Kyushu University Institutional Repository
大麦焼酎製造工程におけるグリセリンの生成機作と その役割に関する研究
大森, 俊郎
https://doi.org/10.11501/3099962
出版情報:Kyushu University, 1994, 博士(農学), 論文博士 バージョン:
権利関係:
第5章 酵母のグリセリン生成に及ぼすグルコース, クエン酸 および脂肪酸の影響
5 - 1 緒言
第4章で全麹もろみにおける酵母のグリセリン高生成要因を検討し, もろみ 中のクエン酸および脂肪酸の濃度の違いが大きく影響していることを述べた。
これまで, グルコース濃度が酵母のグリセリン生成に影響する37 , 4 0 ) ことは 知られていたが, クエン酸や脂肪酸が影響するとの報告はない。
クエン酸は麹に由来し, 焼酎もろみ中に存在するため, 焼酎酵母にはクエン 酸耐性が求められる。 一方, 脂肪酸は菌体を構成する上で必須成分であり, 培 地中に遊離脂 肪酸 が 存 在し た場合, 酵母によ って速やかに取り込まれ,
triglycerideあるいはphospha tid y lch olineのacyl基として代謝, 使用されることな ど酵母の生理に与える影響が大きい9 0 - 9 8 ) 。 また 不飽和脂肪酸は酵母のエ ステル生成に影響する81,83,84) ことが知られており, 飽和脂肪酸は代謝産 物生成に対しても影響する。
本章では培地中のクエン酸濃度が高いほと々酵母菌体内のGPase活性を高め,
その結果酵母のグリセリン生成が向上していること, オレイン酸およびリノー ル酸は酵母のグリセリン生成を向上させ, パルミチン酸は抑制させること, ま た他の醸造用酵母もオレイン酸によってグリセリン生成が向上することを見い だしたことについて述べる。
5-2 実験方法
産呈
大麦焼酎酵母(BAW-6), 鹿児島酵母(Ko), 宮崎酵母(MK), 協会焼酎 酵母2号(SH-4), 清酒酵母協会7号(K-7), 清酒酵母協会9号(K-9), ワイ ン酵母(W-3, OC-2) , ビール酵母Saccharomyces cereνisiae IFO 1167およびウ イスキー酵母Saccharomyces cereνisiae IF02373を使用したO 各酵母のグリセリ ン生成を比較した実験以外はBAW-6を用いたO
グリセリン生成に及ぼすグルコース濃度の影響
YEPD(n)培地(n%グルコース, 10/0酵母エキス, 20/0ポリペプトン)を用いて,
グリセリン生成に及ぼすグルコースの影響を 調べた。 YEPD(2.5)培地は1---10日 目まで毎日2. 50/0量のグルコースを加えたもので並行複発酵モデルとして行った。
YEPD(25)培地は1日目に250/0量のグルコースを添加し, それ以降は添加しなかっ たもので単発酵モデルとして行った。 YEPD(5)培地はし3, 5, 7, 9日目に50/0 量のグルコースを加えたもので並行複発酵, 単発酵の中間モデルとして行っ た。BAW-6 を1X 108cells/mlに調整し た懸濁液を10/0接種し, 300Cで1 2日開発酵 させた。
グリセリン生成に及ぼすクエン酸濃度の影響
YEPD(5)培地を基本培地とし, 脱イオン水の代わりに0.3�3.0%のクエン酸緩 衝液(pH3, 3.5, 4)用いて行った。 また, 基本培地を1M塩酸で pH3, 3.5, 4に
それぞれ調整した培地でも実験を 行った。 発酵は前培養したBAW・6を1X 108 cells/mlに調整した懸濁液を10/0接種し, 30oC, 5日間で、行った。
グリセリン生成に及ぼす脂肪酸の影響
脂肪酸濃度の影響はGCT培地( 100/0グルコース 1 .170/0 Yeast Carbon Base,
0.5%脱脂トリプトン, 0.1% 脱脂albumin )に0.5, 1, 5, 10mMになるようにパ ルミチン酸, オレイン酸およびリノール酸を添加した培地で検討した。 コント ロールは脂肪酸無添加培地とした。
窒素 源が異なる培 地 での脂 肪 酸 の影 響はGC培地(10 0/0グルコース,
1.17%Yeast Carbon Base. 0.10/0脱脂albumin)にそれぞれ0.5mMになるようにパ
ルミチン酸, オレイン酸, リノール酸を添加し, 窒素源として0.5%の塩化アン モニウム, 硫酸アンモニウム, 脱脂トリプトン, 脱脂ポリペプ トンおよび脱脂 カザミノ酸を使用した培地で行った。 各窒素源区の脂肪酸無添加培地をコント ロールとした。
各種醸造用酵母のグリセリン生成はGCT培地に0.5mMになるようにパルミチ ン酸, オレイン酸およびリノール酸をそれぞれ添加した培地で検討したO コン
トロールは脂肪酸無添加培地とした。
それぞれの培地に予め前培養した酵母(1X 108cells/mJ)を10/0接種し, 300Cで 発酵させた。
酵母菌体内酵素およびタンパク質の調製
培養液から遠心分離により酵母を集菌し, さらに脱イオン水で2回洗浄後,
再び遠心分離で集菌した。 集菌酵母にガラスビーズを加え, マ イクロチューブ ミキサー(タイテック,EM-36)を用い 40Cで3時間振動させて細胞を破壊し,
遠心分離して上澄み液を酵素液およびタンパク質含有サンプル液とした。
GlyceroQhosphatase (GPase)活性の測定
高山ら9 9 )の 方 法を 一部 改 変 し GPase活性を測定した。 すな わち ,
450mMリン酸緩衝液(pH 5.0) , 20/0 sodium glycerophosphate およ び酵素液 O.5mlを含む反応液(5.5ml)に防腐のためにトルエン0.2mlを添加し,300Cで24 時間反応させ, 生成するグリセリンをF - Ki t (ベーリンガー ・ マンハイム山之 内)を用いて測定した。 GPase活性は生成したグリセリン量(μg)を酵母菌体 内タンパク質lmg当たりで表した。
Glycerol kinase (G Kase)活性の測定
GKaser舌性の測定はAdlerら1 0 0 )の方法を一部改変して行った。 すなわち,
100mMグリシ ンー水酸化ナトリウム緩衝液(pH9. 5) , 300mM硫化ヒドラジ ン,20mM塩化マグネシウム, 20mM ATP, 10mMグリセリン, 1.4mM NAD+お よびlml当たり10unit含むGlycerol 3-phosphate dehydrogenase (Glyc-3PD,東洋 紛)を含む反応液に酵素液O.lmlを加え,全体で3mlとし, 300Cで20分酵素反 応させ, 増加し たNADHを 340nmで測定 した。 ブランクはグリセリンを加
