高圧力下で結晶成長させたグルコースイソメラーゼ結晶
の常圧下での X 線結晶構造解析
塚本雅之
1、鈴木良尚
*2、櫻庭春彦
3、田村勝弘
2 1徳島大学大学院先端技術科学教育部環境創生工学専攻化学機能創生コース 2徳島大学大学院ソシオテクノサイエンス研究部ライフシステム部門 3香川大学農学部応用生物科学科 *E-mail:[email protected] 第15 回生物関連高圧研究会 20 周年記念シンポジウム抄録集 2008 年 2 月 27 日受付/2008 年 3 月 5 日受理 要旨キャピラリー中にグルコースイソメラーゼ(GI)の過飽和溶液を満たし、0.1 MPa と 100 MPa
下で制御をしながら育成した結晶を用いて、共に0.1 MPa 下で X 線結晶構造解析を行った。 結晶化の制御においては、0.1 MPa、100 MPa 下で共に温度をコントロールすることで、キ ャピラリー中の結晶の数を抑え、X 線結晶構造解析が可能な良質で大きな結晶を作製する ことができた。得られた結晶を両方とも0.1 MPa 下に取り出してX線結晶構造解析を行っ たところ、キャピラリー中に溶液が入ったままで高分解能の構造解析が可能なことが確認 できた。また、溶液共存下での結晶サイズと分解能の相関についても明らかにできた。し かし、構造解析の結果、0.1 MPa と 100 MPa 下で作製した結晶の間で分子構造や水和構造 に明確な変化はなかった。2 つのタンパク質分子を最小二乗法による重ね合わせで、対応 する原子間距離を求めてみると、最大で約1.37 Å、平均で約 0.078 Åの位置の違いしかな かった。また、水和構造においては、観測された水分子の位置について、得られた結果の 中で最大約10 %の位置の違いがあったが、0.1 MPa と 100MPa 下で作製した結晶の間での 有意差は認められなかった。 キーワード:キャピラリー、X 線結晶構造解析、結晶化制御、三次元分子構造、 水和構造 1. はじめに タンパク質の機能は、タンパク質の特異的な三次元分子立体構造に依存している。その中 で圧力変化に対して酵素活性が変化するタンパク質が存在することが報告されている1-5)。こ れは、圧力変化に伴って、基質やタンパク質分子の構造6-8)、もしくはそれらの水和構造8)が 変化することなどによるものと考えられる。これまでに、高圧 NMR の研究で、タンパク質 の分子構造が圧力の変化とともに変化することなどがわかっている 7)。また、超音波音速と 密度の測定による圧縮率の研究から、タンパク質の構造や水和構造に対してリガンド結合が 影響を及ぼすことがわかっている 9)。しかし、構造の変化についての原子レベルでの詳細な 情報を得るためには、更なる高分解能の構造情報が必要であろう。 そこで、我々は上記手法と相補的な方法として、X 線結晶構造解析を用い、高圧力下で変 形したタンパク質分子を構成単位(Fig. 1 )として、それを結晶化という形で固定し、解析する ことで高圧力下のタンパク質の分子構造を明らかにすることを考えた。X 線結晶構造解析を 行うことでより高分解能、高分子量の構造解析ができて、原子レベルで分子構造の議論を行 える。これまでに高圧力を用いたタンパク質のX 線結晶構造解析に関する研究例は、全て高
