00、25.00、12.50、6.25 µg/mL の各濃度のグル コース水溶液を調製し、脱イオン水、5 % Phenol を加え軽く撹拌したのち、濃硫酸を加え速 やかによく撹拌した。これらを放冷後OD490を測定し検量線を作成した。 抽出したLPS 画分及びブランク(蒸留水)に蒸留水、5 % Phenol を加え軽く撹拌したのち、濃
硫酸を加え速やかによく撹拌した。これらを放冷後、iMark Microplate Reader (BIO-RAD, CA,
US) によって OD490を測定し、糖の定量を行った。
2-3-5 タンパク質定量
タンパク質定量には検量線法を用いた。20.00、15.00、10.00、5.00、2.00、1.00 µg/mL の濃
度のBSA 水溶液を調製し、Quick Start Bradford Dye Reagent (BIO-RAD, CA, US)を加えよく
撹拌した。OD595を測定し検量線を作成した。
抽出したLPS 画分のタンパク質量の定量を行った。各サンプル及びブランク(脱イオン水)
21
た。これらのOD595を測定しタンパク質の定量を行った。
2-3-6 SDS-PAGE
糖量を揃えたLPS 画分 10 µL と 5×Sample Buffer (Table 4) 10 µL を混合し、SDS-PAGE を
行った(Table 5 および Table 6)。泳動は 40 mA で行い、dye front がゲルの下端に移動するま
で行った。泳動にはPAGERUN(ATTO, Tokyo, Japan) を用いた。
2-3-7 SDOC-PAGE
Rehuhs らの Deoxycholic acid を用いた方法を一部改編して行った(Rehuhs et al., 1998 )。糖
量を揃えたLPS 画分 10 µL と 2×Sample Buffer(Table 7) 10 µL を混合し、SDOC -PAGE を行
った(Table 8 および Table 9)。泳動は 40 mA で行い、dye front がゲルの下端に移動するまで
行った。泳動にはPAGERUN(ATTO, Tokyo, Japan) を用いた。
2-3-8 過ヨウ素酸処理
糖を開裂させ銀を結合させるために、過ヨウ素酸処理を行った。泳動後のゲルを脱イオン
水で軽く洗浄し、30% EtOH/ 10% 酢酸混合溶液 80 mL を加え 30 min 間振盪した。その後溶
液を捨て、30% EtOH / 10% AcOH/ 0.7% NaIO4 (w/v) aq 80 mL を加え 10 min 間振盪した。そ
22 り返し、ゲル表面に付着した溶液を除去した。
2-3-9 銀染色
泳動したゲルを過ヨウ素酸処理したのち、銀染色試薬(Cosmo Bio, Co., Ltd., Tokyo, Japan)
のプロトコルに従い染色した。平らな容器にゲルを入れ、Pretreatment solution 80 mL を加え
10 min 間振盪した。試薬を捨て、Silver staining solution 80 mL を加え 25 min 間振盪した。試
薬を捨てた後、脱イオン水80 mL で 2 min 間振盪しゲル表面を洗浄した。この洗浄を 3 度繰
り返し、表面のSilver staining solution を除去した。洗浄後、Developer 160 mL を加え適当な
像が得られるまでよく振盪した。適度な像が得られたら速やかにDeveloper を捨て、脱イオ
23 2-4 転写解析
24 2-4-1 酸性処理細胞からの total RNA の抽出
液体培地で培養している野生株を遠心分離 (8000 rpm, 5 min, 25 °C) で集菌し、上清を除
いたのち、回収した細胞をTES-NaOH を除いた BG-11 培地を加え再度遠心し、洗浄した。
この洗浄操作を2 回行い、完全に緩衝能を除いたのち、pH 3.0 の液体培地に植菌して酸性ス
トレスを4 h 与えた。Total RNA は Pinto らのプロトコルに従って抽出した(Pint et al., 2009)。
また、実験に使用する器具等はオートクレーブにより滅菌し、RNase free にした。試薬は
RNase free で調製し、RNase free water (0.1% (v/v) diethl pyrocarbonate)を使用して作成した。
酸性ストレスを与えた細胞を8000 rpm, 5 min, r.t で遠心し回収した。回収したサンプルに 1
mL PGTX (3.92 M phenol, 8.72% (w/v) glycerol, 6.89 mM 8-hydroxyquinoline, 15.58 mM EDTA,
97.56 mM sodium acetate, 0.80 M guanidine thiocyanate, 99.45 mM guanidine hydrochloride, and
2.14% (w/v) Triton X-100)を加え、懸濁した。95 °C で 5 min インキュベートした。すぐに氷
上で5 min インキュベートし、ブロモクロロプロパンを 100 μl 加え、懸濁した。r.t で 15 min
インキュベートした後、15000 rpm, 15 min , 4 °C 遠心した。上清を回収し、2-プロパノール
を500 μl 加え r.t で 10 min インキュベートした後、15000 rpm, 10 min , 4 °C で遠心した。上
清を取り除き70% EtOH を 1 mL 加え、15000 rpm, 5 min , 4 °C で遠心した。上清を取り除き、
乾燥させた。乾燥後、RNase free water を 176 μl 加え、沈殿を溶解させた。DNase を加え 37 °C
で2 h インキュベートした後、PCI (Phenol:Chlorofolm: Isoamylalcohol=25:24:1)を 200 μl 加え、
25
の100% EtOH を加え、-20 °C でインキュベートした後、15000 rpm, 15 min, 4 °C で遠心した。
上清を完全に取り除き、70%EtOH を 1 mL 加え、15000 rpm, 5 min, 4 °C で遠心した。上清を
完全に取り除き、乾燥させた。沈殿を100 μl の RNase free water で溶解した。
2-4-2 逆転写反応
抽出したtotal RNA の量が同じになるように液量を計算し、全量 7 μL になるように dH2O
を加えた。PrimeScript™ RT reagent Kit (Perfect Real Time) (Takara Bio Inc., Shiga, Japan) の
5×PrimeScript Buffer (for real Time)を加え、終濃度 25 pmol の Random 6 mers を加えた。さら
に、PrimeScript RT Enzyme Mix I を 0.5 μl 加えた。すべての操作は氷上で行った。混合した
溶液を37 °C, 15min 処理した後、85 °C, 5 sec で処理した。
2-4-3 リアルタイム RT-PCR
逆転写反応により合成されたcDNA 1μl を鋳型にし、終濃度 10 pmol のプライマー (Table
3)と、Fast SYBR Green Premix Ex Taq (Perfect Real Time) (Takara Bio Inc., Shiga, Japan)を用い
