オートファジーによるRef(2)P/p62の除去を介した
常在菌寛容機構の解明
著者
長井 広樹
学位授与機関
Tohoku University
学位授与番号
11301甲第19205号
URL
http://hdl.handle.net/10097/00129260
博士論文
オートファジーによる
Ref(2)P/p62 の除去を介した
常在菌寛容機構の解明
令和元年度
東北大学 大学院薬学研究科
生命薬科学専攻
長井 広樹
オートファジーによる
Ref(2)P/p62 の除去を介した
常在菌寛容機構の解明
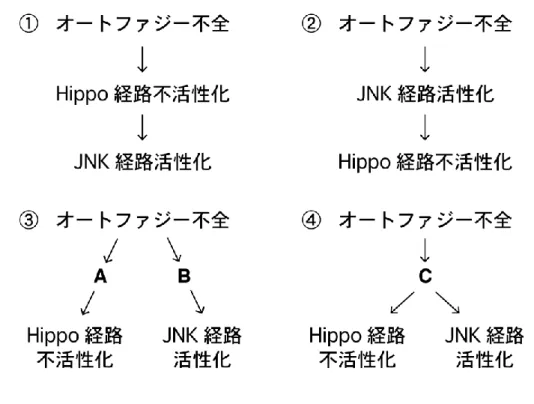
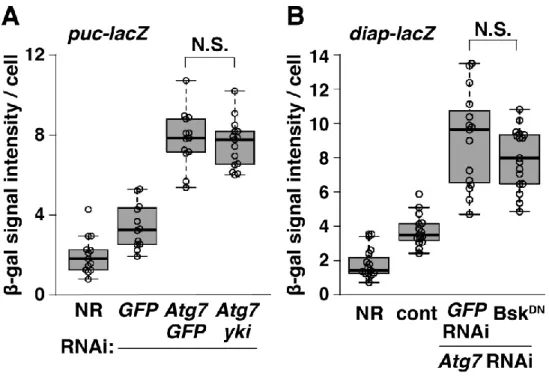
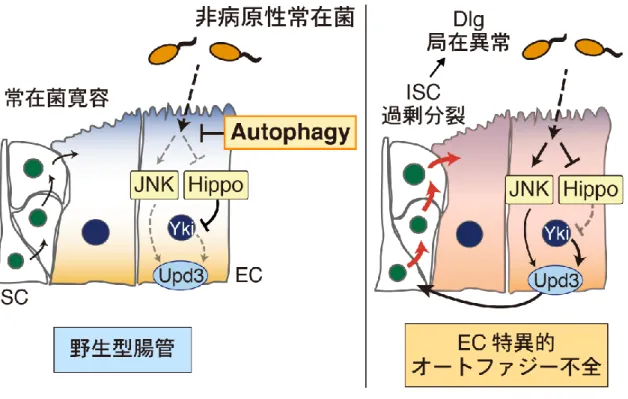
生命機能解析学分野 長井 広樹 腸管には常在細菌が定着しており、宿主は病原性細菌を排除しつつ常在菌に対し て寛容性を示すことで腸管恒常性を維持している。しかし、遺伝子の変異により宿 主の寛容機構が破綻した際など、常在菌は状況依存的に宿主の腸管上皮に炎症を起 こしうる。宿主の遺伝的背景と腸内細菌の相互作用は炎症性腸疾患の発症に寄与す るため、そのメカニズムを解明することは極めて重要な課題である。 近年、そうした宿主側の遺伝的要因に、細胞内分解系であるオートファジーの機 能不全が関与することが報告された。腸管上皮特異的なオートファジー不全モデル マウスを用いた解析から、オートファジー不全が腸内細菌依存的に腸管炎症を亢進 することが示されている。しかし、その発症メカニズムは全く不明である。哺乳類 では、一度腸管炎症が起こると、腸管上皮直下に控える獲得免疫系細胞によって炎 症が増悪されてしまう。これが、腸管上皮におけるオートファジー不全によって生 じる腸管炎症の根本的な原因に迫ることを困難にしていると考えられる。そこで、 本研究では、自然免疫のみで生体防御を行うショウジョウバエ個体をモデル系とし て用い、宿主のオートファジー不全と腸内細菌の相互作用によって腸管恒常性が破 綻するメカニズムを解明することを目的とした。 1. オートファジーは常在菌に対する損傷応答の寛容性に必要である 当研究室では、腸管幹細胞(ISC)から分化した腸管上皮細胞(EC)特異的に オートファジー関連因子(Atg7 など)をノックダウンすることで、腸管炎症誘起 剤DSS 経口摂取時の生存率が低下することが見出されていた。本研究では、この EC 特異的なオートファジー不全個体をモデル系として用いることとした。 DSS は腸管上皮に損傷を与える薬剤であるため、まず EC 特異的オートファジー 不全腸管における損傷応答、すなわち ISC 分裂活性を検討した。その結果、DSS を摂取させずとも、オートファジー不全 EC はサイトカイン分泌を介して ISC 分 裂を亢進させ、腸管上皮の異形成を起こすことが明らかになった。野生型腸管では、 ISC 分裂の亢進は病原菌感染時の損傷修復応答として起こる。オートファジー不全 腸管の ISC 分裂亢進は腸内細菌依存的であった一方で、腸内細菌叢の変化は検出されなかった。驚くべきことに、非病原性の常在菌である Acetobacter 属菌(A. pomorum)による腸内細菌叢の再構築は、オートファジー不全腸管の ISC 過剰分 裂と異形成を起こすのに十分であっ た(Fig.1)。さらに、オートファジー 不全EC では、細胞増殖の抑制性シグ ナルである Hippo 経路が常在菌存在 下で不活性化し、ISC 過剰分裂を起 こしていた。これらの結果から、常在 菌は宿主 EC の損傷応答を刺激して おり、オートファジーが常在菌に対す る過剰な損傷応答を抑制する(寛容性 をもたらす)ことで、腸管恒常性を維 持していると考えられた。 2. オートファジーは Ref(2)P オリゴマーを除去することで寛容性をもたらす オートファジーによる常在菌に対する損傷応答制御メカニズムに迫るにあたっ て、オートファジーの分解基質であるRef(2)P(哺乳類 p62 オルソログ)に着目し た。Ref(2)P は自己相互作用によりオリゴマー化するとともに、シグナル伝達因子 と相互作用してシグナル伝達の足場タ ンパク質として機能する。注目すべき ことに、Ref(2)P は培養細胞において Hippo 経 路 を 不 活 性 化 す る Dachs (atypical myosin)と共免疫沈降され ることが報告されている。そこで、ま ずEC における Ref(2)P と Dachs の局 在を観察したところ、Ref(2)P オリゴマ ーがDachs と共局在していた。また、 EC 特異的オートファジー不全腸管で ISC 過剰分裂によって起こる異形成は、 ref(2)P または dachs のノックダウンに よって緩和された(Fig.2)。これらの結
果から、オートファジー不全EC では Ref(2)P オリゴマーが蓄積することで Dachs により Hippo 経路が不活性化され、過剰な損傷応答が起こると考えられた。さら に、オートファジー不全EC では Ref(2)P と共局在する Dachs のシグナルが増加 したことから、オートファジーがRef(2)P オリゴマーと Dachs タンパク質の両者 を同時に量的制御することで損傷応答を制御していることが明らかになった。 3. 常在菌依存的な ROS 刺激は TCJ 近傍で Ref(2)P オリゴマー形成を起こす 上皮組織の損傷応答誘導には多くの場合、活性酸素種 ROS が寄与する。EC 管 腔側の細胞膜には腸内細菌に対する ROS 産生酵素 DUOX が発現しており、病原 体感染に応じてROS 産生を亢進させることが知られている。オートファジー不全 腸管におけるRef(2)P オリゴマーを介した過剰な損傷応答に DUOX が寄与するか 検討した結果、DUOX をノックダウンす ることで Ref(2)P オリゴマーが減少し、 ISC の過剰分裂もキャンセルされた。一 方で、オートファジー不全はDUOX によ るROS 産生を亢進させなかった。よって、 オートファジー不全は Ref(2)P タンパク 質の蓄積を起こすことで、常在菌存在下 で産生される微量の ROS に対する宿主 の感受性を亢進させると考えられた。実 際に、DUOX 依存的な ROS 産生を誘導 することが知られるuracil を、無菌的に 飼育した個体に経口摂取させると、野生 型腸管では異形成が見られないのに対し、 オ ー ト フ ァ ジ ー 不 全 腸 管 で は 顕 著 な Ref(2)P オリゴマー形成とともに異形成 が生じた。興味深いことに、この時、野 生型腸管でも軽度な Ref(2)P オリゴマー 蓄積が観察され、それらは3 細胞結合部 位(tricellular junction, TCJ)の近傍に 局在していた(Fig.3)。DUOX が産生す
るROS の刺激は管腔から EC へ与えられる。また、TCJ は EC の管腔側に形成さ れる細胞間結合である。そこで、TCJ が Ref(2)P オリゴマー形成に寄与している かを検討した。TCJ の構成因子であるGliotactin(Gli )を EC 特異的にノックダ ウンしたところ、オートファジー不全EC における Ref(2)P オリゴマー形成、Hippo 経路の不活性化がともにキャンセルされた。よって、EC には管腔から与えられる ROS 刺激に応じて Ref(2)P オリゴマーを形成させる特殊な場が存在することが示 された。 以上の結果から、EC においてオートファジーは非病原性の常在菌に対する損傷 応答の寛容性をもたらすことで、腸管恒常性を維持していることが明らかとなった。 さらに、そのメカニズムとして、オートファジーが常在菌依存的なROS 刺激によ って形成される Ref(2)P オリゴマーを除去することで Hippo 経路を制御している ことを示した(Fig.4)。また、常在菌依存的な ROS 刺激は、EC 管腔側の TCJ 近 傍においてRef(2)P オリゴマー形成を誘導し、EC の管腔側領域が常在菌寛容を制 御する場として重要であることを見出した。これらの知見は、宿主のオートファジ ーと腸内細菌の相互作用による腸管恒常性維持機構に、常在菌に対する損傷応答の 寛容性という新たな概念とそのメカニズムを示した。今後、本研究の成果を哺乳類 へと応用することで、炎症性腸疾患の病態の理解と、それに基づく治療戦略の開発 が進むことが期待される。
略語一覧
A911 Acetobacteraceae strain EW911
AJ Adherens junction
A. pomorum Acetobacter pomorum AR Axenically reared Atg Autophagy-related -gal -galactosidase