えない反 応液に酵素液を入れ, 以下同様に操作した。 GKaseによりグリセリ
ンからglycerol-3-ph osphateが生成し , さらにGlyc-3PDにより dihydrox yaceto ne
phosphateが 生成するが, このときNAD+から生成するNADH量を定量するため に, サンプルとブランクとの吸光度差を求め, 予め求めたNADH量と340nmに おける吸光度との関係から, NADH量(μmol)を求めた。 NAD+から1分間 当たりlμmolのNADHが生成する活性を1unitとし, 酵母菌体内 タンパク質 lmg当たりで表した。
酵母菌体内タンパク質の測定
酵母菌体内タンパク質は プロテインアッセイキットII(バイオラッド)を 用いて測定した。 すなわち タンパク質含有液を10倍希釈し, 5倍に希釈し たプロテインアッセイキット試薬5mlにタンパク質含有サンフ。jレ液O.lmlを加 え, 室温で30分間反応させ, 595nmの吸光度を 測定した。 タンパク質量はサ ンプルとブランクとの吸光度差を求め, 予め求めたアル ブミン量と595nm
での吸光度との関係から, タンパク質量(mg)を求めた。 ブランクはサ ンプル液の代わりに脱イオン水O.lmlを用いて同様の操作 を行った。
主主
グルコース, ポリペプトン, albuminおよび高級脂肪酸は和光純薬製, Yeast Nitrogen Base, Yeast Nitrogen Base w/o amino acid and ammonium sulfate, Yeast Carbon Base, トリプトンおよびカザミノ酸はDifco製を使用した。 また, ポリ
ペブ。トン, トリフトン, カザミノ酸およびalbuminは24時間エーテルで脱脂した ものを使用した。
会主
グリセリン, グルコースは第l章1 -2に示した方法で測定した。 吸光度は 分光光度計(目立, U-3210)で、1cmセルを用い, 660nmで、 測定した。 酵母の生 菌数は第3章3-2に示した方法で測定した。
5-3 実験結果
5-3-1 合成培地でのグルコースの影響
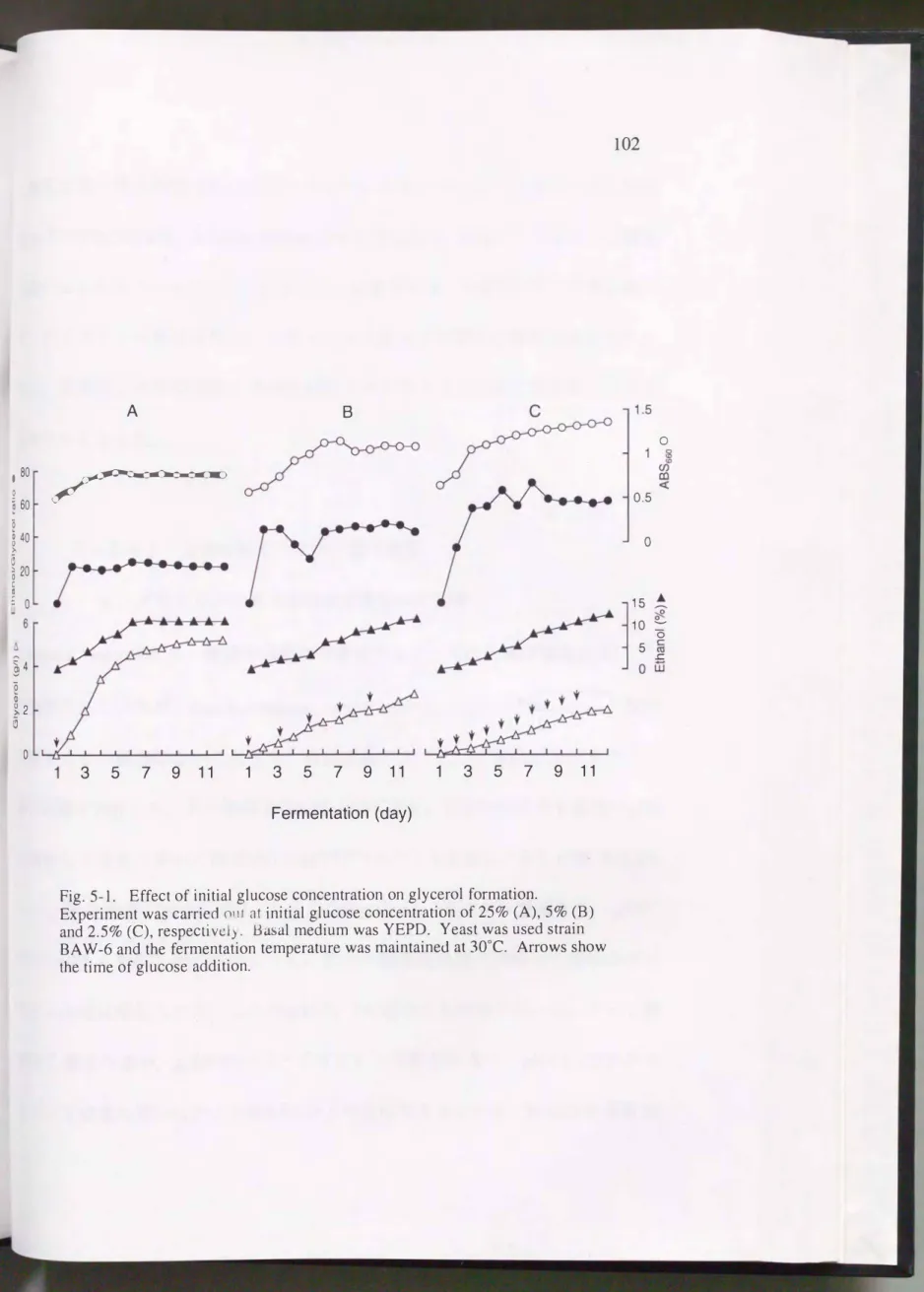
並行複発酵モデルにおいてグ ル コー ス 濃 度 が酵 母 の グ リ セ リ ン 生 成 に 及ぼす影 響を検討した。 単発酵タイプのYEPD(25)培地の場合(Fig.5-1A),
群母の増殖(ABSω)およびエタノール生産は, 4日目および7日目までに それ ぞれ終了し, グリセリン は7日目までに大部分が生成され, その後発酵終了ま で微増した 。 一方 , 並行複発酵タイプの YEPD(2.5)培地の 場合(Fig.5-1C),
菌体量, エタノール グリセリンは12日固まで徐々に増加した。
グリセリンは, YEPD(25), YEPD(5) , YEPD(2.5)培地でそれぞれ5.20, 2.75,
1.98g/
1生成され, 初発グルコース濃度が高いほど増加した。 一方, エタノールfiYEPD(25), YEPD(5), YEPD(2.5)培地でそれぞれ11.2, 12.4, 12.8%生産され,
グリセリンとは違いグルコース濃度が低いほど増加した。 酵母の増殖はグルコー
B
ごωω心l1.5
。
包
0.5
。
申よ�
υ'f�
J\.---I\� �
ム 申市A�一 申 Jf HU会�
3 5 7 9 11 1 3 5 7 9 11 1 3 5 7 9 11 A
nv nv nv nu nu au RU A吟 qL .,
oza』・0ιEwuhzu、ZMWEEPゐ“
/ポ∞∞…
必斗 門〆』
内-、m凶》-n》Lmw川》、AU-mu
。
Fermentation (day)
15
た、10ご
。5
ょ=@Fig. 5-l. Effect of initial glucos_e.C?n?entration on gly�erol �O!�!t���.
Experiment was carried o1Jl at initial glucose concentrat10n of 25%(A),5%(B)
and 2.5% (C), respectivel)・ βasal medium was YEPD. Yeast was used strain
BA\\人6and the fêrmentaiion temperature was maintained at 30oC. Arrows show the time of glucose addition.
ス濃度が高いほど抑制される傾向にあった。 エタノールとグリセリンとの生成
量比はYEPD(25)培地, YEPD(2 .5)培地でそれぞ、れ21.5, 64.6で, グルコース濃度
�)高いほど小さくなった。 以上のように, 初発グルコース濃度が高くなるに従っ てFグリセリン生成は増加し, エタノール生産および酵母の増殖は減少したo iたF単発酵と並行複発酵とでは前者の方がグリセリン生成には望ましいこと が明らかとなった。
5-3-2 合成培地でのクエン酸の影響
a. グリセリン生成と酵母の生菌数への影響
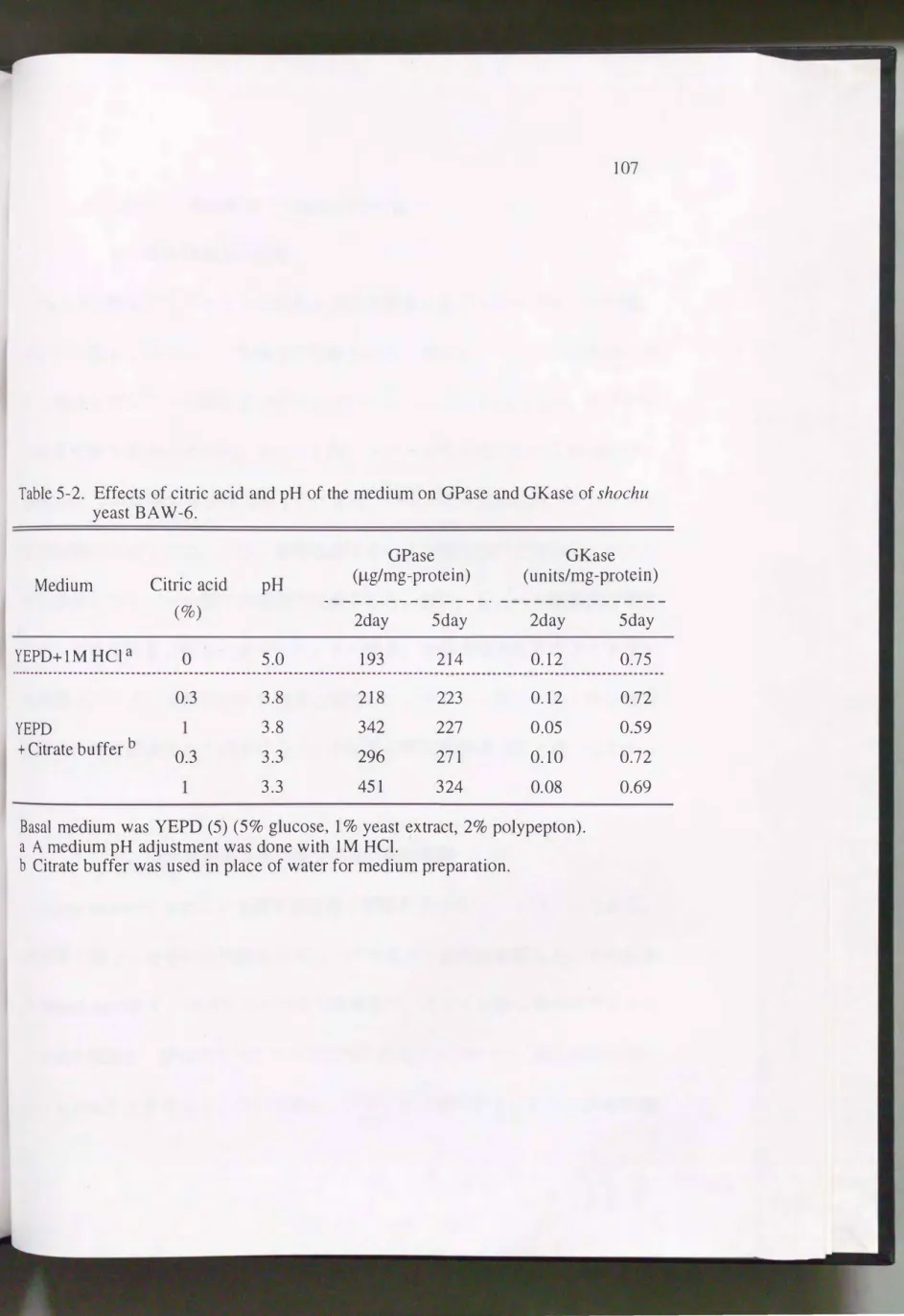
Pichia misoはクエン酸緩衝液添加で多価アルコール の生成 が 増加す る1 0 1 ) と報告されているが, Saccharomyces cerevisiaeのグリセリン生成にクエン酸が 影響するとの報告はないことか ら, 合成培地でグリセリン生成に及ぼすクエン 酸の影響を検討したO その結果をTable5-1a,bに示す。 YEPD(5)培地を塩酸で、pH3 に調整した培地において酵母は0.82g/1のグリセリンを生成し, さらに酵 母は1%
クエン酸緩衝液(pH3)ではこれよりO.34g/1多く, 3%クエン酸緩衝液(pH3)
でO.52g/1多く生成した。 このよ うに, クエン酸濃度の増大に伴って酵母のグリ セリン生成は増加したが, これはpH3.5, 4の場合でも同様であった。 クエン酸 が同じ濃度の場合, pHが低いほどグリセリン生成量は多く, pH3と4でのグリ セリン生成量の違いはクエン酸0.50/0以上の培地で大きかった。 酵母の生菌数は
Table 5-1 a. Effect of citric acid on glycerol formation by shochu yeast BA W -6.