圧力を部分的に用いるものであった。例えば、常圧下で結晶作製したものを用いて、高圧力 下でX 線結晶構造解析を行ったもの10-12)と高圧力下で結晶作製したものを常圧下でX 線結晶 構造解析を行った 13)というようなものである。それらの結果においては、結晶の格子定数が 約 1 %の減少10,11)や微小なタンパク質分子構造の変化 10,12)や水和していると考えられる水分 子の数の増減10∼13)がみられた。例えば、ソーマチンでは49 個の水分子が常圧下で育成した結 晶の構造(282 個)から高圧力下で育成したものの構造(233 個)で減少している13) 。しかし、 いずれの場合もすべての過程を高圧力下で行っていないため、高圧力の効果を純粋に評価で きていないと考えられる。 このことから、我々は①高圧力下の中で結晶を育成し、②高圧力下を維持しながらX 線を 照射するというプロセスを合わせて行うことを目的とした。これを『その場高圧力下X 線結 晶構造解析』とする。実際には、ベリリウム(Be)製高圧セル中にタンパク質溶液を入れた キャピラリーをセットして、静水圧で高圧力をかけて結晶の育成を行う。そして、十分に大 きく成長した結晶がBe 製高圧セル中に存在していると仮定し(Be セル内部は外から見えない ので)、高圧力を維持しながら、Be セルごと X 線を照射して構造解析を実施する。この様な 解析を圧力を細かく変化させて行い、連続的に圧力を変化させていくことに伴うタンパク質 の構造の変化と機能の相関を調べることができれば、圧力による構造変化がフォールディン グ状態の存在確率のみによる不連続なものなのか、それとも連続的なものなのかという問い に解答を与えることができる可能性を秘めている。 本研究では、その場高圧力下 X 線結晶構造解析の途中経過として、1)キャピラリー中で、 GI の過飽和溶液中での0.1 MPa と 100 MPa 下での結晶化制御方法の確立と 2)キャピラリー中
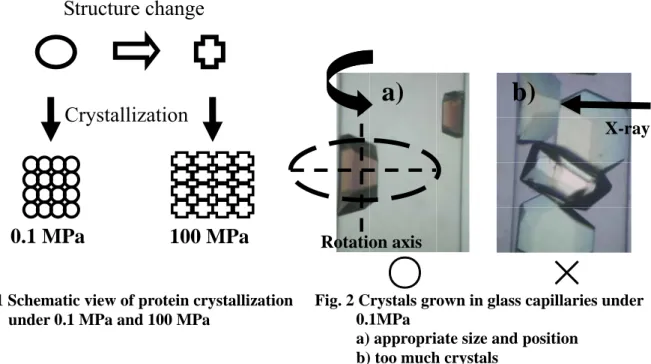
に溶液存在下でのX 線構造解析の実施(Fig. 2)及び 3)0.1 MPa と 100 MPa で育成した結晶中の
常圧下における分子構造の比較・考察結果について報告する。
Structure change
b)
a)
2. 実験方法 2.1. GI の溶液調製 本研究ではモデルタンパク質として、結晶化が高圧力下でコントロールしやすい点を考慮 して、高圧力下で結晶化が促進される GI14.15)を用いた。キャピラリー中で結晶化制御を精密 に行うために、スタート時は常に一定濃度(33 mgmL-1)の GI 水溶液を用いた。その溶液調製のCrystallization
0.1 MPa
100 MPa
Fig. 1 Schematic view of protein crystallization under 0.1 MPa and 100 MPa
Fig. 2 Crystals grown in glass capillaries under 0.1MPa
a) appropriate size and position b) too much crystals
X-ray
○
×
手順を以下に示す。GI の試料原液(Stretomyces rubiginosus ,EC5.3.1.5, Hampton Research Corp.)
