筑波大学大学院博士課程
システム情報工学研究科修士論文
培養神経回路網の長期的観測による
活動特性の経時的変化の解析
向井 嘉崇
(コンピュータサイエンス専攻)
指導教官 椎名 毅
2002 年 2 月
概
要
大脳皮質神経細胞は発達段階において同期バースト現象と呼ばれる全体が同期した自発活 動を示し、これが適切な回路網の形成に大きく関与していると考えられている。この変化を 長期的に連続的に観測するため、測定基板をインキュベータ内に設置し、64 微小電極アレイ 上でラット大脳皮質細胞を培養しながら、同時に活動電位記録を行うことのできるシステム を製作した。培養液はシリンジポンプによって0.1ml/h にて緩やかに潅流し、非常に安定し た溶液状態が維持されている。さらにオンラインにおいて、時系列電位データから活動電位 部分のみを抽出し保存することデータ圧縮を行った。このシステムにおいて5DIV(Days in vitro)から一ヶ月以上に渡る自発活動電位の連続計測に成功し、大脳皮質回路網の成長にお ける詳細な変化を捉えることが可能となった。 バーストの検出は、50msec 窓内の全電極でのスパイク数をカウントし、窓を移動させる ことによって、発火頻度波形を算出した後、閾値を越える部分をバーストとすることで行っ た。さらに成長によるバーストの記録エリアにおける広がりの推移を調査するため、バース トに含まれる各電極のスパイク数を空間パターンとし、k-means 法によりクラスタリングを 行った。 この結果、測定当初から得られていた成長段階での周期的で再現性に良い同期バーストが 培養2 週間目においてに急激な減衰を示すと共に、複雑な空間パターンを示すバーストが活 性化することが明らかになった。これは回路網形成期から回路網成熟期への移行を示すもの と考えられる。目次
第1 章 序論...1 第2 章 培養神経回路網...3 2.1 神経回路網特性の研究...3 2.1.1 神経細胞の基本的性質...3 2.1.1.1 活動電位発生メカニズム...4 2.1.1.2 シナプス後電位...4 2.1.1.3 神経細胞への薬理作用...5 2.1.2 同期バースト現象の発生メカニズム...7 2.2 培養神経回路網の発達過程...8 2.3 微小電極アレイによる活動電位計測...9 第3 章 神経細胞発火パターン検出...10 3.1 活動電位データ...11 3.2 スパイク振幅・幅を用いたスパイクソーティング...15 3.2.1 振幅(Amp)の導出...16 3.2.2 幅(width)の導出...17 3.2.3 スパイク幅と振幅による2次元クラスタリング...18 3.3 細胞内信号伝播経路推定による発火パターン検出法...19 3.3.1 スパイクの検出と切り出し...20 3.3.2 相互マッチングによるスパイクソーティング...20 3.3.3 発火パターンマッチングによる伝搬経路推定...21 3.3.4 神経細胞の発火パターン検出と長期にわたる観測...22 3.4 解析結果...23 3.5 まとめ...28 第4 章 回路網活動長期モニタリング...29 4.1 実験...30 4.1.1 試料作製...30 4.1.2 培養液構成...30 4.1.3 多電極アレイ基板...30 4.1.4 培養液潅流システム...32 4.1.5 電位記録システム...35 ハードウェア...374.1.5.5 データ圧縮...37 4.1.5.6 インキュベータノイズ除去...39 4.2 同期バースト現象解析...41 4.2.1 バーストの検出...41 4.2.2 バーストのクラスタリング...43 4.3 結果...46 4.3.1 長期的電位測定システムの安定性...47 4.3.2 活動電位波形...49 4.3.3 スパイク数の推移...54 4.3.4 バースト間隔の推移...54 4.3.5 バースト内スパイク数の推移...55 4.3.6 バースト幅の推移...55 4.3.7 バースト空間パターンの推移...60 4.3.8 同期バースト現象の減衰...60 4.3.9 電極基板上の培養神経細胞...61 4.4 考察...65 4.5 まとめ...66 第5 章 結論...67 謝辞...68 参考文献...69 付録...72 A NTTmain データ形式...72 B 圧縮ファイル mlt のデータ形式...73
図目次
図 3-1 微小電極アレイ基盤(1 ブロック構成)...12 図 3-2 電極番号(1 ブロック構成)...12 図 3-3 培養液交換システム...13 図 3-4 活動電位波形...14 図 3-5 波形のパラメータの名称...15 図 3-6 うねりによる同スパイクの違い...16 図 3-7 スパイク幅の導出...17 図 3-8 クラスタリングによる波形の分離の例...18 図 3-9 多電極を用いたスパイクソーティングの流れ...19 図 3-10 幅と振幅によるクラスタリング結果 ...24 図 3-11 スパイクソーティング結果 ...25 図 3-12 分離された活動電位波形...26 図 3-12 推定された神経細胞の信号伝搬の様子...26 図 3-13 発火間隔分布の推移...27 図 4-1 微小電極アレイ基板(2 ブロック構成)...31 図 4-2 電極アレイ付シャーレ...31 図 4-3 電極番号(2 ブロック構成)...31 図 4-4 培養液潅流システム...33 図 4-5 培養液潅流システム(模式図)...34 図 4-6 長期活動電位記録システム...36 図 4-7 ノイズ標準偏差算出...37 図 4-8 スパイクパラメータと抽出領域...38 図 4-9 インキュベータノイズ...40 図 4-10 発火頻度算出法模式図 ...44 図 4-11 バーストのパラメータ...45 図 4-12 活動電位記録過程...46 図 4-13 ノイズ標準偏差の推移 ...48 図 4-14 ノイズ標準偏差の推移(G0,D3 電極) ...48 図 4-15 5DIV から 12DIV までの活動電位波形...50 図 4-16 13DIV から 20DIV までの活動電位波形...51 図 4-17 23DIV から 51DIV までの活動電位波形...52 図 4-18 自発活動電位波形...53図 4-19 64 電極別スパイク数の推移...56 図 4-20 全電極のスパイク数の推移...57 図 4-21 スパイク数の推移(3 次元表示)...58 図 4-22 バースト間隔の推移...59 図 4-23 バースト内のスパイク数の推移...59 図 4-24 バースト幅の推移...59 図 4-25 バーストの空間パターンの推移...62 図 4-26 同期バーストの減衰...63 図 4-27 電極基板上の培養神経回路網...64
第1章
序論