Glycerol formed (g/l)
pH Citric acid concentration (%) of medium
『司・ー司司・ーーーーーーーーーーーーーーーーーーー晶画ーーーーーーーーー・・・ーーーー--晶画ーーーーーー司ーーーー喧ー圃ーーーー-ーー唱ーーーー骨唱Fー--ーー-ーーー・・ー---ー司ーー
a 0 b 0.3 b 0.5 b 1 b 2 b 3
3 0.82 0.96 1.08 1.16 1.31 1.34
3.5 0.76 0.92 0.95 1.07 1.15 1.28
4 0.72 0.88 0.91 0.95 1.05 1.19
Table 5-1 b. Effect of citric acid on variable yeast cells.
pH
Variable yeast cells ( x 107 Iml) Citric acid concentration (0/0) of medium
a 0 b 0.3 b 0.5 b 1 b 2 b 3
3 5 4
司3
5.4 5.6 6.1
3.1 3.1 4.1
2.6 2.6 3.4
1.2 2.1 2.8
0.9 1.7 2.4
0.7 1.1 1.5 Basal medium was YEPD (5) (50/0 glucose, 1 % yeast extract, 2% polypepton).
a A medium pH adjustment was done with 1M HCl.
b Citrate buffer was used in place of water for medium prep紅ation.
クエン酸濃度が高いほど, またクエン酸濃度が同じであれば pHが低いほど少な かっfこO
全麹もろみは麹歩合330/0もろみよりもクエン酸濃度は高く, pHは低い。 この ことから, 全麹もろみは麹歩合330/0もろみに比べると酵母がグリセリンを生成 するのに適した環境であることが推察される。
b. GPaseおよび、GKase活性 に及ぼす影響
グリセリンはglycerol-3-phosphate ( Glyc-3P) を基質としてGPaseによる脱リ ン酸化で 生成され, GKaseによるリン酸化で代謝される (Fig.5-2)。 そのため,
GPaseおよび、GKaseはグリセリン生成 に重要と考えら れるので, 両酵素の活性 に 及ぼすクエン酸濃度の影響を検討した。 そ の結果 をTable5-2に示す。 GPase活 性は, クエン酸濃度が高いほど高く, またクエン酸濃度が同じで あれば, pHが 低い方がGPase活性は 高かったO 全麹もろみ(クエン酸1 0/0, pH3 .3 ) に相当す る培地での2日目のGPase活性は, 麹歩合330/0もろみ(クエン酸0.30/0, pH3.8) に相当する培地でのGPase活性のおよそ 2.1倍であった。 また, GKase活性 は 5 日 目の方が高く, クエン酸濃度が高い培地で低い傾向にあったが, 培養条件によ るGKase活性の違いは ほとんどなかった。 以上の結果, クエン酸濃度の増加が 酵母のGPase活性を向上させ, グリセリン生成を増加させ ていることが示唆さ れた。
Glucose
V---
w
FDP
DHAP TPI
;
ENADH 主NAD+ GAP Glyc-3P-VT・'V7・
Glycerophosphatase (GPase)
Glycerol kinase (GKase)
G ・v ・- v ・ v ・
-v'lvM 一一戸』
UH+ DD AA NN
Glycerol
Fig.5-2. Biochemical synthesis of glycerol in Saccharomyces cerevisiae.
Abbreviations: FDP, Fructose 1,6-diphosphoglycerate GAP, Glyceraldehyde 3-phosphate DHAP, Dihydroxyacetonephosphate Gly-3P, Glycerol 3-phosphate
TPI, Triosephosphate isomerase.
Table 5-2. Effects of citric acid and pH of the medium on GPase and GKase of shochu
yeast BA W -6.
GPase GKase
Medium Citric acid pH (μg/mg -protein) (units/mg-protein)
ー一一ーー一一ーーーーーーーー- 一一一一一一一一一一一一一一一
(%) 2day 5day 2day
YEPD+IM HCla 。 5.0 193 214 0.12
0.3 3.8 218 223 0.12
YEPD 3.8 342 227 0.05
+ Citrate buffer b 0.3 3.3 296 271 0.10
3.3 451 324 0.08
Basal medium was YEPD (5) (5% glucose, 1 % yeast extract, 20/0 polypepton).
a A medium pH adjustment was done with 1M HCl.
b Ci甘ate buffer was used in place of water for medium preparation.
5day 0.75
0.72 0.59 0.72 0.69
5-3-3 合成培地での脂肪酸の影響
a. 脂肪酸濃度の影響
Fig.5-3に酵母のグリセリン生成および酵母菌体収量に及ぼすパルミチン酸,
オレイン酸およびリノール酸濃度の影響を示す。 酵母のグリセリン生成はオレ イン酸およびリノール酸の添加区ともコントロールよりも増加した。 グリセリ ン生成が最も多かったのは, オレイン酸, リノール酸の添加区とも0.5mMで,
lmM以上で、は減少する傾向にあった。 また, パルミチン酸添加区でグリセリン 生成は抑制され ていた。 一方, 菌体収量はオレイン酸添加区で増加し, パルミ チン酸およびリノール酸の添加区では減少した。特に, リノール酸濃度の増加 に伴って菌体収量は顕著に減少した。 その結果, 乾燥菌体当たりのグリセリン 生成量は, リノール酸添加区で顕著に増加した。 オレイン酸, パルミチン酸添 加区での乾燥菌体当たりのグリセリン生成量は脂肪酸無添加区と違いはなかっ
b. 窒素源が異なる培地での脂肪酸の影響
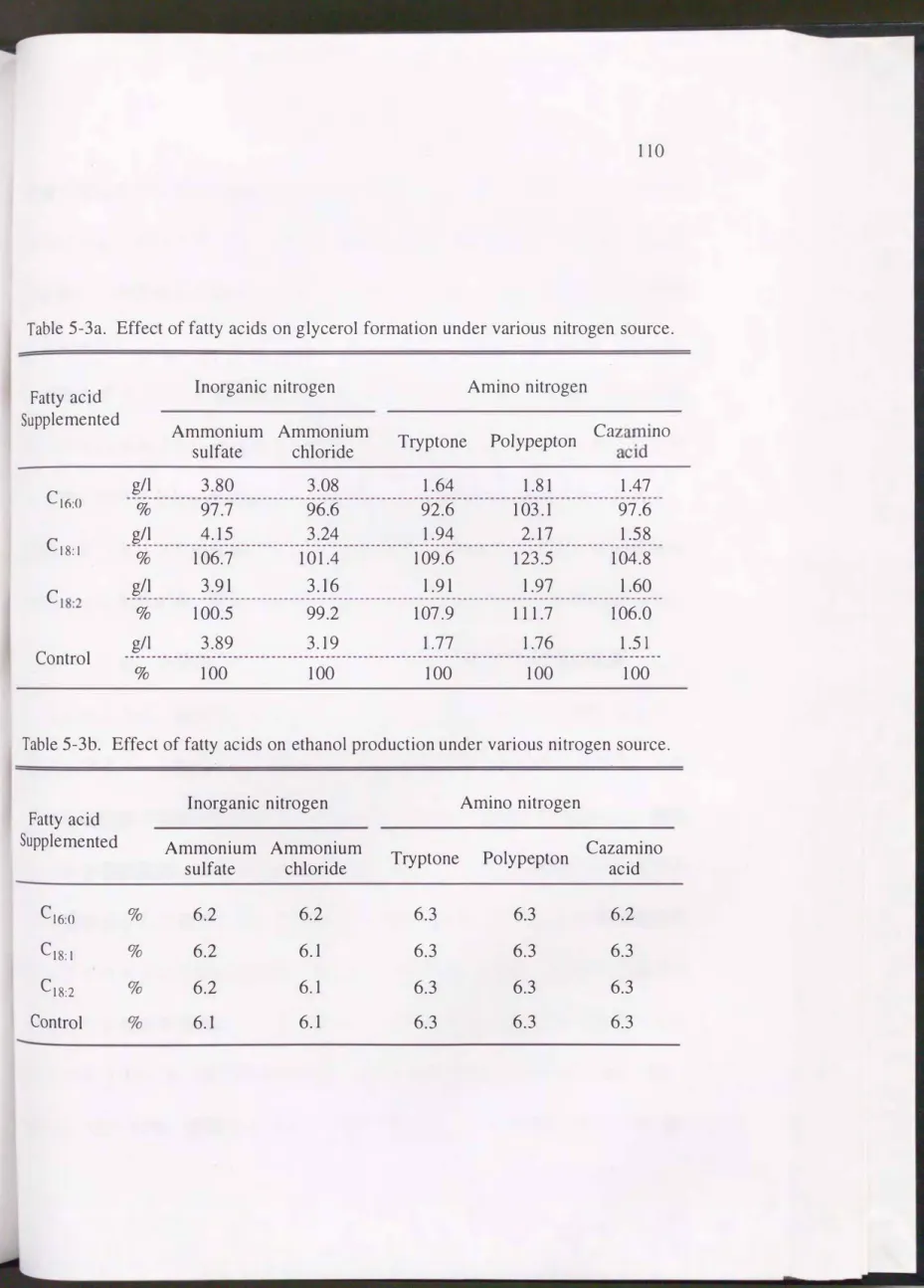
Pichia misoのグリセリン生成は窒素源の影響を受ける1 0 1 • 1 0 2 ) ことから,
窒素源が異なる培地に脂肪酸を添加し, グリセリン濃度を比較した。 その結果 をTable5-3aに示すO 供試したすべての窒素源で, オレイン酸は酵母のグリセリ ン生成を促進し, 特にポリペプトンの添加区ではコントロール(脂肪酸無添加) よりも20%以上多かった。 これに対し, パルミチン酸はポリペプトン以外の窒
2.0
1.8
1.6
(一\O)一O」ωO弘一。
1.4 0.3
0.2
(O)
HZO一ω〉〉=oobo
0.1 1.2 1.0 0.8 0.6 (HZ90〉〉=00と℃'O\窃)
b一〉一ちコ℃O」a一O」00去の
5
Fatty acid concentration (mM)
4 3
。 2
Fig.5-3. Effects of fatty acid concentration on dry cell weight and glycerol formation by shochu yeast BAUF-6.