Bsk Basket CR Conventionally reared DAP Diaminopimelic acid DAPI 4',6-diamidino-2-phenylin
dole
Df Deficiency diap death-associated inhibitor of apoptosis Dlg Discs-large Dpp Decapentaplegic DSS Dextran sulfate
sodium
dTRAF2 Drosophila TNF-receptor associated factor 2
DUOX Dual oxidase EB Enteroblast
EC Enterocyte EGFR Epidermal Growth Factor Receptor
ERK Extracellular Signal- regulated Kinase
FLP Flippase
Ft Fat GFP Green fluorescent protein Gli Gliotactin HM Hypomorphic
IEC Intestinal epithelial cell IFN Interferon
IL Interleukin Imd Immune deficiency ISC Intestinal Stem Cell JNK c-jun N-terminal kinase
Lgl Lethal giant larvae LIR LC3-Interacting Region LPS Lipopolysaccharide PB1 Phox and Bem1
PBS Phosphate buffered salts
PCR Polymerase chain reaction
PFA Paraformaldehyde PGN Peptidoglycan PGRP Peptidoglycan-
recognition protein
PH3 phospho-Histone H3
Puc Puckered Ref(2)P Refractory to sigma P ROS Reactive oxygen species rRNA Ribosomal ribonucleic acid
SNP Single Nucleotide Polymorphism
SJ Septate junction
TA Transit-amplifying cell TJ Tight junction TCJ Tricellular junction TLR Toll-like receptor TNF Tumor necrosis factor Upd Unpaired
UAS Upstream activation sequence
UBA Ubiquitin-associated
UPR Unfolded protein response
WT Wild type
目次
要旨 略語一覧 目次 序論 1. 宿主と腸内細菌の相互作用による腸管恒常性維持 1 2. 炎症性腸疾患クローン病とオートファジー 10 2-1. 炎症性腸疾患クローン病 10 2-2. オートファジーの機能とメカニズム 12 2-3. ショウジョウバエにおけるオートファジーの保存 15 2-4. オートファジー不全が起こす腸管炎症に関するこれまでの知見 16 3. モデル生物としてのショウジョウバエ腸管の有用性 25 3-1. ショウジョウバエ腸管と哺乳類腸管の形態的類似性 25 3-2. ショウジョウバエ腸管における抗菌応答と常在菌寛容 26 3-3. ショウジョウバエ腸管上皮の損傷修復機構の哺乳類との類似性 30 4. 本研究の目的と当研究室における取り組み 37 本論 第一章 EC 特異的なオートファジー不全腸管では非病原性の常在菌依 存的に密着結合構成因子の局在異常が起きる 45 第一節 EC 特異的オートファジー不全モデルを有用性確認および DSS 感受性に関与する遺伝子のゲノム網羅的スクリーニング 46 第二節 EC 特異的なオートファジー不全は密着結合構成因子の局在異 常を起こす 54第三節 オートファジー不全による Dlg 局在異常は腸内細菌依存的であ る 60 第四節 第五節 第六節 第七節 オートファジー不全腸管の腸内細菌叢は菌の構成・量ともに野 生型と比べてほとんど変化しない 単一の非病原性常在菌による腸内細菌叢の再構築はオートファ ジー不全腸管のDlg 局在異常を起こすのに十分である 持続的なEC 特異的オートファジー不全が Dlg 局在異常を起こ す 考察 62 64 66 69 第二章 オートファジー不全腸管では非病原性常在菌に対してISC 分裂 が過剰亢進することでDlg 局在異常が起こる 75 第一節 EC 特異的オートファジー不全腸管では常在菌依存的に ISC 分 裂が亢進する 77 第二節 EC 特異的オートファジー不全はupd3 を介して ISC 過剰分裂お よびDlg 局在異常を起こす 79 第三節 ISC 分裂の過剰亢進が Dlg 局在異常を起こす 83 第四節 第五節 EC 特異的オートファジー不全は ISC の EC への分化には異常 を起こさない 考察 86 92 第三章 オートファジー不全EC では常在菌に対して損傷応答が誘導さ れることで過剰なISC 分裂が起こる 99 第一節 オートファジー不全 EC では常在菌に対し Hippo 経路の不活性 化とJNK 経路の活性化が起きる 103 第二節 Hippo 経路不活性化・JNK 経路活性化の両者がオートファジー 不全腸管のISC 過剰分裂に寄与する 109
第三節 オートファジー不全による Yki や JNK の活性化は互いの転写活 性に依存せずに起こる 114 第四節 考察 117 第四章 常在菌依存的な Ref(2)P のオリゴマー化がオートファジー不全 腸管の過剰な損傷応答を起こす 120 第一節 オートファジー不全腸管における Ref(2)P のオリゴマー化は常 在菌由来の刺激によって促進される 122 第二節 Ref(2)P オリゴマーの蓄積が腸管恒常性を破綻させる 124 第三節 第四節 オートファジー不全腸管における Ref(2)P を介した過剰な損傷 応答にはDachs および dTRAF2 が関与する Ref(2)P オリゴマーは EC において Dachs と共局在する 133 143 第五節 考察 146 第五章 オートファジーは Ref(2)P を除去することで常在菌依存的な ROS 刺激に対する感受性を制御する 151 第一節 第二節 DUOX による ROS 産生がオートファジー不全腸管の恒常性破 綻のトリガーである 腸内細菌への抗菌ペプチド応答および Nrf2 系はオートファジ ー不全腸管の恒常性破綻に関与しない 153 157 第三節 オートファジー不全は DUOX による ROS 産生を亢進しない 163 第四節 オートファジー不全腸管では ROS 感受性が亢進している 165 第五節 オートファジー不全腸管ではDUOX を介して Hippo 経路不活 性化プラットフォームが蓄積する 167 第六節 考察 171
第六章 TCJ が DUOX 依存的な ROS 刺激に応じた Ref(2)P オリゴマー 形成の場となる
第一節
第二節
DUOX 依存的な ROS 刺激は TCJ 近傍で Ref(2)P オリゴマー形 成を起こす
TCJ は DUOX 依存的な ROS 刺激に対する Ref(2)P オリゴマー 形成に必須である 182 185 第三節 TCJ はオートファジー不全 EC における Hippo 経路不活性化に 必須である 193 第四節 第七章 第一節 第二節 第三節 第四節 考察 管腔側から与えられるROS 刺激に応じた Ref(2)P オリゴマー形 成に関与する遺伝子の網羅的探索 H2O2経口投与時のオートファジー不全個体の個体死はref(2)P に完全に依存している スクリーニングのストラテジーと途中経過 Ref(2)P オリゴマーを TCJ に限局させる遺伝子変異はオートフ ァジー不全腸管のDlg 局在を回復させる 考察 195 198 199 205 212 217 結論と展望 224 材料と方法 235 参考文献 249 発表論文リスト 270 謝辞 271
序論