脳は極めて高機能な情報処理器官である。脳に働いている神経細胞(ニューロン)の数は 1012個という膨大なものであり、その回路網は1015個ものシナプスを仲介する接点で構成さ れ複雑な処理を可能としている。特に脳がノイマン型コンピュータと大きく異なる点は自身 の活動に依存して機能が変化し、処理と学習を同時に行うことができる、超並列なメモリベ ース型のアーキテクチャーであるということである。一般的には神経回路網は構成する素子 である神経細胞が活動すると同時に活動した神経細胞同士を結合しているシナプス強度が変 化することで、処理と同時に記憶・学習が行われる。特に短期記憶を司る海馬や高度な情報 処理を可能とする大脳皮質等の組織は高頻度な刺激を与えることで、長期増強や長期抑制と いった可塑性を引き起こすことが明らかにされている[1][2][3]。ヒトの脳では胎児の2から3ヶ 月頃から大脳皮質の形成が始まり、4ヶ月目から急激な神経突起の伸長とシナプス形成が進 むことで大脳はさらに増大していく。このような外見的な脳の成長と共にネットワークレベ ルで起こっている学習機能の獲得がどのような形で進行するのかは脳形態学的に非常に興味 深いテーマである。回路網自体が、自分自身の活動の結果に依存し、時系列的に自己組織化 され進化していくと考えられているが[4]、これらのメカニズムが明らかとなれば、人工的に 脳の形成を制御し、構築することができる可能性を秘めている。 これらの解明に対して、脳波計測などのin vivo での測定では、対象が膨大な数の神経細 胞による複雑な回路網で解析が困難であり、幼児期からの測定は難しい。またガラス電極に よる方法や光学的に染色する方法では細胞組織の損傷は避けられず、長期的な計測は困難で ある。脳組織スライスを対象とした場合、胎児段階から採取することが困難であると共に神 経細胞の集合電位として計測されるため個々の神経細胞レベルでの解析に適さない。 そこで、本研究ではラット大脳皮質から単離した神経細胞を微小電極アレイ上に培養した 培養神経回路網を実験対象として用いることにした。埋め込み型微小電極アレイを利用して 細胞外信号を記録する手法は1979 年に Gross により開発されて以来、他の測定手法と組み 合わせることができるという汎用性の面から多くの研究に用いられるようになった。また、 培養された電極基板上の低密度な2次元平面状の回路網はその構造自体はランダムとなるが、 互いにシナプス結合を形成し、基本的な回路網の機能は保持しているとされている。特にこ極数分の空間分解能を有した神経回路電位測定が可能とできることである[5][6]。 これまで、大脳皮質神経回路網は神経突起が成長することで互いにシナプス結合を行うと 共に、神経細胞のランダムな発火から、同期バースト現象(synchronized bursting)と呼ばれ る、周期的な回路網全域的に非常に同期した一連の発火を生じるようになること、さらに成 熟段階にでは複雑な発火パターンを示すことが報告されている[7]。しかしながら、これらの 報告は微小電極アレイを用いても滅菌及び溶液状態の維持の点から、同一回路網の長期観測 が難しいために、培養期間が異なる別々の回路網からの統計的解釈での結果であった。同期 バースト現象を含め、回路網の発達における特性変化を詳細に調査するには、同一の回路網 において長期的に培養しながらの連続的な電位記録することが必須であると考えられる。こ のような背景の下、近年長期測定システムに関する報告[8][9]がされているが、未だ詳細な発 火パターンの推移を計測した報告は無い。 また、微小電極アレイによる測定は細胞外の膜電位を測定するため、多細胞の電位が折り 重なり得られることから、培養環境中のバースト発火では一神経細胞の活動電位を検出する ことが難しく、個々の神経細胞における発火パターンが成長によってどのように変化するの かを解析することは困難であるという問題がある。しかしながら、個々の神経細胞レベルで の発火パターンとその成長による推移を得ることができれば、脳形態学的に大きな知見を得 ることできる。 よって、本研究ではまず、微小電極アレイ基板上に培養された回路網から個々の神経細胞 の発火パターンを精度良く検出する手法を提案し、50 日培養された成熟段階の回路網から、 抽出された個々の神経細胞の発火パターンが日々の培養によってどのように推移するか調査 した。この結果、成熟段階では大きな変化は無いが、培養液交換による発火パターンの揺ら ぎが観測された。 そこで、測定系をインキュベータ内へ設置するとともに、回路網への培養液潅流システム の構築、及び活動電位データ圧縮を行うことで、同一回路網の成長に伴う特性変化を長期的 に安定して調査することを試みた。このシステムにおいて、培養開始後5日目(5 days in vitro (DIV))から 50 日目に渡る自発活動電位の連続計測に成功し、回路網形成時における発火パ ターンの変化を詳細に捉えることが可能となった。その結果、5DIV から同期したバースト が得られ、それに賛同するスパイク数が増加すると共に同期性が向上し、13DIV でピークを 迎えた。しかし、その後、減衰に転じ 15DIV 付近で消失すると共に、それとは全く別な複 雑なパターンのバーストが出現し、それに連なるスパイク数が急激に増加していく結果とな った。これは、非常に同期したバーストを示す培養中期段階から、複雑な発火を示す成熟段 階への移行過程を捉えたものである。 本論文では2章において、まず、神経細胞の基本的機能及び培養神経回路網の同期バース ト現象について現在明らかになっている点を述べる。3章においては微小電極アレイ基板か ら得られる多細胞の活動電位が混在した電位波形から、一神経細胞内での信号伝播経路を推 定し、神経細胞の発火特性を精度良く求める手法について述べる。さらに、4章においては 培養神経回路網活動の長期記録システムの構築、同期バースト現象の解析手法の提案を行い、 得られた結果と考察を示す。最後に5章においては今後の課題を含めて結論を述べる。
第2章
培養神経回路網
2.1 神経回路網特性の研究
2.1.1
神経細胞の基本的性質
ここでは脳を構成する基本的素子である神経細胞の基本的な性質を述べる。神経細胞は介 在している場所によってその形が少しずつ異なっているが、一般的に細胞核を中心部に持っ ている細胞体と呼ばれる部分と、それからでる多数の木の枝のような樹状突起、さらに細胞 体から1 本だけ長く伸びた軸索と呼ばれるケーブルから構成される。ニューラルネットワー クは神経細胞同士が樹状突起と軸索とのシナプスを介する結合により成り立ち情報処理を行 っている。基本的な信号の流れ次のとおりである。多数の神経細胞からの入力を樹状突起に よって受け、その信号を細胞体で加算し、その電位がある一定の閾値を越えたとき、出力信 号である活動電位を発生する。活動電位は軸索を通り末端のシナプスへと伝わる。シナプス において電位変化が生じることによって、神経伝達物質が放出される。放出された物質は樹 状突起側に存在しているレセプター蛋白と結合することによって、イオンチャンネルを開閉 させ、イオンチャンネルを介して物質の流入・流出が起こり、シナプス後細胞の膜電位に変 化が生じる。 このようにして、多くの分岐した樹状突起に同時に多数のシナプスからの信号が入り、こ れが突起中を伝わる過程で積算され神経細胞の膜電位となる。このように多数の入力信号の 影響を加算する作用を空間的加算と呼ぶ。又スパイクの到来によって膜電位の変化した部分 へその変化が減衰する前に次のスパイクが到来すれば電位の変化はまだ残っている膜電位と の変化に加えられる。この作用を空間的加算と呼ぶ。このように空間的、時系列的に多くの 他の神経細胞の信号を加算し、その結果を他の神経細胞へ伝搬することで処理を行っている。2.1.1.1
活動電位発生メカニズム
