3番染色体の逆位により誘導されるEVI1遺伝子と
GATA2遺伝子の発現の異常は巨核球増多を伴う白血
病を発症させる
著者
山岡 彩香
学位授与機関
Tohoku University
学位授与番号
11301甲第19147号
URL
http://hdl.handle.net/10097/00129292
博士論文
3 番染色体の逆位により誘導される
EVI1 遺伝子と GATA2 遺伝子の発現の異常は
巨核球増多を伴う白血病を発症させる
東北大学大学院医学系研究科医科学専攻
生体機能学講座 医化学分野
山岡 彩香
目次 1.要旨 ・・・・・・・・・・・・・・・・・・・・・・・・ 3 2.研究背景 ・・・・・・・・・・・・・・・・・・・・・・・・ 6 3.研究目的 ・・・・・・・・・・・・・・・・・・・・・・・・ 9 4.研究方法 ・・・・・・・・・・・・・・・・・・・・・・・・ 10 5.研究結果 ・・・・・・・・・・・・・・・・・・・・・・・・ 22 6.考察 ・・・・・・・・・・・・・・・・・・・・・・・・ 39 7.結論 ・・・・・・・・・・・・・・・・・・・・・・・・ 49 8.参考文献 ・・・・・・・・・・・・・・・・・・・・・・・・ 50 9.表 ・・・・・・・・・・・・・・・・・・・・・・・・ 54 10.謝辞 ・・・・・・・・・・・・・・・・・・・・・・・・ 56 11. 図
1.要旨 3 番染色体の長腕 q21(3q21)と q26(3q26)との間の逆位もしくは転座を伴う急性骨髄 性白血病(AML)は、予後不良の白血病である。この白血病は、血小板数が正常から 高値を示すこと、骨髄中に異形成を伴う巨核球が増殖していることが特徴的である。 3 番 染 色 体 の 逆 位 も し く は 転 座 が 生 じ る こ と で GATA2-distal hematopoietic enhancer(G2DHE)は EVI1 遺伝子に近接し、その発現を活性化することが白血病の原
因となっている。一方、G2DHE は GATA2 遺伝子から離れるために GATA2 遺伝子の
ヘテロ欠失が起こる。このように、白血病を引き起こす遺伝子異常は解明されてい るが、EVI1 遺伝子と GATA2 遺伝子の発現異常が白血病の発症にどのように関与し ているのかは依然として明らかにされていない。本研究室では、G2DHE による EVI1 遺伝子高発現を再現したトランスジェニックマウス(3q21q26-EVI1 マウス)、GATA2 遺伝子のヘテロ欠失を再現した Gata2+/gfp マウス、さらにその両者を再現した 3q21q26-EVI1::Gata2+/gfp マウスが樹立されている。本研究では、白血病の発症メカ ニズムを解明するために、上記のマウスを元にEVI1 遺伝子と Gata2 遺伝子の発現 をsingle cell で同時にモニターできるマウスを樹立して解析を行った。白血病発症
前の12 週齢マウスの骨髄を解析したところ、EVI1 遺伝子と Gata2 遺伝子の両方を 高く発現している細胞群(High population)が存在していた。この High population は、
EVI1 遺伝子と Gata2 遺伝子の両方の発現が高く誘導されている細胞群であり、連続
的なコロニー形成能を有していた。また、造血幹細胞・前駆細胞(HSPC)が濃縮され ていることから、High population が白血病の起源であると考えられた。前述のよう
に、3q21q26 白血病は骨髄中に異形成を伴う巨核球が増殖しているのであるが、High
population には HSPC の他に巨核球も濃縮されていた。このように、High population
に HSPC と巨核球が濃縮されていたことから、白血病発症前の 12 週齢マウスにお いてEVI1 遺伝子と Gata2 遺伝子の発現異常が HSPC と巨核球にどのような影響を 与えているのかを解析した。その結果、EVI1 遺伝子の高発現により、HSC の分化 は骨髄球系前駆細胞と巨核球系前駆細胞に傾いており、一方でリンパ球系前駆細胞 への分化は減少していた。さらに、EVI1 遺伝子の高発現に加えて Gata2 遺伝子の発 現低下が加わることで、巨核球系前駆細胞への分化により傾いていた。最後に、白 血病発症後のマウスの解析を行った。すると、3 番染色体逆位による EVI1 遺伝子高 発現とGata2 遺伝子のハプロ不全の両方を再現したときに、血小板と巨核球増多を
伴う白血病が再現された。以上より、3 番染色体逆位による EVI1 遺伝子高発現と
Gata2 遺伝子のハプロ不全が同時に存在することが、巨核球系への分化を促進し、
2.研究背景 3 番染色体の長腕 q21(3q21)と q26(3q26)との間の逆位もしくは転座は、骨髄異形症 候群(MDS)や急性骨髄性白血病(AML)の原因となる。そして、3q 変異を伴う AML はAML の 1-2%にみられ、予後不良であることが知られている 1-6。3q 異常を伴う 白血病では、2 つの遺伝子すなわち EVI1(MECOM)遺伝子と GATA2 遺伝子に発現の 異常が生じている。3 番染色体に逆位もしくは転座が生じると、3q21 に存在してい
るGATA2 遺伝子エンハンサー(GATA2-distal hematopoietic enhancer : G2DHE)が 3q26
のEVI1 遺伝子の近傍に位置するようになる7-9。3q 異常が生じた AML 細胞では、
EVI1 遺伝子の発現は G2DHE により誘導され上昇する一方で、GATA2 遺伝子の発現
はG2DHE を失うために発現が減少する。しかしながら、これらの遺伝子発現の異 常がどのように白血病発症に関わっているのかは不明である。 3q AML の骨髄は形態学的に多様で、骨髄単球性白血病や巨核芽球性白血病の像に 似ている 10, 11。さらに、異形成の巨核球が観察されることが多く、7-22%の患者は 血小板増加を伴う 11,12。末梢血には、巨大で顆粒の少ない血小板と裸核巨核球が見 られる13。このように、3q 変異を伴う AML と MDS は、巨核球の異常を伴ってい
る。これまでに、白血病発症のメカニズムを解明しようと、いくつかの EVI1 遺伝 子を過剰発現させたマウスモデルが樹立されてきた14-17。しかしながら、これらの マウスのうち、ヒト3q AML の特徴である巨核球と血小板の増多を呈しているもの は存在していなかった。それゆえにEVI1 遺伝子と Gata2 遺伝子の発現の異常が、 どのように巨核球増多や予後不良に関係しているのかを明らかにするには、別のマ ウスモデルが必要であった。当研究室では以前に、2 つの BAC クローンを組み合わ せることによりEVI1 遺伝子と G2DHE を含む 196kb のヒト 3q21q26 逆位アリルを 遺伝子導入した 3q21q26-EVI1 マウスを作成している 9。このマウスでは、G2DHE により誘導されるヒト EVI1 遺伝子の発現は、造血幹細胞・前駆細胞において高く なっており、最終的に白血病を発症した。骨髄細胞の解析により、3q21q26-EVI1 マ ウスでは B220+c-Kit+Gr1-芽球様細胞が白血病発症能を有しており、この芽球様細 胞はGr1+骨髄球に分化することができていた(分化型白血病)。しかしながら、この 3q21q26-EVI1 マウスでは、マウス内因性の Gata2 遺伝子に変化はない。そのため、
Gata2 遺伝子のハプロ不全を再現するために、3q21q26-EVI1 マウスと Gata2+/gfpマウ
遺伝子のハプロ不全が加わることで、白血病は悪性化することが明らかになった18。 すなわち3q21q26-EVI1::Gata2+/gfp白血病マウスの骨髄では、B220+c-Kit+Gr1-芽球様 細胞はGr1+骨髄球に分化することが出来なくなり、B220+芽球様細胞が骨髄内で増 殖していた(未分化型白血病)。また、3q21q26-EVI1::Gata2+/gfp白血病マウスのもうひ とつの特徴として、約30%は分化型でも未分化型でもない白血病を呈することが分 かったが、この参考論文では解析はされていなかった。
3.研究目的 一般的に白血病は、造血細胞の分化に異常が生じてその段階の細胞が蓄積すること、 造血細胞の増殖が異常に促進すること、アポトーシスに異常が生じ、本来アポトー シスを起こすべき異常な細胞が死滅しないことにより発症する 19。3q21q26 白血病 では、白血病を引き起こす原因遺伝子は明らかにされたが、発症メカニズムは依然 として不明である。本研究では、造血細胞におけるEVI1 遺伝子と Gata2 遺伝子の 発現の異常が引き起こす3q21q26-EVI1::Gata2+/gfpマウスの白血病発症メカニズムを 明らかにすること、さらに巨核球と血小板の増多を伴う3q21q26-EVI1::Gata2+/gfp白 血病の特徴を調べることを目的とした。
4.研究方法 動物実験 3q21q26 マウス(line B)9とGata2+/gfpマウス20は、以前に報告されたものを使用した。 全てのマウスの取り扱いは、東北大学における動物実験等に関する規定に従った。 マウスの飼育は、室温 20±3℃、12 時間明暗周期(午前8時から午後8時を明期と した)、および特定病原体除去環境下にて行い、水および餌は自由摂取とした。ま た実験に際しては、できる限りマウスに苦痛を与えないよう配慮し、著しい体重減 少や移動障害といった病的兆候を示していないことを確認し、安楽死にはイソフル ランによる麻酔の上、頚椎脱臼を用いた。白血病発症の長期観察に際しては、定期 的に採血を行い末梢血白血球数の増加傾向(>5×104/μl)が認められた時点で解析を 行うようにした。 ゲノム DNA の調整および PCR 法による遺伝子型の決定 マウスの尻尾を50mM NaOH で 1 時間溶解後、1M HCl で中和し、15000rpm、1 分 間遠心した上清をPCR 反応に用いた。PCR 反応条件は 94℃20 秒、60℃20 秒、72℃