Symbols: 0 , palmitic acid;口, oleic acid; ß , linoleic acid.
Table 5-3a. Effect of fatty acids on glyceroI formation under various nitrogen source.
Fattyacid Supplemented
Inorganic nitrogen Amino nitrogen
Ammonium Ammonium
sulfate chloride Tryptone
。n md ac za QU C
n 0 6EL P ρiu p vd '・EAo PA
C16:O g/l 3.80 3.08 1.64 1.81 1.47
% 97.7 96.6 92.6 103.1 97.6
g/l 4.15 3.24 1.94 2.17 1.58
0/0 106.7 101.4 109.6 123.5 104.8
g/l 3.91 3.16 1.91 1.97 1.60
C18:1
「,Lnxu nし
% 100.5 99.2 107.9 111.7 106.0
g/l 3.89 3.19 1.77 1.76 1.51
0/0 100 100 100 100 100
Control
Table 5-3b. Effect of fatty acids on ethanol production under various ni甘ogen source.
Fattyacid Inorganic nitrogen Amino nitrogen
Supplemented Ammonium Ammonium Cazamino
sulfate chloride Tryptone Polype pton
acid
C16:O % 6.2 6.2 6.3 6.3 6.2
C18:1 % 6.2 6.1 6.3 6.3 6.3
C18:2 % 6.2 6.1 6.3 6.3 6.3
Control % 6.1 6.1 6.3 6.3 6.3
素源添加区でグリセリン生成を抑制させる傾向にあった。 一方, リノール酸は
トリプトン, ポリペプトン, カザミノ酸を窒素源とした場合にグリセリン生成 を促進し, 硫安および塩化アンモニウムを窒素源とした場合にはほとんど影響 しなかった。
酵母のグリセリン生成が最も多かった窒素源は硫安で3.9---4.2g/l, 次いで塩 化アンモニウムで3.1---3.2g/1で、あった。 トリプトン, ポリペプトンおよびカザ
ミノ酸でのグリセリン生成は1.5---2.2g/1で, アミノ態窒素を窒素源とした場合,
酵母のグリセリン生成は低下した。 Table5-3bに示すようにエタノール濃度は グリセリン生成が多い硫安 塩化アンモニウムの添加区で0.1---0.2%低かった。
c. 各種醸造用酵母のグリセリン生成に及ぼす脂肪酸の影響
以上のように 焼酎酵母BAW-6のグリセリン生成はパルミチン酸, オレイン 酸およびリノール酸によって影響されることが明らかとなった。 そこで, この ような脂肪酸の影響が他の醸造用酵母に対してはどうなのかを検討した。 醸造 もろみ中の窒素源はアミノ態窒素であることから, ここでは脱脂したトリプト ンを窒素源として使用した。 その結果をTable5-4�こ示すO オレイン酸は菌株に よってグリセリン生成の増加率に違いはあるものの, 供試したすべての酵母の グリセリン生成を促進した。 すなわち, オレイ ン酸添加区のグリセリン生成 は, コントロール(脂肪酸無添加区)に比べ焼酎酵母(BAW-6, MK, Ko,
SH-4)で4---120/0 清酒酵母(K-7, K-9)で2---30/0, ワイン酵母(W-3, OC-2)
Table 5-4. Glycerol formation by Saccharomyces cerevisiae in fatty acid supplemented media.
Strain
BAW-6 MK
Ko SH-4
K-7 K-9 明人3
OC-2 IF01167 IF02373
Glycerol (g/l) Ratio (%)
Control C16 C18:1 C18:2
1.92 1.81 2.15 2.12
100 94.3 112.0 110.4
2.10 1.92 2.19 2.29
100 91.4 104.3 109.0
2.82 2.52 3.02 3.30
100 89.4 107.1 11 7 .0
2.52 2.45 2.75 2.61
100 97.2 109.1 103.6
2.83 2.73 2.88 2.70
100 96.5 101.8 95.4
2.42 2.28 2.50 2.37
100 94.2 103.3 97.9
3.38 3.37 3.49 3.14
100 99.7 103.3 92.9
2.36 2.36 2.46 2.36
100 100 104.2 100
1.58 1.58 1.68 1.62
100 100 106.3 102.5
2.12 2.11 2.26 2.34
100 99.5 106.6 110.4
Note
Shochu Shochu
Shochu Shochu
Sake Sake Wine Wine Beer Whisky
Medium: 10% glucose, 1.170/0 yeast carbon base, 0.1 % defatted albumin, 0.5% defatted tryptone and 0.5 mM palmitic acid (C16:0),
oleic acid (C18:1) or linoleic acid (C18:2), Temperature was maintained at 30 oC.
で3---4%, ビール酵母(IF01167) ウイスキー酵母(IF02373)で60/0それぞ れ増加したO これに対し, パルミチン酸添加区のグリセリン生成は, コントロー ルに比べ焼酎酵母, 清酒酵母で3---11%減少し ワイン酵母, ビール酵母, ウイ スキー酵母でほ とんど違いはなかった。 また リノール酸添加 区でのグリセリ ン生成は, コントロールと比べ焼酎酵母で4---17%, ビール酵母で30/0, ウイス キー酵母で10%それぞれ増加したのに対し 清酒酵母およびワイン酵母W-3で 逆に2---8%減少した。
5-4 考察
第4章で, 大麦焼酎もろみで はグルコースが低濃度であるために, 全麹もろ みでの酵母のグリセリン高生成 には, グルコース濃度はほとんど関与していな いと推察した。 グルコース量を同じにした並行複発酵モデルと単発酵モデルに おいて, 酵母のグリセリン生成を比較した結果, 単発酵モデルの方がグリセリ ン生成は多くなった。 また, 同じ並行複発酵モデルでも初期のグルコース濃度 が高いほどグリセリン生成は増加し, これは前章Table4-1の結果と一致する。
梅本ら1 0 3 )は 塩存在下でグリセリンが高生成される場合酵母の増殖も抑
制され, 増殖阻害がグリセリン生成の増加に直接結びっくものではないことか ら, 塩のような電解質が単なる増殖阻害剤としてのみ働いているのではないと 報告している。 クエン酸は低濃度でも酵母のグリセリン生成を促進し, さらに
酵母の増殖も 阻害 し(Table5-1a,b) , この 現象 は梅本らI 0 3 ) の報告と一致し ていた。 第4章Table4-1, Table4-2で、述べたように, グルコースは3%以下では グリセリン生成に影響しなかったのに比べ クエン酸は低濃度でも 影 響した (Table5-1 a)。 クエン酸によるグリセリン高生成メカニズムは現在のところ詳 細には明らか ではない が , クエン酸が塩と同じ電解質であるということも比較 的低濃度で 影響した原因ではないかと考えられた。 田嶋らI 0 4 )は, 塩存在下 ではtriosephosphate isomerase (TPI)の作用 の阻害によりdihydroxyacetonephos
phate (DHAP) が 供給さ れ, その結果, glycerol-3-phosphateさら にグリセリン の生成が 促進されると推定している。 Table5-2に示すように, 合成培地におい てクエン 酸による酵母 のグ リセ リン 生成促進の原因 を検討したと こ ろ,
g防1)砂ycωerl刈01トト-ふ.