て10 μL に Fill up 後 95 °C 20 sec で 1 cycle 反応させた後、95 °C 30 sec, 60 °C 30 sec を 40
cycle 行った。蛍光測定は 1cycle ごとに行った。反応中の DNA 増幅量の変化は蛍光検知に
よりリアルタイムに追跡し、そのPCR 曲線をモニタリングした。得られた Ct 値を内在性コ
26
たりの量に揃えた値をΔΔCt 値とした。PCR は 1 反応ごとに 2 倍になることを用いて、発現
量の比較は、2(-ΔΔCt)の式で算出した。
2-4-4 タンパク質の抽出
野生株の酸性処理は2-17.と同様の方法を用いた。pH 3.0 に 4 h 曝した野生株を遠心分離
(8000 rpm, 5 min, 25 °C) で集菌して上清を除き、Lysis Buffer(20 mM Tris-HCl (pH 8.0), 150 mM
NaCl)を 1 mL 加えて懸濁した。懸濁液を遠心し(15000 rpm, 5 min, 25 °C)、上清を完全に取り
除いた。沈殿に Lysis Buffer 200 µL を加えて懸濁し、ビーズを加えて FP101A Fast Prep
(Thermo)で 3 回細胞破砕を行った。その後、3300 rpm, 5 min, 25 °C で遠心し、上清を回収し、
さらにビーズを完全に取り除くために6000 rpm, 5 min, 25 °C で遠心して上清を回収した。
溶液の吸光度を測定し濃度をそろえた。
濃度調整したサンプル12 µL に 5×Sample Buffer(Table 4) 3 µL を加え 95 °C で 5min インキ
ュベートした。
2-4-5 メンブレンへの転写
切り出したメンブレンをメタノールで親水化処理した後、転写Buffer(12 mM Tris-HCl(pH
8.0), 9.8 mM Glycine, 1/5 容 MeOH)に 30 min 程浸した。半乾式法を用いて 10V, 30 min でゲル
27
ドライブロッティング装置(BIO-RAD, CA, USA)を用いた
2-4-6 抗原抗体反応
転写したメンブレンをブロッキング剤(Table 10)に浸し、冷所で一晩インキュベートした。
ブロッキングしたメンブレンを一次抗体に浸し、r.t で 1.5 h、一次抗体反応を行った。一次
抗体にはanti-Slr2019 from rabbit を 5%スキムミルク溶液で 1/60000 に希釈した。一次抗体反
応後、TBS-T で 5 min 洗浄し、さらに 15 min の洗浄を 2 回行った。メンブレンの洗浄後、
r.t で 1 h、二次抗体反応を行った。二次抗体は ECL Anti-rabit IgG, Horseradish Peroxidase Linked
Whole Antibody from donkey (GE Healthcare, Buckinghamshire, UK)を 5%スキムミルク溶液で
1/5000 に希釈した。二次抗体反応後、上述したようにメンブレンの洗浄を行った。
2-4-7 検出
ECL Advance Western Blotting Detection Kit, SolutionA 200 µL と SolutionB 200 µL を混合し、
メンブレンの転写面にかけた。メンブレンを37℃で 10 min インキュベートしたのち、溶液
を完全に除いた。LAS-4000UVmini (Fuji film, Tokyo, Japan).を用いて 150 min 露光した。
2-4-8 抗体の作製
抗体の作製はオペロン バイオテクノロジー株式会社に依頼し、Slr2019 の配列に対して特
28
ド候補からポイントの高い C+NLAQDIQHDLRLDA この配列を選択し、ペプチド合成を依
頼した。合成後、rabbit に合成ペプチドを注射し、49 日後に採血し、抗血清(anti-Slr2019 from
rabbit)として用いた。
2-5 吸光度測定
集めた細胞の濁度を分光光度計(V-630Bio; JASCO, Tokyo, Japan)で測定した。固定波長(490
nm, 595 nm, 600 nm, 730 nm)で測定した。
2-6 ICP-AES
pH 8.0 および pH 6.0 で培養した Synechocystis 500 ml を遠心分離 (8000 rpm, 5 min, 25 °C)
で集菌し、乾燥器で1 週間乾燥させた後、乾燥重量を測定した。13.5 M HNO3(有害金属測定
用)と 30% H2O2(原子吸光分析用)を用いて湿式灰化を行った後、容量分析用 1 M HCl 10 mL
を加えて超音波洗浄機により完全に溶解した。サンプルはNo.5C のろ紙(Toyo Roshi Kaisha,
Ltd., Tokyo, Japan)を用いて濾過した。ICP-AES の測定は Optima 5300 DV(PerkinElmer Inc., MA,
USA)で行った。ICP 分析用多元素混合標準液 W-I、W-II 、W-V(Wako Pure Chemical Industries,
29 2-7 透過型電子顕微鏡(TEM)による解析 細胞形態及び細胞表層の厚さを計測するため、TEM による解析を三重大学電子顕微鏡室 の小川覚先生に依頼し、解析を行った。脱水操作の90%EtOH 洗浄までは 4 ℃環境で操作し た。 サンプル50 mL 回収後、3%グルタルアルデヒドに一晩つけて前固定し、リン酸緩衝液で 10min 洗浄を 3 回行った。その後、2%四酸化オスミウムに 3 h つけて後固定し、リン酸緩 衝液で15 min 洗浄を 2 回行った。洗浄後、30%、50%、70%、80%、90%に希釈した EtOH で 10 min、2 回かけて徐々に脱水した。その後、樹脂とアセトンを 1:1 に混合した混合液に 1 晩浸透し、100%樹脂に交換し 60 min 浸透し、新しい 100%樹脂に交換し 90 min 振盪を 3 回 行った。平板包理板に試料と樹脂を添加した後、r.t で 12 h、40℃で 12 、60℃で 48 h かけ て重合させたものをサンプルとした。そのサンプルを透過型電子顕微鏡を用いて観察した。 2-8 Nile Blue A 染色
Tsang らの方法に従って行った(Tsang et al., 2013)。液体培地で 7 日間培養した野生株と
slr2019 株 1.5 mL を遠心分離(16000 g, 5 min, 25 °C)により集菌し、上清を除いた。そこに 1% ホルムアルデヒドBG-11 溶液(TES なし) 200 µL を加え、懸濁した。r.t で 20 min インキュベ
ートした後、16000 g, 5 min, 25 °C で遠心分離して上清を完全に除いた。500 µL の BG-11(TES
BG-30
11(TES なし) 200 µL で懸濁し、この溶液 99 µL と DMSO を溶媒に用いた 0.001% Nile Blue
A 溶液で 30 min 染色し、観察した。観察にはシステム顕微鏡 BX53 (Olympus Corporation,
Tokyo, Japan)を、蛍光フィルタには U-FBNA(励起フィルタ;470-495 nm,
吸収フィルタ;510-550 nm, ダイクロイックミラー;505 nm)を用いた。
2-9 アライメント
目的遺伝子の配列は、NCBI (http://www.ncbi.nlm.nih.gov/) から入手した。アライメントは、
31
BG-11
Final conc.