20 秒のサイクルを 40 サイクルで行った。プライマーは、human EVI1 遺伝子につい ては 5’-TCTTTCCTGATTCCATCCACCT-3’と 5’- AGTGGCCAGATGTCAAACCA-3’ を用いた。3q21q26-tdTomato 遺伝子については 5’- CCTGTACGGCATGGACGAGC-3’ と 5’-AGTGGCCAGATGTCAAACCA-3’ を 用 い た 。 GFP 遺 伝 子 に つ い て は 5’-CCTGGTTCCCAAGACACAGTAG-3’と 5’-GGGTGCTGCGCATTCAATAC-3’を用 いた。 3q21q26-tdTomato ターゲット断片の作成 tdTomato 遺伝子と Frt シークエンスを隣接させたアンピシリン(Amp)耐性遺伝子 (FrtAmp)を含む断片を作成した。tdTomto-polyA/pBS-SK プラスミドを作成するため に、tdTomto 遺伝子と polyA を BamHI と EcoRV の間に挿入した(図 1A)。FrtAmp フ ラグメントは、pFrtAmp プラスミドを用いた PCR により作成した( TomatoFrtAmp(f) と TomatoFrtAmp(r)のプライマーセットは表 3 に示す)。FrtAmp フラグメントは、 In-Fusion HD ク ロ ー ニ ン グ キ ッ ト (Clontech) を 用 い た 相 同 組 み 換 え に よ り tdTomto-polyA/pBS-SK プ ラ ス ミ ド の Sal Ⅰ サ イ ト に 挿 入 し 、
tdTomto-polyA-FrtAmp/pBS プラスミドを作成した。3q21q26-tdTomato ターゲットフ ラグメントは、tdTomto-polyA-FrtAmp/pBS プラスミドを用いた PCR により作成し た(3q21q26tdTomto(f)と 3q21q26tdTomto(r)のプライマーセットは表 3 に示す) 。PCR にはPrimeSTAR Max DNA ポリメラーゼ(Takara)を用い、PCR 産物は Gel Extraction Kit(QIAGEN)で純化した。
3q21q26-tdTomato BAC の作成
3q21q26-tdTomato BAC を 作 成 す る た め に 、 3q21q26-EVI1 BAC9 を 改 編 し た 。
3q21q26-EVI1 BAC 中の EVI1 遺伝子の翻訳開始点に tdTomato 遺伝子を挿入するため に相同組み換えを行った 21,22。Flp 組み換え誘導のために E.coli.の EL250 系を用い
た21。この系は、採取するに先立ち42℃の振とうウォーターバス内において追加で
15 分間誘導した。BAC DNA と目的とする構築は、Gene Pulser(BIO-RAD)を用いて 1.8kV, 25uF, 200W の電気穿孔(electroporation)で形質導入した。抗生剤により選別す
るために、クロラルフェニコール25ug/mL とアンピシリン 100ug/mL を LB 培地に
リコンビナーゼを誘導した。BAC DNA は、Nucleo-bond BAC DNA preparation kit (Macherey-Nagel)を用いて純化した。純化した BAC DNA は電気穿孔による Flp リコ ンビナーゼをすることはせず、DH10B 系に形質導入した。
3q21q26-tdTomato BAC の評価
相同組み換えをサザンブロット解析で確かめた(図 1C)。BAC DNA を BamHI もしく は EcoRI の制限酵素で切断し、電気泳動しナイロン膜に転写した。ナイロン膜は、
ジ ゴ キ シ ゲ ニ ン(DIG)でラベルされた DNA プローブ(DIG DNA Labeling and Detection Kit(Roche)を用いた)で反応させ、その後、AP で標識した抗 DIG 抗体で反 応 さ せ 、NBT-BCIP 基 質 を 用 い て 可 視 化 し た 。 3q21q26 BAC の 鋳 型 と し た 5’-TTCCAGGCAGGATTCAACTC-3’と 5’-TGGTGACAAATGGAAAGCTG-3’のプラ
イマーセットによるPCR 産物をプローブとして利用した(図 1B オレンジの四角)。
Flp による組み換えは PCR により確認した。
BAC DNA を PI-SceⅠの分解により直線化し、フェノール-クロロホルム抽出とエタ ノール沈殿により純化した。直線化した BAC 構築を BDF1 受精卵に注入した。こ のトランスジェニックマウスをC57BL/6 マウスと 8 世代掛け合わせ、純系化した。 ジ ェ ノ タ イ ピ ン グ は 、 5’-CCTGTACGGCATGGACGAGC-3’ と 5’-AGTGGCCAGATGTCAAACCA-3’のプライマーによる PCR で決定した。トラン ス ジ ー ン の コ ピ ー 数 は ヒ ト と マ ウ ス の EVI1 遺 伝 子 第 7 エ ク ソ ン の 5’-ACAAGCCAAGACCAGCCCCTGG-3’ と 5’-TGGTTTTTTCGAGGCTCAGTC-3’ の プラーマーによるqPCR で決定した(図 1D)。 血液学的解析 頬採血によって50-200μl の末梢血を採取し、全自動血球計算機 MEK-6258(日本光 電)を用いて血球数算定を行った。末梢血像は塗抹標本をライトギムザ染色で染色 し形態を観察した。骨髄細胞はFACS AriaⅡにより細胞を表面マーカーで識別して 採取したものをサイトスピンによりスライド標本として、ライトギムザ染色で染色 し形態を観察した。
コロニーアッセイ
FACS AriaⅡで表面マーカーにより識別した細胞 1000 個を 1000μl の IMDM 培地 (Iscove’s MDM with 25mM Hepes) へ採取した。そのうち 250μl を 2.5ml のメソカル トGF M3434(Stem Cell Technologies)へ添加し混合した後、シリンジと 18G 針を用い
て1ml ずつ 3cm ディッシュ(2枚)へまいた。メソカルト GF M3434 には、あらかじ
めStem cell factor, IL-6, IL-3, Erythropoietin, Transferrin を加えた。1週間後にコロニ ーカウントを行い、コロニー形成能を評価した。さらに、コロニー形成がみられた ものは1.7ml の IMDM 培地を加えてピペッティングを行い回収する操作を3回繰り 返して回収し、細胞数をカウントした。2500 個の細胞を 2.5ml のメソカルト GF M3434 へ添加し、初代培養と同様の操作で継代を行い、1週間後にコロニーカウン トを行い、リプレーティング能を評価した。 フローサイトメトリー解析 骨髄あるいは末梢血から単一細胞懸濁液を調整し、Histopaque-1083(Sigma)により脱 核赤血球を除いた後、非特異的染色を防ぐため精製FcRⅡ/Ⅲ抗体と反応させ、その
後に各抗体(1:200)による染色を行った。PI(propidium iodide)により死細胞の染色を 行った後、FACS AriaⅡ、FACS Verse を用いて FACS 解析を行った。抗体のリスト は表1 に示した。データ処理は FlowJo ソフトウェア(Tree Star)で行った。
使用した抗体の組み合わせは以下に示す。 コロニー染色 図5. 巨核球 図6. 好中球と単球 B220 PE-Cy7 Gr1 Bv510 c-Kit APCeFluor CD41 APC CD61 BV421 CD41 APC CD61 BV421 c-Kit APCeFluor B220 APCCy7 CD3 APCCy7 NK1.1 PECy7 CD115 APC Ly6G BV510
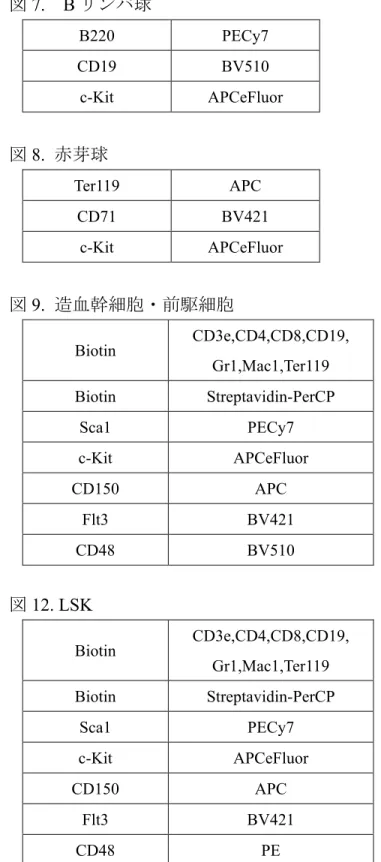
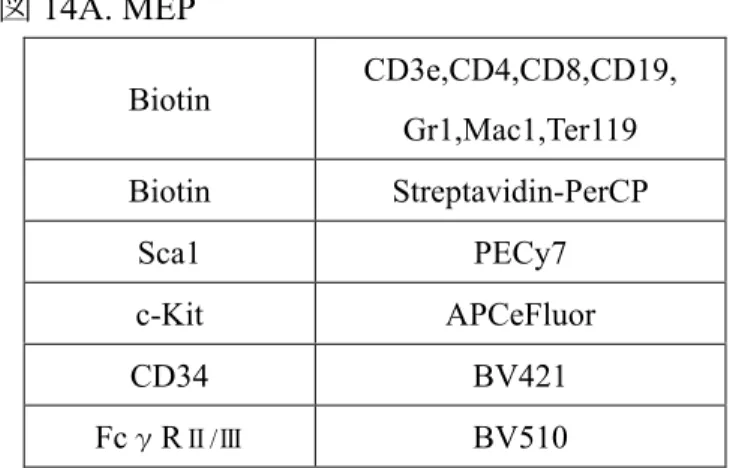
図7. B リンパ球 図8. 赤芽球 図9. 造血幹細胞・前駆細胞 図12. LSK B220 PECy7 CD19 BV510 c-Kit APCeFluor Ter119 APC CD71 BV421 c-Kit APCeFluor Biotin CD3e,CD4,CD8,CD19, Gr1,Mac1,Ter119 Biotin Streptavidin-PerCP Sca1 PECy7 c-Kit APCeFluor CD150 APC Flt3 BV421 CD48 BV510 Biotin CD3e,CD4,CD8,CD19, Gr1,Mac1,Ter119 Biotin Streptavidin-PerCP Sca1 PECy7 c-Kit APCeFluor CD150 APC Flt3 BV421 CD48 PE
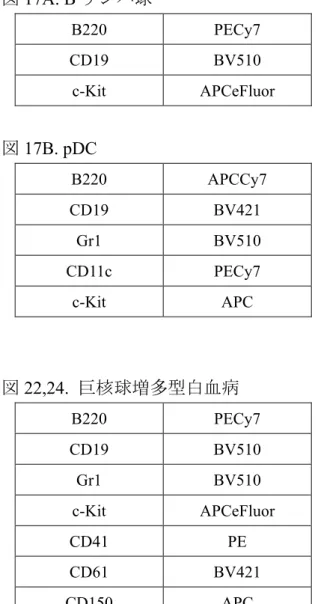