により増加し, この 結果は田嶋らI 0 4 )の結果と符合する。 また, 第4章 Table4-3で述べたように クエン酸は高濃度のグルコース存在下でも酵母のグ
リセリン生成を促進したことから, クエン酸が酵母のグリセリン生成を促進す るのは大麦焼酎もろみ特有のことではなく, 初期グルコース濃度が高 い米焼酎 全麹もろみあるいはクエン酸を含む培地に一般的に見られる現象と考えられた。
オレイン酸は供試した10種類の醸造用酵母のグリセリン生成を促進し, リノー ル酸は焼酎酵母 ビール酵母お よびウイスキー 酵母のグリセリン生成を促進し たが, 清酒酵母やワイン酵母のグリセリン生成を阻害した。 焼酎, ピール , ウ イスキーの原料であるこ条大麦や破砕米は清酒原料米やブドウと 比べるとリノー
jレ酸が多く, そのもろみや糖化液も清酒もろみなどと比べるとリノール酸は多
いと推定される。 焼酎酵母やビール酵母などはこのような脂肪酸が多い環境で も十分にその特性が発揮できる菌株と考えられ, このような酵母だけがリノー ル酸存在下で、もグリセリン生成が向上したことは興味深い。
不飽和脂肪酸は酵母のエステル生成を阻害し 飽和脂肪酸は促進する作用が あり83,84) , 全麹仕込みは麹中に不飽和脂肪酸が多いことから, エステル 生成には好ましくない1 0 ) 。 酵母のグリセリン生成向上を目的に, もろみ中の オレイン酸あるいはリノール酸の濃度を高めることは 一方で酵母のエステル 生成を阻害することから 結果的には焼酎製品そのもののエステル濃度は低下
する1 0 ) 。 そのため 焼酎製品の香気を改善するためには, 低い麹歩合でグリ セリン濃度だけを増加させるような仕込み条件が望ましいと考えられた。
無機態窒素を用いた培地ではアミノ態窒素を用いた培地に比べエタノール生 産は低下し, グリセリン生成は増加した。Lutstorf and Megnet 4 2 ) はADHアイ ソザイムの一部欠損株にグリセリンを顕著に生産する株を見いだし, ADH活性 の低下でアセトアルデヒドからエタノールへの還元反応に伴って消費さ れる NADHが細胞内で過剰になり, これがDHAPからGlyc-3Pへの還元反応で、消費さ れたために酵母のグリセリン生成が増加したと推定している。 次章で詳細に述 べるが, 酵母のADH�舌性はアミノ態窒素培地よりも無機態窒素培地の方が低かっ た。 これは, 無機態窒素培地で酵母のグリセリン生成が増加したことと符合す る。また, 無機態窒素を窒素源として使用した場合エタノール生産が低下する
のはグルコースからのアミノ酸生合成が盛んになるため1 0 5 )といわれており、
エタノール生産の低下はグリセ リン生成に影響したと考えられたが、 この点に ついては次章で詳細を述べる。
5-5 小括
酵母はグルコース濃度が高いほどグリセリンを高生成し 生菌数は 減少した。
また, クエン酸濃度が高いほど 酵母のグリセリン生成は増加し 生菌数は減少 した。培地中のクエン酸は菌体内のGPase活性を向上させた。
オレイン酸は焼酎酵母BAW-6のグリセリン生成を最も促進し, 次いでリノー ル酸であった。 また, パルミチン酸はグリセリン生成を抑制し た。 菌体当たり のグリセリン生成量はリノ ール酸添加区で増加し パルミチン酸およびオレイ ン酸の添 加区で はコントロールと同程度であっ た 。 オレイン酸は焼酎酵母 BAW-6以外にも9種類の醸造用酵母のグリセリン 生成を増加させ 特に焼酎酵 母(BAW-6, MK, Ko, SH-4) , ビール酵母(IFO 1167 )およびウイスキー酵 母(IF0237 3)で顕著であった。 リノール酸は焼酎酵母 ビール酵母およびウ イスキー酵母のグリセリン生成を 促進したが, 清酒酵母(K-7, K-9)およびワ イン酵母(W-3)のグリセリン生成は阻害した。
また, グリセリン生成は窒素源の影響を受け, 酵母はアミノ態窒素よりも無 機態窒素で、グリセリンを高生成した。
第6章 アミノ酸アナログ耐性株のグリセリン生成
6 - 1 緒言
酵母が生成する主要な香気成分であるイソブタノール イソアミルアルコー ル,酢酸イソアミル, ß-フェネチルアルコール司 酢酸ß-フェネチ ルなどは Ehrlich経路の他に, アミノ酸の生合成に伴って生成される。 例えば, イソブタ
ノールはバリン, イソアミルアルコールおよび酢酸イソアミルはロイシン , ß-フェネチルアル コールおよび酢酸ß-フェネチルはフェニルアラニンの生合 成に伴う産物である。
アミノ酸代謝系が変異したアミノ酸アナログ耐性株は 特定のアミノ酸生合 成のフィードバック阻害が解除され フィードバック阻害が解除されたアミノ 酸を継続して過剰生産1 0 6 )するため, そのアミノ酸に由来する香気成分を高 生成する。 イソアミルアルコールやß-フェネチルアルコールは酒類の重要 な香 気成分であることから, ロイシン やフェニルアラニンのアナログを使い, こ れらを高生成するアミノ酸のフィードバック阻害を解除した変異株 の取得 が 行われている1 0 7 -1 1 0 )。
本章では, アミノ酸生合成が活発になるとalcohol dehydrogenase (ADH)活 性が低下し, 余剰のNADHがグリセリン生成系で消費されてグリセリン生成は 増加するとの作業仮説に基づいて, 焼酎酵母BAW-6のアミノ酸アナログ耐性 変異株からグリセリン高生成株の取得を試みた結果について述べる。
6-2 実験方法
酵母
第2章に示した大麦焼酎用酵母BAW-6を親株として使用した。
菌体内alcohol dehydrogenase (ADH)活性
菌体内ADH活性に及ぼす培地窒素源の影響を調べた実験は, 10%グルコース,
1.17% Yeast Carbon Baseを基本培地とし, 窒素源として, 0.50/0の硫安, 塩化アン
モニウム, トリフトンおよびカザミノ酸の中からいずれか一種類を加えた培地
を用い,
250Cで酵母を培養した。アミノ酸アナログ耐性株の 菌体内ADH活性を調べた実験は, YEPD(2)培地を 用い, 250Cで酵母を培養した。 菌体内酵素液は培養液から第5章5-2に示し た方法により調製し、 ADH活性測定に用いた。
ADH活性の測定 は60mMビロリン酸ナトリウム緩衝液(pH8.5), 0.7Mエタ ノールおよび0.5mM NAD+になるょっに濃度を 調整した反応液3mlに10倍 希釈した酵素液O.lmlを添加し , 300Cで30分間反応させ NAD+から生成す る NADH量(μmol)を340nmでのサンプルとブランクとの吸光度差から求めたo ADH活性 は1分間当たり1μmolのNADHが生成される活性を1unitとし, 酵母菌体 内タンパク質lmg当たりで表した。
酎
性株の分離焼酎酵母をYEPD(2)培地で30oC, 24時間培養し, 対数増殖期の細胞を滅菌水
で3回洗浄後, ethy 1 methanesulfonate (EMS)で1時間振とう処理し, チオ硫酸 ナトリウムで中和後, 各種スクリ ーニング培地に塗布し, 300Cで培養後, 増殖 したコロニーを耐性株として分離した。 スクリーニング培地は, 2%グルコース,
m寒天, 0.67% Yeast Nitrogen Base w/o amino acidを基本培地とし, 5ふ5-trifluoro leucine
(TFL)
200ppm, p-fluoro DL-ph e nylalanine (FPA) 500ppm, canavanine(CAN)
100ppm, cerulenin (CER) 50ppmをそれぞれ培地に添加し, 使用した。グリセリン生成試験
グリセリン生成試験はYEPD(10)培地(100/0グルコース , 10/0酵母エキス , mポリペプトン)を用い 300Cで4日間発酵させて行った。
香気生成試験
香気生成培地①(10%グルコース, 0.65% Yeast Nitrogen Base w/o amino acid) および香気生成培地③(10%グルコース, 1.170/0 Yeast Carbon Base, 0.5%カザミ ノ酸)に予め前培養した酵母液を2%接種し, 300Cで4日開発酵させた。 発酵後,
TFL耐性株については, 培養液5gをバイアルに取り, 密封後800Cで40分間保 持し, ヘッドスペースガスlmlをヘッドスペースガス分析システム(島津製作 所, HSS-GC)に供与した。 GC分析は次に示す方法で、行った。 すなわち, カラ ムにDBWAX(ゆ0.25mmX 30m, 膜厚0.25μm, J&W)を使用し, 700Cで5分間 保持後, 10oC/minで1700Cまで昇温させ, 5分保持する条件で行った。 注入口温 度, 検出器温度は2000Cとし, 検出器はFIDを使用した。 キャリア ーガスはヘリ
ウムを使用し, 流速0.7ml/min, スプリット比10: 1で行った。
FPA耐性株は培養液5gに内標準物質として10g/1の安息香酸エチノレ50μlを添加 し, エーテル抽出後, 抽出液を濃縮し, 1μlをGC (目立, 263-80型)に供し,
ß-フエ不チルアルコールおよび酢酸ß-フェネチルの分析を行った。 カラムは
G初o (ゆ1.2mm X 40m, 膜厚1μm, 化学品検査協会)を用い, カ ラム温度
1000Cから30C/minで、2000Cまで昇温させるプログラムで、行った。 注入口温度,
検出器温度は3000Cとし, 検出器はFIDを使用した。 キャリアーガスはヘリ ウムを使用し , 流速40ml/min, サンプル注入量2μlとした。
分析
もろみ中のグリセリン, グルコースの定量は第1章1-2に, エタノールの 定量は第3章3-2に示した方法で行った。
6-3 実験結果
6-3-1 窒素源が異なる培地での酵母菌体内
alcohol dehydrogenase �舌性
これまで, ADH活性が低下するとNADHが細胞内で過剰になり, これが dihydroxyacetonephosphateからglycerol-3-phosphateへの還元反応で、消費されるた
めに酵母のグリセリン生成は増加する4 2 )と報告されている。 第5章5-3- 3で硫安, 塩化アンモニウムを窒素源として用いた場合, 酵母のグリセリン生
成が増加したことを述べた。 そこで窒素源の異なる培地での酵母菌体内alcohol dehydrogenase (ADH)の 活性変化を調べたo Fig.6-1に示すように, ADH活性は 発酵中常にカザミノ酸が最も高く, 次いでトリプトン, 塩化アンモニウム, 硫 安の順番であった。一方, グリセリン生成量は硫安が最も高く, 次いで塩化ア ンモニウム, トリフトン, カザミノ酸の順番であった。すなわち, 酵母のADH 活性が低いほどグリセリン生成量が多かった。以上のように, 窒素源は酵母菌 体内のADH活性に影響し, その結果グリセリン生成量に違いが生じたものと考 えられた。
アミノ態窒素を窒素源として使用した場合 , アミノ酸が直接菌体内に取り込 まれ, 取り込まれたアミノ酸のフィードバック阻害により酵母菌体内でのアミ ノ酸生合成が押さえられるのに対し、 無機態窒素を窒素源として使用した場合,
グルコースからのアミノ酸生合 成が盛んになるという違いがある。 このことか ら, 著者は窒素源によって酵母のグリセリン生成に違いが生じたのは菌体内で のアミノ酸生合成が影響していると考えた(Fig.6-2)。すなわち, 無機態窒素添 加区では酵母が自らアミノ酸を生合成する分だけエタノール生成に回るグルコー ス量が減少し, ADH活性は低下する。 その結果, エタノール生成系で消費され るはずのNADHが過剰になり, これがグリセリン生成系で消費されるためにグ リセリン生成が向上したのではないかということである。
(一\窃)一O」ωo去の
5
4
3
2
a)
b)
0.8
民U
λ斗
内/』
ハU
ハU
ハU〈CZWUFO』且lmrchE3V〉t〉ZOUZO《
。
5 4
Fermentation day 3
2 5
4 Fermentation day
3 2
Fig. 6-1.