神経細胞の活動電位の実態は主にナトリウムイオンとカリウムイオンの不均一分布に基づ く細胞内外の電位さが瞬時に反転することによって生じる脱分極である。通常、神経細胞は ATP(アデノシン3リン酸)をエネルギー源として、細胞膜外では Na イオン濃度が高く、 細胞膜内ではK イオン濃度が高い状態を維持している。よって細胞膜内外において電位差が 生じ、細胞内では細胞外に対して約-60mV が平衡状態となっている。 軸索膜にはNaイオンのみを通す Naチャンネルと K イオンのみを通す K チャンネルが存 在し、これらのイオンチャンネルは電位依存性のチャンネルである。膜電位がある一定の閾 値を超えると最初にNa チャンネルが開ことによって、細胞膜内に Na イオンが流入し、膜 電位が約+40mV まで急激に上昇する。さらに少し遅れて K チャンネルが開くことで細胞内 中のK イオンが流出し、急激に膜電位は下降する。この間、約 1msec が活動電位であり、 この発生機構を記述した数学モデルに Hodgikin-Huxley の式がある[10]。さらに膜電位変化 は近隣の Na チャンネルを開閉させることによって、軸索内の部位が次々と脱分極すること で伝搬してゆく。一度、活動電位を発生した場所は約1msec 秒の興奮できない不応期となる こ と で 、 信 号 が 軸 索 内 を 逆 に 伝 搬 す る こ と は 無 い 。 こ の 軸 索 を 伝 わ る 電 位 も Hodgikin-Huxley の式から導き出せる[11]。 信号が軸索を伝わる速さはミエリン鞘と呼ばれる軸索を保護する鞘で覆われている部分で は数百 m/sec、大脳皮質のように覆われていない部分でも数m/sec と高速であるが、シナプ スでは化学物質による信号の伝搬を行うため、この間1msec 程度の遅れが生じ、情報処理に かかる時間のかなりの部分がこのシナプスで費やされている。2.1.1.2
シナプス後電位
神経伝達物質が樹状突起側のレセプターに連結することにより、イオンチャンネルが開き シナプス後細胞の膜電位が変化することは前に述べた。膜電位が上昇させる働きを持つもの を興奮性、下降させる働きを持つものを抑制性と呼ぶ。大脳皮質系における興奮性の神経伝 達物質はグルタミン酸が主である。グルタミン酸が放出された場合、グルタミンレセプター は陽イオンを通過させるチャンネルの一部分を構成しおり、グルタミン酸が結合することに よって陽イオンが流入する。その結果、膜電位がプラス側に傾き、これを興奮性シナプス後 電位(EPSP:Excitatory Post-Synaptic Potential)と呼ぶ。また、抑制性の神経伝達物質 のGABA(γ-aminobutyric acid)が放出され、塩素イオンのチャンネルを構成する GABA レセプターと連結すると、塩素イオンが流入することで、膜電位がマイナス側に傾く。これ を抑制性シナプス後電位(IPSP:Inhibitory Post-Synaptic Potential)と呼ぶ。この EPSP とIPSP の総和が閾値を超えると神経細胞は活動電位を発生させる。大脳皮質系での興奮性レセプターであるグルタミン酸レセプターは大きく分割すると non-NMDA (N-methyl-D-aspartate) 型と NMDA 型に分かれる。non-NMDA 型受容体は、 さらにキスカル酸受容体(AMPA 受容体)とカイニン酸受容体にわけられ、これらは、Na+、
シナプス前の神経細胞が高度に発火すると、まず、non-NMDA 型受容体が活性化されナト リウムイオンチャンネルを作動させることで大きな EPSP を引き起こす。これにより、 NMDA 型受容体をブロックしている Mg2+が除去され、ナトリウムイオン、カリウムイオン と共にカルシウムイオンの流入が引き起こされ、non-NMDA 型に比べ小さいが長く持続す るEPSP を発生させる。さらに、このカルシウム流入が細胞内セカンドメッセンジャーを活 性化させ、長期増強を引き起こすと考えられている。 胚令初期には細胞表面には両受容体が存在するが、NMDA 型受容体のみのシナプス部位 が多数存在し、発生過程においてnon-NMDA 受容体を伴った EPSC の大きなシナプスが出 現するという報告[12]がある。また、NMDA 型受容体は NR1 サブユニットと 4 種類サブユニ ット(NR2A-NR2D)から形成されており、これらサブユニットの発現状態が胎児段階と成熟 段階で異なる報告もある[13]。また、GABA 作動型のレセプターにおいても、発達段階に大き な影響を与えているとの報告がある[14]。 これら各種レセプターの発現状態が発生過程で生じる高度に同期したバーストを引き起こ す源となっていると考えられる。
2.1.1.3
神経細胞への薬理作用
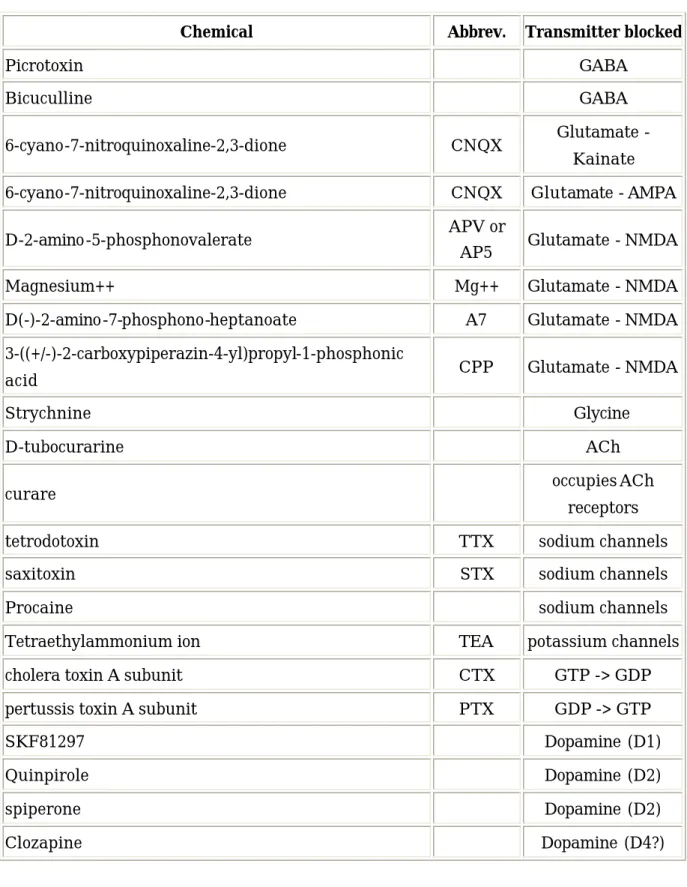
伝達物質のレセプターに直接反応して作用を与える薬理作用を持つものには大きくアゴニ ストとアンタゴニストに分けることができる。アゴニストは伝達物質のレセプターに結合し て、伝達物質と質的に同じ反応を受容細胞に引き起こすものであり、アンタゴニストは伝達 物質のレセプターに結合し、その働きを阻害するものである。神経細胞の発火を観測しなが ら、これらの薬を用いることにより、その神経細胞へのシナプス結合がどのような種類のも のであるのか特定することができる。神経生理の実験に良く用いられるアンタゴニストを表 2-1 に載せる。また、GABA のアゴニストとして muscimol がある。表 2-1 神経伝達物質のアンタゴニスト
Chemical Abbrev. Transmitter blocked
Picrotoxin GABA
Bicuculline GABA
6-cyano-7-nitroquinoxaline-2,3-dione CNQX Glutamate - Kainate 6-cyano-7-nitroquinoxaline-2,3-dione CNQX Glutamate - AMPA D-2-amino-5-phosphonovalerate APV or
AP5 Glutamate - NMDA Magnesium++ Mg++ Glutamate - NMDA D(-)-2-amino-7-phosphono-heptanoate A7 Glutamate - NMDA 3-((+/-)-2-carboxypiperazin-4-yl)propyl-1-phosphonic
acid CPP Glutamate - NMDA
Strychnine Glycine
D-tubocurarine ACh
curare occupies ACh
receptors
tetrodotoxin TTX sodium channels
saxitoxin STX sodium channels
Procaine sodium channels
Tetraethylammonium ion TEA potassium channels cholera toxin A subunit CTX GTP -> GDP pertussis toxin A subunit PTX GDP -> GTP
SKF81297 Dopamine (D1)
Quinpirole Dopamine (D2)
spiperone Dopamine (D2)
2.1.2
同期バースト現象の発生メカニズム
発達段階の神経回路網では、全体的なバースト発火を繰り返す現象が見られる。さらに、 このバースト発火は回路網全体において同期性が高く、周期的に発生する。つまり、回路網 の活動は全体が同期して発火する活動フェーズと殆ど発火が見られない静止フェーズに分か れ、この2つのフェーズが交互に生じる現象を同期バースト現象と呼ぶ。