は、結晶を含む懸濁溶液なので40 ℃の恒温槽に入れ結晶を 30 分間溶解させる。その後。微
小な結晶等の不純物をフィルター(Whatman SYRINGE FILTER 0.45 μm 直径 13 mm)を用いて
ろ 過 す る 。 そ の ろ 液 を 緩 衝 溶 液 (6 mM Tris(Hydroxymethyl)Aminomethane, 120 mgmL-1
Ammonium Sulfate, 1 mM Magnesium Sulfate, pH = 7.0 ± 0.05)で希釈し、仕込み溶液を調製する。
仕込み溶液の濃度は、溶液の一部を緩衝溶液で 20~100 倍に希釈し、分光光度計(HITACHI U-2001)で吸光度測定することにより仕込み溶液の濃度を決定する。Hampton Research 社の 製品データによると濃度が0.96 mgmL-1の時に280 nm の波長で吸光度が 1 となることから、 得られた吸光度を用いて計算することで濃度を決定することができる。 2.2. 結晶の育成方法 本研究では、単結晶を回転させて X 線結晶構造解析を行うために、X 線の照射するポイン トに結晶が一つだけ存在し、その結晶を通ってビームに平行な回転面内に、他の結晶が存在 しないようにする必要がある(Fig. 2 a) )。結晶が X 線の照射される方向に重なってしまうと (Fig. 2 b) )、その結晶からの反射が元の結晶からの反射に重なることで単一の構造を解析する ことができなくなり、構造を決定することが不可能となる。この問題を解決するには、まず 結晶の数を制御することが必要となる。結晶数を制御するためには、結晶の核生成を制御す る必要がある。結晶成長過程は、過飽和溶液中に臨界核という微結晶が生成する核生成の段 階と、その臨界核上に分子が結合していく成長の段階に分けられる。両方とも、過飽和状態 で反応の駆動力が与えられた状態でのみ起こるが、一般的に核生成の方がより大きな駆動力 を必要とする。その関係をFig. 3 に示す。 Fig. 3 Control of supersaturation based on a phase diagram.
The black line presents a solubility curve. Nucleation can occur above dashed line.(A) Stage of nucleation (B) Stage of crystal growth
結晶成長の駆動力は、過飽和度σ = ln(C/Ce) (C は溶液の濃度、Ceは溶解度)で示される。Fig.3 に示すような相図において、σ は温度が低く、濃度が高いほど大きくなる。よって、ある一 定時間内に核生成が起こる領域は、図の左上になり、その境界が破線のように引けるはずで ある。この破線と溶解度曲線にはさまれた領域は、核生成は起こさないが結晶は成長する準 安定領域となる。この相図上で状態を変化させることで結晶化が制御できる。例えば、点 A の条件で一定時間経過させることで核生成が起こる。そこから温度を上昇させることで点 B の条件にするとσ が減少して結晶は成長しかしないようになる。実際には C = 25 ~ 28 mgmL-1 の溶液が入った GC を常圧下と 100 MPa 下におき、恒温槽中で結晶が核生成する状態(常圧 下: 22 ~ 23 ℃、100 MPa 下: 29 ℃)で一定時間経過後に、徐々に温度を上昇させて結晶の成 長だけが進む状態B(常圧下: 25 ~ 26 ℃、100 MPa 下: 32 ℃)で結晶の育成を行った。 2.3. X 線結晶構造解析 X 線結晶構造解析装置は、R-axis Ⅶ(Rigaku)を用いて行った。X 線発生装置のターゲットは
銅で、波長は1.5418 Åである。GC を用いた一般的な X 線回折実験は、結晶を別の環境相で 作製して、その中から大きく良質な結晶を選択し、GC 中にセットしてほとんど周りに溶液が ない状態で行うものである。しかし、その場高圧力下X 線結晶構造解析を行うためには、溶 液を通して常に結晶が加圧されている必要があるので、溶液が満たされた GC 中に直接結晶 を作製して、溶液共存下でX 線回折実験を行う必要がある。理想的には、上述した結晶の育 成方法で、Fig. 2 a)のような状態を作り出すことが必要である。今回の撮影方法は、すべてカ メラ距離が 150 mm で 1 °振りを 1 分間で行い、180 °回転させて回折データを Crystal Clear(Rigaku)というソフトウェアで取得して解析する。 3. 結果と考察 3.1 核生成の制御 本研究の結晶の育成方法を用いることで、結晶の数を制御することができた。例として、 溶液濃度が約27 mgmL-1で温度制御(常圧下22 ℃ 4 時間,23 ℃ 3 時間, 25 ℃ 2 日間)を行った ものの経過をFig. 4 a)に示す。構造解析に適した大きくて良質な単結晶を育成することに成 功した。同じ溶液濃度で常圧下 22 ℃ を 2 日間という一定温度以外何も制御しない状態で結 晶化を行うとFig.4 b)のようになった。この状態では構造解析できない。さらに、高圧力下で も同様に制御することができた。なおこの方法においては、結晶がキャピラリーの壁面に付 着して成長するためにその場に固定されており、X 線結晶構造解析を行う際に、手間のかか る結晶のマウント作業を省略できるというメリットもある。