100×BG-11
NaNO
317.60 mM
MgSO
4・7H
2O
0.31 mM
CaCl
2・
2H
2O
0.25 mM
citric acid
0.031 mM
Na
2EDTA
2.8 μM
Trace Metal Mix
H
3BO
346.26 μM
MnCl
2・4H
2O
9.10 μM
ZnSO
4・7H
2O
0.765 μM
Na
2MoO
4・2H
2O
1.61 μM
CuSO
4・
5H
2O
0.316 μM
Co(NO
3)
2・
6H
2O
0.170 μM
TES-NaOH
10 mM
Ferric ammonium citrate
6 mg/l
Na
2CO
3189 μM
K
2HPO
4175 μM
Table 1 Synechocystis sp. PCC6803の生育培地 (pH8.0)
濃度
Mg
2+3.1 mM
Ca
2+2.5 mM
Mn
2+910 μM
Zn
2+7.7 μM
Cu
2+3.2 μM
Co
2+1.7 μM
Table 2 金属イオンの濃度
32 P rim er pa ir P rim er s equ en ce (5' → 3' ) O bj ec t R ef er en ce sl l155 8 1 C C G A A T T C G G C C A T T G A A A A A C C A G T G G for c on st ra ct of m ut an t T hi s w or k sl l155 8 2 C C G G A T C C G G C T T T C A T C A A C A A T T T T C for c on st ra ct of m ut an t T hi s w or k sl l155 8 3 C C G G A T C C T C C A G C C G G A A T A A T C A C A G for c on st ra ct of m ut an t T hi s w or k sl l155 8 4 C C C T G C A G T G T T T G C C C T G G G C A T T A A G for c on st ra ct of m ut an t T hi s w or k sl r201 9 F G A G C T C A C A G T G A T G C C T for c on st ra ct of m ut an t T hi s w or k sl r201 9 R A A G C T T T G A G C C A G T A G G C T for c on st ra ct of m ut an t T hi s w or k K pn I-C m -F G G T A C C T T C A G G C G T A G C A C C A G for c on st ra ct of m ut an t T hi s w or k K pn I-C m -R G G T A C C T C G G C A C G T A A G A G G T T C for c on st ra ct of m ut an t T hi s w or k ch ec k-sl r201 9 F T A G C C A G G T G C A A G G for c on st ra ct of m ut an t T hi s w or k ch ec k-sl r201 9 R T T T C T T C A G A T C C A C for c on st ra ct of m ut an t T hi s w or k sl l127 6 u p-f C C T T C C C A A G C G G A A G C C A G T C G C A for c on st ra ct of m ut an t T hi s w or k sl l127 6 L f-re v- up-r A C A T C A G A G A T T T T G A G A C A C A A C G T G G C T C A A C C A G G C T A G A A T G G G C for c on st ra ct of m ut an t T hi s w or k sl l127 6 R r-re v-dn -f C A C C A A C T G G T C C A C C T A C A A C A A A G C T C T G C T A G A A A A T A T T G C C A A T for c on st ra ct of m ut an t T hi s w or k sl l127 6 dn -r A A C A G T G G T G G T T C T C C A A T A A C C G for c on st ra ct of m ut an t T hi s w or k sl l172 5 u p-f C C A A A G G A G A T T T A G C A A T G C G G G A for c on st ra ct of m ut an t T hi s w or k sl l172 5 L f-re v- up-r A C A T C A G A G A T T T T G A G A C A C A A C G T G G C T T T A T C T A T C A C T T T T A A C T for c on st ra ct of m ut an t T hi s w or k sl l172 5 R r-re v-dn -f C A C C A A C T G G T C C A C C T A C A A C A A A G C T C T T G A T G G C C G A A A T T T T C T C for c on st ra ct of m ut an t T hi s w or k sl l172 5 dn -r T T A C C C C A T T G C T A A T T T G C T T C C A for c on st ra ct of m ut an t T hi s w or k sl r114 9 u p-f A T T A A C A G T T A A A T T G G C C C A A A G A for c on st ra ct of m ut an t T hi s w or k sl r114 9 L f-re v- up-r A C A T C A G A G A T T T T G A G A C A C A A C G T G G C T GGAGAAAGAAAT GAT T GAC for c on st ra ct of m ut an t T hi s w or k sl r114 9 R r-re v-dn -f C A C C A A C T G G T C C A C C T A C A A C A A A G C T C T T