この現象は発達段階のラットのLGN(lateral ganiculate nucleus)において生じていて、 その形成に重要な役割を担っていることが1991 年に M. Meister らによって報告された[4]。 さらに、哺乳類の海馬、大脳皮質等の脳組織において生じていることも報告されている。周 期的に発生する非常に同期バーストはラットや猫、猿等においてのin vivo 実験[15]、in vitro でのスライス[16]、培養神経回路網においての両方で確認されている。これらの報告により、 同期バースト現象が自己組織化的な中枢神経系における記憶、学習、認識といった基本的な 脳の情報処理機能の獲得において重要な役割を担っていると考えられているが、このメカニ ズムについては未知な部分が多く残されている。 一般的に海馬、大脳皮質の組織において、同期バースト現象は細胞内のカルシウム濃度変 動と密接な関係があることが多数報告されている。特に大脳皮質の2次元培養神経回路網に おいてはNMDA型レセプターをブロックしている Mg2+を除外した低マグネシウム環境下で 非常に周期的な同期バースト現象を引き起こすことが分かっている。さらにCa2+濃度と同時 に活動電位を測定することで同期バースト現象は緩やかなカルシウム変動と同期して周期的 に発生していることがH. P. C. Robinson らにより報告された [17]。これら回路網全体に伝わ る神経回路網のバースト及びカルシウム変動は NMDA レセプターのアンタゴニストである APV を投与することにより阻害されることから、同期バースト現象の伝播はギャップジャン クションや物質の拡散を介したものではなく、多シナプス結合を介して回路網全体に広がる と考えられている[18]。 バーストの発生源はバーストごとにランダムであり、回路網を紫外線レーザーにより切断 することで分割された部位は別々の周期でバースト現象を繰り返すようになる。また、不応 期時に電極アレイからの刺激により引き起こされたバーストは部分的に伝搬し、1秒から30 秒間隔で周期的に刺激を与える実験では刺激間隔が小さいと伝搬速度が遅く、バーストに参 加するスパイク数も減少することから、同期バースと現象はシナプス前細胞の自発発火のレ ベル、ネットワーク結合、興奮と回復度合いに同期バーストは支配されていると考えられる [19]。神経細胞のランダムな発火にこれらのパラメータをモデリングし、シミュレーションを 行った試みでは、周期的な同期バースト現象に近いパターンが得られた[20]。しかしながら、 細胞内カルシウム変動を考慮したモデリングには至っていない。
2.2
培養神経回路網の発達過程
成長過程による同期バーストを含めた特性変化の研究報告は 1996 年において微小電極基 板上の培養神経回路網を用いたH. Kamioka によって成された[7]。これはラット17 日目の胎 児から抽出された大脳皮質神経細胞の培養における変化を調査したものである。培養初期段 階では回路網はランダムな自発発火のみとなり、他への信号伝播は見られないが、培養 4-5 日において0.5-1mm/s の遅い信号の伝播が見られ、バースト間隔は 50-100sec であった。2 週目からは多くのニューロンが参加した非常に同期したバーストが見られ、伝播速度も 1-10cm/s に増加し、バースト間隔は 40sec 程度に減少している。さらに培養 30 日目以降で は複雑なパターンを示し、それ以上の培養日数においても同様だった。この例では複雑なバ ーストパターンとして長く持続するタイプ(450msec)と短いタイプ(100msec)が同時に観測 されている。培養初期においては細胞膜中にイオンチャンネル及びレセプターは形成されて いるが、シナプス結合が殆ど無いために他に伝播しない。しかし、その後、成長に従いシナ プス結合を形成していくが、シナプスが未熟で結合も少数であるために神経伝達物質の放 出・吸収が遅く、長く持続するバーストとなり、シナプス後電位が加算されるまで多くの時 間を必要とするので伝播速度は小さい。さらにシナプス小胞の数も少なく、一度のバースト によって費やされてしまい、神経伝達物質の充填のために長い休止期間が必要となる。これ らのバーストは低マグネシウム濃度下で起こる同期バーストと同じくAPV, TTX, muscimol に抑制され、NMDA 型を介したカルシウム変動を伴い、これがシナプス結合発達のトリガ となっていることが示唆された。 また、培養30 日目以降の複雑なパターンに APV を投与したところ、NMDA 型の遅いバ ースト部分のみが阻害されることから、非NMDA 受容体が成長していることが示唆された。 さらに、TTX を各成長段階において適応すると、非常に同期したバーストは抑制されずに 成熟段階のような長く持続するバーストと類似し、さらに CNQX を投与しても阻害されな かったが、APV を投与すると消失している。これは non-NMDA 型レセプターの発達が同期 した発火に依存していることを示唆している。さらに回路網成長期には抑制性のシナプス結 合は興奮性のシナプス結合よりも後に生じているという報告がある。これを考慮すると回路 網初期では発火を助長させられ、Hebb の学習規則に則ったシナプス形成が抑制性も含めて 進行するのではないかという仮説が立てられる。 以上の理由から、同期バースト現象が回路網形成に重要な役割を担っていると考えられて いる。しかし、これらの結果は各培養日数の異なる回路網による統計的な解釈である。同一 神経回路網を連続的にモニタリングできれば、以上より、重要なパラメータとなっている、 バースト持続時間、バーストに参加するスパイク数、バーストの伝播速度、バースト間隔の 微細な変化を捉えることができる。これにより、同期生の高いバースト現象から複雑化して いく過程における現象を解明できると考えられる。2.3
微小電極アレイによる活動電位計測
培養神経回路網を測定対象とした場合、神経細胞の活動電位を測定する手法として、パッ チクランプ法、光学膜電位法、微小電極アレイを用いる手法の大きく3つに分けることがで きる。一般的に神経生理の実験に用いられるのが、ガラス管微小電極によるパッチクランプ 法である。この手法は1つの神経細胞に直接ガラス間を挿入しガラス管内と細胞内を電通さ せて細胞内電位を測定する手法である。この手法は S/N 比が非常に良く、EPSP・IPSP を 含めた一細胞の内部電位を記録するのに非常に適するが、神経回路網活動の空間的な伝播を 捕らえることはできない。また近年よく用いられる光学的膜電位記録法は膜電位の伝播と共 に変化する色素を観測することで高い空間分解能が得られるが低S/N 比である。上記の両手 法は回路網を測定する上でそれぞれ大きなメリットを持っているが、数十日に渡る長期的な 測定としては神経回路網になるべく影響を与えず、非侵襲な測定を行う必要がある。ガラス 管微小電極は細胞膜を傷つけ、光学膜電位記録法は色素自体が細胞にとって毒性を持つため、 これらの手法は長期的な測定に用いることができない。 細胞に影響を与えない記録法として最も適しているのが、1979 年に Guenter W. Gross が 開発した埋め込み微小電極アレイによる細胞外記録法である。この手法はあらかじめガラス 板に微小多電極をフォトレジスト加工によりプリントし、電極上に試料を載せ培養すること で、非侵襲な計測を可能としている。多電極アレイ基板上で神経細胞を培養すると、神経細 胞は神経突起を2 次元平面状に伸ばし、回路網を形成していく。この伸張していく軸索・樹 状突起を含めた細胞膜外の電位を微小電極で多点記録することが可能である。微小電極アレ イによる測定はS/N 比も比較的良く、他電極の配置によってある程度の空間分解能を有する ことによって、神経回路網の伝播経路を考慮した解析が可能となる。 微小電極アレイは神経細胞膜外にあるため、計測される活動電位は細胞膜外に流入・流出 する主にナトリウムとカリウムイオンが引き起こす過渡的な電流が体積導体中に作る微小な 電位となる。この電位は細胞膜内の約 100mV、1msec から大きく減衰し、振幅、約百μV オーダー、帯域として約300Hz∼6kHz を有する信号となる。この得られる活動電位は細胞 と電極の位置関係に依存し、距離が近ければ振幅も大きくなり、距離が遠ければ振幅は小さ くなる。一般的に 10μm離れた位置にある細胞の信号は検出されないとされる。しかしなが ら、細胞外で計測する非侵襲な手法である反面、一電極で記録される活動電位は複数の神経 細胞の活動電位の重ね合わせとなる。とくに培養液環境中での測定は活動電位の重なる頻度 が高い。回路網形成中の同期バーストの解析においては、2.1.2 節で述べたが、神経細胞ごと での発火タイミングは考慮されていない。電極から重なって得られるスパイクを発生源の神 経細胞ごとに精度よく分類できれば、それらの発火パターンの推移、及び各細胞間をシナプ スを介して空間的に伝播する様子が詳細に捉えられ大きな知見を得る可能性がある。 次章では微小電極アレイから得られる複数同時測定を考慮した解析を行うことによって、 個々の神経細胞の発火パターンを精度よく得る手法を述べる。第3章