1. 宿主と腸内細菌の相互作用による腸管恒常性維持 生物は常に細菌やウイルスなどの微生物と接しており、それら異物からの刺 激に暴露されている。病原体が体内に侵入した際には、細胞や組織が損傷を受 けるため、宿主は病原体を排除するとともに病原体から受けた損傷を修復する 機構を有している。一方で、外界と接している上皮組織には、無数の細菌が宿 主から排除されることなく常在している。これは、常在菌と宿主が互いの生存 に有利になるような共生関係を確立した結果であり、細菌側が自身の病原性を 抑制し、宿主側が細菌に対する排除応答を抑制することで成立している。この ような宿主と常在菌の共生関係は、特に腸管上皮において顕著に発達している。 ほぼ全ての後生動物の腸管には腸内細菌が定着している。我々ヒトの腸管管 腔にも、およそ 100 兆と推定される、一個体の総細胞数をはるかに上回る数の 腸内細菌が常在している(1)。腸内常在菌は宿主の感染防御に重要な役割を果 たしていることが近年続々と明らかにされている。例えば、腸内常在菌は外来 の病原菌と管腔内の限られた代謝物を巡って競合し、また抗生物質を産生する ことで病原菌の定着を阻害する(2)。このような直接的な感染防御に加え、常 在菌は宿主の免疫応答を調節することでも病原菌感染防御に寄与する。例えば、 常在菌の定着は宿主の腸管上皮細胞からの抗菌ペプチドRegIIIγ の産生を促し、 Enterococcus faecalis などの感染に対する抵抗性を上昇させる(3)。また、腸 管上皮直下の粘膜固有層には多数の免疫系細胞が控えているが、自然免疫系細 胞であるマクロファージにおける IL-1前駆体の合成も常在菌によって促進さ れる。これは、病原菌感染に応じて即座に成熟型IL-1を分泌し免疫応答を活性 化させることを可能にしている(4)。さらに、腸内常在菌は宿主の免疫応答のみならず、代謝など、個体の健常状態を維持するために必須の現象にも必要と されることが明らかにされている(5)。 このように腸内常在菌は宿主の恒常性維持に極めて重要な役割を果たしてい るが、宿主が常在菌から恩恵を受けるためには、常在菌に対して病原菌感染時 に起こすような防御応答を抑制し、常在菌を管腔内に定着させることが必要で ある。腸管上皮細胞は、腸管に感染した病原菌に対しては抗菌ペプチドや活性 酸素種(reactive oxygen species, ROS)を分泌することでこれを排除する。し かし、これらの抗菌応答は常在菌に対しては誘導されず、いわゆる常在菌寛容 (免疫寛容)の状態となっている。抗菌応答の寛容性は、①腸管上皮と腸内細 菌を隔てる粘液層(6)、②腸管上皮による物理的バリアーと細菌認識分子の発 現制御(7)、③腸管上皮下に控える制御性 T 細胞による免疫抑制(1)によって 確立される。 粘液層を構成する主要な因子は糖タンパク質 Mutin である。Mutin は多量の 糖鎖修飾を受け粘性を増すため、Mutin の集積によって粘液層が形成される。 腸管上皮では、分泌性細胞の1 種である Goblet 細胞(杯細胞とも呼ばれる)が Mutin を管腔側へと分泌することで粘液層の形成に寄与している。粘液層は腸 管上皮と腸内細菌を隔てる障壁として機能している。宿主の免疫応答は細菌が 宿主の腸管上皮細胞あるいは粘膜固有層の免疫系細胞に認識されることで活性 化するため、粘液層は常在菌に対する免疫寛容の第一線を担っていると言える。 また、粘液層は抗菌ペプチドや IgA 抗体の濃度が高まっており、病原菌に対す る感染防御の場としても重要である。興味深いことに、常在菌が分泌する酪酸 などの短鎖脂肪酸はMutin の分泌を亢進させることが知られており、常在菌も 粘液層の維持、そして自身に対する免疫寛容の形成に寄与していると考えられ ている(6)(Fig.1-1A)。 仮に常在菌が粘液層を突破しても、宿主の腸管上皮細胞は免疫応答を過剰に活
性化させないシステムを有している。第一に、腸管上皮細胞は隣り合う細胞同 士が細胞間結合によって密接に結合することで、常在菌の体腔への透過を防ぐ 強固なバリアーを形成している。これにより、常在菌が粘膜固有層に控える免 疫系細胞によって認識されることを防いでいる。第二に、腸管上皮細胞では、 細菌を認識して自然免疫を活性化させるToll-like receptors(TLRs)の apical 側(管腔側)への発現が制限されている。生物種によって分子種に差があるも のの、例えばヒト小腸上皮細胞では、細菌の構成成分であるリポポリサッカラ イド(LPS)やフラジェリンを認識する TLR4 や TLR5 が管腔側に発現せず、 基底膜側にのみ発現している(7)。ただし、腸管上皮組織の中でも免疫応答に 特化した領域であるパイエル板では、TLRs が apical 側に発現しているため、 粘液層を突破した常在菌あるいは病原菌に対して全く免疫応答が活性化しない わけではない(7)。このような TLR 局在は、常在菌に対する免疫応答を最小限 に止めることに寄与していると考えられている(Fig.1-1B)。 粘液層や腸管上皮細胞における自然免疫に関する免疫寛容機構に加え、腸管 上皮直下の粘膜固有層では、獲得免疫系である制御性 T 細胞(Treg)による免 疫寛容機構が働いている(Fig.1-1A)。Treg はエフェクターT 細胞や B 細胞に 作用することで、自身が認識する抗原に対する免疫応答を抑制する。常在菌由 来の抗原に富む腸管上皮組織では、これらの抗原に対してTreg が優先的に誘導 される。これは、腸管上皮細胞や、粘膜固有層に存在する間質細胞が分泌する サイトカインによって、抗原提示を行う樹状細胞がTreg を選択的に誘導する能 力を獲得するためである。常在菌が分泌する短鎖脂肪酸もまた、樹状細胞のTreg 誘導能に寄与している。通常Treg は胸腺で分化誘導されるが、腸管においては 胸腺と独立してTreg を誘導するための機構が備わっていることで、常在菌寛容 に貢献している(1)。
以上のように、腸管上皮組織には常在菌に対する抗菌応答を抑制するための 多段階の機構が備わっているが、病原菌感染時の腸管上皮の防御応答には、抗 菌応答に加えて、損傷修復応答が重要である。すなわち、病原菌が産生する毒 素や、宿主自身の炎症応答によって損傷を受けた腸管上皮を修復し、バリア機 能を維持する必要がある。腸管上皮では分裂能を持つ細胞は限られており、哺 乳類では腸管幹細胞(intestinal stem cell, ISC)と、ISC の非対称分裂によっ て新生される前駆細胞(transit-amplifying cell, TA)のみが分裂能を持つ (Fig.1-2)。前駆細胞は、主に消化産物の吸収を担う腸管上皮細胞(enterocyte, EC)、または Goblet 細胞のような分泌性細胞へと分化し、これら分化した細胞 種は分裂能を持たない。したがって、腸管上皮の損傷を修復するためには、損 傷を受けた EC や分泌性細胞がアポトーシスによって上皮から脱離するととも に、ISC や TA の分裂が活性化することで、失われた上皮細胞を補う必要がある (8)。前述の腸管上皮細胞基底膜側での TLR 発現は ISC 分裂の誘導にも寄与し ており、損傷によって管腔内の常在菌や病原菌が基底膜側へ移行すると、これ をTLR が認識して ISC へ分裂シグナルが伝達される(7)。このように自然免疫 応答は病原菌感染時の損傷修復にも寄与している。一方で、ISC 分裂の過剰な 活 性 化 は 腸 管 上 皮 の 過 形 成 (hyperplasia、上皮組織の 肥大化) や異形成 (dysplasia、細胞の形態異常による組織構造の変化)、最悪の場合はガン化に結 びつく。したがって、病原菌感染によって損傷を受けた際には ISC 分裂を活性 化させる必要があるが、非損傷時には ISC 分裂の活性を制御することが重要で ある。常在菌は腸管上皮に損傷を与えないため、宿主の損傷応答を活性化させ ることはないと考えられているが、宿主側に常在菌に対する損傷応答を抑制す る機構が備わっているのかは不明である。 上述してきたように、宿主には腸内常在菌に対する免疫寛容が備わっており、
常在菌側も通常は宿主に対して病原性を発揮せず、宿主の恒常性維持に寄与し ている。しかし、腸内常在菌は常に宿主にとって有益な存在ではなく、宿主と 常在菌の関係は状況依存的なものであることが明らかにされている(1, 9)。例 えば、持続的あるいは過剰な抗生物質の投与は常在菌の多様性や菌数を減少さ せるが、この時、常在細菌の1 種であるClostridium difficile の腸内細菌に占め る割合が増加すると、大腸炎が起こりうる。また、免疫抑制性サイトカインで あるIL-10 を欠損したマウスや、獲得免疫系細胞を欠損した Rag2 ノックアウト マウスでは、常在細菌 Helicobacter hepaticus によって腸管炎症や大腸ガンが 多発する。これらのことから、腸内常在菌の中には、腸管内の環境変化や宿主 の遺伝的要因によって、状況依存的に病原性を発揮する菌種が存在すると考え られる。このような常在菌は、宿主に害を与えない共生細菌(symbiont)に対 して、病原性共生細菌(pathobiont)と定義されている(10)。健常時、pathobiont はsymbiont と比べて腸内細菌叢に占める割合が小さい。こうした健常な腸内細 菌叢の状態はsymbiosis と呼ばれる。逆に、常在菌の多様性が失われ pathobiont の割合が増加した状態や、腸内細菌の総数が減少あるいは増加した状態は dysbiosis と呼ばれる。 Dysbiosis は宿主の腸管恒常性破綻と密接に結びついており、前述の通り pathobiont の増加は腸管炎症を引き起こす。また、腸内細菌数の減少は外来の 病原菌との競合力を低下させ感染率を高める。逆に腸内細菌の増加は、宿主に 与えられる常在菌由来の免疫活性刺激(抗原など)の増大によって、炎症反応 を亢進させる。腸管炎症には獲得免疫系が寄与することが古くから知られてい るが、近年では自然免疫系も炎症に深く関わっていることが明らかにされてい る。例えば、前述したRag2 ノックアウトマウスにおける Helicobacter hepaticus 依存的な腸管炎症には、TLR シグナルによる自然免疫系の過剰な活性化が寄与 することが示されている(11)。このように、腸管炎症においては、健常時は抑