a) b)
0.1 mm
Fig. 4 Crystal growth at 0.1 MPa in GC with (a)) and without (b)) nucleation control 3.2 結晶の大きさと溶液共存下での分解能 タンパク質のX 線結晶構造解析を実施して、三次元分子構造を決定していく際に分解能が 非常に重要になる。分解能の値が4.0~3.5 Åでアミノ酸残基の空間的な位置を議論することが でき、2.5 Å程度で残基の側鎖の向きを大まかに決定することができる。そして、2.0Å近く のデータを得ることができると正確に側鎖の構造を決定し、水和水の議論もできるようにな る 16)。タンパク質の結晶は、タンパク質分子自身が大きいことや水を多く含むような結晶が 生成するために、結晶が小さいと十分な回折強度を得ることができない。このことを改善す るためには、大きな結晶を作製することがタンパク質分子の規則的な配列のできる数を多く するためには必要である。そこで、本研究においては、微小な変化や水和構造の変化を考慮 することが必要になってくると考えて、2.0 Å近くのデータを得るために必要な結晶の大きさ を決定することにした。
Fig. 5 L indicates the length of crystal. t indicates the thickness of crystal
0.1mm
L
t
Fig.5 の L は結晶の長さ、t を結晶の厚みと考えて、結晶の大きさと分解能の関係を Table1.に 示した。その結果から、A 結晶より大きければ、2.5 Åよりもより高分解能のデータを得るこ とが可能で、B 結晶程度の大きさで高分解な値がより多く、データとして現れてくることが わかる。タンパク質の構造の変化を考えていく際に、微小な変化も考慮していかなければい けないことや水和に関することを議論していくためにも0.3×0.3×0.2 mm3以上の結晶を作製 することができればいいのではないかと考える。Table1. Size of crystal and resolution
L (mm) t (mm)
Resolution (Å)
A 0.13
0.08 2.5
B 0.28
0.15 2.1
C 0.33
0.16 2.1
D 0.44
0.22 2.0
3.3 グルコースイソメラーゼの三次元分子構造と水和構造の比較Crystal Clear(Rigaku)や HKL package17) というソフトウェアで X 線結晶構造解析の回折デー
タを取得して、データの処理を行う。そして、構造解析はCCP4 package18)のソフトウェアを 用いて行った。得られた回折データのセットを使い、分子置換法によって位相を決定し、構 造精密化を行った。 まず、タンパク質の分子構造自身については、0.1 MPa と 100 MPa 中で結晶育成したものの 0.1 MPa 中での構造には明確な変化はなかった(Fig.6 a, b)。二つの構造を最小二乗法による重 ね合わせで、対応する全原子間距離を求めてみると、最大で約1.37 Åの位置の違いで、平均 として約 0.078 Åであり、あまり大きな変化ではない。これまでの研究において、私達とほ ぼ同じような形で高圧力下で結晶育成して常圧下でX 線結晶構造解析を行っているソーマチ ンにおいても主鎖の位置の変化の平均が0.134Å、その他の原子では 0.140Åとなっている13)。 著者らは、それぞれ微小な変化であり、明確な変化ではないと述べている。また、逆に常圧 下で作った結晶を高圧力下で構造解析を行った結果では、ニワトリ卵白リゾチームで全原子 の位置の変化の平均が0.2 Åであった10,11)。最も大きく変化しているところでは、約1 Åにな るとされているが、全体としては明確な構造の変化はないと考えられる。ただし、このよう な微小な変化が実は大きな機能変化をもたらすかも知れないという可能性についてはさらな る研究が必要となる。 次に、分子構造自身だけでなく、高圧力下での水和構造に関する変化についても考えなけ ればならない。タンパク質分子が変化する可能性があるなら、それに伴う水和構造も同時に
変化するはずである。今回のデータから、構造解析によって決定した水分子の数と位置に関 する比較を行った(Fig.6 c, d)。今回の構造解析においては、水分子の数においては、0.1 MPa と100 MPa の値を比較したときに、回折データの比較するものによっては変わらなかったり、 多かったりと有意差がほとんどないと考えられる。その位置に関しても、0.1 MPa 同士を比べ たときの位置の違いの割合と0.1 MPa と 100 MPa で比べたときの位置の違いの割合も約 10 % 程度でほとんど変わらないので、これも有意差があると考える事は難しい。これまでの研究 では構造のデータを平均で出していると考えられる。ニワトリ卵白リゾチームでは、常圧下 (水分子 151 個)から比べて高圧下(水分子 163 個)での構造で 12 個の水分子の増加と水分子 とタンパク質分子の距離の違いなどについて議論している 10,11)。それらの結果を見ると、わ ずかに分布の違いがあることがわかる。また、ソーマチンでは49 個の水分子が常圧下で育成 した結晶の構造(282 個)から高圧力下で育成したものの構造(233 個)で減少している。その位 置の違いは、高圧力下での水分子の位置が常圧下と比べて違うところが約9 %とされている 13)。この水和構造の評価は、非常に難しいと考えられる。その水分子の位置決定は、構造解 析のソフトウェアの計算によって算出しているために、回折データのセットによって変化す るとされている 19)。更に、X 線結晶構造解析においては、水分子のことを議論していこうと すると分解能の高い値が必要となってくるだろう。そのために、今後の研究結果において、 水和構造の変化に関しては十分な注意が必要であるが、0.1MPa と 100MPa 下における X 線結 晶構造解析の回折データに現れてくるかもしれない違いを議論していくことはできるものと 考えられる。 c) d) b) a)