神経細胞発火パターン検出
回路網を構成する神経細胞の個々の発火特性の成長による変化を捉えるには各スパイクの 発火時間の間隔をヒストグラムにし、それを発火パターンとして解析することができる。し かし、多電極アレイを含めた細胞膜外記録法において、個々の神経細部が発火した時間を求 める場合に問題となるのが、1つ以上の神経細胞の活動電位が折り重なるようにして得られ てしまうことである。それは1電極に1つ以上の細胞が接している場合に起こり、電極の大 きさが大きいほど、多くの神経細胞の電位が加算され得られてしまう。よって、各神経細胞 の発火パターンを得るには、多数の細胞の活動電位が折り重なった一時系列データから、細 胞ごとにスパイクを分類する必要がある。これをスパイクソーティングと呼ぶ。 電極上の神経細胞はそれぞれ電極と細胞との接触度及び距離が異なるため、電極から得ら れるスパイク波形により各細胞をキャラクタライズすることが可能となる。これら活動電位 波形は短時間に変化することはなく、一電極上で得られたスパイク波形が同じなら、それは 同じ細胞が発生したスパイクであると見なせる。 一般的なスパイクソーティングはスパイクの幅とスパイク振幅の2次元クラスタリングを 形成させることにより視覚的に分離する手法が主に使用される。最近ではテンプレートマッ チング[23]やウェーブレット[24][25]を用いた波形の相関を考慮する手法も提案されているが、バ ーストのように極度に波形が重なる場合には波形の相関がとれないために分離は難しい。可 塑性の研究で行われる実験ではMg2+濃度を高くすることで、同期バーストのような自発活動 を抑えているが、培養環境下での測定ではバーストとして活動電位が得られるので、波形同 士が重なる場合が多い。 しかしながら、電極アレイ基板上に伸張した軸索を含め、神経細胞内を伝播する信号を多 電極で捉えることにより、その信号伝播経路を推定でき、さらにその細胞の発火時間を精度 よく求めることが可能である。 この章では、まず、従来用いられるスパイク幅と振幅を用いた2次元クラスタリングによ るスパイクソーティングの手法を示す。次に多電極アレイから得られる同一神経細胞のスパ イクを検出し、多電極に渡る信号の伝播経路推定を行うことで、従来手法では分離不可能で あったスパイクの分類が可能となることを示す。さらに、50DIV 以降の成熟期における神経 細胞の発火パターンの推移を示し、結果を述べる。3.1 活動電位データ
活動電位データは予め回路網を50 日間培養後、32 日間に渡り合計 13 回計測したものを 用いた。電極基盤は図 3-1 に示すように1ブロック構成であり、電極サイズは 30μm 四方、 電極中心間隔は180μm である。また、各電極に対応する電極番号を図 3-2 に示した。培養 液交換は図 3-3 に示すようにシリンジにて古い培養液を抜き、新しい培養液を注入すること で遠隔的に行い、毎週2回、0.5ml ずつ手動で行った。 微小電極アレイで得られた微小な活動電位はアンプにより2万倍に増幅され、サンプリン グ周波数 25kHz、16bit 量子化で計測される。計測時間は 1 フレームあたり 16384 サンプ リングポイントで240 回、合計 157 秒であり、64 電極同時に記録される。図 3-4 に 51DIV において、64 電極の中でよくスパイクを得られている電極の波形を載せる。図 3-4-A は 150 秒間の各電極で得られた活動電位波形を示し、全体に活動が高まるバースト状態と全体が静 まる状態が繰り返し起こっていることが分かる。図 3-4-B は活動が高まった状態の一部分を 150msec 窓で載せる。電極アレイを用いる細胞外記録法では細胞内電位と対照的にマイナス 側に鋭いピークとなってスパイクが計測されている様子が分かる。振幅の異なる波形が混在 している。また、B1ch では波形が重なる場合も見受けられる。前にも述べたが、このよう に振幅の異なるスパイクは別々の神経細胞からの信号であるとでき、これを分離することに よって、神経細胞レベルでの解析が可能となる。図 3-1 微小電極アレイ基盤(1 ブロック構成)
図 3-2 電極番号(1 ブロック構成) 500μm
図 3-3 培養液交換システム
64 microelectrode array
incubator
図 3-4 活動電位波形 -100μV 100μV
51 DIV
-100μV 100μV 150msec 150secA
B
B1ch D0ch E0ch F0ch G0ch H0ch B1ch D0ch E0ch F0ch G0ch H0ch3.2 スパイク振幅・幅を用いたスパイクソーティング
一般に網膜の視細胞の応答のスパイク分離に適応されている手法として、クラスタリングよるスパ イクソーティングがある。2次元平面状にスパイクの振幅を縦軸、スパイクの幅を横軸としてプロットし ていくと、いくつかのクラスタが形成される。それを視覚的にある閾値を決めることにより分類すること ができる。分類されたクラスタは特徴的なスパイクの形を代表するものであり、クラスタ数がその電極 上の検出された細胞数だと推定することができる。今回、次のように振幅と幅を求めることにした。便 宜上スパイク波形のそれぞれのパラメータに名称をつけた。図 3-5 にスパイクのパラメータの名称の 図解を載せる。 図 3-5 波形のパラメータの名称3.2.1
振幅(Amp)の導出
Base-line(基線)からマイナス側のピークであるValley-pointまでの差の絶対値を振幅(Amp)と定め た。つまり、測定値
x
n(
n
=
1
,
2
,...,
n
)
において、threshold
x
n<
(3-1)となる
x
nにおいて最小のx
nをValley-point とし、Valley-point の時間から±Spike-size/2 分(サンプ リングポイント数)をスパイク波形として抽出するようにした。図 3-6 に示すようにうねりがある場合、同 じスパイクでもValley-point が違ってくる。よって、ノイズによる影響を小さくし、スパイクの形に重点を おくと、スパイク自体の平均値(Base-line)からValley point までの変異を振幅値としたほうが妥当な 振幅値(Amp)を得ることができる。以下のように振幅値を求めた。∑
=− +=
Spike size size Spike nX
Valley Po nsize
Spike
Baseline
2 _ 1 _ 2 1 _ int_
1