T A T C C T A C C G G G A A T C T C for c on st ra ct of m ut an t T hi s w or k sl r114 9 dn -r A G A T G A C T A C A T C A G C G G T G C C C C G for c on st ra ct of m ut an t T hi s w or k 4K -L f A G C C A C G T T G T G T C T C A A A A T C T C T G A T G T for c on st ra ct of m ut an t S at o, 2 009 4K -L r G A G A A A T C A C C A T G A G T G A C G A C T G A A T C C for c on st ra ct of m ut an t S at o, 2 009 4K -R f A A G C T T T T G C C A T T C T C A C C G G A T T C A G T C for c on st ra ct of m ut an t S at o, 2 009 4K -R r A G A G C T T T G T T G T A G G T G G A C C A G T T G G T G for c on st ra ct of m ut an t S at o, 2 009 R T -r npB F G G A G T T G C G G A T T C C T G T C A for qR T -P C R U ch iy am a et al ., 2012 R T -r npB R A C T G T T T A C T G G T T G C T G T T T T C T A for qR T -P C R U ch iy am a e t al ., 2012 sl l155 8-R T F C G G A C G A C A A C G G C A A A for qR T -P C R T hi s w or k sl l155 8-R T R A T C C C C T C C C A G G T C G for qR T -P C R T hi s w or k R T -P C R sl r201 9 F A C G G G G C G G C T G A T G for qR T -P C R T hi s w or k R T -P C R sl r201 9 R C A A A A A A T G C T C C A C C A A T C A C for qR T -P C R T hi s w or k sl l127 6 Fw A T T G C C A T T G T G C C C C for qR T -P C R T hi s w or k sl l127 6 Rv C C C G T T C C C C C A C A T A G G for qR T -P C R T hi s w or k sl l172 5 Fw G C T G G T A G C G A T G G T G A C T T T for qR T -P C R T hi s w or k sl l172 5 Rv C A A C A G C C A A G C G G C A T T for qR T -P C R T hi s w or k sl r114 9 Fw A A T C G G G C T T T A C G G C for qR T -P C R T hi s w or k sl r114 9 Rv G G G T T T G A T G G T G A A G T T for qR T -P C R T hi s w or k T abl e 3 本研究で 用い た プ ラ イ マ ー
33
Tris-HCl(pH6.8)
0.303 M
SDS
10%
β-mercaptoethanol
10%
5M Sucrose
10%
BPB
少量
Table 4 5×Sample Bufferの組成
Separate-Gel (10%) ml Stacking-Gel (4%) ml
30% アクリルアミド溶液
4.29
0.65
1.5 M Tris-HCl(pH8.8)
3.25
―
0.5 M Tris-HCl(pH6.8)
―
1.25
10% SDS
0.13
0.05
dH
2O
5.42
3.05
10% APS
0.07
0.04
TEMED
0.01
0.008
合計
13 ml
5 ml
Table 5 SDS-PAGEのゲル組成
Tris
25 mM
SDS
0.35 mM
Glycine
190 mM
34
Tris-HCl (pH6.8)
0.635 M
BPB
149 nM
Glycerol
10%
Table 7 2×Sample Bufferの組成
Separate-Gel (18%) ml Stacking-Gel (4%) ml
30% アクリルアミド溶液
7.8
0.66
1.5 M Tris-HCl(pH8.8)
3.12
―
0.635 M Tris-HCl(pH6.8)
―
1.0
dH
2O
2.08
3.34
10% APS
0.05
0.025
TEMED
0.01
0.005
合計
13 ml
5 ml
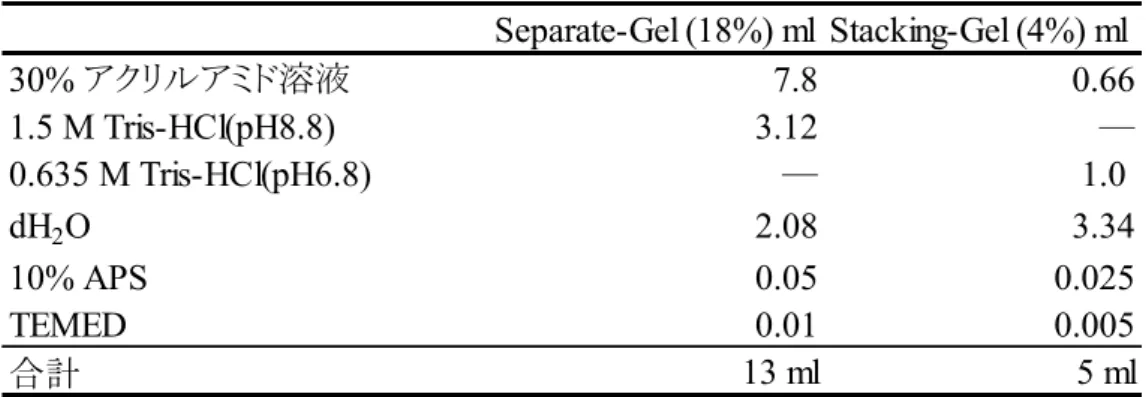
Table 8 SDOC-PAGEのゲル組成
Tris
37 mM
Sodium Deoxycholate
6.4 mM
Glycine
290 mM
Table 9 SDOC-PAGE running buffer 組成
スキムミルク
0.2 g
TBS-T
10 ml
TBS-T
Tris
20 mM
NaCl
0.14 M
Tween20
0.00%
Table 10 ブロッキング剤の組成
35
第
3 章 結果
3-1-1 sll1558 の転写解析 以前に、当研究室で行われた酸性ストレス下での網羅的な転写解析の結果、転写が促進さ れた遺伝子を約30 個同定した(Ohta et al., 2005)。これらのうち、σ 因子などのストレス応答 に関与することが報告されている遺伝子を除いて、先行研究ですでに機能が明らかにされている、変化の大きかった遺伝子としてsll1558 に着目した。Kyoto Encyclopedia of Genes and
Genomes (KEGG:http://www.genome.jp/kegg/) で は 、 sll1558 は Mannose-1-phosphate
guanylyltransferase をコードする遺伝子として登録されていたが、近年、前田らによって
UDP-glucose pyrophosphorylase をコードすることが明らかになった (Maeda et al., 2014)。
UDP-glucose pyrophosphorylase は gulucose-1-phosphate に UTP を付加することによって