Effects of nitrogen source on
ADHactivity (a) and glycerol formation (b) of
shochuyeast
BA W -6.Symbols: 0, Ammonium sulfate;ム, Ammonium chloride;・, Tryptone;,. , Casamino acid.
Higher alcohol ___ Higher
_____acetates � alcohols
�印刷c acid ー圭一 A似aldehyde α-Keto acids ト
l
『司司司 ...- Ethanol
Glucose
-�メ』
Alcohol dehydrogenase
Feed back inhibition Amino acids
Fig.6-2. Biosynthetic pathway of ethanol, glycerol and amino acids in Saccharomyces cerevisiae.
6-3-2 各種耐性株のグリセリン生成
6-3-1に述べたように, アミノ酸生合成が活発になる とグリセリン生成 が増加するとすれば, アミノ酸を継続的に生合成するアミノ酸アナログ耐性株 からグリセリン高生成株を取得する ことが可能と考えられた。 そこで、 大麦焼 酎用酵母BAW-6を親株として アミノ 酸代謝系変異株のアミノ 酸ア ナログ
(TFL,
FPA)耐性株を取得し これらの耐性株のグリセリン生成を検討した。また, アルギニンの取り込みが欠損したCAN耐性株1 1 1 • 1 1 2 )および脂肪酸 生合成系の一部が変異したCER耐性株1 1 3 )についても比較したO すべての 変異株ともEMS処理後, それぞれのスクリーニング培地に塗布し, T孔および FPAの耐性株はそれぞれ50株, CANおよびCERの耐性株はそれぞれ 100株を取 得した。 Table6・1に各種耐性株のグリセリンおよびエタノールの生成量を示す。
グリセリン生成量の平均値はTFL耐性株で2.10g/l, FPA耐性株で、2.07g/l, CANお よびCERの耐性株でそれぞれ1.60g!l , 1. 59g/1で、あった。 エタノールはTFLおよび FPAの耐性株の方がCANおよびCERの耐性株および親株より 0.10/0ほど低かった。
Fig.6-3に親株BAW-6に対する各耐性株のグリセリンの増加率を示す。 CANお よびCERの耐性株では親株よりグリセリン生成が低下した株が多かったのに対 し, TFLおよびFPAの耐性株は高頻度でグリセリンを高生成した。 すなわち,
T札耐性株では取得した50株中48株が親株よりグリセリンを多く生成し, 最優 秀株は親株のおよそ1.8倍であった。 FPA耐性株では取得した50株中42株が親株 よりもグリセリン生成量が多く, 最優秀株はTFL耐性株と同様に親株のおよそ
Table 6-1. Glycerol formation and ethanol formation of amino acid analogue strains of shochu yeast BA W-6.
Glycerol (g/l) Ethanol (%)
Mutant a
Min. Max. Ave. Min. Max.
TFL 1.54 2.91 2.10 5.0 5.5
FPA 1.52 2.95 2.07 4.9 5.5
CAN 1.28 1.89 1.60 5.1 5.6
CER 1.23 2.17 1.59 5.0 5.5
BAW-6 (parent) 1.61
a TFL; 5,5,5-trifluro-leucine, FPA; p-fluoro DL-phenylalanine,
CAN; canavanine, CER; cerulenin.
Ave.
5.2 5.2
5.3 5.3 5.3
c) CAN b)FPA
d) CER a)TFL
20
10
nunu
nu
nu
nu
2 1
4 2
℃ω芯一0255窃』O』ω岳ヒコZ
。
。 40
20
。 60 80
Glycerol increased from the parent strain (0/0) 40
20
Fig. 6-3. Glycerol formation of arnino acid analogue strain of shochu yeast BA W -6.
1.8倍であった。一方, CANおよびCERの耐性株では, 親株よりもグリセリンを
高生成した株はそれぞれ100株中10株以下であった 。以上のように, アミノ酸
代謝系が変異したアミノ酸アナログ耐性株は親株よりもグリセリンを高生成す る株が高い確率で取得できた。
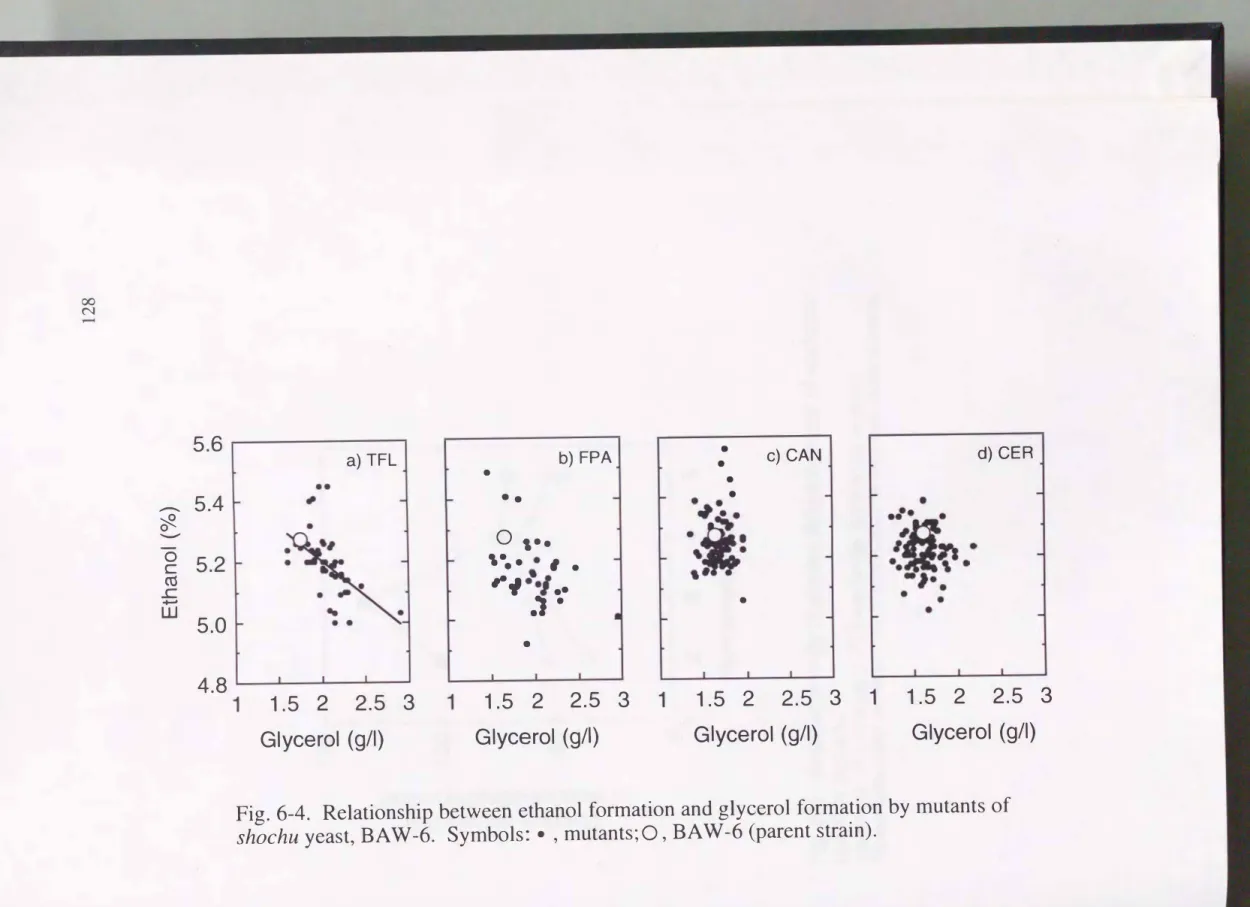
6-3-3 エタノールとグリセリンとの関係
Fig.6-4にYEPD(10)培地で、のエタノール生成量とグリセリン生成量との関係を 示す。CANおよびCERの耐性株ではエタノールとグリセリンとの明確な関係が 認められなかったのに対し, アミノ酸代謝系変異株であるTFLおよびFPAの耐 性株ではグリセリン生成が多い株ほどエタノール生成は低下し エタノール生 成とグリセリン生成とは負の相関が見られた。特にTFL耐性株は(6-1)式に回 帰することができた。
Ethanol (0/0) = - 0. 23 X glycerol (g/l) + 5.66 r=0.65 (6-1)
Fig.6-5にTFLおよびFPAの耐性株でグリセリンを高生成したそれぞれ8株の ADH活性の経時変化 を示す。発酵期間中, TFLおよびFPAの耐性株のADH活 性は親株BAW-6のそれよりも低く推移し, 特に2, 3日目では明らかな違いが見 られた。 また 親株は3日目に最大活性を示し 4. 5日目には急激に低下した のに対し, TFLおよびFPAの耐性株は4日目に最大活性を示すまで徐々に増加し,
貯性の変化にも違いが見られた。耐性株の最大活性は親株のそれのおよそ50%
まで低下した。以上のことから、 ADH活性の低下がエタノール生成低下をもた
5.6 d) CER
ー
ー
ー
...l.