(3-2) | |X _ int Baseline Amp= Valley Po − [V] (3-3)スパイクは約 1msec の波形であるので Spike-size は約 20 から40 ポイント(25kHz で 0.8msec∼ 1.6msec)が妥当である。 図 3-6 うねりによる同スパイクの違い Spike-size Amp Base-line
3.2.2
幅(width)の導出
スパイクと閾値が交差するところのスパイク間隔を幅(width)と定めた。電圧の急激な降下から電 圧の急激な上昇までのスパイク波形は約は 1msec 程度である。25kHz サンプリングにおいては、 0.04μsec の分解能となり、クラスタリングには不十分である。よって、サンプリング周期よりも更に正 確に求めるために幅の算出は、図 3-7 のように直線近似を施し、サンプリングポイント間を補間する ことで行う。つまり、サンプリング周波数をf[Hz]とすると、スパイクとなる最初の時間tiは以下のように 表される。]
[
/
1
1 2t
f
Hz
t
−
=
(3-4)|
)
(
|
threshold
V
t
1a
=
−
,b
=
|
V
(
t
2)
−
threshold
|
(3-5) b a b ) ( 2 1 2 + × − − =t t t ti (3-6) 最終の時間t
oは以下のように表される。]
[
/
1
3 4t
f
Hz
t
−
=
(3-7)c
=
|
threshold
−
V
(
t
4)
|
,d
=
|
V
(
t
3)
−
threshold
|
(3-8) d c d ) ( 4 3 3 − − × + =t t t to (3-9) よって幅(width)は以下のように表される。 i ot
t
width
=
−
[s] (3-10) 図 3-7 スパイク幅の導出3.2.3
スパイク幅と振幅による2次元クラスタリング
上記のように求めた幅(width)を横軸、振幅(Amp)を縦軸とし 2 次元平面状にプロットすると、図 3-8 のようになる。この図において、この場合、クラスタA とクラスタB に分離されていると視覚的に評 価できる。つまり、細胞 A と細胞 B が計測電極に接していることが推測できることになる。 しかしながら、スパイク幅と振幅は波形の特徴の一部であるため、異なる細胞のスパイクでも 2 次 元平面上このようにプロットすると接近しており、同じクラスタのように見えてしまう場合が生じる。特に 実データを当てはめて行ってみると、視覚的に分離できない、つまり、プロットがまばらでクラスタを 形成しないことも多く見受けられ、この場合にはスパイクの分離は不可能である。 図 3-8 クラスタリングによる波形の分離の例3.3 細胞内信号伝播経路推定による発火パターン検出法
多電極アレイを用いた計測では、多電極に渡り同じ神経細胞の活動電位を計測できる場合 がある。解析の流れは、最初に一電極における時系列データに対しスパイクの振幅相関と波 形相関の両方を用いた相互マッチングによるスパイクソーティングを行う。次ぎに分離され た各スパイク系列の発火タイミングにおけるマッチングを行うことで、多電極に渡る同一神 経細胞の検出と伝播経路を推定する。さらに、多電極における伝搬遅延を考慮することで個々 の細胞ごとに発火時間を高精度で得ることができる。 本節ではこれら解析手法の提案を行い、長期的観測データに適応することにより、個々の 細胞の発火パターンが長期的にどのように変化するか解析を試みた。 この信号処理の手順について示すと図 3-9 のようになる。以下、各々について概説する。 図 3-9 多電極を用いたスパイクソーティングの流れスパイクの検出と切り出し
スパイクソーティング
発火パターンマッチングによる伝搬経路推定
遅延パターンによる神経細胞のスパイク検出
3.3.1
スパイクの検出と切り出し
活動電位波形の振幅を閾値処理することにより、スパイク部分を切り出す。閾値ηをマイ ナス側に超えた部分で振幅が最大の時刻tfを発火時刻とする。このtfを中心に一定幅 2τ でスパイクの切り出しを行う。切り出されたスパイク波形はマイナスピークを t=0 として、 S(t)(-τ ≦t ≦ τ)のように表す。今回は τ=0.4msec とした。3.3.2
相互マッチングによるスパイクソーティング
各電極で得られたスパイク群に対して、同一の細胞からのもの同士にグループ分けする。 従来、スパイクソーティングはスパイクの幅と振幅の 2 次元拡散図を元に行うものが一般的 であるが[1]、ここでは、細かな波形の特徴まで考慮することで、より厳密なスパイクソーテ ィングを行う。即ち、スパイク Sa(t)とスパイク Sb(t)について、波形の相関係数 Cshape(a,b) と、振幅の相関値 Camp(a,b)を次式で求める。∑
∑
∑
+ − = + − = + − = = τ τ τ τ τ τ i b i a i a b shape i S i S i S i S b a C ) ( ) ( ) ( ) ( ) , ( 2 2 (3-11){
}
{
(0), (0)}
max ) 0 ( , ) 0 ( min ) , ( b a b a amp S S S S b a C = (3-12) これらが各閾値 ηshape、ηampを超えた場合、この2つのスパイク Sa,Sbはマッチングし同 じ細胞からのものと判断する。これを全てのスパイクに対して総当りに行う。しかし、Sa、Sb、 Scにおいて、Saと Sb、また Sbと Scがマッチングしているが、Saと Scはマッチングしていない ということが起こりえる。本研究ではこのとき、Saと Scは同じ細胞であると識別することに した。つまり似たスパイク同士は閾値により決められた範囲で手を繋ぐようにしてグループ 化されることになる。これは連続的な発火の振幅の減衰に対するスパイク波形の微小な連続 的変化を考慮するものである。 今回は高速化のため、最初に得られる 500 のスパイクに対し相互マッチングを行い、スパ イクグループを得た後、新しく検出されるスパイク波形 Snに対して各グループの平均スパイ ク波形 Savrとの距離d(n,avr)を以下の式で算出する。∑
=− − = τi τ Sa i Sb i b a d( , ) ( () ())2 (3-13)この距離が最小になるグループに加えることとした。グループに加える際にはグループの スパイクにおける距離の標準偏差の2倍以内に収まることを条件としている。さらに全ての スパイクを一旦分離した後、各グループの平均波形においてもう一度相互マッチングさせ、 最後にグループ同士を連結しスパイクソーティングを終了する。これを全 64 電極において行 った。グループ化されたスパイク群を系列と呼ぶことにする。
3.3.3
発火パターンマッチングによる伝搬経路推定
各系列は1つの細胞から出されたスパイクであると仮定できる。電極間で信号が伝搬する 場合、ある遅延を持って伝搬するので同じ発火パターンが各電極で得られることになる。よ って全 64 電極で得られた系列で発火パターンの一致するものを検出することで、電極間にお い て 伝 搬 経 路 を 推 定 する 。 系列 a か ら 発 生 した numa 個 の ス パ イ ク の発 火 時 間 をte,1 , te,2 , … , te,numaとする。発火遅延tlateを-4msec から+4msec まで変異させた時のパター
ン a とbの相関係数 Ctime(a,b,tlate)は以下の式で算出し、閾値η3を超えたとき、パターン a,
b はマッチングしているとし、Ctime(a,b,tlate)が最大になるときのtlateをパターン a, b の相対
的な発火遅延とした。 b a num i num j b late i a late num num t t t g t b a C a b + − = 2