制されているはずの宿主の免疫応答が過剰に活性化している。しかしながら、 重要な問題として、宿主の常在菌に対する過剰応答や常在菌のdysbiosis が、腸 管炎症の原因となっているのか、あるいは炎症の結果として起きているのか、 その因果関係は明らかにされていない(12, 13)。これは、腸管炎症によって腸 管上皮のバリア機能が低下すると、管腔内の抗原が粘膜固有層へ透過すること で免疫系が活性化し、炎症を激化させるとともに、腸内細菌叢を変化させてし まうためである。また、腸内細菌叢が病原性の高い構成に変わる (つまり dysbiosis が起こる)ことでも、炎症応答が亢進してしまう。このように dysbiosis と宿主の炎症応答が互いに影響を与え合うことで、腸管炎症の根本的な原因に 迫ることが困難となっている。この問題を解決することは、宿主と腸内常在菌 の相互作用による腸管恒常性維持機構の理解を深めるとともに、根治療法の存 在しない炎症性腸疾患に対する新たな治療法を確立する上で極めて重要である。
Fig.1-1 哺乳類腸管における免疫寛容機構
(A)腸管上皮を構成する Goblet 細胞は Mutin を含む粘液を放出することで粘 液層を形成する。粘液層は常在菌が腸管上皮細胞と直接的に接触しないための 障壁となっている。Goblet 細胞の Mutin 産生は腸内常在菌が分泌する短鎖脂肪 酸によって促進されることが知られている。また、短鎖脂肪酸は粘膜固有層に おける制御性 T 細胞(Treg)の誘導も促進する。腸管管腔には常在菌由来の抗 原が豊富に存在し、これを受容した樹状細胞(dendritic cell, DC)は T 細胞の 分化を誘導する。この時、短鎖脂肪酸によってエフェクターT 細胞ではなく Treg
への分化が促進される。Treg はエフェクターT 細胞や B 細胞を抑制することで 常在菌に対する免疫応答を抑制している。なお、画像は参考文献(6)より一部 改定(文字の拡大、日本語訳)している。 (B)ヒト小腸上皮細胞における TLR 発現の模式図を示した。TLR2, TLR3, TLR4, TLR5 は腸管上皮細胞の apical 側(管腔側)には発現しない。なお、本 Figure にはパイエル板は示していないが、パイエル板では apical 側に TLR が 発現している。画像は参考文献(7)のものを一部改定している。
Fig.1-2 哺乳類腸管上皮の細胞系列
哺 乳 類 の 腸 管 上 皮 を 構 成 す る 細 胞 系 列 の 模 式 図 を 示 し た 。 腸 管 幹 細 胞 (Intestinal Stem Cell, ISC)は非対称分裂によって ISC 自身と前駆細胞 (Transit-Amplifying cell, TA)を生み出す。TA も分裂能を持つが、その後腸 管上皮細胞(Enterocyte, EC)や分泌性細胞へ分化していく。分泌性細胞には 自然免疫応答を担うパネート細胞(Paneth Cell, PA)や粘液産生を担うゴブレ ット細胞(Goblet Cell, GC)、また図には示していないが、ホルモン分泌を担う Enteroendocrine cell や、寄生虫に対する免疫応答の活性化を担うタフト細胞 (Tuft cell)が存在する。ISC, TA, PC は絨毛よりもさらに底部の腸陰窩に位置 しており、EC や GC は絨毛に位置している。
2. 炎症性腸疾患クローン病とオートファジー 2-1. 炎症性腸疾患クローン病 腸管上皮組織の損傷に付随して免疫系による炎症反応の活性化が見られる疾 患を総称して炎症性腸疾患と呼ぶ。炎症性腸疾患は厳密には、病原体感染や薬 剤など損傷の原因が特定できる特異性炎症性腸疾患と、原因が特定されていな い非特異性炎症性腸疾患に分類されるが、一般的に炎症性腸疾患と言うと非特 異性のものを指すことが多い。本論文でも、非特異性炎症性腸疾患を炎症性腸 疾患と呼ぶこととする。 炎症性腸疾患の代表例としてクローン病が挙げられる。クローン病は口腔から 肛門の消化管全域において非連続性(病変部と非病変部が混在している)の炎 症を引き起こす慢性の炎症疾患である(12)。病変部では腸管壁の潰瘍が見られ、 症状の悪化とともに腸閉塞、腸穿孔が生じ、消化管からの大出血が生じること もある。日本においてクローン病の罹患者数は増加の一途をたどっており、1976 年では128 人だったのに対し、2014 年では 42397 人であり、人口 10 万人あた り約 33 人がクローン病を患っていることが報告されている(14)。また欧米で は人口10 万人あたりの罹患者数が約 200 人と日本の 6 倍近くに上っている。 クローン病は国内のみならず世界的に見ても効果的な治療法の確立が強く望 まれる疾患であるが、クローン病発症の分子機構、特に根本的な発症原因は未 だ不明であり、根治療法は存在しない。クローン病の治療は薬による化学療法 が一般的だが、免疫抑制剤などの対症療法であるため、投薬を行っても寛解と 再発を繰り返してしまう。近年では、クローン病における炎症反応に炎症性サ イトカインTumor Necrosis Factor-(TNF-)が寄与することから、抗 TNF- 抗体や、TNF-の受容体との結合を競合阻害する受容体製剤が開発され、寛解 状態の長期維持が可能となった(15)。また、クローン病のリスク遺伝子であ
るIL-23R, JAK2, STAT3(いずれも Th17 細胞の分化誘導に寄与)に対する阻 害薬でクローン病の寛解導入効果が認められ(16)、抗 TNF-抗体に続く治療 戦略として注目されている。しかしながら、これらの療法では本来は生体の防 御応答に機能する免疫反応を抑制するため、副作用として感染症や肺炎が起こ る危険性がある。したがって現在でも、より効果的で安全性の高いクローン病 治療薬の開発が求められており、そのためにはクローン病の発症メカニズムの 解明が必要である。 クローン病の発症には環境的な要因と遺伝的な要因の両方が関与すると考え られている。環境的な要因としては腸内細菌の寄与が大きく、種々のクローン 病モデルマウス(TNF-高発現マウスなど)における腸管炎症の病理スコアは、 無菌的に飼育することでほぼ完全に回復することが明らかにされている(17, 18, 19 ) 。 ま た 、 ク ロ ー ン 病 罹 患 者 の 腸 内 細 菌 叢 で は 、 symbiont と さ れ る Bifidobacterium 属(ビフィズス菌)、Lactobacillus 属(乳酸菌)が減少し、 病原性大腸菌の1 種である Adherent-invasive E. coli などが増加する dysbiosis が検出される。前述の TNF-高発現マウスにおいても dysbiosis が見られ、こ の個体の糞中の菌を、無菌的に飼育したTNF-高発現マウスに経口投与すると、 炎症が再発することが確かめられている(17)。しかしながら、dysbiosis がク ローン病の発症原因であるのか、炎症の増悪に寄与しているのかは未だに不明 である。 クローン病の遺伝的リスクに関して、近年ゲノムワイド関連解析によるクロー ン病罹患者と健常者の一塩基多型(SNPs)の解析が報告され、注目を集めてい る。クローン病罹患者で多く見られるSNP には、前述したIL-23R などの免疫 関連遺伝子に加え、細胞内分解系であるオートファジーに関与する ATG16L1, NOD2, IRGM が含まれていた(20, 21)。