UDP-glucose を合成する酵素である(Fig. 1-2)。UDP-glucose は Cellulose (Römling, 2002)や
Glycogen (Alonso et al., 1995)、Galactose 代謝(Holden et al., 2003)の生合成、さらに heteroglycan
や糖タンパク質(Silbert et al., 1995; Maeda et al., 2014)、糖脂質(Sandhoff et al., 1992)における
グルコース部分の付加に用いられている。これらは、細胞表層を構築するために必要な生理
物質である。
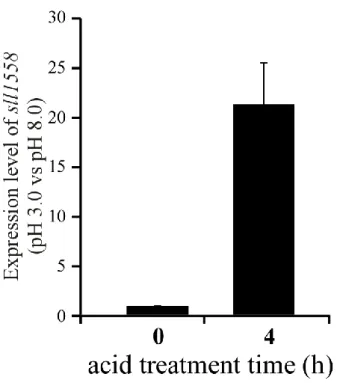
36 は4 時間の酸処理により約 20 倍転写量が増加した(Fig. 3-1)。この結果から、sll1558 は酸性 ストレス耐性に関与することが示唆された。 Mitschke らによる RNA-Seq 解析によると、sll1558 は低温(15 ºC)や鉄、リン酸欠乏ストレ スなど、多くのストレス下において転写量が増加することが報告されている(Mitschke et al., 2011)。さらに、UV-B 照射によって、sll1558 の転写が促進されることも先行研究で明らか にされている(Huang et al., 2002)。これらの結果から sll1558 は酸性ストレスだけではなく、 様々なストレス耐性に関与することが示唆されている。
37
Fig. 3-1 qRT-PCR による酸性ストレス下での sll1558 の転写解析
対数増殖期まで培養したSynechocystis を pH 3.0 で 4 時間処理した。縦軸は 0 h における
転写量を基準とした、相対転写量を示す。独立に3 回実験を行い、その標準偏差を SD とし
38 3-1-2 sll1558 株の構築
Synechocystis における酸性ストレス耐性と sll1558 との関係を明らかにするために、sll1558 をKmrカセットで置換したsll1558 欠損株の構築を試みた(Fig. 3-2)。
使用したプライマーをTable 3 に記載した。sll1558 の上流 400 bp と下流 600 bp を PCR で
増幅し、上流断片はEcoRI、 BamHI で処理し、下流断片は BamHI, HindIII で処理した。EcoRI,
BamHI で処理した高コピープラスミドである pUC19 に上流断片を Ligation により挿入し、 JM109 に形質転換して pUP1558 を得た。pUP1558 を BamHI, HindIII で処理して下流断片を
挿入し、JM109 に形質転換して pUP1558DN を得た。さらに pUP1558DN を BamHI で処理
し、pUC4K から切り出した Kmrカセットを挿入、形質転換してp1558 を得た。Synechocystis
とp1558 を相同組換し、20 µg/ml の Km を含む寒天培地で選別した。
先行 研究で Synechocystis はゲノムを複数保持していることが明らかにされている
(Labarre et al., 1989)。また、UDP-glucose pyrophosphorylase はシアノバクテリアに広く保存さ
れており、UDP-glucose の供給に必須であることから、生育や生存に必須の酵素であること
が一般的に知られている(Maeda et al., 2014)。
そこで、すべてのゲノムのsll1558 が Kmrカセットで置換されているかを確認するために、
野生株とsll1558 株からゲノムを抽出し、PCR を行った(Fig. 3-3,4)。
39 ころ、野生株では1 本のバンドが、sll1558 株では 2 本のバンドが確認できた(Fig. 3-4A)。Fig. 3-3A, 3-4A で示すように相同組換領域を含む sll1558 は 2167 bp であり、Kmrが挿入された 時の2234 bp よりも短い。さらに、sll1558 株には野生株と同じ 2167 bp にバンドが存在する ことから、sll1558 株のゲノムには sll1558 と Kmrの両方のパターンが存在することが明らか になった。 次に、sll1558 遺伝子内にアニールするプライマーを用いてそれぞれのゲノムを鋳型とし てPCR を行った(Fig. 3-3B)。野生株と sll1558 株の両方に約 200 bp のバンドが確認でき (Fig. 3-4B)、sll1558 株に sll1558 が残留していることが示された。最後に、sll1558 の上流領域と Kmr カセットまたは Kmr カセットと sll1558 下流領域にアニールするプライマーを用いて PCR を行った(Fig. 3-3C, D)。どちらも sll1558 株にのみバンドが確認できた(Fig. 3-4C, D)。 これらの結果からsll1558 株の少なくとも一部は Kmrカセットが置換されていることが確認 できた。 以上の結果より、sll1558 株は、sll1558 遺伝子を完全に欠損しておらず、sll1558 遺伝子は 完全欠損できないことが示唆された。そのため、UDP-glucose の供給に必要な UDP-glucose pyrophosphorylase は、必須の酵素であり、その酵素をコードする slll1558 も生存に必須な遺 伝子であると考えられる。 遺伝子を完全に欠損させることができなくても、複数あるゲノムのうちの一部が Kmrカ セットに置き換わることで、鋳型となるsll1558 が減少し、転写される mRNA の総量も減少
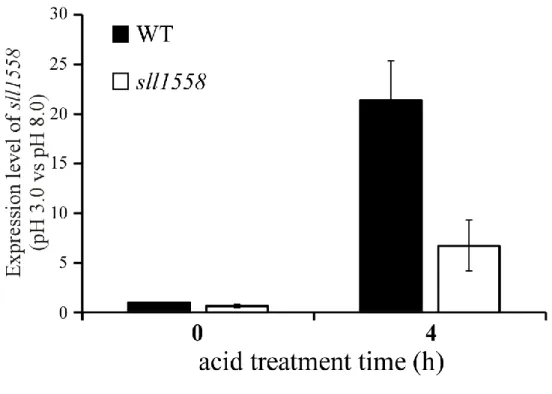
40 する。そのため、ストレス感受性を示すこともある。そこで、sll1558 株における sll1558 の mRNA の転写量を qRT-PCR によって測定した。 野生株とsll1558 株に pH 3.0 で 4 時間ストレスを与え、転写量を比較した(Fig. 3-5)。スト レスを与えていない0 h では野生株と変異株の転写量にほとんど差が見られなかった。しか し、4 h では野生株の転写量が約 20 倍に増加しているにも関わらず、sll1558 株は転写量が 5 倍程度であった。この転写量の違いが sll1558 株の酸性ストレスへの感受性の違いを引き 起こす可能性が考えられる。
41 Fig. 3-2 p1558 の構築方法
42 Fig. 3-3 sll1558 株の確認方法
野生株およびsll1558 株の欠損確認におけるプライマーのアニール部位を図示した。図中
43 Fig. 3-4 sll1558 株の確認
sll1558 が欠損していることを確認するために PCR を行った。アガロースゲル電気泳動後、 エチジウムブロマイドで染色し、UV で発色させた。(A)-(D)は Fig. 3-3 に図示したプライマ