図14A. MEP 図14B. 巨核球 図15. 赤芽球 図16. 好中球と単球 Biotin CD3e,CD4,CD8,CD19, Gr1,Mac1,Ter119 Biotin Streptavidin-PerCP Sca1 PECy7 c-Kit APCeFluor CD34 BV421 FcγRⅡ/Ⅲ BV510 CD41 APC CD61 BV421 c-Kit APCeFluor Ter119 APC CD71 BV421 c-Kit APCeFluor B220 APCCy7 CD3 APCCy7 NK1.1 PECy7 CD115 APC Ly6G BV510
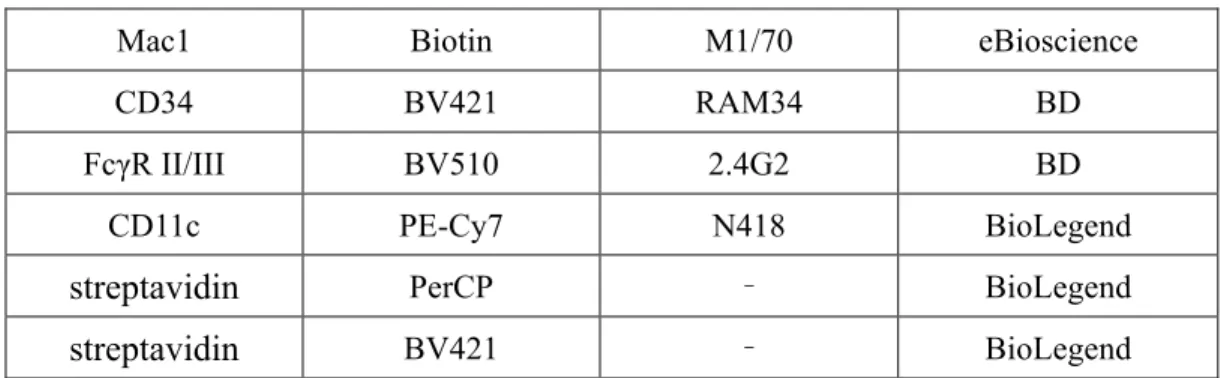
図17A. B リンパ球 図17B. pDC 図22,24. 巨核球増多型白血病 骨髄の細胞数と割合 マウスの両大腿骨と両頸骨から、3ml の PBS を入れたシリンジと 23G 針を用いて 骨髄を流出させ、ほぐした。骨髄細胞を含む3ml の PBS を、15ml チューブに入れ B220 PECy7 CD19 BV510 c-Kit APCeFluor B220 APCCy7 CD19 BV421 Gr1 BV510 CD11c PECy7 c-Kit APC B220 PECy7 CD19 BV510 Gr1 BV510 c-Kit APCeFluor CD41 PE CD61 BV421 CD150 APC
た3ml の HISTOPAQUE-1083 の上に乗せ、1500rpm で 30 分間遠心した。中間に浮
遊する分離された白血球の層を1000ul のピペットマンで 3 回取り出し(計 3ml)、6ml
のPBS を入れた 15ml チューブに入れ、1500rpm で 5 分間遠心した。上清を除いて
からPBS で 500ul に合わせ、この細胞液を 10ul とトリパンブルー10ul を混合して
から CountessⅡ(invitrogen)で生存細胞数をカウントした。FACS の展開で得られた
(図12B)各分画の割合(%)を掛けて、細胞数を計算した。
定量的 RT-PCR
FACS AriaⅡで表面マーカーにより識別した細胞 10,000 個を 1ml セパゾール RNA Ⅰ Super G(ナカライテスク)へ採取し、全 RNA を精製した。cDNA 合成は ReverTra Ace qPCR RT Master Mix with gDNA Remover(TOYOBO)を用いて行った。定量的リア ルタイム PCR は Quanto Studio 6 Flex Real time system を用いて行い、試薬は THUNDERBIRD Probe qPCR Mix(TOYOBO)及び KAPA SYBR FAST qPCR Kit Master Mix Universal(KAPA BIOSYSTEMS)を用いた。プライマーの配列は表 2 に示した。 ヒトEVI1 遺伝子のプライマー、プローブは Life Technologies 社の Taqman® Gene
Expression Assay Hs01115407_m1 を用いた。
統計解析
ドットブロット、棒グラフは平均値±標準偏差を示した。群間の比較にはt 検定を
5.研究結果 (1) 3q 変異により誘導される EVI1 遺伝子の発現をモニターできるマウスを樹 立した EVI1 遺伝子は正常の状態では、造血幹細胞において高く発現しており、細胞の維持 や分化の決定に働いている。このことから、G2DHE により EVI1 遺伝子が異常に高 く発現している細胞に分化と増殖の異常が生じ、3q 白血病の起源となっていると考 えた。そこで、G2DHE による EVI1 遺伝子の発現をモニターするために、3q21q26-
EVI1 BAC アリルの EVI1 遺伝子のエクソン 3 翻訳開始点に tdTomato 遺伝子を挿入
した3q21q26-tdTomato BAC を作成した(図 1A)。サザンブロットにより相同組換を 確認した(図 1C)。BAC を受精卵に注入し 3 ラインを得た(line A, B, C)。形質導入さ
れたコピー数をqPCR で確認したところ、2-5 コピーであった(図 1D)。以下の実験
では、3 コピーの導入遺伝子を持つ line A のマウスを用いた。3q21q26-tdTomato マ
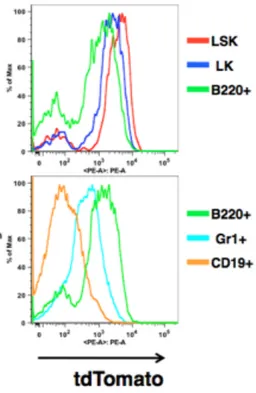
ウスにおいて tdTomato 蛍光の発現を解析したところ、LSK 分画で最も高く次いで
LK 分画で高く、B220+細胞でやや高い一方で Gr1+細胞や CD19+細胞で発現が低か った(図 2)。これは 3q21q26 マウス(line B)において BAC 由来 human EVI1 の発現を
qPCR で測定した結果9と一致しており、G2DHE による EVI1 遺伝子の発現をモニ
ターできていると考えられた。
(2) 3q21q26-tdTomato レポーターと Gata2-GFP レポーターを持つマウスの解析 により、EVI1 遺伝子と Gata2 遺伝子の発現が共に高値である High 細胞群の
存在を明らかにした
骨髄細胞において、G2DHE による EVI1 遺伝子の発現と Gata2 遺伝子の発現を同時
にモニターするために、3q21q26-tdTomato とヘテロ Gata2-GFP ノックインノックア ウトマウス(Gata2+/gfpマウス)をかけ合わせて、Gata2+/gfp::3q21q26-tdTomato マウスを
得た(以下、Gata2+/gfp::Tom マウスとする)。Gata2+/gfpマウスは、内因性 Gata2 遺伝
子の第 2 エクソンに GFP 遺伝子が挿入されているので、Gata2 遺伝子の代わりに
GFP が発現する20。EVI1 遺伝子の高発現による影響も調べるため、Gata2+/gfp:: Tom
マウスと3q21q26- EVI1 マウスをかけ合わせて 3q21q26-EVI1::Gata2+/gfp::Tom マウス
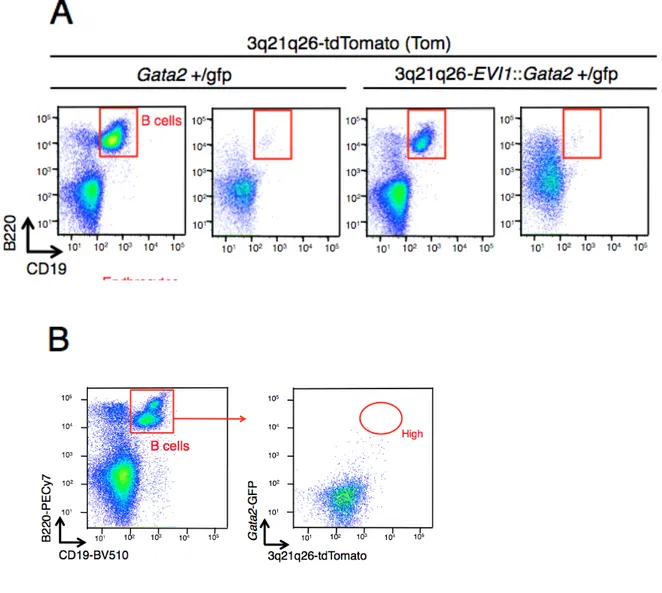
を得た。3q 白血病の発症メカニズムを明らかにするために、白血病発症前である 12 週齢の Gata2+/gfp::Tom マウスと 3q21q26-EVI1::Gata2+/gfp::Tom マウスの骨髄細胞
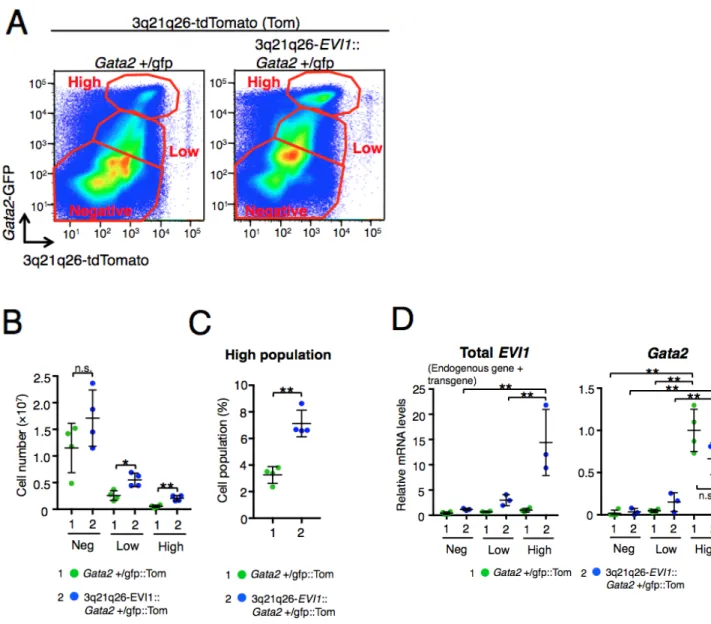
を解析した。tdTomato と GFP の発現には相関が見られ、G2DHE による EVI1 遺伝
子の発現はGata2 遺伝子の発現と相関することが示唆され、さらに tdTomato と GFP
の両方を高く発現している細胞集団が存在していることが分かった(図 3A)。このこ とから、tdTomato と GFP の発現の高さで High, Low, Negative 群で分割した。それぞ れの細胞群でEVI1 遺伝子 mRNA と Gata2 遺伝子 mRNA の qPCR を行った。EVI1
遺伝子は3q21q26-EVI1 由来の human EVI1 遺伝子とマウス内因性 Evi1 遺伝子の合 計であるが、3q21q26-EVI1::Gata2+/gfp::Tom マウスの High 細胞群では tdTomato 蛍光
が高値であると共にEVI1 遺伝子の発現も高くなっていることを確認した(図 3D 左)。
また、Gata2+/gfp::Tom マウスと 3q21q26-EVI1::Gata2+/gfp::Tom マウスにおいて High
細胞群では GFP 蛍光が高値であると共に Gata2 遺伝子の発現も高くなっているこ
と を 確 認 し た ( 図 3D 右 ) 。 Gata2+/gfp::Tom マ ウ ス に 比 べ て 3q21q26-EVI1::Gata2+/gfp::Tom マウスでは、High, Low 群で骨髄細胞数が増加してお
り(図 3B)、さらに骨髄に占める High 細胞群の割合が増加していた(図 3C)。以上よ り、EVI1 遺伝子の高発現は High 細胞群を蓄積させることが示された。
(3) 3q21q26-EVI1::Gata2+/gfp::Tom マウスの High 細胞群は連続的コロニー形成能 を有する
それぞれの細胞群の複製能を調べるため、High, Low, Negative 細胞群をソーティン
グしてコロニーアッセイの実験を行った。培地はSCF, IL-3, IL-6 を含み、7 日毎に
コロニー数をカウントし、コロニーが生えていた場合はリプレーティングした。1 週後では、Gata2+/gfp::Tom マウスと 3q21q26-EVI1::Gata2+/gfp::Tom マウスの High 細
胞群でコロニー形成が見られた(図 4A. 1st の緑の棒グラフ(Gata2+/gfp::Tom)と青の棒
グラフ(3q21q26-EVI1::Gata2+/gfp::Tom))。これらの細胞群に連続的な複製能があるの か を 確 か め る た め に リ プ レ ー テ ィ ン グ を 行 っ た と こ ろ 、
3q21q26-EVI1::Gata2+/gfp::Tom マウスの High 細胞群は少なくとも 4 週後まで複製能
を有していることが示された(図 4A, 青い棒グラフの 2nd, 3rd, 4th)。一方で、
Gata2+/gfp:: Tom マウスでは 2 週後の時点でコロニーはほとんど形成されなくなって
いた(図 4A, 緑の棒グラフの 2nd)。これらの結果から、EVI1-/Gata2- High 細胞群は
コロニー形成能があり、さらにEVI1 遺伝子が高発現を伴うことで、High 細胞群は
過程でHigh 細胞群の tdTomato(EVI1)と GFP(Gata2)の発現はどうなっているのかを