2-2. オートファジーの機能とメカニズム オートファジーは細胞内の成分をリソソーム(酵母や植物では液胞)へと輸 送して分解する、生物種を越えて高度に保存された細胞内分解系である(22)。 オートファジーは、オートファゴソームと呼ばれる二重膜で細胞内成分を隔離 し分解へと導くマクロオートファジー、リソソーム膜が湾曲することで細胞内 成分を直接分解するミクロオートファジー、そして分子シャペロンと結合した 基質タンパク質がリソソームへと輸送されるシャペロン介在性オートファジー に大別される。このうち、マクロオートファジーが最も主要であり、以後、本 論文ではマクロオートファジーを指して「オートファジー」と表記する。 オートファジーの分解基質はタンパク質、核酸、脂質、オルガネラ、そして細 胞内に侵入した病原菌と多岐に渡る。特に、ユビキチン–プロテアソーム系では 分解困難な、タンパク質凝集体やミトコンドリアといった巨大な細胞内成分を 分解できることが大きな特徴である。オートファジーによる細胞内成分の分解 は恒常的に機能しており、細胞の構成物質を常に新しいものに作り変えるター ンオーバーを担っている。一方で、オートファジーは細胞に与えられる様々な ストレスに応じて誘導され、例えば、栄養飢餓時には積極的にタンパク質を分 解することでアミノ酸を供給する。また、細胞内に侵入した細菌に対してもオ ートファジーが誘導され、菌を分解することで免疫応答にも寄与している。そ の他、オートファジーはガンや細胞死など、生体の様々な現象に関わることが 明らかにされている(22)。また、ショウジョウバエ個体を用いた解析から、 オートファジー関連因子Atg7 の機能が低下した変異個体では、酸化ストレス誘 起剤であるparaquat(N,N’-dimethyl-4,4’-bipyridinium dichloride)の経口摂 取による個体死が亢進することが示されており(23)、オートファジーが酸化ス トレス応答としても重要であることが知られている。 オートファジーには多数のATG(autophagy-related)遺伝子が関与し、オー
トファジーによる細胞内成分の分解は次のような段階を経る(Fig.1-3)(22, 24, 25)。まず、オートファジー誘導に際して、ATG13 が脱リン酸化を受けること で ATG13 を含む ULK1 複合体(ULK1-Atg13-FIP200-ATG101)が形成され る(induction step)。ULK1 複合体によって、主に小胞体膜上に存在するホス ファチジルイノシトール3 キナーゼ(PI(3)K)複合体(Beclin1, ATG14, VPS34 などから構成される)が活性化することで、隔離膜(phagophore)という扁平 な膜構造が形成される(initiation step)。このとき隔離膜の膜成分の供給には、 ATG 遺伝子の中で唯一の膜タンパク質である ATG9 が寄与すると考えられてい る。隔離膜は徐々に伸長され湾曲していき、膜の両端が融合することで、いわ ゆるオートファゴソームとなる。
隔離膜の伸長過程(elongation step)には ATG12-ATG5-ATG16L1 複合体に よる LC3(ATG8)のホスファチジルエタノールアミン(PE)修飾が重要であ る。PE 修飾を受けていない LC3(LC3-I)は細胞質中に溶解しているが、PE 修飾を受けたLC3(LC3-Ⅱ)は隔離膜に取り込まれる。LC3-Ⅱはオートファゴ ソームがリソソームと融合し内容物の分解が行われるまでオートファゴソーム 膜に存在するため、オートファゴソームのマーカーとして利用される。LC3 の PE 修飾には ATG12-ATG5-ATG16L1 複合体に加え E1 様タンパク質 ATG7 お よびE2 様タンパク質 ATG3 が寄与する。更に、ATG7 は E2 様タンパク質 ATG10 とともにATG12-ATG5-ATG16L1 複合体形成にも必要となる。 オートファゴソームの形成は細胞質中の至る所で起こる一方で、リソソーム は主に核近傍に局在していることが知られている。したがって、完成したオー トファゴソームは、細胞内に張り巡らされた微小管を伝って核近傍へと輸送さ れる必要がある(26)。この輸送は、小胞輸送促進タンパク質 Rab7 を介した オートファゴソームとDynactin-Dynein モーターの相互作用によって担われる。 そして、オートファゴソーム側の Q-SNARE タンパク質 STX17 と、リソソー
ム側のR-SNARE タンパク質 VAMP8 が相互作用することで、オートファゴソ ームとリソソームが融合し、オートファゴソーム内の基質が分解される(24)。 クローン病のリスク遺伝子として同定されたATG16L1, NOD2, IRGM のう ち、ATG16L1 は上述のとおりオートファゴソーム形成に必須な役割を果たす。 また、NOD2 と IRGM は、細胞内に侵入した病原菌に対するオートファジー誘 導 に 機 能 す る 。IRGM は 培 養 細 胞 に お い て 細 胞 内 に 侵 入 し た 結 核 菌 (Mycobacterium tuberculosis)に対するオートファジー誘導に必要とされる ことが2006 年に報告されたが(27)、当時オートファジー誘導のトリガーとな る因子は不明であった。当研究室ではショウジョウバエ個体を用いた解析から、 病原体センサーであるペプチドグリカン認識タンパク質 PGRP-LE が細胞内に 侵入したリステリア菌Listeria monocytogenes を細胞質で認識し、オートファ ジーを誘導することを明らかにした(28)。この発見に続いて、NOD2 が細胞 内の赤痢菌(Shigella flexneri )およびリステリア菌を認識してオートファジ ーを誘導するセンサーであること(29)、IRGM が NOD2 および ATG16L1 と 相互作用することでオートファジー誘導に寄与することが明らかにされている (30)。 通常オートファジーは非選択的に細胞内の物質を包み込むが、細胞内の細菌 に対しては病原体センサーによって特異的に細菌を認識し、その周囲にのみオ ートファゴソームを形成して選択的な分解を行う。また、タンパク質凝集体や、 損傷を受けたミトコンドリアに対しても、オートファジーによる選択的な分解 が起こる(31)。このような選択的オートファジーには、アダプタータンパク 質p62/SQSTM1(以下 p62)が寄与する。p62 はユビキチン化タンパク質と相 互作用するUBA ドメインと、オートファゴソーム膜上の LC3-Ⅱと相互作用す るLIR ドメインを有している。細胞内で異常に凝集したタンパク質や損傷を受
けたミトコンドリアの膜タンパク質はユビチキン化を受けるため、これに p62 が UBA ドメインを介して結合することができる。p62 は更に LIR ドメインを 介してオートファゴソームと相互作用することで、凝集タンパク質の選択的分 解を可能にしている。この時、p62 自身もオートファゴソームに取り込まれ分 解される。つまり、p62 は選択的オートファジーのアダプターであると同時に、 選択的オートファジーの分解基質でもある(Fig.1-4)。オートファジーが不全 になると細胞内で p62 タンパク質が蓄積するが、p62 は自己相互作用が可能な PB1 ドメインを有しているため、蓄積した p62 はオリゴマー化することで凝集 体を形成しうる。オートファジー不全による p62 凝集体の蓄積や不良ミトコン ドリアの蓄積はガンや神経変性に寄与することが知られており(22)、選択的 オートファジーは病原体の排除という自然免疫機能のみならず、広く生体の恒 常性維持に寄与している。 2-3. ショウジョウバエにおけるオートファジーの保存 上述したオートファゴソーム形成に関与する基本的な因子は、本研究で用い るショウジョウバエにも全て保存されている(Fig.1-5)(32)。1 点異なること として、LIR ドメインを持つ選択的オートファジーのアダプター分子が挙げら れる。哺乳類ではp62 に加えて、NDP52, OPTN, NBR1 など多くのアダプター 分子が同定されている(31)。これらの分子は、タンパク質凝集体など同じ基質 と結合する重複性を持ち、特に NBR1 は p62 と同様に PB1 ドメインと UBA ドメインを持ち類似性が高い。これに対し、ショウジョウバエが持つアダプタ ータンパク質は、p62 オルソログである Ref(2)P のみである。Ref(2)P は p62 と 同様に神経細胞において凝集体形成を起こすことが知られており(33)、機能的 にも類似している。よって、ショウジョウバエは Ref(2)P/p62 に焦点を当てた 解析を行いやすいモデルであると考えられる。
2-4. オートファジー不全が起こす腸管炎症に関するこれまでの知見