ーセット(A)-(D)に対応している。Marker にはそれぞれ、A;λ-BstPI, B-D ;Gene Ladder 100 を
44
Fig. 3-5 qRT-PCR による野生株および sll1558 株における sll1558 の転写解析
対数増殖期まで培養したSynechocystis を pH 3.0 で 4 時間処理した。縦軸は 0 h における
発現量を基準とした、相対発現量を示す。Black bar は野生株、White bar は sll1558 株を示す。
45 3-1-3 酸性ストレス下での生育
Sll1558 の転写上昇と酸性ストレス耐性との関連性を明らかにするため、構築した sll1558
株と野生株を通常培養条件であるpH 8.0 および酸性ストレス条件である pH 6.0 で生育させ
た(Fig. 3-6)。pH 8.0 では、sll1558 株と野生株に有意な差は認められなかった(Fig. 3-6A, B)。
しかし、pH 6.0 では、sll1558 株は野生株と比較して有意に生育が抑制された(Fig. 3-6A, C)。 前項で述べたsll1558 株における Real-time PCR の結果と合わせると、pH 8.0 では、野生株 と比較して転写量に差が見られなかったことから、生育においても有意な差は認められな かったが、pH 6.0 では sl1558 株の転写量が低下したために、生育が抑制されたと考えられ る。sll1558 株では UDP-glucose の産生量が野生株よりも低下していると予想される。この ため、glucose を利用する細胞外多糖や、UDP-glucose から合成される膜脂質の 1 つである Sulfoquinovosyldiacylglycerol(SQDG)の合成に影響を及ぼし、酸性ストレス耐性が低下する可 能性が示唆された。
46 Fig. 3-6 野生株および sll1558 株の酸性ストレス下での生育 野生株およびsll1558 株を pH 8.0 と pH 6.0 の液体培地(A)および寒天培地(B, C)で生育さ せた。生育曲線はOD730を測定し、その値をプロットしたものである。寒天培地では1, 1/10, 1/100 と希釈した。独立に 3 回実験を行った。(A)●:野生株 pH 8.0、■:sll1558 株 pH 8.0、○: 野生株pH 6.0、□:sll1558 株 pH 6.0、(B) pH 8.0、(C) pH 6.0
47 3-1-4 様々な環境ストレス下での生育
酸性ストレス耐性に関与する遺伝子としてこれまでに、Na+/H+対向輸送体の NhaS4 や、
光誘導性プロトン放出に関与するPxcA などが報告されている (Wang et al., 2002; Sonoda et
al., 1998)。これらは塩ストレスや CO2,、NO3-の取り込みにも関与することが報告されてい る。また他にも、酸性ストレス耐性に関与する脂質輸送タンパク質のSltA1 や Slr0967、Sll0939 な ど も 塩 や 浸 透 圧 ス ト レ ス 耐 性 に 関 与 す る こ と が 明 ら か に さ れ て い る(Tahara et al., 2012;Uchiyama et al., 2014)。 さらに前述したように、Sll1558 は細胞表層の構築に関わっている。細胞表層は様々な外 部ストレスに最も曝されている細胞の一部であることから、様々なストレス耐性と密接に 関わっている。 Sll1558 が酸性以外のストレス耐性に及ぼす影響を確認するために、sll1558 株を様々な環 境ストレス下で生育させた。sll1558 株と野生株を無機イオン過剰または欠乏下で生育させ
たとき、有意な差は認められなかった(personal communication)。0.5 M NaCl を B-11 培地に添
加した高塩濃度では有意に生育が抑制された(Fig. 3-7A)。高浸透圧(0.5 M sorbitol)ストレス下
においても、sll1558 株は有意に生育が抑制された(Fig. 3-7B)。さらに、低温(20℃)または高
温(40℃)ストレスにも sll1558 株は野生株よりも感受性を示した(Fig. 3-7C, D)。以上の結果か
48
Synechocystis は 塩 や 浸 透 圧 ス ト レ ス に 曝 さ れ る と 、 適 合 溶 質 と し て Sucrose や glucosylglycerol(GG)を合成する(Reed and Stewart, 1985)。Sucrose は Fluctoes-6P に UDP-glucose
が作用してsucrose phosphate を経て合成されるが、GG は glycerol-3-phosphate に ADP-glucose
が作用してglucosylglycerol phosphate を経て合成される(Pade and Hagemann, 2014)。先に述べ
たように、Sll1558 は UDP-glucose の合成に関与しているため、sll1558 株では UDP-glucose が減少し、適合溶質であるSucrose の合成ができない、または合成量が減少したために、塩 や浸透圧ストレスに感受性を示した可能性がある。 また、Mitschke らによる RNA-Seq 解析によると、低温(15 ºC)ストレス下における sll1558 の転写量は対数増殖期における転写量と同等である(Mitschke et al., 2011)。しかし、sll1558 株は低温環境下で生育が抑制されたことから、sll1558 の転写量の低下が低温ストレス耐性 に影響した可能性が示唆された。高温(42 ºC)ストレス下においては sll1558 の転写量が低下 している (Mitschke et al., 2011)。
Synechocystis に 重 要 な 膜 脂 質 で あ る monogalactosyldiacylglycerol (MGalDG) と digalactosyldiacylglycerol (DGalDG)の合成には、diacylglycerol (DG)から一度 monoglucosyl
diacylglycerol (MGluDG)を経由するが、このとき glucose の供与体となるのが UDP-glucose で
ある。また、SQDG は UDP-glucose から UDP-6-sulfoquinovose を経由して合成される。その
ため、UDP-glucose は MGalDG、DGalDG、SQDG の合成に関与しており (Awai et al., 2014)、
49 された可能性がある。シロイヌナズナでは膜脂質の一つである DGalDG の合成酵素 (digalactosyldiacylglycerol synthase 1)である DGD1 をコードする遺伝子変異である dgd1-2 や dgd1-3 を持つ変異株の解析から、脂質組成の変化が熱ショックタンパク質(HSP)の発現抑 制に関与することが明らかにされたため、脂質組成が熱耐性の獲得に重要な役割を持つこ とが報告されている (Chen et al., 2006)。