2.5
」圃
2
ょ
1.5
ー
ト- 42.5
ム
2
....L
1.5 ト
•
3 Glycerol
(g/I)
3 Glycerol
(g/I)
3 2.5 Glycerol
(g/り
Fig.
6-4.Relationship between ethanol formation and glycerol formation by mutants of
shochuyeast,
BA W -6.Symbols:・, mutants; 0,
BA W -6(parent strain).
ト .
'11 �・
ト _'
G
配れ ・う. 7 ・
c) CAN
ー ー
-・
..や縛
•
ト ト b)FPA
ー ー
•
••
。‘旬
、..._. , •
,." .-;・i
..3・
•
ト
←
a) TFし 5.4
5.2
(ポ)一0CC工日
5.0 トー
4.8 2.5 3 1.5 2
Glycerol
(g/I)
1.5 2
150
100
50
ωωccωOO』刀、戸工ω℃一O工00一の平O
(JCb一〉一日ocω〉一芯一
ω江
。 4 5
Fermentation day 3
2
Fig.6-5. Relative activity of alcohol dehydrogenase of analogue reslstant mutants.
Symbols: 0, TFL resistant�ム, FP A resistant�・,BAW-6 (parent strain).
These data represent the mean value of results obtained from eight different strains.
らし, その結果, アミノ酸代謝系変異株はグリセリンを高生成したものと推定 された。
6-3-4 アミノ酸生合成系に由来する香気成分と
グリセリンとの関係
TFL耐性株はロイシン, FPA耐性株はフェニルアラニンをそれぞれ高生成し,
これに由来するイソアミルアルコールおよびß-フェネチルアルコールの生成が 増加する。 前項で述べたように T孔およびFPAの 耐性株のADH活性は親株 よりも低下し, 著者はこれがアミノ酸生合成が活発になったためと考えて いる。 そこで、 イソアミルアルコールおよびß-フェネチルアルコールの生成 量とグリセリン生成 量と の関連を調べた。 最小培 地を用いた香気 生成培地
①での結果をFig.6-6に示すO グリセリン生成量の増大に伴って, TFL耐性株で はイソアミルアルコールが増加し, FPA耐性株ではß-フェネチルアル コール が増加し, それ ぞれ正の相関が見られた。 香気成分とグリセリンとの関係は TFL耐性株で(6-2)式, FPA耐性株で(6-3)式 の直線にそれぞれ回帰できた。
Isoamyl alcohol (mg/l) = 75 X glycerol (g/l) - 225 r = 0.77 (6-2)
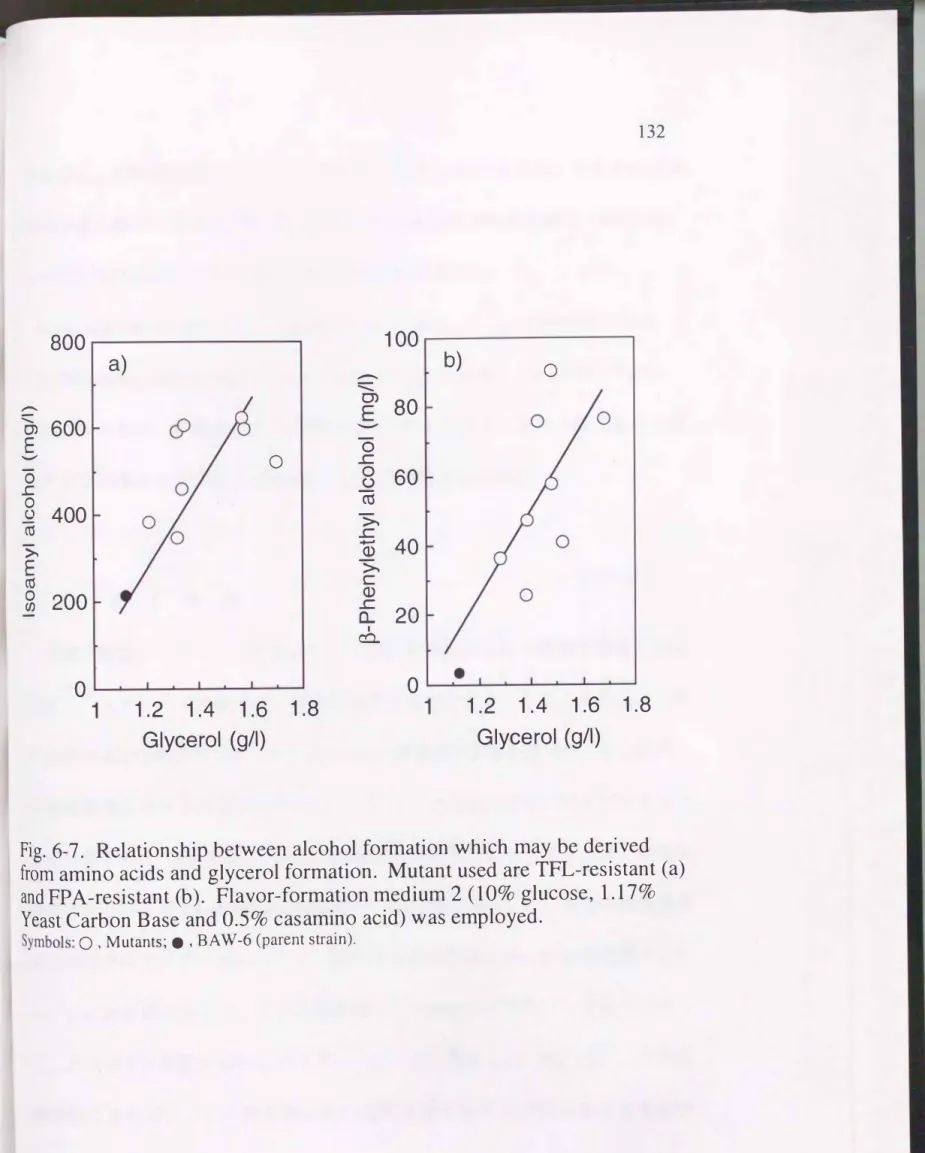
。-Phenylethyl alcohol (mg/l) = 220 X glycerol (g/l) - 845 r = 0.72 (6-3) 一方, 香気生成培地⑦での結果をFig.6-7 �こ示すO 香気生成培地①と同様に,
グリセリン生成量の 増大に伴ってTFL耐性株ではイソアミルアルコール生成量
600
a) b)
".-.、
、0、、3
,・句、 E
ε。】 400 。 2� 400 r 。。
工0 0
/
0国 エ4
〉
圃二" ω
、
ω
ご価 。200 吾200
ωエニa_
• 巴ユ
自 。
4
56
74
56
7Glycerol (g/I) Glycerol (g/I)
Fig.6-6. Relationship between alcohol formation which may be derived from amino acids and glycerol formation. Mutants used are TFL-resistant
(a)
and FPA-resistant (b). Flavor-formation medium 1 (10% glucose, 0.65九
Yeast Nitrogen Base w/o amino acid) was employed . .syrnbols: 0, Mutants;・, BA W-6 (parent strain).800 100
b) 。
80
60
40
20 (一\。ε)一O工00一ω一弘£ω一KACω工止,c
。 a)
600
400
200 (-bE)一O工00一国王ECOω一
1.8
Fig.
6-7.
Relationship between alcohol formation which may be derived from amino acids and glycerol formation. Mutant used are TFL-resistant (a) and FPA-resistant (b). Flavor-formation medium 2(10%
glucose,1.179も
Yeast Carbon Base and
0.5%
casamino acid) was employed.�ymbols: 0 , Mutants;・, BA W -6 (parent strain).