ゲノムワイド関連解析から同定されたATG16L1 の T300A 変異は、クローン 病の発症率をヘテロ接合で1.4 倍、ホモ接合で 1.9 倍に上昇させる(34)。この T300A 変異によって、ATG16L1 タンパク質は Caspase-3 による分解が促進さ れ、オートファジー不全が起こることが明らかにされている(35)。ATG16L1 がクローン病のリスク遺伝子として同定されて以来、Atg16L1 欠損によるクロ ーン病モデルマウスを用いた解析が精力的に行われてきた。Atg16L1 ノックア ウトマウスはAtg5 や Atg7 のノックアウトマウスと同様に生後 1 日で致死とな る(36)。そのため、ジーンターゲッティングによるAtg16L1 低発現マウスや、 骨髄キメラマウス、腸管上皮特異的なノックアウトマウスを用いたin vivo 解析 が行われてきた。 血球系細胞特異的にAtg16L1 を欠損させたキメラマウスから単離したマクロ ファージでは、LPS 刺激依存的に炎症性サイトカイン Interleukin-1(IL-1) やIL-18 の過剰産生が起こる(36)。また、この血球細胞特異的Atg16L1 欠損 キメラマウスでは、デキストラン硫酸ナトリウム(DSS、経口投与により腸管 炎症を誘発する化合物)経口摂取時の腸管炎症が亢進し、コントロール個体と 比べて生存率が低下する。さらに、この生存率の低下はDSS とともに IL-1や IL-18 に対する中和抗体を投与することでキャンセルされることが示されてい る。これらのことから、Atg16L1 の欠損は、腸管上皮直下の粘膜固有層に浸潤 しているマクロファージからの異常なサイトカイン分泌によって腸管炎症を亢 進させると考えられている(Fig.1-6A)。
一方で、Cadwell らは、Atg16L1 の全身性低発現変異マウス(ATG16L1HM
マウス)を樹立して解析を行なった(18, 37)。このマウスはAtg16L1 ノック アウトマウスと異なり、発生や成長の過程には異常が生じないため、個体レベ ルでの解析が可能である。ATG16L1HMマウスではクローン病において炎症の頻
発する小腸末端部や大腸の上皮細胞で異常は見られなかったが、腸陰窩(crypt) に存在するパネート細胞で異常が見られた。パネート細胞はα-Defensin などの 抗菌ペプチドを顆粒分泌する自然免疫に重要な細胞であり、ATG16L1HMマウス ではパネート細胞からの顆粒分泌に異常が生じていた。パネート細胞からの分 泌 異 常 は ATG16L1 に変異を持つクローン病罹患者においてもみられ、 ATG16L1HM マウスを用いた解析結果が疾患の病態と一致することが示唆され ている(37)。また、ATG16L1HMマウスではDSS 摂取時に腸管の筋層肥大や リンパ球凝集といったクローン病における特徴的な組織病変が見られた。こう したパネート細胞の顆粒分泌異常やDSS 摂取時の組織病変は腸内細菌依存的で あるとともに、マウス飼育環境中に存在するmurine norovirus(MNV)の腸管 への定着に依存していた(18)(Fig.1-6B)。ATG16L1HM マウスでは全身で オートファジー不全が起こっているが、Villin-Cre-LoxP システム(38)により、 腸管上皮を構成する細胞(Intestinal epithelial cell, IEC)特異的にAtg16L1, Atg7, Atg5 をノックアウトした解析も報告されている。これらのマウスでは Atg16L1HMマウスと同様にパネート細胞の顆粒分泌異常が生じることから、パ ネート細胞の異常が腸管外の組織におけるオートファジー不全の影響ではなく、 腸管上皮におけるオートファジー不全によって起きていることが示されている (39)。 パネート細胞の機能不全や腸管炎症の亢進は、ER ストレス応答に寄与する Xbp1 を IEC 特異的にノックアウトすることでも生じる(40)。また、Xbp1ΔIEC マウスの腸管上皮ではオートファジーの誘導が見られ、IEC 特異的に Xbp1 と Atg16L1 をノックアウトすることで、腸管炎症のさらなる増悪とともに ER ス トレスの亢進やNF-B の活性化が観察されている。これらの結果は ER ストレ スの蓄積が起こす腸管炎症をオートファジーが抑制していることを示唆してい るが、Atg16L1ΔIECマウスにおける腸管炎症の亢進がER ストレス依存的である
かは不明である。
以上のように、オートファジー不全は炎症性サイトカインの発現上昇、パネ ート細胞の分泌異常、ER ストレスとの関連が報告されているが、それらが腸管 炎 症 の 原 因 と な っ て い る か は 明 ら か で な い 。 さ ら に 、pathobiont である Helicobacter hepaticus や Toxoplasma gondii の感染実験では、Atg16L1ΔIEC
によってこれらの病原体感染による腸管炎症が亢進する一方で、NF-B の活性 化やER ストレスの亢進は見られない(19, 41)。Atg16L1ΔIECマウスでは、腸 内細菌依存的に TNF によるアポトーシス誘導が亢進することで、pathobiont 感受性が亢進することが示されている(Fig.1-6C)。しかし、これらの解析は 強制感染系であるため、pathobiont が腸管炎症の原因であるかは不明である。 これらの報告から、宿主側のオートファジーが腸内細菌に対する腸管恒常性 維持に機能していることは明らかである。しかしながら、オートファジー不全 によって腸管恒常性が破綻する根本的な原因が何であるのか、またその分子メ カニズムはどのようなものであるのかは解明されていない。マウスでの詳細な メカニズム解析が困難な理由として、① 一度腸管炎症が起こると粘膜固有層に 控える獲得免疫系の過剰な免疫応答によって炎症応答や dysbiosis が増悪され、 オートファジー不全による炎症の原因と結果を区別することが困難であること (12, 13)、② Atg16L1ΔIECマウスなどの遺伝学的な操作を施したマウスの作製 は時間を要し、オートファジー不全を起こした上で別の遺伝子変異を合わせ持 つマウスの作製など、遺伝学的手法が困難であることが挙げられる。特に、前 者の問題はクローン病の治療法を確立する上でも障壁となっており、炎症の結 果起きている現象(つまり炎症を増悪させるような現象)を抑えても、炎症そ のものは根治できないため、根本的な治療は困難である。
オートファジー誘導によってオートファゴソームが形成され、リソソームと の融合によって内容物が分解されるまでの基本的なメカニズムをまとめた。 (1)飢餓などによってオートファジーが誘導されると、ATG13 が脱リン酸化 を受けてULK1 複合体が形成される(induction)。ULK1 複合体は PI(3)K 複合 体を活性化し、隔離膜形成を起こす(nucleation)。隔離膜成分の供給には ATG9 が寄与する。 (2)ATG12-ATG5-ATG16L1 複合体が LC3 の PE 修飾を行い、隔離膜が伸長、 湾曲していく。ATG12-ATG5-ATG16L1 複合体形成や LC3 の PE 修飾は ATG7、 ATG3、ATG10 によって媒介される(elongation)。 (3)隔離膜の両端が融合し,2 重膜構造を有するオートファゴソームが完成す る(completion)。このステップにも ATG12-ATG5-ATG16L1 複合体が寄与す る。 (4)オートファゴソーム膜上の Syx17 とリソソーム膜上の Vamp8 を介して両 者が融合し、内容物が分解される。 なお、本Figure は参考文献(24, 25)のものを一部改定したものである。
Fig.1-4 p62 を介した選択的オートファジーの概念図 細胞内で生じたミスフォールドタンパク質はユビキチン化を受ける。p62 は UBA ドメインを介してユビキチン化タンパク質と相互作用するとともに、PB1 ドメインを介した自己相互作用によって集積する。さらに、p62 は LIR ドメイ ンを介してLC3-II と相互作用することで、オートファゴソームを選択的に誘導 する。選択的オートファジーのターゲットは、ミスフォールドタンパク質の他 にも、損傷を受けたミトコンドリアや細胞内に侵入した病原菌など多岐に渡る。 本Figure は参考文献(42)より一部改定した。
Fig.1-5 オートファジー関連遺伝子のショウジョウバエへの保存
ショウジョウバエ(Drosophila melanogaster)、酵母(Saccharomyces cerevisiae)、ヒト(Homo sapience)でのオートファジーの保存性を表にまと めた(32)。オートファジーの induction, nucleation, elongation, fusion に関わ る遺伝子は高度に保存されている。選択的オートファジーのアダプターについ ては、ショウジョウバエに保存されているのは p62 オルソログである Ref(2)P のみである。
Fig.1-6 オートファジー不全と腸内細菌の相互作用による恒常性破綻
(A)IEC におけるオートファジー不全は、腸内細菌や腸管感染性のウイルス依 存的にパネート細胞からの顆粒分泌に異常を起こす。