∑ ∑
=1 j=1 ( , , , ) ) , , ( ⇒ ⇒ < − = 0 1 | | ) , ( 1 2 2 1 else t t t t g τ (3-14) 今回、求められた相対的な系列の遅延時間は最大のものでも約 0.56msec である(図 3-13)。 神経細胞同士のシナプスを介する伝搬は 1msec 以上の遅延が生じるので、これら同系列の伝 搬はシナプスを介するものでなく、同細胞の軸索が多電極において接して得られる信号伝搬 である。3.3.4
神経細胞の発火パターン検出と長期にわたる観測
今回の計測したサンプルは 50DIV のもので、軸索が十分成長した後とすると、多電極にお ける伝搬遅延はでは一定であると仮定できる。このため、推定される多電極に渡る同じ神経 細胞の活動電位の伝搬遅延を考慮することで、その細胞の発火時間を精度良く求めることが できる。さらに発火間隔分布をヒストグラム化することで発火パターンが一ヶ月に渡りどの ように変化したのかを可視化した。この手法についてここで述べる。 推定された細胞のスパイクが N 個の電極にそれぞれ遅延時間 N late late late1, 2,L, で伝搬した とする。これを遅延パターンとする。これら N 個の電極で得られている全スパイクの発火時 間を te,1 , te,2 , … , te,nume(e は電極番号1∼N,numeは全スパイク数)とすると、時間t における遅延パターンと N 電極での全発火パターンとの相関は N late t t g t f N e num i ei e e∑ ∑
= = − = 1 1 (, , ) ) ( (3-15) で算出することができる。閾値 η4を設定しこれを超えた時間をこの細胞の発火時間とする ことで多電極への軸索を考慮した精度良いスパイク検出が可能となる。3.4 解析結果
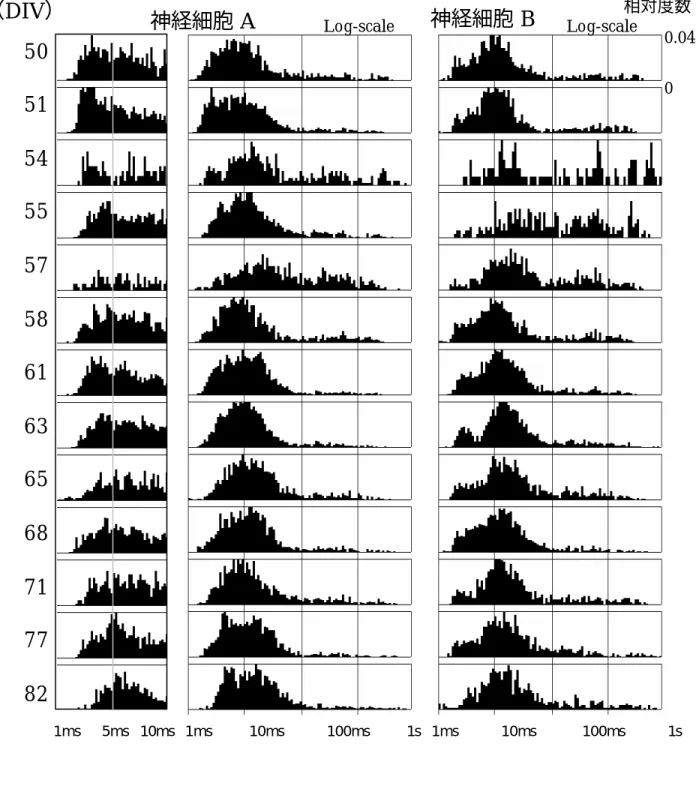
実験データを 3.2 に示した手法により、スパイク振幅・幅を用い2次元クラスタを形成さ せたものを図 3-10に示す。0μV付近で検出されているものはノイズである。C3電極や H2、 H3 電極等ではクラスタが形成され、視覚的に分離することが可能であることが分るが、そ の他の電極では分離が難しい。特にE0 電極においては視覚的には1つのクラスタに見える。 しかしながら、今回提案した手法を用いることによって2つのクラスタの集合に分割するこ とが可能であった。図 3-11 に今回提案した手法により分類された例として E0 電極の結果を 示す。一クラスタとして1つの神経細胞としてその発火間隔分布を求めた場合、1msec 以下 にも分布が得られている。神経細胞には不応期があるため、1msec 以下の高頻度な間隔で発 火することは無く、この分布は2 つ以上の神経細胞からのものであると推定され、2 次元ク ラスタを用いた分離では対応できないことが示される。しかしながら、今回提案した手法で は、E0 電極では神経細胞 A と神経細胞 B の2つに分離することが可能であり、それぞれの 分布が高精度で得られている。 神経細胞A は D0,E0,E1,F1,G0,H0 電極での伝播遅延を用いてのマッチングにより検出さ れたもので、神経細胞B は B0,B1,D0,E0,E1 電極を用いて検出されたものである。それぞれ の活動電位は図 3-12 に示すようにある一定の遅延をもって各電極で検出されているのが分 かり、それを用いることで、細胞A と細胞 B に分離することができた。特に B1ch の細胞 B のように1電極における活動電位が歪んだ場合においても他の電極でマッチングするので、 検出することができる。 さらに、信号伝播経路推定により2電極以上で計測された神経細胞の数は今回の回路網に おいて神経細胞A と神経細胞 B を含め4個であった。これを最初に発火が検出された電極を 基準に伝搬の様子と遅延時間を電極上に表したものを図 3-13 に載せる。一般的に電極間隔 が大きいほどやはり遅延が大きくなり、電極位置を考慮して伝播速度を求めると、約数m/ 秒で神経突起中を伝播していることが分かる。神経細胞 A、B のみに注目し、50DIV から 80DIV までの発火間隔分布の推移を図 3-14 に示す。特に神経細胞A では 1msec から 10msec までの分布を標準スケールで示している。 神経細胞A,B とも、50DIV からの推移ということもあり、成長による大きな変化は見られ ない。しかしながら、54DIV, 57DIV では分布が大きく異なって得られている。これは培養 液交換における影響が顕著にでた結果である。しかしながら、神経細胞A においては 50DIV
では発火間隔が2.5msec 付近が最も頻度が高いが 61DIV では 2.7msec付近、82DIV では 5.6msec
付近と、日数を経るごとに発火間隔のピークが右に遷移している様子が観察され、発火間隔が大 きくなってくるという傾向が結果として得られた。これは回路網として減衰過程にあると考えら れるが、詳細なメカニズムは明らかではない。
図 3-10 幅と振幅によるクラスタリング結果
0.8 [msec]
80 [μV]
図 3-11 スパイクソーティング結果 0 0.4 [msec] 0 60[μV] 神経細胞A 神経細胞B スパイク幅 スパイク幅 振幅 振幅 60[μV] 0.4 [msec] 発火間隔分布(全スパイク) 20[msec] 0 0.1 相対度数 0.1 0 20[msec] E0 ch E0 ch 51 DIV
図 3-12 分離された活動電位波形 図 3-13 推定された神経細胞の信号伝搬の様子 神経細胞A 神経細胞B 神経細胞C 神経細胞D 0.12 0.24 0.40 0.28 0.44 0.36 0.20 0.12 0.40 0.28 0.56 0.04 0.08 0.24 (数字は遅延時間を表す[msec]) 25msec -100μV 100μV B0ch B1ch D0ch E0ch E1ch F1ch G0ch H0ch 細胞A 細胞A 細胞A 細胞B 細胞B
図 3-14 発火間隔分布の推移
50
51
54
55
57
58
61
63
65
68
71
77
82
(
DIV)
1ms 10ms 100ms 1s 0.04 0神経細胞
A
神経細胞
B
1ms 5ms 10ms 1ms 10ms 100ms 1s Log-scale Log-scale 相対度数3.5 まとめ
神経回路網を培養した状態では回路網は自発バーストを繰り返し、個々の細胞の活動電位 が重なり合うことが多く、従来のスパイクソーティングでは対応できなかった。しかし、本 研究で提案した電極間で伝搬経路を推定し、その伝搬遅延を考慮してスパイクを再検出する 手法により、個々の神経細胞の発火パターンを精度良く検出することが可能となった。 注射器による培養液交換システムにより得られた50DIV から 82DIV のデータに本手法を 適用し、各神経細胞の発火間隔分布を抽出した。その成長による推移に大きな変化は見られ なかったが、しかしながら、その分布は大きく揺らぐ場合があり、詳細な神経細胞の発火パ ターンの推移を詳細に捉えることはできていない。この段階での回路網は成熟期に相当する ため、安定に推移すると考えられ、これら分布の揺らぎは手動による培養液交換により急激 に培養液環境が変化するために起こるものだと推測される。 第4 章では、シリンジポンプによる微量な培養液の潅流システムによる培養初期の成長段 階からの安定な活動電位測定について述べる。第4章
回路網活動長期モニタリング