確かめるために、コロニーを回収しフローサイトメトリー解析した(図4B)。これ
を数値化したところ、3q21q26-EVI1::Gata2+/gfp::Tom マウスの High 細胞群は、1 週後
にEVI1-/Gata2- High の割合が約 50%と低下した(図 4C. 青のドット Passage number
1)が、2 週後から 4 週後にかけては約 70%で維持されていた(図 4C. 青のドット Passage number 2,3,4)。一方で、Gata2+/gfp:: Tom マウスの High 細胞群コロニーは 1
週後のEVI1-/Gata2- High の割合が約 30%に低下していた(図 4C. 緑のドット Passage
number 1)。コロニーをリプレーティングする際に FACS 解析した像を図 4D-G に示 す。Gata2+/gfp::Tom マウス、3q21q26-EVI1::Gata2+/gfp::Tom マウス共に、tdTomato(EVI1)
とGFP(Gata2)の発現が低下した細胞は Gr1+細胞であり、High 細胞群に含まれる造 血幹細胞・前駆細胞(図 9A)が Gr1+細胞に分化したと考えられた。リプレートする 際は、すべてのコロニーを回収して細胞数をカウントし、1 ディッシュ当たり 1000 個をまいた。図4E, 1 段目で見られていた Low/Negative 分画が、1 週間後の図 4F,1 段目には見られなくなっていたが、これは Gr1+細胞に分化した細胞は連続的なコ ロニー形成能を失うためと考えた。High 分画中の CD41+CD61+巨核球は 4 週に渡
って維持されており、EVI1 遺伝子の高発現と Gata2 遺伝子の発現が巨核球を維持し
たと考えられた。図4E, 4F において、High 分画でも c-Kit の発現がないのは、培地
にstem cell factor が含まれており、その受容体である c-Kit がダウンレギュレーショ ンしたためと考えた。
以上から、3q21q26-EVI1::Gata2+/gfp::Tom マウスの High 細胞群は、G2DHE による
EVI1 の発現と Gata2 の発現を高く保った状態で連続的にコロニーを形成できるこ とが示され、このことから細胞集団が白血病の起源となっていると考えられた。 (4) EVI1-/Gata2- High 細胞群は造血幹細胞・前駆細胞(HSPC)と巨核球を含む High 細胞群は連続的コロニー形成能を有することが示されたことから、どのような 細胞が High 細胞群に含まれているのかを調べるために、造血細胞の表面マーカー で染色してフローサイトメトリーを行い、それぞれの血球が High 細胞群と
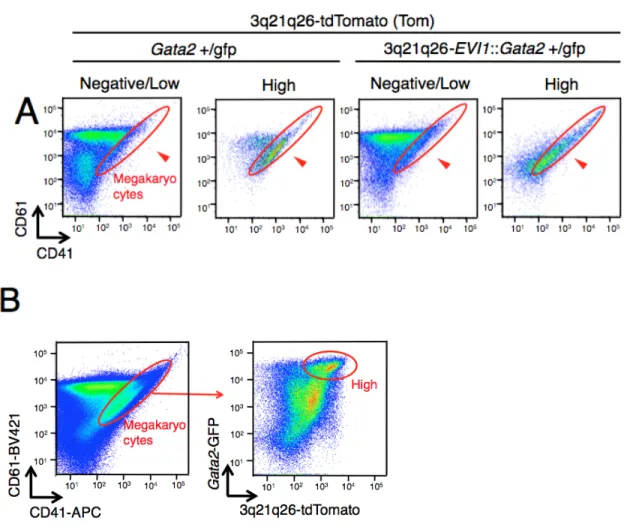
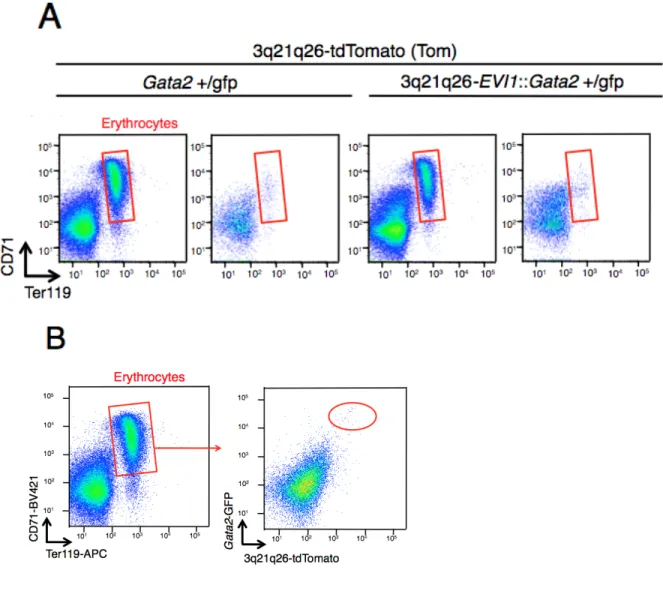
Negative/Low 細胞群のどちらに含まれるかを解析した。すると、Gata2+/gfp::Tom マ
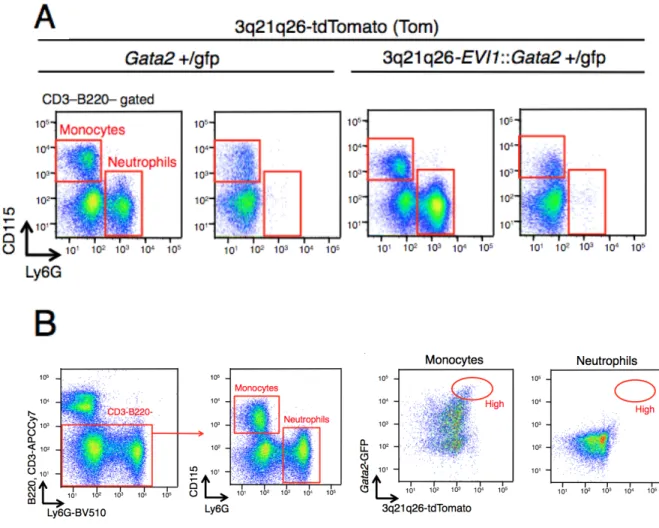
ウ ス 、3q21q26-EVI1::Gata2+/gfp::Tom マ ウ ス 共 に 、 High 細 胞 群 に は 巨 核 球 (CD41+CD61+)が濃縮されていた(図 5A)。一方で、好中球(CD3-B220-Ly6G+)、B リ
ンパ球(B220+CD19+)、赤芽球(CD71+Ter119+)は、大部分が Negative/Low 細胞群に 存 在 し て お り( そ れ ぞ れ 図 6A, 7A, 8A) 、 単 球 (CD3-B220-CD115+) も 多 く は Negative/Low 細胞群に存在していた(図 6A)。HSPC がどこに含まれているかを見る ためにc-Kit と Sca1 で染色したところ、LSK,LK は High 細胞群に含まれていた(図 9A)。また、3q21q26-EVI1::Gata2+/gfp::Tom マウスの LSK は Gata2+/gfp::Tom マウスに 比べて、よりHigh 細胞群に濃縮されていた(図 10B)。また、CD41+CD61+細胞の c-Kit の発現を見たところc-Kit(-)細胞を含んでいることから、幹細胞ではなく巨核球と考 えられた(図 10C)。これらの結果から、EVI1-/Gata2- High 細胞群は、HSPC と巨核 球を豊富に含んでいることが示された(図 10D)。 (5) G2DHE により誘導される EVI1 遺伝子の高発現により、MPP2(巨核球系・ 赤芽球系への分化傾向を示す前駆細胞)と MPP3(骨髄球への分化傾向を示 す前駆細胞)が増殖する これまでのところで、モニターマウスを用いて造血細胞における EVI1 遺伝子と
の影響が単独で、または協調して白血病発症に及ぼしている影響を調べるために、
12 週齢の 4 種類のジェノタイプマウス:WT, Gata2+/gfp , 3q21q26-EVI1, 3q21q26-EVI1::
Gata2+/gfpを用いて骨髄細胞の解析を行った(図 11)。LSK は大部分が High 細胞群に
含まれていた(図 9A,10B)ことから、LSK を解析することとした。マウス骨髄細胞を Biotin - Gr1, Mac1, CD3e, CD4, CD8, CD19, Ter119 で染色し、解析するときには成熟
骨髄球、成熟リンパ球、成熟赤芽球を除いてから展開した(図12A)。3q21q26-EVI1 マウスと3q21q26-EVI1::Gata2+/gfpマウスにおいては、WT マウスと Gata2+/gfpマウス と比べて、LSK の細胞数が著明に増加しており(図 12B 上段、図 12C )、このこと から、G2DHE による EVI1 遺伝子の高発現は HSPC を増加させることが示された。 次に、LSK 内を HSC と MPP に分けて(図 12B 中段、下段)、それぞれの分画の細胞 がLSK に占める割合(図 13A)と細胞数(図 13B)について解析した。WT と Gata2+/gfp 間においては、LSK に占める割合も細胞数も同程度であった(図 13A,B)。ひとつ例 外があり、Gata2+/gfpにおいてはWT より LT-HSC の割合が減少していたが、これは 以前に報告されていたことと同様であった23。LT-HSC の細胞数は WT と Gata2+/gfp で同程度であることから、Gata2 遺伝子のハプロ不全のみでは HSC と MPP の蓄積
に大きく影響しないことが示された。3q21q26-EVI1 マウスは WT マウスと比べて、 LT-HSC と ST-HSC の LSK に占める割合は減少し(図 13A )、細胞数は同程度である(図 13B)ことから、G2DHE による EVI1 遺伝子の高発現は、造血幹細胞の増殖は引き起 こしていないことが示唆された。また、巨核球に分化傾向のあるMPP2 と骨髄球に 分化傾向のあるMPP3 は LSK に占める割合、細胞数ともに著明に増加していた(図 13A, 13B)。一方で、リンパ球に分化傾向のある MPP4 に関しては、細胞数は同程度 であるものの、割合は著しく減少していた(図 13A, 13B)。これらの結果から、G2DHE により誘導されるEVI1 遺伝子の高発現が生じると、MPP2 と MPP3 の細胞が増殖し、 MPP4 は減少することが示された。3q21q26-EVI1 マウスと 3q21q26-EVI1::Gata2+/gfp マウスを比較した場合、後者でMPP2 の割合がわずかながら増加している(図 13A) 他に、差を認めなかった。 (6) G2DHE による EVI1 遺伝子の高発現は、巨核球と血小板の産生を促進させ る High 細胞群は HSPC に加えて巨核球が含まれていることから(図 10C)、次に EVI1
遺伝子とGata2 遺伝子の発現の異常が巨核球分化に影響を与えているのかを調べた。
3q21q26-EVI1 マウスと 3q21q26-EVI1::Gata2+/gfpマウスでは、WT マウス、Gata2+/gfp マウスと比べて、骨髄中の巨核球・赤芽球前駆細胞(MEP)と巨核球(CD41+CD61+) の細胞数が増加していた(それぞれ図 14A 下段と 14C、14B 上段と 14D)。また、巨
核球におけるc-Kit+の未熟な巨核球の割合が増加していた(図 14B 下段と 14E)。さ
らに、3q21q26-EVI1 マウスと 3q21q26-EVI1:: Gata2+/gfpマウスでは、末梢血の血小板
数の増加がみられたものの、この2 ジェノタイプ間での差は見られなかった(図 14F)。 これらの結果から、3q 変異による EVI1 遺伝子の高発現は、MEP が増加をもたらし、 巨核球と血小板の産生を促進することが示唆された。以上より、3q21q26-EVI1 マウ スと3q21q26-EVI1:: Gata2+/gfpマウスでは、ヒト3q 白血病に特徴的な巨核球と血小 板の増加というフェノタイプ11,12を再現できていると考えられた。 (7) G2DHE による EVI1 遺伝子の高発現は、リンパ球への分化を抑制する 次に、EVI1 遺伝子と Gata2 遺伝子のハプロ不全が他の造血細胞の分化に影響を及ぼ しているのかを調べた。骨髄中の赤芽球(CD71+Ter119+)数はすべてのジェノタイプ
で同程度であり(図 15A と 15B)、末梢血の赤血球数は 3q21q26-EVI1 マウスと 3q21q26-EVI1:: Gata2+/gfpマウスでやや減少していた(図 15C)。好中球(Ly6G+)数と単 球(Ly6G-CD115+)数はすべてのジェノタイプで同程度であった(それぞれ図 16A 中 段と16B、図 16A 下段と 16C)。B リンパ球(B220+CD19+)の細胞数は、WT マウス、