シアノバクテリアNostoc commune では、UV-B を照射した培養液から産生された EPS の
量が、通常の培養液の3 倍以上に増加することが報告されている(Ehling-Schulz et al., 1997)。
これはUV-B によって EPS 合成が刺激され、UV-A/B を吸収する mycosporines というオリゴ
糖が供給されることを意味している。このことから、EPS 合成は UV-B 損傷を軽減する可能
性がある。また、mycosporines は構造中に glucose を含むため(Böhm et al., 1995)、UV-B 耐
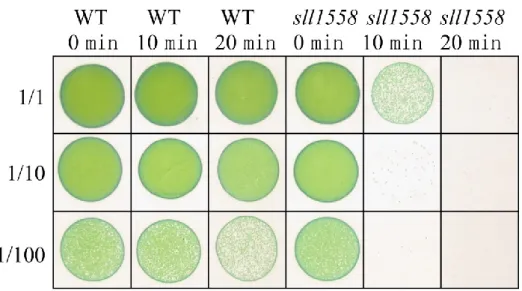
性にsll1558 が関与する可能性がある。さらに、UV-B 照射の 2 時間後、sll1558 の発現が増 加することも先行研究で明らかにされている(Huang et al., 2002)。 そこで、UV 損傷保護への Sll1558 の寄与を調べるために、UV-B ストレス後の細胞生存率 を測定した(Fig. 3-8)。10 分または 20 分の UV-B 照射は sll1558 株の生存率を顕著に低下させ た。この結果から、Sll1558 が Synechocystis における UV-B 保護への重要な役割を果たすこ とを示した。
細胞の外側に存在するEPS は、周囲の環境中に遊離される多糖のことである(Pereira et al.,
50
成する糖の約20%が Glucose であることから(Schmidt et al., 1980)、CPS や EPS にも glucose
が含まれていることが予想される。これらのことから、sll1558 株では、細胞外多糖の構造
51
Fig. 3-7 野生株および sll1558 株の様々な環境ストレス下での生育
(A, B)野生株および sll1558 株を(A) 0.5 M NaCl (B) 0.5 M Sorbitol の寒天培地で生育させ
た。(C, D)野生株および sll1558 株を(C) 20ºC (D) 40 ºC で生育させた。OD730を測定し、濁度
52
Fig. 3-8 野生株および sll1558 株に対する段階的な UV 照射の影響
対数増殖期まで培養した野生株およびsll1558 株を OD730 = 1 に合わせ、UV を 0 min, 10
53 3-1-5 TEM による細胞表層および形態の解析
Synechocystis を含むグラム陰性細菌の細胞表層は細胞質側から、内膜(IM)、ペプチドグリ カン(PG)、外膜(OM)、リポ多糖(LPS)、そして莢膜多糖(CPS)から構成されている(定家ら 2004,
Fig. 1-3)。LPS は内膜にアンカーされている lipid A に core oligo saccharide、O-antigen が結合
しており、CPS は細胞近傍をカプセル状に覆っている。
枯草菌では、糖脂質合成に関与する UDP-glucosyltransferase をコードする ugtP 欠損株は
細胞が屈曲して膨張することが報告されている(Matsuoka et al., 2011)。Sll1558 が合成する
UDP-glucose は糖脂質合成などの細胞表層の構築に関与するため(Vilches et al., 2007; Leipold
et al., 2007)、sll1558 株においても細胞の形態や表層に変化が起こる可能性を考えた。これを 確認するために、透過型電子顕微鏡(Transmission Electron Microscope; TEM)により細胞を観
察した(Fig. 3-9)。
はじめに、細胞形態の観察と細胞長の計測を行った(Fig. 3-9A, 3-10A)。細胞形態において
は、pH 8.0 と pH 6.0 のどちらにおいても野生株と sll1558 株に変化は見られなかった(Fig.
3-9A)。野生株の細胞長は、pH 8.0; 1805.23±89.37 nm, pH 6.0; 2413.48±143.11 nm、sll1558 株の
細胞長はpH 8.0; 1986.28±119.59 nm, pH 6.0; 2275.07±156.62 nm であった(Fig. 3-10A)。野生株
とsll1558 株のどちらにおいても、酸性ストレス下で細胞長が約 1.2 倍になり、有意に大き
54 も報告があり、一致している(Uchiyama et al., 2015)。これらの結果から、Sll1558 に合成され るUDP-glucose の存在は、細胞の形態および大きさに影響を与えないことが示された。 次に、細胞表層の厚さを計測した(Fig. 3-9B, 3-10B)。pH 8.0 では、野生株と sll1558 株の細 胞表層の厚さに有意な差は見られなかった(41.4 ± 2 nm and 41.2 ± 1.8 nm; Fig. 3-10B)。一方、 酸性ストレス下では、野生株の細胞表層の厚さが減少したが、sll1558 株には変化が見られ なかった(39.5 ± 2.1 nm and 34.4 ± 1.3 nm; Fig. 3-10B)。 以上の結果から、Synechocystis は細胞表層の密度を高くすることにより、酸耐性に適応す ることが示唆された。
55 Fig. 3-9 野生株および sll1558 株の形態と細胞表層の TEM による観察 野生株およびsll1558 株を pH 8.0 および pH 6.0 の液体培地で 7 日間培養し、TEM により 観察した。(A)スケールバーは 1µm (B) スケールバーは 200 nm
(B)
(A)
56 Fig. 3-10 野生株と sll1558 株における細胞長および表層の厚さの計測 細胞長(A)および表層の厚さ(B)を TEM により計測した(n = 20)。*、**は pH 8.0 と pH 6.0 における有意差を表す(Student’s t-test; *P<0.05)。
(A)
(B)
57 3-1-6 細胞外多糖の構造解析
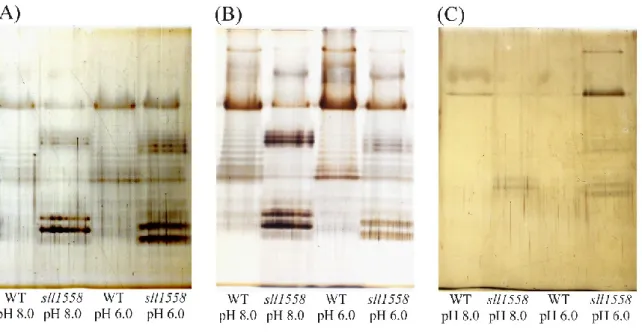
sll1558 株では、酸性ストレス下において、細胞表層の厚さに変化が見られなかった。そ のため、表層の構造に差異があるのではないかと考えた。Schmidt らによると、細胞表層を
構成するLPS には glucose が 21.8% / dw 含まれている(Schmidt et al., 1980)。また、Microcystis
aeruginosa NIES-87 の LPS のコア多糖 は glucose や rhamnose、mannose、galactose、xylose を
含み、O-antigen は glucose のみで構成されていることも報告されている(Fujii et al., 2012)。