(B)血球細胞におけるオートファジー不全を起こしたマウスから単離したマク ロファージでは、LPS 刺激に応じた IL-1発現が上昇する。腸内細菌由来の LPS 刺激によって粘膜固有層のマクロファージが IL-1 を過剰産生することで、腸 管炎症に寄与している可能性が考えられている。
(C)IEC 特異的オートファジー不全は、Helicobacter hepaticus や Toxoplasma gondii といった日和見病原体を強制感染させた際の細胞死を腸内細菌依存的に 亢進する。この細胞死は TNF-α 刺激によって起こることが示されているが、 TNF-α を産生する細胞種は特定されていない。
(A)と(B)は参考文献(43)より、(C)は参考文献(44)より一部改定して いる。
3. モデル生物としてのショウジョウバエ腸管の有用性 3-1. ショウジョウバエ腸管と哺乳類腸管の形態的類似性 宿主と腸内細菌の相互作用による腸管恒常性維持メカニズムを解析する上で、 ショウジョウバエは有用なモデル生物となる。ショウジョウバエ腸管にも無数 の常在菌叢が定着しているが、その構成は比較的シンプルで、ヒトでは 500 種 以上の菌が腸管に定着している一方、野生に生息するショウジョウバエでは約 30 種である。実験室で飼育される場合は 5-20 種と、更に単純化されている。こ のシンプルさに加え、無菌的な飼育法や単一の菌種での腸内細菌叢の再構築と いった実験手法も確立されており、ショウジョウバエは宿主とsymbiont あるい はpathobiont の相互作用を解析しやすいモデルである(45)。 ショウジョウバエでは、ほ乳類の食道、胃、小腸、大腸に相当する器官がそれ ぞれforegut, crop, midgut, hindgut と呼ばれ、ほ乳類の各組織と同等の機能を 有している(46)(Fig.1-7A)。ほ乳類の小腸とショウジョウバエの midgut を比較すると、ショウジョウバエにはほ乳類のような絨毛構造こそないものの、 単層の上皮構造を持つ点は同一である(Fig.1-7B)。また、上皮を構成する細 胞種もよく類似している(Fig.1-7C)。各細胞種は分裂能を持った幹細胞 (Intestinal stem cell, ISC)から新生される。ISC は非対称分裂によって ISC と分化能を持った前駆細胞(ほ乳類: Transit-amplifying cell, TA、ショウジョ ウバエ: Enteroblast, EB)に分裂する。Fig.1-2 でも示した通り、ほ乳類では ISC から新生された TA は腸管上皮細胞(Enterocyte, EC)または、分泌性の細胞 であるパネート細胞(Paneth cell)やゴブレット細胞(Goblet cell)へ分化す る。同様にして、ショウジョウバエでは、ISC から新生された EB は腸管上皮 細胞(EC)または分泌性細胞である Enteroendocrine cell(EE)へ分化する。 1 点異なることとして、哺乳類では前駆細胞 TA も分裂能を持っているが、ショ
ウジョウバエではISC のみが分裂能を有している。この特徴は ISC 分裂に焦点 を当てた解析を容易にしており、2006 年に成虫の midgut における ISC が同定 されて以来(47, 48)、ショウジョウバエ midgut は組織幹細胞分裂による損傷 修復機構の解明に大きく貢献してきた(49)。本研究でも midgut を観察して おり、以下、midgut を指して「腸管」と表記する。 3-2. ショウジョウバエ腸管における抗菌応答と常在菌寛容 細胞系列に加えて、腸内細菌に対する応答もショウジョウバエと哺乳類でよ く類似している。哺乳類の腸管上皮と同様、ショウジョウバエ腸管においても 病原菌に対して抗菌ペプチドや活性酸素種ROS が産生され、病原菌の排除に機 能している(Fig.1-8)。無脊椎動物であるショウジョウバエは獲得免疫を持たず、 自然免疫のみで生体防御を行う。この性質から、ショウジョウバエは個体レベ ルで自然免疫に焦点を当てた解析を行いやすく、自然免疫応答の基本原理と、 その生理学的意義の解明に貢献している。腸管上皮を含む上皮組織では血球系 の免疫細胞とは自然免疫機構が異なる面もあり、この点に関してもショウジョ ウバエで先駆的な報告がなされてきた。 抗菌ペプチド発現を誘導するシグナル経路として、ショウジョウバエではToll 経路とImd 経路があり、それぞれほ乳類では TLR 経路と TNF 経路として保存 されている(50)。Toll 経路は真菌の構成成分である-glucan や、主にグラム陽 性菌が持つリジンタイプのペプチドグリカン(PGN)を認識することで活性化 する。Toll 受容体を介して細胞内へシグナルが伝達されると、転写因子 Dif / Dorsal(NF-B)が活性化することで drosomycin などの抗菌ペプチド発現が 誘導される。一方で、Imd 経路は主にグラム陰性菌が持つ DAP(ジアミノピメ リン酸)タイプのPGN を認識して活性化する。この時、PGN 認識タンパク質 PGRP-LC や PGRP-LE が DAP タイプ PGN を認識し、細胞膜直下に存在する
Imd(ほ乳類では RIP)を活性化する。これによって、dTAK1(TAK1)や IKK 複合体などを介してシグナルが伝達され、転写因子 Relish(NF-B)が活性化 し、diptericin などの抗菌ペプチドの発現が誘導される。Relish は抗菌ペプチド 発 現 を 誘導 す ると 同 時に 、Imd 経路の負の制御因子である PGRP-SC や PGRP-LB の発現も誘導することで、免疫応答の過剰な活性化を抑制しているこ とが明らかにされている(50)。なお、ほ乳類では TLR 経路や TNF 経路が活性 化すると、IL-1 などの炎症性サイトカインが発現誘導される。 ショウジョウバエ腸管においては主にEC が抗菌ペプチド応答を担っている。 Imd 経路の因子の変異体では病原菌の腸管感染時に抗菌ペプチド発現を誘導で きず、生存率が低下することから、Imd 経路が腸管においても抗菌ペプチド産 生経路として機能していることが示されている(51)。上皮組織では抗菌ペプチ ドがToll 経路ではなく Imd 経路によって誘導される例が多数知られており(51, 52, 53)、Toll 受容体変異体の腸管上皮においても、真菌の腸管感染に応じた抗 菌ペプチド発現が見られる。一方で、Imd 経路の細胞内シグナル伝達因子の変 異体では真菌の腸管感染時の抵抗性が低下する(53)。これらのことから、腸管 上皮における抗菌ペプチド応答へのToll 経路の寄与は小さく、Imd 経路が主要 な抗菌ペプチド産生経路だと考えられている。このように、ショウジョウバエ 腸管上皮では少なくともImd 経路によって病原菌に対して抗菌ペプチドが誘導 されているが、非感染時には(つまり常在菌に対しては)抗菌ペプチド発現は 誘導されない。 驚くべきことに、腸内常在菌の定着は、Imd 経路の転写因子 Relish の活性化 を誘導し、Imd 経路の負の制御因子である PGRP-SC や PGRP-LB の発現を上 昇させる一方で、抗菌ペプチドの発現は上昇させないことが報告されている (54)。この常在菌寛容は、個体発生に必須なホメオボックス遺伝子である Caudal が 、 抗 菌 ペ プ チ ド 遺 伝 子 配 列 上の CDRE ( Caudal-protein DNA
recognition element)に結合し、Relish による転写誘導を阻害することによっ てもたらされている(54, 55)。Caudal の発現を抑制した腸管では、常在菌に対 す る 抗 菌 ペ プ チ ド の 発 現 が 上 昇 す る と と も に 、symbiont で あ る Acetobacteraceae strain EW911 ( A.911 ) の 減 少 と pathobiont で あ る Gluconobacter sp. strain EW707 の増加といった dysbiosis が見られる。また、 Caudal ノックダウン個体は寿命が短縮するが、腸内細菌叢を複数の symbiont (Acetobacter pomorum, A.911, Lactobacillus plantarum, L. brevis )で再構 築することによって、この寿命の短縮がキャンセルされる。したがって、宿主 免疫系の常在菌寛容によって symbiosis を維持することは、ショウジョウバエ においても個体の生存に重要であることが明らかにされている(54)。