Gata2+/gfpマウスと比べて3q21q26-EVI1 マウスと 3q21q26-EVI1::Gata2+/gfpマウスで著 しく減少していた(図 17A と 17C)。B220+CD19-分画には B リンパ球に分化傾向の あ る 細 胞 の 他 に 形 質 細 胞 様 樹 状 細 胞 (pDC) も 含 ま れ る の で 24 、
pDC(B220+CD19-CD11c+Gr1+) 分 画 の 細 胞 数 の 変 化 も 調 べ た 。 pDC 細 胞 数 は 3q21q26-EVI1 マウスと 3q21q26-EVI1:: Gata2+/gfpマウスで減少していた(図 17B 下段
と17D)。胸腺細胞数も 3q21q26-EVI1 マウスと 3q21q26-EVI1:: Gata2+/gfpマウスで減
少しており(図 17F)、EVI1 遺伝子の高発現が T リンパ球の分化にも影響を及ぼして
いることが示唆された。これらの結果から、EVI1 遺伝子の高発現はリンパ球産生の
抑制に働くことが示された。白血病の発症機序を図18 に示した。3q 変異により EVI1
遺伝子が高発現すると、造血前駆細胞の増殖と巨核球への分化の偏りが生じ、一方
3q21q26-EVI1 マウスと比較して、Gata2 遺伝子のハプロ不全が MPP2 への分化をわ ずかに促進させていたものの、巨核球や血小板の増加は見られなかった。
Gata2 ハプロ不全において、Gata1 遺伝子の発現が減少しているかを確認するために、 WT マウス、Gata2+/gfp マウス、3q21q26-EVI1 マウス、3q21q26-EVI1::Gata2+/gfpマウ
スの巨核球でのGata1 遺伝子の発現を解析した。成体マウスの巨核球を用いた解析
では、巨核球の細胞数が少ないために、信頼できる結果が得られなかったので、胎
児肝臓からトロンボポエチンによって培養した巨核球(CD41+細胞)を用いて EVI1,
Gata2, Gata1 の mRNA 発 現 量 を 解 析 し た 。 3q21q26-EVI1 マ ウ ス と
3q21q26-EVI1::Gata2+/gfpマウスでトランスジーンによるEVI1 遺伝子の高発現がみら れ(図 19A)、3q21q26-EVI1 マウスではそれに伴う Gata2 遺伝子発現の上昇が見られ た(図 19B)。3q21q26-EVI1::Gata2+/gfpマウスでは、3q21q26-EVI1 マウスと比較して Gata2 遺伝子発現の低下傾向がみられたが、有意差は見られなかった(図 19B)。こ の細胞においてGata1 遺伝子の発現を解析したところ、Gata2 遺伝子と同様に低下 傾向がみられたものの、有意差は得られなかった(図 19C)。現時点では、Gata1 の 巨核球形成への関与は不明である。
(8) 3q 白血病において Gata2 遺伝子のハプロ不全が血小板と巨核球の増加を伴 う白血病を発症させる 3q21q26-EVI1 マウスと 3q21q26-EVI1::Gata2+/gfpマウスは白血病を発症するが、WT マウスと Gata2+/gfpマウスは発症しない 18。EVI1 遺伝子の発現異常が白血病発症前 から巨核球増多を引き起こしていたことから、白血病を発症した 3q21q26-EVI1 マ ウスと3q21q26-EVI1:: Gata2+/gfpマウスで末梢血の血小板数を調べた。マウスの末梢 血の血算を測定し、白血球数が5×10^4 個/μl 以上を白血病発症とみなして安楽死 し、解析した。3q21q26-EVI1 マウスでは同週齢の WT マウスと比べて、ほとんどの マウスで血小板が低値であったのに対し、3q21q26-EVI1:: Gata2+/gfpマウスでは血小 板が低値のマウスの他に正常〜高値のマウスも存在していた(図 20A)。次に、骨髄 中におけるCD41+巨核球の割合を調べた。コントロールとして解析した WT マウス の骨髄に占めるCD41+細胞の割合は 10 匹の平均で 1.97%であったことから、白血 病マウスを骨髄に占める巨核球の割合で、High (10%以上) -, Medium(2-10%)-, Low(2% 未 満 )-Megakaryocyte に 分 け た と こ ろ 、 3q21q26-EVI1 マ ウ ス で は 、 High-Megakaryocyte 白血病が 25 匹中 1 匹(4%)であったが、3q21q26-EVI1::Gata2+/gfp
マウスでは25 匹中 6 匹(24%)が High-Megakaryocyte 白血病であった(図 20B)。ま た、骨髄中の巨核球の細胞数の平均値は 3q21q26-EVI1 マウスで 7.07×105 個、 3q21q26-EVI1::Gata2+/gfp マウスで 2.17×106個であり 3q21q26-EVI1::Gata2+/gfpマウ スで多かった。 これらの結果から、EVI1 高発現白血病では Gata2 遺伝子のハプロ不全が伴うことで、 巨核球と血小板が正常〜高値の白血病を発症することが示唆された。巨核球の多少 が EVI1 高発現白血病のトランスフォーメーションと発症に関係しているのかを調 べ る た め に 、3q21q26-EVI1 マ ウ ス の Low-Megakaryocyte 白 血 病 (n=21) と High-Megakaryocyte 白 血 病 (n=1) 、 3q21q26-EVI1::Gata2+/gfp マ ウ ス の Low-Megakaryocyte 白血病(n=15)と High-Megakaryocyte 白血病(n=6)について、誕生 から白血病発症までのコホートを調べた(図 21)。3q21q26-EVI1::Gata2+/gfp マウス High-Megakaryocyte 白血病の発症までの期間は同マウスの Low-Megakaryocyte 白血 病と同程度であったが、3q21q26-EVI1 マウス Low-Megakaryocyte 白血病と比べると 有意に短くなっていた。これは、以前に明らかにした、3q21q26-EVI1 マウスよりも 3q21q26-EVI1::Gata2+/gfpマウスの方が早期に白血病を発症する、すなわち Gata2 遺
伝子のハプロ不全は白血病のトランスフォーメーションに関与する 18 という結果 に 合 致 し て い た 。 し か し な が ら 、3q21q26-EVI1::Gata2+/gfp マ ウ ス の Low- と High-Megakaryocyte 白血病を比べた場合に発症までの差は見られなかった。
(9) 高血小板を伴う 3q21q26-EVI1::Gata2+/gfpマウス白血病は、ヒト3q 変異 AML の特徴を有する
High-Megakaryocyte type の 3q21q26-EVI1::Gata2+/gfpマウス白血病の特徴を明らかに
することとした。末梢血のスメアではヘテロな像を呈しており(図 22A)、大きい核 と青い細胞質を持つ芽球様細胞(白い矢印)、分葉核を持つ骨髄球様細胞(黄色い 矢印)、そして大きい核と淡青で不定型の細胞質を持つ巨核球様細胞(黒い矢印) が観察された。また、巨大な血小板(矢印)も認めた。これらの末梢血の形態学的 特徴は、ヒト 3q 変異を伴う白血病の形態と似ていた。続いて、骨髄細胞のフロー サイトメトリーと形態学的解析を行った。骨髄中には、CD41+CD61+巨核球が増殖 していたが、同時にCD41-細胞も増殖しており、CD41-分画には、B220+Gr1-細胞、 B220-Gr1+細胞、B220-Gr1-細胞が含まれていた(図 22B)。それぞれの分画で未分化
マーカーである c-Kit の発現を見たところ、CD41-B220+Gr1-分画で c-Kit の発現が 高く(図 22C)、これは 3q21q26-EVI1::Gata2+/gfp Low-Megakaryocyte 白血病と同様で あった18。それぞれの分画で骨髄細胞の形態を見てみると、CD41+CD61+分画には 偽足を伴う細胞が含まれおり(図 22D)、CD41-B220+Gr1-分画は芽球様細胞が CD41-B220-Gr1+分画には分葉核を持つ骨髄球様細胞が含まれていた(図 22D)。 CD41+CD61+B220-Gr1-, CD41-B220+Gr1-, CD41-B220-Gr1+, CD41-B220-Gr1-分画そ れぞれのEVI1 遺伝子と Gata2 遺伝子の発現を調べた(図 22E)。CD41+CD61+巨核球
において、EVI1 遺伝子と Gata2 遺伝子の発現が最も高く、CD41-B220+Gr1-分画で もEVI1 遺伝子と Gata2 遺伝子の発現が高くなっていた。一方で、CD41-B220-Gr1+ 骨髄球細胞ではEVI1 遺伝子と Gata2 遺伝子の発現は低かった。 白血病幹細胞を同定するため、またCD41+CD61+細胞が白血病細胞由来であるか否 か を 検 討 す る た め に 、 巨 核 球 増 多 型 白 血 病 を 発 症 し た マ ウ ス の 骨 髄 細 胞 を CD41+B220-Gr1-分画、CD41-B220+Gr1-分画、CD41-B220-Gr1+分画に分離し、移植 実験を行った(図 23A)。それぞれの分画の細胞 5 万個を放射線照射した WT マウス に移植したところ、CD41+CD61+細胞を移植したマウスは白血病を発症しなかった。