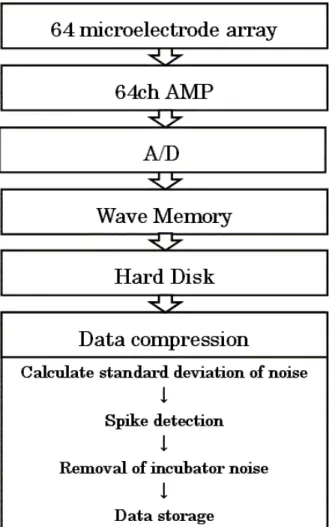
微小電極アレイが1979 年に開発されてから、in vitro での多くの実験に使用されてきた。 しかしながら、その使用方法の多くは対象試料に対して非侵襲であるという最大の長所を生 かしたものではなく、パッチクランプ法やカルシウム濃度モニタリング等と複合しての使用 される場合である。また可塑性の研究においては成熟段階の回路網を使用するために電極ア レイ上の対象試料を実験に合致した日数だけインキュベータ内にて培養して使用するといっ た使用法であり、培養初期段階から活動電位を連続的に得た報告はない。 この最大の理由としては微小電極アレイがいかに非侵襲であっても、アンプやAD 変換器 といった装置を接続した電位記録と同時に神経細胞の形成が順調に進むような培養環境の維 持が困難であったためだと考えられる。特に培養液環境においては浸透圧、pH、温度が回路 網に大きな影響を与え、さらに滅菌を保った状態で数十日に渡る測定は困難である。 本研究ではこの問題を打開するため、培養環境の維持においては、一般に用いられている CO2インキュベータを用いることにした。これにより、回路網は他のサンプルと同様の雰囲 気下で培養される。さらにシリンジポンプによって非常に緩やかに培養液が潅流するシステ ムの構築を行うことで安定な培養液環境を作り出した。インキュベータ内の微小電極アレイ から連続的に得られる膨大な電位データは圧縮して記録されるように設計し、5DIV より計 40 日間の活動電位測定を行った。 さらに解析手法に関しては発達段階で生じる同期バーストを検出し、2.2 節で述べたよう に同期バーストの重要なパラメータである、バースト持続時間、バーストに参加するスパイ ク数、バースト間隔を定量化すると共に、バーストが伝播した空間パターンを用いることで バーストのクラスタリングを行う手法を提案する。 以上により同期バーストをキャラクタライズし、その各パラメータが成長によってどのよ うに推移していくかを調査した。その結果、得られた知見、考察を含め、回路網の長期的モ ニタリングシステムについて本章で述べる。4.1
実験
4.1.1
試料作製
18 日胚の Wistar ラットから大脳皮質を摘出し 0.02%のパパイン酵素(Boehringer)を加え 36℃の湯漕に約 20 分間入れた後、数回のピペッティングによって神経細胞を単離した。あ らかじめポリD-リジン、ラミニンでコーティングした埋め込み 64 電極アレイ上で 5 日間培 養させた。4.1.2
培養液構成
使用した培養液はDMEM(Dulbecco’s Modified Eagle’s Medium: Gibico)に熱処理によ って不活性化させた 5%牛胎児血清(Gibco)、5%非働化ウマ血清、2.5μg/ml のインシュリ ン(Sigma)にペニシリン/ストレプトマイシンを 5?40U/ml を加えたものである。これを、グ リア細胞のみをあらかじめ培養したシャーレ内に1 昼夜保持し、conditioned medium とし たものを潅流液とした。
4.1.3
多電極アレイ基板
基板に石英ガラスを用い透明導電性材料のITO(Indium-tin-oxide)で電極パターンを作 り、表面をフォトレジストで被覆したものを用いた。測定点の電極の先端のみ絶縁膜を取り 除き、界面インピーダンスを下げるために白金をコーティングした。この電極基板上にガラ ス枠を取り付け、培養液を入れるようにし電極付シャーレとした。これは測定基板への結合 のためのコネクタ電極がついている(図 4-2)。 電極アレイ基板は8×4 電極を 1 ブロックとする間隔 500μm の 2 ブロック構成、電極サ イズは30μm 四方、電極中心間隔は 180μm のものを用いている(図 4-1)。また、電極番 号を図 4-3 に示すように取り扱う。図 4-1 微小電極アレイ基板(2 ブロック構成) 図 4-2 電極アレイ付シャーレ 図 4-3 電極番号(2 ブロック構成)
500μm
180μm
4.1.4
培養液潅流システム
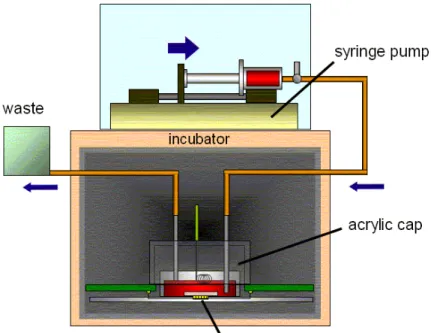
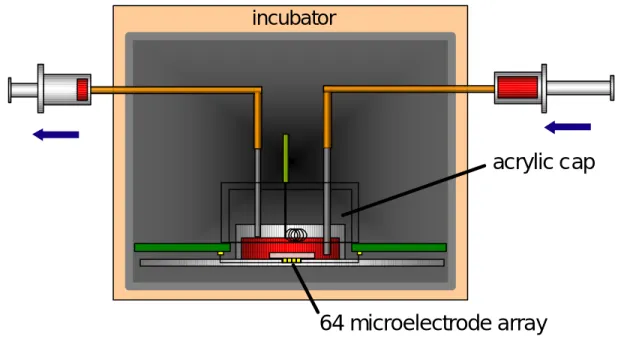
in vitro での計測は外部溶液の物質濃度環境を実験に適合するようにコントロールするこ とが容易である反面、その環境を長期的に維持することはこれまで困難であった。特に回路 網の発火パターンが外部溶液に対して敏感に反応するため、安定に培養中の回路網から活動 電位計測を行うためには外部溶液である培養液を回路網に対し無襲侵に交換することが必須 となる。 本研究においてはシリンジポンプを用い、非常に緩やかな流量で培養液を潅流させるシス テムを製作した。そのシステムの写真および模式図を図 4-4 図 4-5 に示す。微小電極付ガラ ス板を固定・結合する基板部分を別に製作し、インキュベータ内に直接設置した。また、培 養液注入・排出用の管が付いたアクリルキャップを作製し電極付きシャーレに被せた。これ によりインキュベータ内での培養液の蒸発を防ぐことができ、キャップの内径はシャーレの 外径よりも僅かに隙間があることで、CO2が基板の下側から通気するようになっている。さ らにキャップには64 微小電極における参照電極も取り付けている。 注入用管は培養液の入ったテルモシリンジ(10ml)にチューブで接続し、シリンジポンプに よって、流量0.1ml/h にて培養液を注入した。培養液が満たされたチューブは導電性が高い ためインキュベータのハムノイズを拾ってしまう。これを防ぐため、チューブはシャーレ側 で一度点滴による絶縁を行っている。さらに排出用の管は減圧に保ち、注入用管よりも先端 を上に設置した。注入によって培養液面が増加すると排出用針の口に接するので、古い培養 液は自動的に排出される。また、これによってシャーレ内の培養液量を一定に保っている。 このシステムによって培養液が自動的に潅流し、安定状態が保たれている。培養は一般的 な培養環境であるCO2が10%、37℃の飽和水蒸気雰囲気下で行った。図 4-4 培養液潅流システム シリンジポンプ 全体写真 インキュベータ内 測定基盤と注入・排出用管付キャップ



![図 3-11 スパイクソーティング結果 0 0.4 [msec] 0 60[μV] 神経細胞 A 神経細胞B スパイク幅 スパイク幅 振幅 振幅 60[μV] 0.4 [msec] 発火間隔分布(全スパイク) 20[msec] 0 0.1 相対度数 0.1 0 20[msec] E0 ch E0 ch 51 DIV](https://thumb-ap.123doks.com/thumbv2/123deta/6701740.705409/31.894.146.739.322.928/スパイクソーティングスパイクスパイクV発火間隔分スパイクDIV.webp)
![図 3-12 分離された活動電位波形 図 3-13 推定された神経細胞の信号伝搬の様子 神経細胞A神経細胞B神経細胞C神経細胞D0.120.240.400.280.440.360.200.120.400.280.560.040.080.24 (数字は遅延時間を表す[msec]) 25msec-100μV100μVB0chB1chD0chE0chE1chF1chG0chH0ch細胞A 細胞A 細胞A 細胞B 細胞B](https://thumb-ap.123doks.com/thumbv2/123deta/6701740.705409/32.894.160.728.135.1072/信号様子神経細胞A細胞B神経細胞C神経細胞D時間表すVVBB.webp)