Fig. 6 Comparison of ball and stick model structure at 0.1 MPa(a) and 100 MPa(b) Comparison of Hydration structure at 0.1 MPa(c) and 100 MPa(d)
4. まとめ 本研究は、その場高圧力下X 線結晶構造解析を実施して、高圧力下でのタンパク質の構造 を直接決定する準備段階である。得られた研究結果を以下にまとめる。 ① ガラスキャピラリー中で、0.1 MPa と 100 MPa 下で結晶化の制御方法が確立できた。 ② ガラスキャピラリー中で溶液が入ったままで高分解能の構造解析が可能なことが確認で きた。 ③ 結晶サイズと溶液共存下での分解能の相関を決定した結果、0.3×0.3×0.2mm3以上の結晶が 必要であることがわかった。 ④ 今回の常圧下での X 線結晶構造解析において、得られた分子構造及び水和構造に明確な 変化はなかった。 しかし、今回は、すべての過程を高圧力下で行ってはいないために、高圧力下での構造を 維持していないと考えられる。今後の研究におけるその場高圧力下での実験の結果によって 最終的な判断を必要とするだろう。また、微小な構造の変化に対する評価についても考えて いく必要があるだろう。 謝辞 本研究の一部は、文部科学省科学研究費補助金若手研究(B)課題番号 16760014 及び 19760009 を使用して行いました。 参考文献 [1] 巻本彰一, 谷口吉弘, (1989) 酵素反応への圧力効果 蛋白質 核酸 酵素. 34, 105-112
[2] Gross, M. and Jaenicke, R. (1994) Proteins under pressure. The influence of high hydrostatic pressure on structure, function and assembly of proteins and protein complexes. Eur. J. Biochem. 221, 617-630
[3] Makimoto, S., Wada, A. and Y. Taniguchi, (1996) Thermal and Pressure Inactivation of Subtilisin BPN. J. Soc. Mat. Sci. Japan. 45, 274-279
[4] Mozhaev, V. V., Lange, R. and Kudryashova, E. V. (1996) Application of High Hydrostatic Pressure for Increasing Activity and Stability of Enzymes. Biotechnology and Bioengineering, 52, No2, 320-331
[5] Northrop, D. B. (2002) Effects of high pressure on enzymatic activity. Biochimica et Biophysica Acta 1595, 71-79
[6] 谷口吉弘, (1989) 蛋白質の高圧変性. 蛋白質 核酸 酵素. 34, 98-104
[7] Akasaka, K. (2003) Highly Fluctuating Protein Structures Revealed by Variable-Pressure Nuclear Magnetic Resonance. Biochem. 42, 10875-10884
[8] 月向邦彦, (2004) Effect of Hydration on the Volume and Compressibility of Protein Molecules. Netsu Sokutei. 31, 186-193
[9] 月向邦彦, (1996) 硬い蛋白質と軟らかい蛋白質 ‐圧縮率から見た構造のゆらぎ‐. 蛋白質 核酸 酵素. 41, 2025-2036
[10] Kundrot, C.E. and Richards, F.M. (1987) Effect of hydrostatic pressure on the solvent in crystals of hen egg-white lysozyme. J. Mol. Biol. 193, 157-170
[11] Kundrot, C.E. and Richards, F.M. (1988) Effect of hydrostatic pressure on the solvent in crystals of hen egg-white lysozyme. J. Mol. Biol. 200, 401-410