UDP-glucose は細胞外多糖の glucose 供与体であるため、sll1558 株は正常な細胞外多糖が産 生できない可能性が考えられる。そこで、野生株およびsll1558 株の LPS をフェノール抽出 法により抽出し、SDS-PAGE によって分離した。 親水性LPS 画分では O 側鎖糖ユニットのリピート配列と考えられるラダー状のバンドを 確認した(Fig. 3-11A)。LPS の主要なバンドの位置は、野生株では高分子量側、sll1558 株で は低分子量側に現れ、見え方が大きく異なっていた。また、野生株、sll1558 株ともに、酸性 ストレスによるLPS への影響は見られなかった(Fig. 3-11A)。 さらに、CPS、RPS を抽出し、SDS-PAGE によって分離した。LPS と同様に、野生株と sll1558 株ではバンドパターンが大きく異なることが確認できた(Fig. 3-11B, C)。野生株では CPS、RPS ともに酸性ストレスによる影響はほとんど見られなかったが、sll1558 株は酸性ス トレス下でCPS がわずかに減少し、RPS は増加していた(Fig. 3-11B, C)。
58
正常な LPS を持つ野生株は分子量が大きいために、高分子量側でバンドを確認できたと
考えられる。さらに、1 つの菌種が産生する LPS の O-antigen は複数種類にのぼることから、
検出されるバンドはシングルではなくラダー状になる(Li and Wang, 2011)。
これらの結果から、sll1558 株は UDP-glucose が欠損しているために、正常な LPS を合成
できないことが示唆された。さらに、CPS や遊離多糖(RPS; release polysaccharide)を含む EPS
を抽出し、SDS-PAGE により構造を確認した(Fig. 3-11B, C)。野生株と sll1558 株における CPS とRPS のバンドパターンもまた LPS のときと同様の結果となった。 これまでの結果をまとめると、sll1558 株は UDP-glucose の合成量が低下しているために 正常なLPS が合成できず、酸性ストレスに感受性を示すことが示唆された。 Sll1558 は糖の合成に関わるタンパク質であることから、sll1558 株の LPS だけでなく CPS、 RPS も野生株と異なることが明らかになった。また、酸性ストレス耐性には細胞外多糖が重 要であることも示された。そこで、細胞外多糖のストレス耐性への影響を個別に比較するた めに、LPS 欠損株を構築し、LPS のみによる酸耐性への影響を確認することにした。
59
Fig. 3-11 野生株および sll1558 株の LPS, CPS, RPS の解析
野生株およびsll1558 株を pH 8.0 および pH 6.0 の液体培地で培養した。LPS はフェノール
60 3-2-1 アミノ酸配列比較による MsbA オーソログの同定 3-1 では Sll1558 変異によって LPS や CPS の構造が変化し、sll1558 株が酸性ストレスを はじめとする、様々な環境ストレスに感受性を示すことを明らかにした。しかし、LPS と CPS のどちらが酸耐性に寄与しているは明らかにできていない。 そこで次にLPS のみを欠損させ、LPS がストレス耐性に影響を及ぼすかを検討した。
LPS 欠損株を構築するために、KEGG PATHWAY を用いて Synechocystis における LPS 合
成経路を探索した(http://www.genome.jp/kegg/pathway.html)。Synechocystis の LPS 合成経路に
はO-antigen や lipid A 合成に関与する酵素(LpxA, LpxC, LpxD, and LpxB)がいくつか保存さ
れていたため、lipid A に着目した。
大腸菌のMsbA は 内膜に存在する ABC transporter であり、(Karow and Georgopoulos, 1993)
LPS の lipid A を内膜の inner leaflet から outer-leaflet に輸送することが先行研究で明らかに
されている(Polissi and Georgopoulos 1996; Zhou et al. 1998; Sperandeo et al. 2008 )(Fig. 1-4)。
Synechocystis が MsbA と 相 同 性 の 高 い タ ン パ ク 質 を 持 つ か 、 NCBI の Blast search (http://blast.ncbi.nlm.nih.gov/Blast.cgi)を用いて検索した。その結果、Slr2019(36% identities and
60% similarities)、Slr1276 (33% identities and 56% similarities)、Sll1725 (32% identities and 53%
similarities)、Slr1149 (31% identities and 52% similarities) (Table 11)の 4 つ存在することが明ら
61
MsbA は膜貫通ドメインと ATP 結合ドメインそれぞれ 1 つからなる ABC タンパク質であ
り、ATP 結合ドメインには、アミノ酸配列がよく保存された 4 ヶ所の領域(Walker A、C motif、
Walker B、switch)が存在する(植田, 2005)。一方で、膜貫通ドメインに保存されている配列は
ABC タンパク質に特有の EAA を含むモチーフが知られているのみである(植田, 2005)。
Synechocystis の 4 つのタンパク質は、MsbA の膜貫通領域である 31 番目から 298 番目に 該当する配列前半部分の相同性は低いが、配列後半部分の Walker A、C motif、Walker B、
switch はよく似ている。また、それぞれの膜貫通領域の数は Slr2019;5、Sll1276;4、Sll1725;3、 Slr1149;5 であった(Table 11)。MsbA の膜貫通領域の数は 5 で奇数回なので、Slr2019、Sll1725、 Slr1149 は N 末 C 末が MsbA と同じ向きに存在しており、似たトポロジーを持つことが示 唆されている。 この結果から、Synechocystis の 4 つのタンパク質は、MsbA とよく似た特徴を持つことが 明らかになり、lipid A flippase である可能性が示唆された。
62 Fig. 3-12 MsbA ホモログのアラインメント
(A)MsbA, Escherichia coli (B) Slr2019, Synechocystis 6803 (C) Sll1276, Synechocystis 6803 (D)
Sll1725, Synechocystis 6803 (E) Slr1149, Synechocystis 6803
63