一方で、CD41-B220+Gr1-分画を移植したマウスは高率に白血病を発症した。白血 病を発症したレシピエントマウスでドナー由来の骨髄細胞について見てみたとこ ろ、巨核球増多型白血病もしくは未分化型白血病であった。これらの結果から、巨 核 球 増 多 型 白 血 病 で 見 ら れ て い る 巨 核 球 は 白 血 病 幹 細 胞 で は な く 、 CD41-B220+Gr1-分画の細胞を起源に持つ白血病細胞であると考えられた。 3q21q26-EVI1 マウス群にも巨核球増多型白血病が 1 匹認められた(図 20AB)。まず 本個体が、3q21q26::Gata2+/gfpマウスでみられる高巨核球型白血病と同様のものであ るかの確認を行った。末梢血のスメア像(ライトギムザ染色)と骨髄のフローサイト メトリー解析の結果を図 24 に示した。この個体の表現型は、3q21q26::Gata2+/gfpマ ウスでみられる巨核球増多を伴う白血病の表現型とよく類似しており、このことか ら 3q21q26-EVI1 マウスでも少数ではあるが、3q21q26::Gata2+/gfpマウスでみられる 高巨核球型白血病と同様の白血病を発症することが示された。
6. 考察 これまでにも、EVI1 を発現しているレトロウイルスを形質導入した細胞を骨髄に移 植する方法やSca1 プロモーターにより EVI1 遺伝子の発現を誘導するトランスジェ ニックマウスで14-17EVI1 遺伝子を高発現させた AML や MDS のマウスモデルが樹 立されていた。これらのモデルマウスは白血病を発症することはあっても、巨核 球・血小板の増加というヒト 3q 白血病の重要な特徴を呈していなかった。本研究 において、3q21q26-EVI1::Gata2+/gfpマウスの 24%で巨核球・血小板の増加を伴う白 血病を発症しており、ヒト3q 白血病に近いモデルマウスを樹立できた。 白血病は、造血細胞の分化に障害が生じ、異常な造血幹細胞・前駆細胞が蓄積する ことで、発症の母体となる 19。3q21q26 マウスでは、白血病の発症メカニズムが分 かっていなかったため、発症機序を明らかにするために、レポーターマウスを樹立 し、白血病発症前の12 週齢マウスの骨髄の解析を行った。すると EVI1-/Gata2- High 細胞群が白血病発症前から蓄積しており、この細胞群は連続的なコロニー形成能を 有していることから、白血病の起源となる細胞が含まれていると考えられた。また、 High 細胞群には、HSPC のみでなく巨核球も含まれており、巨核球で G2DHE によ
るEVI1 遺伝子の誘導が起こることが 3q 白血病の巨核球増多を引き起こす原因のひ とつと考えた。 Gata2+/gfpマウスのLT-HSC や ST-HSC 等の未熟細胞群分画で High 細胞の比率を測定 した図10B において、GFP の発現が high でない細胞が 20-30%存在するが、LT-HSC やST-HSC の GATA2 発現に heterogeneity があると考えている。従来、造血幹細胞・ 前駆細胞における転写因子発現解析は細胞集団の平均値で行われていたが、近年 single cell の遺伝子発現解析が行われるようになり、造血幹細胞・前駆細胞の転写 因 子 発 現 は heterogeneity で あ る こ と が 明 ら か に な っ た 25,26。 造 血 幹 細 胞 の heterogeneity は、起源(中胚葉)の違いやニッシェ(血管内皮ニッシェもしくは造骨ニ ッシェ)の違い、細胞周期の違いなどから生じると考えられる。また本マウスモデ ルでは、Gata2 遺伝子の型アリルに GFP がノックインされているため、もう一方の アリルの Gata2 が発現している可能性も考えられる。一方で、本研究では High 細 胞をノックインマウス由来のGata2-GFP だけでなく、トランスジェニックマウス由 来の3q21q26-tdTomato の蛍光を用いて定義している。ノックインマウスと比べてト ランスジェニックマウスでは、位置効果(トランスジーンが挿入された周辺領域か
らの影響)のために、本来発現すべき細胞であるにも関わらず、トランスジーンが
発現しない場合がある。このマウスでは位置効果による3q21q26-tdTomato 発現の抑
制も関与している可能性がある。
CD41 が myeloid-biased adult hematopoietic stem のマーカーとなるとの報告もある。 しかし、図5A の CD41+CD61+細胞の c-Kit の発現を見たところ CD41+CD61+細胞
はc-Kit-細胞を含んでいる(図 10C)ことから、幹細胞ではなく、巨核球であると結論
づけた。GATA2 は、造血幹細胞・前駆細胞、赤血球・巨核球前駆細胞で発現してお
り、造血の初期段階を制御している。また、巨核球が分化する間に高く発現してお
り27,28、巨核球の最終分化にも関与している29。過剰に発現しているEVI1 は Gata2 発現を上昇させ(fetal liver の結果より)、Gata2 の発現が上昇することで、前駆細胞
は赤血球への分化が抑制され巨核球への分化が促進する30。12 週齢マウスの解析で
3q21q26-EVI1 マウス、3q21q26-EVI1::Gata2+/gfpマウス共に巨核球・血小板が増加し
ているのはGata2 の上昇によるもので、12 週齢以降に変異が加わることで白血病発
症後には両者に差が生じると考えた。
Gata2+/gfp)の骨髄・末梢血の解析を図 3,4 に示した。12 週齢の時点では、EVI1 高発
現の影響が顕著にみられている。血球の分化に及ぼす影響は図4L に示したように、
MPP2, MEP, 巨核球、末梢血の血小板の増加と MPP4, リンパ球の減少である。巨核 球増加の一方で、MEP が増加するものの赤芽球と赤血球が増加していない。EVI1
はGata2 遺伝子のプロモーターに結合してエンハンサーとして働く31が、過剰に発
現しているEVI1 は Gata2 発現を上昇させ、Gata2 の発現が上昇することで、前駆細
胞は赤血球への分化が抑制され巨核球への分化が促進する30と考えた。これまでに
作られたモデルマウス14-17も、造血前駆細胞の増殖、そして赤血球系細胞とリンパ
球系細胞の減少を示しており、これは今回の 3q21q26-EVI1 マウスの結果と一致し
ていた。
EVI1 は Gata2 発現を上昇させ、Gata2 の発現が上昇することで、前駆細胞は赤血球
への分化が抑制され巨核球への分化が促進する30。また、GATA2 は EPO のプロモ
ーター領域に結合することでその活性を抑制する 32 ことから赤血球が増加しない
と考えられる。またEVI1 は GATA1 を抑制する作用があることが示されている。赤
巨核球への分化を進める要因である可能性が考えられる。 12 週齢の時点でも、WT と Gata2+/gfp マウスとの間、3q21q26-EVI1 マウスと 3q21q26-EVI1::Gata2+/gfp マウスとの間で、造血幹細胞・前駆細胞の細胞数、全骨髄 細胞数に占める割合においてほとんど差は見られておらず、白血病発症前(白血病 発症に必要な遺伝子変異(セカンドヒット)が入る前)には Gata2 ハプロ不全の影響は 小さいと考えられる。Gata2 ハプロ不全が巨核球増多型白血病の発症を促進するメ カニズムについては、セカンドヒットがある状態で解析しなければ解明できないと 考えられ、現在、セカンドヒットの同定を検討している。 白血病を発症したマウスを見てみると、3q21q26-EVI1::Gata2+/gfp マウスにおいて巨 核球・血小板の増多を伴う白血病を発症している割合が高く、EVI1 遺伝子の高発現 にGata2 遺伝子ハプロ不全が加わることで、このフェノタイプが惹起されることが 分かった。しかしながら、EVI1 遺伝子の高発現と Gata2 遺伝子ハプロ不全が白血病 を発症させる分子メカニズムは不明である。12 週齢の段階では、3q21q26-EVI1 マ ウスより3q21q26-EVI1::Gata2+/gfpマウスでLSK 細胞に占める MPP2 細胞の割合がわ
ずかながら上昇している他に差が見られない。Gata2+/-マウスは明らかな問題はなく 生存できるが、6-12 週齢になると造血幹細胞の増殖能は野生型マウスと比べて劣っ ている23。12 週齢以降に 3q21q26-EVI1::Gata2+/gfpマウスに何らかのセカンドヒット が加わり、巨核球・血小板増加のフェノタイプが起こると考えられる。また、Gata2 とGata1 の発現の変化による可能性も考えられる。Gata2 の発現上昇は、赤血球と 巨核球の分化を制御しているGata1 の発現を上昇させる。Gata1 の発現は高値であ ると赤血球の分化を促進するが、低値であると巨核球の分化を促進する 33。Gata2 遺伝子ハプロ不全がGata1 発現を低下させることで、巨核球への分化を促進させる
ことが考えられる。さらに、Gata2 は Gata1 の非存在下で巨核球形成に働くが、Gata2
が遺伝子を制御することは細胞周期と巨核球分化に関連している34。これらの結果
から、Gata2 遺伝子ハプロ不全は EVI1 遺伝子を高発現している白血病細胞が巨核球
に分化するのを促進すると考えられる。
先行研究(Katayama et al., 2017)で、3q21q26-EVI1::Gata2+/gfpマウスでは、3q21q26-EVI1
マウスと比較して未分化型の白血病の頻度が増加することを示した。しかし、