1.4 1.6
Glycerol (g/I)
1.2
。 •
1.2 1.4 1.6 1.8
Glycerol (g/I)
。
が増加し, FPA耐性株ではß-フェネチルアルコールが増加し, それぞれ正の
相関が見られたO 香気成分とグリセリンとの関係はTFL耐性株で(6-4)式,
FPA耐性株で、(6-5)式の直線にそれぞれ回帰できた。
Isoamyl alcohol (mg/l) = 522 X glycerol (g/l) - 243 ß-Phenylethyl alcohol (mg/l) = 188 X glycerol (g/l) - 145
r = 0.73 (6-4) r = 0.77 (6-5) 以上のように, 培地中のアミノ酸の有無にかかわらず, アミノ酸生合成に由 来する香気成分とグリセリン生成とには正の相関が見られた。
6-4 考 察
前章で酵母のグリセリン生成がアミノ態窒素添加区 に比べ無機態窒素添加区 で多く, エタノ ール生産は逆に無機態窒素培地で少なかったことから, はじめ に菌体内ADH活性とグリセリン生成に及ぼす窒素源の影響を調べた。 その結果,
無機態窒素添加区でのADH活性は低く, さらにADH活性が低いほどグリセリン 生成は多かった(Fig.6-1)。 これはADH活性の低下によってグリセリンが高生 成されたという点で、Lutstorf and Megnet4 2 )の推論と符合した。 著者は無機態窒 素添加区ではグルコースからアミノ酸の生合成が活発になってADH活性および エタノール生成が低下し, その結果蓄積したNADHがグリセリン生成系に回っ て, グリセリン生成が高められるのではないかと推論した(Fig.6-2)。 この作 業仮説によれば, アミノ酸を継続的に過剰生成するアミノ酸代謝系変異株は
ADH活性が低下し, さらにグリセリンを高生成すると考えられ, アミノ酸アナ ログ耐性株からグリセリン高生成株の 取得を試みた。
焼酎酵母BAW-6を親株として4種類の耐性変異株を取得し, グリセリン生成 を比較したところ, アミノ酸代謝系が変異したT孔およびFPAの耐性株のみ高 い頻度で グリセリン高生成株が得 られ た(Fig.6-3)。 また, TFLおよびFPAの 耐性株においても グリセリン生成が多い株ほどエタノール生成 の低下 が見られ (Fig.6-4) , さらに菌体内ADH活性 は親株よりも低く推移した(Fig.6-5)。
取得したアミノ酸アナログ、耐性株の中にはEMS変異処理によりADHアイソザ イムが一部欠損し, そのためにADH活性が低下した株が含まれる可能性もある。
しかし, アミノ酸生合成が原因となってADH活性が低下し, そのためにグリセ リン生成が向上したのであれば フィードバック阻害を解除したアミノ酸に由 来する高級アルコールとグリセリン生成とは正の相関が見られるはずである。
そこで窒素源が異なる香気生成培地①および香気生成培地②で アミノ酸に由来 するイソアミルアルコールやß-フェネチル アルコー ルの 生成 とグリセリン 生成との関係を調べた(Fig.6-6, Fig.6-7)。酵母は香気生成培地①の窒素源 が硫安であることから培地からアミノ酸を取り込めず 香気生成培地② の窒素 源がカザミノ酸であることから培地からアミノ酸を取り込むことができる。 い ずれの条件であっても アミノ酸に由来する香気成分とグリセリン生成とは正の 相関が見られた。 以上の ように, フィードパック阻害が 解除されたアミノ酸に
由来する香気成分がグリセリンと正の相関が見られたことから, アミノ酸生合
成がADH活性の低下に影響し, その結果グリセリンが高生成されたものと推察 されたO
以上の結果をTable6-2 �こまとめた。 硫安, 塩化アンモニウムを窒素源として 用いた場合, カザミノ酸, トリプトンを窒素源として 用いた場合よりもADH活 性は低く, グリセリン生成は多かった。 この結果から, 著者はグリセリン生成 に影響するADH活性の違い42)はアミノ酸生合成に関連している, すなわち アミノ酸生合成が活発になるとADH活性が低下し その結果グリセリンが高生 成されたと考えた(Pig.6-2)。 この仮説に基づいて, アミノ酸代謝系が変異し たアミノ酸アナログ耐性株からグリセリン高生成株を分離した。 また, これは アミノ酸アナログ耐性株がアミノ酸を継続的に生合成した1 0 7 - 1 1 0 )ためにグ リセリンを高生成したものと推定された。
アミノ酸アナログ耐性株を使用して焼酎醸造を行った場合, アミノ酸に対応 したアルコールや酢酸エステルのもろみ中の濃度が増加し, 官能に特徴ある焼 酎が得られる1 5 )と報告されている。 アミノ酸アナログ耐性株からグリセリン を高生成する株が高頻度で得られることから, これまで醸造に応用されてきた TFLおよびFPAの耐性株もグリセリンを高生成する株であった可能性が高い。
このことから, アミノ酸アナログ耐性株を用いた焼酎醸造はもろみ中の香気成 分を増加させ, さらにグリセリン濃度の増加により香気成分の留出を高めると いう2つの効果があり, その結果特徴ある焼酎が得られるものと考えられた。
Table 6-2. Strain phenotype and nitrogen source which influence glycerol formation.
Conditions Biosynthesis b C d d
of Alcohol Ethanol Glycerol
amino acid dehydrogenas formation formation
Unknown 42)
Unknown +42) Alcohol dehydrogenase-partial
deficient mutant 42)
Ammonium sulfate a
Ammonium chloride ++ +
Casamino acid a
Tryptone 十 + +
+1 10-1 13)
Amino acid analogue mutants +
a These results was obtained from Fig. 6-1.
b + + ... active, + .. . active only some arnino acids, :t • • • inactive.
c + ... high,一 ...low.
d + ・ ・ ・ mcrease,一 ... decrease.
一般にADH活性の低下した株は増殖力あるいは発酵力が低下している可能性
が高く , 実用上問題である。 著者は塩化ナトリウム耐性を指標にして, ADH
活性は親株と遜色なく, GPase活性が高まったグリセリン高生成株を取得した
が, これについては次章で述べる。
6-5 小括
ADH活性はアミノ態窒素添加区より無機態窒素添加区で低 く グリセリン生
成は無機態窒素添加区で、高かった。 この結果 から アミノ酸生合成が活発にな るとADH活性およびエタノール生成が低下し, グリセリン生成が向上するので はないかと推論し アミノ酸代 謝系が変異したアミノ酸アナログ耐性株のグリ セリン生成を調べた。 その結果, T礼およびFPAの耐性株 から高い確率で 親株 よりもグリセリ ンを高生成する株が得られ 最優秀株は両耐性株とも親株のお よそ1.8倍のグリセリンを生成した。 また 対象として行ったCANおよびCERの 耐性株は親株より もグリ セリン生成が低下した株が多かった。 TFLおよびFPA の耐性株ではグリセリン生成が多い株ほどエタノール生成量は 低下する傾向に あり, 菌体内ADH活性は親株よりも低く 推移した。 また, アミノ酸代謝系変異 株のグリセリン生成量はフィードバック阻害が解除されたアミノ酸に由来する イソアミルアルコールやß-フエネチルアルコールの生成量とそれぞれ正の相関 が認められた。 以上の結果, アミノ酸代謝系変異株は親株よりADH活性が低下
し, その結果グリセリンを高生成したが, これは変異株がフィードバック阻害
が解除されたいくつかのアミノ酸を継続的に生合成したためと推察されたO
第7章 塩化ナトリウム耐性を強化した 焼酎酵母 からの
グリセリン高生成株の取得
7 - 1 緒言
前章でアミノ酸代謝系が変異した アミノ酸アナログ耐性株 は, alc oh ol
dehydrogen ase (ADH )活性の低下によりグリセリンを高生成 した ことを述べ
たO焼酎酵母に求められる基本的性質はクエン酸耐性があり, アルコール生産 性が高いことで あることから, ADH 活性が低下 した株は焼酎醸造においては実 用的ではないと考えられた 。 これまで, グリセリンを高生成する変異株として,
allyl alcohol耐性39,41-44)あるいはpyrazole耐性3 9 , 4 1 )株 などに見られる ADHアイソザイム一部欠損株, 呼吸欠損株3 9 , 4 1 ) が報告されている。 しかし,
前述した ようにADHアイソザイム一部欠損株はアルコール生産が低下すること 39), 呼吸欠損株は増殖が遅れることなど の理由から, 焼酎醸造上好ま しくな
いと考えられた。
一方, Zygosaccharomyces rouxii, Debariomyces hanseniiは塩に対する耐性を有 しており, Saccharomyces cereνlSlaeよりもグリセリンを高生成する1 1 4司1 2 0 ) ことが知られている。 また , Saccharomyces cereνlSlaeも塩存在下でグリセリン を高生成する1 0 3 , 1 2ト1 2 4 ) ことが報告されているo Matsutaniら12 5 ) は,
清酒酵母協会7号(K-7, Saccharomyces cerevisiae)からethyl methanesulfonate (EMS)処理によって塩化ナトリウム耐性株 を取得し, 電子顕微鏡観察の結果
塩化ナトリウムに対する耐性は細胞が小さくなることで得られることを報告し ている。 しかしながら, グリセリンなどの発酵生産物の濃度向上を目的として,
Saccharomyces cereνlSlaeの塩化ナトリウム耐性を強化したという報告はない。
本章は, 塩化ナトリウム耐性を強化したおccharomyces cereνlSlaeがグリセリ ンを高生成することを明らかにし さらに18%塩化ナトリウム耐性株TK-2の醸 造特性について述べる。
7-2 実験方法
原料大麦
第l章1 -2に示した。
大麦麹
第2章2-2に示した。
酵母
第2章2-2に示した大麦焼酎用酵母BAW-6を使用した。
酵母菌体内酵素およびタンパク質の調製
YEPD(5)培地(5%グルコース, 10/0酵母エキス, 20/0ポリペプトン) , 300Cで 振とう培養した培養液から, 第 5章5-2に示した方法で酵母菌体内酵素およ びタンパク質を調製した。
Alcohol dehydrogenase (ADH)活性の測定
ADH活性は第6章6-2に示した方法で測定したO
Glycerophosphatase ( G Pase)活性の測定
GPase活性は第5章5-2に示した方法で測定した。
Glycerol kinase ( G Kase)活性の測定
GKase活性は第5章5-2に示した方法で測定した。
酵母菌体内タンパク質の測定
酵母菌体内タンパク質は第5章5-2に示した方法で測定した。
グリセリン生成試験
グリセリン生成試験はYEPD(5)培地を用い, 300Cで4日開発酵させた後, グ リセリン濃度を測定した。 クエン酸存在下でのグリセリン生成を調べた実験で は, YEPD(10)培地(10%グルコース, 1%酵母エキス, 2%ポリペプトン)を基 本培地として0.3%クエン酸緩衝液(pH5)を用いて調製したCit-YEPD(10)培 地を用い, 300Cで4日開発酵させた後, グリセリン濃度を測定した。 コントロー ル(YEPD(10)培地 )は1Mの塩酸でpH5に調整した。
増殖特性
バイオフォトレコーダー(アドバンテック, TC-I06)を用い, 最小培地(2%グ ルコース, 0.65% Yeast Nitrogen Base w/o amino acid)を基本培地として増殖特性