抗菌ペプチドに加え、EC の抗菌応答には病原菌に対する ROS 産生が寄与す る。EC には腸内細菌依存的に ROS 産生を起こす膜貫通タンパク質として DUOX(dual oxidase)と NOX(NADPH oxidase)が発現している。NOX に よるROS 産生は、symbiont であるLactobacillus plantarum によって促進さ れる。すなわち、L. plantarum が分泌する L-lactate が EC 内に取り込まれ、 NADH 合成に利用されることで、NOX の活性が上昇する(56)。NOX は細胞 内へ向けてROS を産生し、抗酸化応答を担う Keap1-Nrf2 系を恒常的に活性化 していると考えられている(57, 58)。
一方で、DUOX は EC の細胞外、つまり管腔に向けて ROS(H2O2)を産生す
る。DUOX の細胞外ドメインには H2O2 を次亜塩素酸 HOCl に変換する
peroxidase-homology domain があり、HOCl が腸内細菌に対して強い殺菌作用 を発揮する(59)。DUOX は腸管上皮における抗菌応答に大きく寄与している が、ROS に耐性を持つ菌に対しては Imd 経路による抗菌ペプチド応答が必須と される(53)。このように、DUOX による ROS 産生と抗菌ペプチド応答の両者
が揃うことで腸管上皮の抗菌応答が盤石化している。
ショウジョウバエ腸管上皮では、H2O2をO2とH2O に分解する分泌性カタラ
ーゼであるIRC(immune-regulated catalase )が発現している。IRC をノッ クダウンすることでDUOX により産生される ROS の除去が不全になるととも に、菌感染に対する抵抗性が低下する。これは、DUOX が産生する ROS が病 原菌の排除に必須である一方で、過剰なROS は宿主にとって有害であることを 示唆している(59, 60)。したがって、健常時の常在菌の過半数を占める symbiont に対して、DUOX による ROS 産生を活性化させない機構が必要であり、これ は腸内細菌の分泌するuracil の量によって制御されることが知られている(61, 62, 63)。uracil を受容する宿主側の受容体は同定されていないものの、EC が uracil を感知すると、DUOX の発現および ROS 産生活性が上昇する。この時、 DUOX の活性上昇は Hedgehog シグナルを介した小胞体からの Ca2+の放出と、
TOR(target of rapamycin)シグナルによる NADPH 産生の促進によってもた らされる(62, 63)。Symbiont は uracil 分泌量が少ない一方で、pathobiont や 外来の病原菌はuracil を多量に分泌するため、宿主はこの uracil 分泌量の違い を利用して、病原菌に対し選択的にDUOX による ROS 産生を誘導することが できる(Fig.1-8)。Uracil 刺激に応じたDUOX 依存的な ROS 産生誘導は、シ ョウジョウバエのみならずヒトの上皮細胞培養系や線虫個体にも保存されてい る(61)。なお、細菌は、菌密度などの周囲の環境を察知する(quorum-sensing) ことで増殖サイクルや毒性を調節しており、uracil の分泌を quorum-sensing に利用していることが示唆されている(64)。したがって、symbiont は本来必 要な uracil 分泌を抑制することで、宿主の腸管内への定着を可能にしていると 考えられている(61)。 抗菌ペプチドやROS による抗菌応答に加え、ショウジョウバエ腸管上皮と腸
内細菌の間には、囲食膜(Peritrophic matrix)と呼ばれる、ほ乳類腸管の粘液 層に相当する粘膜バリアーが存在する(50)。囲食膜にはペプチドグリカン分解 酵素などの抗菌物質が存在し、物理的防御機能に加えて、免疫応答の場として も機能する(50)。このように,ショウジョウバエの腸管はヒトと同様に、抗菌 ペプチドやROS による化学的防御機構と、囲食膜による物理的防御機能という 複数の防御機構を備えている。 3-3. ショウジョウバエ腸管上皮の損傷修復機構の哺乳類との類似性 哺乳類、ショウジョウバエともに腸管の恒常性を維持する上で、腸管幹細胞 ISC による上皮細胞のターンオーバーが非常に重要な役割を果たす(49, 65)。 腸管は宿主の細胞が常在菌だけでなく外界からの病原体や刺激物とも接触する 場であり、他の組織と比較して腸管上皮細胞が頻繁に損傷を受ける。腸管上皮 の大半を占めるのはEnterocyte(EC)であり、分裂能を持たない EC は自身の 細胞内で損傷を修復できない場合、アポトーシスなどによって腸管上皮から脱 落する(66)。このような損傷刺激に応じて ISC は分裂を活性化し、EC の脱 落によって腸管上皮のバリア機能が損なわれないように上皮細胞を新生する。 哺乳類、ショウジョウバエともに、ISC 分裂の活性は近傍に存在する細胞種 から分泌されるサイトカインによって制御されている。ほ乳類では、基底膜直 下の間質細胞(筋繊維芽細胞、平滑筋細胞、免疫系細胞など)や、ISC と同様 に腸陰窩に存在するパネート細胞が主にISC 分裂を制御している。同様にして、 ショウジョウバエでも基底膜を覆う筋層や、前駆細胞 Enteroblast(EB)から サイトカインが分泌される。また、ショウジョウバエ腸管上皮では EC も ISC と近接しており(Fig.1-7)、EC からも ISC 分裂を促進するサイトカインが分 泌される(Fig.1-9)。 腸管上皮細胞のターンオーバーは病原体感染などの外的な損傷刺激を受けて
いなくとも活発で、ショウジョウバエではおおよそ4 日、マウスではおおよそ 5 日で全ての上皮細胞が入れ替わるとされている(8, 67)。非感染時では、ショ ウジョウバエISC は IL-6 様サイトカインである Unpaired 1(Upd1)をオート クラインすることによって、非損傷時の上皮細胞のターンオーバーを起こして いる(49, 65)。Upd ファミリーは JAK/STAT 経路のリガンドであり、受容体 Domeless に結合することで STAT による遺伝子発現を誘導する。また、筋層か らはWnt 経路のリガンドである Wingless や、EGFR シグナルのリガンドであ るVein が分泌され、ISC 分裂活性を支えている。Wingless が受容体 Frizzled と結合すると、転写共役因子-catenin が活性化することで Wnt 経路のターゲ ット遺伝子発現が誘導される。また、Vein は EGFR に結合することで、 Ras-MAPK カスケードを介して ISC 分裂を活性化させる。これらに加え、非感 染時のターンオーバーには、細胞死に応じてEC から分泌される Spitz や Keren といったEGF が寄与することも報告されている(67)。
これに対し、損傷時のISC 分裂には、主に EC から分泌される Upd3 や、EB から分泌される Upd2 が寄与する。Upd2, Upd3 は直接 ISC に作用して JAK-STAT 経路を活性化させるとともに、筋層にも作用して Vein の産生を誘 導する(Fig.1-8)。これにより、Upd2/3 は腸管上皮が損傷した際にシグナルを 増幅しつつ ISC 分裂を効率的に活性化させることができると考えられている (67)。特に、腸管上皮に占める割合が高い EC は損傷を受けやすく、Upd3 は腸管損傷時の恒常性維持に必須の役割を果たしている。損傷に応じて Upd2 や Upd3 の発現を誘導するためには、損傷刺激を受容し、これらのサイトカイ ン発現を誘導するシグナル経路を活性化する必要がある。これまでに、病原体 感染やDSS の経口摂取時に EC において upd3 発現を誘導するシグナル経路と して、Hippo 経路や JNK 経路が報告されている(68, 69, 70)。しかし、これ らのシグナル経路が損傷に対してどのようなメカニズムで活性化するのかは明
らかにされていない。
こうしたショウジョウバエ腸管の ISC 分裂システムは、哺乳類にも保存され ている。哺乳類腸管の基底膜直下に存在する間質細胞は Wnt(ショウジョウバ エ:Wingless)や IL-6(ショウジョウバエ:Upd1/2/3)を分泌することで ISC 分裂を促進する。また、腸陰窩において ISC と近接しているパネート細胞では Wnt や EGF(ショウジョウバエ:Spitz, Vein, Keren)の高い発現が見られ、 in vivo における ISC の分裂能や、in vitro におけるオルガノイド培養にパネー ト細胞が必須であることが明らかにされている。ショウジョウバエ腸管と同様 Hippo 経路が DSS 摂取時の ISC 分裂制御に寄与しており、Hippo 経路下流の転 写因子 YAP の活性化によって Wnt 経路の活性化が誘導されることが明らかに されている(65)。