め、このマウスの白血病幹細胞を含む細胞画分(B220+Gr1-)において Gata2 の発現を 解析したところ、未分化型以外(骨髄球への分化能を示す)の白血病マウスの B220+Gr1-細胞と比較して、Gata2 遺伝子の発現が低下していることが示された。 このことから突発的に起こる Gata2 遺伝子の発現低下が 3q21q26-EVI1 マウスにお ける未分化型白血病発症の原因であると考えられた。このことから、血小板高値の 3q21q26-EVI1 白血病マウスも Gata2 の発現が減少し、この表現型になった可能性が 考えられる。現在、3q21q26-EVI1 マウスで血小板高値を伴う白血病を発症する個体 が1 個体しか得られていないので、Gata2 遺伝子の発現は解析できていないが、今 後、同様の個体が得られた場合には、解析を行う予定である。 本研究で用いたGata2+/gfpマウスはすべての分化段階の細胞でGata2 遺伝子の発現が 低下している。一方で、-77 エンハンサー(G2DHE)を欠失したマウスでは、LSK で はGata2 遺伝子の発現低下はみられず、MEP 以降の段階で Gata2 遺伝子の低下が示
されている。このことから、特に MEP 以降の分化段階で Gata2 遺伝子の発現低下
が白血病発症に関与している可能性が考えられる。しかし、先行研究では、LSK の
みられたMPP2 でも低下している可能性は残る。そのため、MPP2 を巨核球系列の
細胞での細胞でGata2 ハプロ不全の影響がでる可能性を考えて、今後の研究は進め
ていきたいと考えている。
3q21q26-EVI1::Gata2+/gfpマウスのLow-と High-Megakaryocyte 白血病を比べた場合に
発症までの差は見られなかった(図 21)。3q21q26-EVI1::Gata2+/gfp Low 群では、骨髄
は B220+Gr1-c-Kit+細胞で占められた未分化型白血病である。一方で巨核球増多型
白血病は、CD41+CD61+細胞、B220+細胞、Gr1+細胞が混在した白血病である。先 行論文(Katayama et al., 2017)では、3q21q26-EVI1 白血病マウスで TypeⅠ(骨髄におけ る未分化細胞(B220+細胞)の割合が 80%以上)が最も早期に白血病を発症しており、 次いで TypeⅡ(未分化細胞:40-80%)が発症し、TypeⅢ(未分化細胞が 40%以下であ り、その他は Gr1+細胞で占められる)は発症が最も遅いことが示されている。巨核 球増多型白血病では、Gr1+細胞が骨髄細胞の約 30-70%を占めているにも関わらず、 未分化型白血病(Low 群)と同程度に発症が早いという結果は、巨核球が巨核球増多 型白血病の発症を促進する可能性があると考えた。 ヒト3q AML には巨核球と血小板の増加を呈する症例があるが11,12、その病態的な
意義はわかっていない。正常状態において、造血幹細胞・前駆細胞はニッシェとい
う微小環境下で維持されている35。骨髄内には、骨芽細胞ニッシェと血管ニッシェ
があるが、巨核球は血管壁から偽足を伸ばして血小板を産生する他に、様々な因子
を 分 泌 す る こ と で 血 管 ニ ッ シ ェ を 維 持 ・ 制 御 し て い る 。 例 え ば 、 巨 核 球 は
CXCL4(platelet factor 4)や TGF-β(transforming growth factor β1)を分泌して HSC を 休止状態(homeostatic quiescence)に保つ働きがある36,37。また、化学療法といったス
トレス下では、巨核球はFGF-1(fibroblast growth factor 1)を分泌して HSC 休止を保つ
働きもある37。これらのことから、巨核球増多型白血病では、白血病幹細胞が休止 の状態で維持されている仮説や、ヒト 3q 白血病においては白血病幹細胞が休止の 状態であるために化学療法が効かないという仮説を考えている。他にも、巨核球が 産生する血小板由来増殖因子(PDGF)などの因子は血管ニッシェに影響を与える 38。 固形癌のようにAML の骨髄でも血管新生が生じているが、巨核球は、血管新生を 促進する因子(VEGF, TGFβ, b-FGF)を分泌し、血管新生を亢進させていると考えられ る35。このように、増殖している巨核球・血小板が白血病の悪性化に寄与する仮説 を考えている。しかしながら、TGF-β や PDGF などの巨核球が産生する物質を測定
していないため、仮説の域を出ていない。そのため、今回、考察には仮説の一つと して記述した。まずは巨核球が白血病の悪性化の原因となっているか否かを明らか にすることが最優先課題であると考えており、そのため、条件付きジフテリア毒素 受容体発現マウスと巨核球特異的Cre マウスを組み合わせた巨核球枯渇実験を検討 している。3q21q26-EVI1::Gata2+/gfp の前白血病及び白血病マウスにおいて、巨核球 を枯渇させた場合に造血前駆細胞の蓄積が抑制されるか、また白血病は抑制される かを検討したい。
7.結論
3 番染色体の逆位により誘導される EVI1 遺伝子の発現は、造血幹細胞・前駆細胞と 巨核球で高い。
3 番染色体の逆位により誘導される EVI1 遺伝子の高発現と Gata2 遺伝子ハプロ不全 の両方が起こることで、巨核球と血小板の増多を伴う白血病が発症する。
8.参考文献
1. Byrd JC, Mrozek K, Dodge RK, et al. Pretreatment cytogenetic abnormalities are predictive of induction success, cumulative incidence of relapse, and overall survival in adult patients with de novo acute myeloid leukemia: results from Cancer and Leukemia Group B (CALGB 8461). Blood. 2002;100(13):4325-4336.
2. Slovak ML, Kopecky KJ, Cassileth PA, et al. Karyotypic analysis predicts outcome of preremission and postremission therapy in adult acute myeloid leukemia: a Southwest Oncology Group/Eastern Cooperative Oncology Group Study. Blood.
2000;96(13):4075-4083.
3. Cui W, Sun J, Cotta CV, Medeiros LJ, Lin P. Myelodysplastic syndrome with inv(3)(q21q26.2) or t(3;3)(q21;q26.2) has a high risk for progression to acute myeloid leukemia. Am J Clin Pathol. 2011;136(2):282-288.
4. Haferlach C, Bacher U, Haferlach T, et al. The inv(3)(q21q26)/t(3;3)(q21;q26) is frequently accompanied by alterations of the RUNX1, KRAS and NRAS and NF1 genes and mediates adverse prognosis both in MDS and in AML: a study in 39 cases of MDS or AML. Leukemia. 2011;25(5):874-877.
5. Lugthart S, Gröschel S, Beverloo HB, et al. Clinical, molecular, and prognostic significance of WHO type inv(3)(q21q26.2)/t(3;3)(q21;q26.2) and various other 3q abnormalities in acute myeloid leukemia. J Clin Oncol. 2010;28(24):3890-3898.
6. Rogers HJ, Vardiman JW, Anastasi J, et al. Complex or monosomal karyotype and not blast percentage is associated with poor survival in acute myeloid leukemia and myelodysplastic syndrome patients with inv(3)(q21q26.2)/t(3;3)(q21;q26.2): a Bone Marrow Pathology Group study. Haematologica. 2014;99(5):821-829.
7. Morishita K, Parganas E, William CL, et al. Activation of EVI1 gene expression in human acute myelogenous leukemias by translocations spanning 300-400 kilobases on chromosome band 3q26. Proc Natl Acad Sci U S A. 1992;89(9):3937-3941.
8. Gröschel S, Sanders MA, Hoogenboezem R, et al. A single oncogenic enhancer rearrangement causes concomitant EVI1 and GATA2 deregulation in leukemia. Cell.
9. Yamazaki H, Suzuki M, Otsuki A, et al. A Remote GATA2 Hematopoietic Enhancer Drives Leukemogenesis in inv(3)(q21;q26) by Activating EVI1 Expression. Cancer Cell. 2014;25(4):415-427.
10. Fonatsch C, Gudat H, Lengfelder E, et al. Correlation of cytogenetic findings with clinical features in 18 patients with inv(3)(q21q26) or t(3;3)(q21;q26). Leukemia. 1994;8(8):1318-1326.
11. Secker-Walker LM, Mehta A, Bain B. Abnormalities of 3q21 and 3q26 in myeloid malignancy: a United Kingdom Cancer Cytogenetic Group study. Br J Haematol. 1995;91(2):490-501.
12. Grigg AP, Gascoyne RD, Phillips GL, Horsman DE. Clinical, haematological and cytogenetic features in 24 patients with structural rearrangements of the Q arm of chromosome 3. Br J Haematol. 1993;83(1):158-165.
13. Bitter MA, Neilly ME, Le Beau MM, Pearson MG, Rowley JD. Rearrangements of chromosome 3 involving bands 3q21 and 3q26 are associated with normal or elevated platelet counts in acute nonlymphocytic leukemia. Blood. 1985;66(6):1362-1370.
14. Buonamici S, Li D, Chi Y, et al. EVI1 induces myelodysplastic syndrome in mice. J
Clin Invest. 2004;114(5):713-719.
15. Louz D, van den Broek M, Verbakel S, et al. Erythroid defects and increased retrovirally-induced tumor formation in Evi1 transgenic mice. Leukemia. 2000;14(11):1876-1884.
16. Yoshimi A, Goyama S, Watanabe-Okochi N, et al. Evi1 represses PTEN expression and activates PI3K/AKT/mTOR via interact37. Katayama S, Suzuki M, Yamaoka A, et al. GATA2 haploinsufficiency accelerates EVI1-driven leukemogenesis. Blood.
2017;130(7):908-919.
17. Ayoub E, Wilson MP, McGrath KE, et al. EVI1 overexpression reprograms hematopoiesis via upregulation of Spi1 transcription. Nat Commun. 2018;9(1):4239
18. Katayama S, Suzuki M, Yamaoka A, et al. GATA2 haplosufficiency accelerates EVI1-driven leukemogenesis. Blood. 2017;130(7):908-919
20. Suzuki N, Ohneda O, Minegishi N, et al. Combinatorial Gata2 and Sca1 expression defines hematopoietic stem cells in the bone marrow niche. Proc Natl Acad Sci U S A. 2006;103(7):2202-2207.
21. Brandt W, Khandekar M, Suzuki N, Yamamoto M, Lim KC, Engel JD. Defining the functional boundaries of the Gata2 locus by rescue with a linked bacterial artificial chromosome transgene. J Biol Chem. 2008;283(14):8976-8983.
22. Khandekar M, Suzuki N, Lewton J, Yamamoto M, Engel JD. Multiple, distant Gata2 enhancers specify temporally and tissue-specific patterning in the developing urogenital system. Mol Cell Biol. 2004;24(23):10263-10276.
23. Rodrigues NP, Janzen V, Forkert R, et al. Haploinsufficiency of GATA-2 perturbs adult hematopoietic stem-cell homeostasis. Blood. 2005;106(2):477-484.
24. Nakano H, Yanagita M, Gunn MD. CD11c(+)B220(+)Gr-1(+) cells in mouse lymph nodes and spleen display characteristics of plasmacytoid dendritic cells. J Exp Med. 2001;194(8):1171-1178.
25. Moignard V, Gottgens B, et al. Characterization of transcriptional networks in blood stem and progenitor cells using high-thoughput single-cell gene expression analysis, nature
cell biology. 2013;15:363-373
26. Crisan M, Dzierzak E. The many faces of hematopoietic stem cell heterogeneity, The
Company of Biologists. 2016;143:4571-4581
27. Terui K, Takahashi Y, Kitazawa J, Toki T, Yokoyama M, Ito E. Expression of transcription factors during megakaryocytic differentiation of CD34+ cells from human cord blood induced by thrombopoietin. Tohoku J Exp Med. 2000;192:259-73
28. Pronk C, Bryder D, et al. Elucidation of the phenotypic, functional, and molecular topography of a myeloerythroid progenitor cell hierarchy. Cell Stem Cell.2007;428-442 29. Visvander JE, Crossley M, Hill J, Orkin SH, Adam JM.The C-terminal zinc finger of GATA-1 or GATA-2 is sufficient to induce megakaryocytic differentiation of an early myeloid cell line. Mol Cell Biology. 1995;15:634-41
30. Ikonomi P, Noguchi C, et al. Overexpression of GATA-2 inhibits erythroid and promotes megakaryocyte differentiation. Exp. Hematol. 2000;28:1423-1431
hematopoietic stem cell proliferation through GATA-2 expression. The EMBO Journal. 2005;24:1976-1987
32. Imagawa S, Suzuki N, Ohmine K, et al. GATA suppresses erythropoietin gene expression through GATA site in mouse erythropoietin gene promotor, Int J Hematol. 2002;75:376-81
33. Gutierrez L, Tsukamoto S, Suzuki M, et al. Ablation of Gata1 in adult mice results in aplastic crisis, revealing its essential role in steady-state and stress erythropoiesis. Blood. 2008;111(8):4375-4385.
34. Huang Z, Dore LC, Li Z, et al. GATA-2 reinforces megakaryocyte development in the absence of GATA-1. Mol Cell Biol. 2009;29(18):5168-5180.
35. Psaila B, Lyden D, Roberts I. Megakaryocytes, malignancy and bone marrow vascular niches. Journal of Thrombosis and Haemostasis. 2011;10:177-188
36. Bruns I, Lucas D, Pinho S, et al. Megakaryocytes regulate hematopoietic stem cell quiescence through CXCL4 secretion. Nat Med. 2014;20(11):1315-1320.
37. Zhao M, Perry JM, Marshall H, et al. Megakaryocytes maintain homeostatic quiescence and promote post-injury regeneration of hematopoietic stem cells. Nat Med. 2014;20(11):1321-1326.
38. Italiano JE, Richardson JL, Patel-Hett S, et al. Angiogenesis is regulated by a novel mechanism: pro- and antiangiogenic proteins are organized into separate platelet alpha granules and differentially released. Blood. 2008;111(3):1227-1233.
9. 表
表1.フローサイトメトリー解析に用いた抗体リスト
Epitope Conjugate Clone Company
B220 PE-Cy7 RA3-6B2 TONBO
B220 APC-Cy7 RA3-6B2 BD CD19 Biotin 1D3 TONBO CD19 BV510 1D3 BD Gr1 BV510 RB6-8C5 BioLegend Gr1 Biotin RB6-8C5 eBioscience Ly6G BV510 1A8 BD
CD115 APC AFS98 BioLegend
CD41 PE MWReg30 BD
CD41 APC MWReg30 BioLegend
CD61 BV421 2C9.G2 BD
CD71 BV421 RI7217 BioLegend
Ter119 APC TER-119 BD, eBioscience
Ter119 Biotin TER-119 eBioscience
c-Kit APC-eFluor 780 2B8 eBioscience
Sca1 PE-Cy7 D7 BD Flt3 BV421 A2F10 BioLegend Flt3 PE A2F10 BioLegend CD150 APC TC15-12F12.2 BioLegend CD48 PE HM48-1 BioLegend CD48 BV510 HM48-1 BD
CD3e Biotin 145-2C11 eBioscience
CD3 APC-Cy7 17A2 BioLegend
CD4 Biotin GK1.5 BioLegend
Mac1 Biotin M1/70 eBioscience
CD34 BV421 RAM34 BD
FcγR II/III BV510 2.4G2 BD
CD11c PE-Cy7 N418 BioLegend
streptavidin PerCP – BioLegend
streptavidin BV421 – BioLegend
表2.定量的RT-PCR に用いたプライマー配列
Gene Forward primer Reverse primer
EVI1 ACAAGCCAAGACCAGCCCCTGG TGGTTTTTTCGAGGCTCAGTC
Gata2 CCTGGTTCCCAAGACACAGTAG GGGTGCTGCGCATTCAATAC
Gapdh TGCACCACCAACTGCTTAG GGATGCAGGGATGATGTTC
表3. BAC の組み換えに用いたプライマー配列 Primers Sequences TomatoFrtAmp(f) TATCGATACCGTCGAAGGGGCGGATCCTGAAGTT TomatoFrtAmp(r) CCCCCTCGAGGTCGAGCTCAAGCTTCGAA 3q21q26tdTomato(f) GAGTAGTTGCAGACATTGCGCCGGGAGAGGAGCTTCTGCTGTTCATG GTGAGCAAGGGCGAGG 3q21q26tdTomato(r) GCGATGATAAGGTGATAAGGAGGGTGGCGTGAGTGGTACTAACGCTC AAGCTTCGAATTC
10. 謝辞
本研究を行うにあたり、素晴らしい研究の環境を与えて下さり、ご指導頂いた医化
学分野の山本雅之教授に感謝いたします。また、直接ご指導頂いた医学部ラジオア
イソトープセンターの鈴木未来子講師に感謝いたします。その他、ご協力を頂きま
図 1. 3q21q26-tdTomato BAC トランスジェニックマウスの樹立
(A) tdTomato 蛍光がEVI1 遺伝子の翻訳開始点に挿入されており、G2DHE により
tdTomato 蛍光が発現する。(B) 3q21q26-tdTomato BAC 構築。EVI1 3rd エキソン
の翻訳開始点にtdTomato 遺伝子を挿入した。プローブ(オレンジの四角)と制限
酵素により切断される位置(B: BamHI, E: EcoRI)を示す。(C) 3q21q26-tdTomato BAC のサザンブロット。(D) lineA,B,C のコピー数。
図 2. 3q21q26-tdTomato レポーターマウスは G2DHE によるEVI1 遺伝子 の発現をモニターできている
tdTomato 蛍光の発現を解析したところ、LSK 分画で最も高く次いで LK 分画で高