[12] Collins, M.D., Hummer, G., Quillin, M.L., Matthews, B.W. and Gruner, S.M. (2007) Structural Rigidity of a Large Cavity-containing Protein Revealed by High-pressure Crystallography. J. Mol. Biol. 367, 752-763
[13] Charron, C., Robert, M.C., Capelle, B., Kadri, A., Jenner, G., Giege, R. and Lorber, B. (2002) X-ray diffraction properties of protein crystals prepared in agarose gel under hydrostatic pressure. J.
Crystal Growth. 245, 321-333
[14] Visuri, K., Kaipainen, E., Kivimaki, J., Niemi, H., Leisola, M. and Palossari, S. (1990) A New Method for Protein Crystallization Using High Pressure. Bio/Technology 8, 547-549
[15] Suzuki, Y., Sazaki, G., Matsui, T., Nakajima, K. and Tamura, K. (2005) High-Pressure Acceleration of the Growth Kinetics of Glucose Isomerase Crystals. J. Phys. Chem. B. 109, 3222-3226
[16] 後藤祐児, 桑島邦博, 谷澤克行 [編], タンパク質科学 構造・物性・機能, 化学同人, 2005, 36-37 [17] Otwinowski, Z. and Minor, W. (1997) Processing of X-ray diffraction data collected in oscillation
mode. Methods Enzymol. 276, 307-326
[18] Collaborative Computational Project, (2004) Number 4, Acta Crystallogr. D 50 760-763
X-ray structure analysis of glucose isomerase molecules
in crystals prepared under high pressure
Masayuki Tsukamoto
1, Yoshihisa Suzuki
*2, Haruhiko Sakuraba
3Katsuhiro Tamura
21
Department of Chemical Science and Technology, The University of Tokushima, 2-1 Minamijosanjima, Tokushima 770-8506, Japan
2
Department of Life System Institute and Technology and Science, The University of Tokushima, 2-1 Minamijosanjima, Tokushima 770-8506, Japan
3
Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, 2393 Ikedo, Miki-cho, Kida-gun, Kagawa 761-0795, Japan
Proceedings of the 15th Symposium for Japanese Research Group of High Pressure Bioscience and Biotechnology
Received 27 February 2008/Accepted 5 March 2008
Abstract
Three-dimensional structures of glucose isomerase molecules in crystals prepared from supersaturated solutions in capillaries under 0.1 and 100 MPa were determined by X-ray crystallography under 0.1 MPa. With accurate temperature control, we obtained a few large crystals of good quality in the capillaries under 0.1 and 100 MPa. X-ray structures with high quality data sets were successfully obtained without removing solution around the crystals. We also determined a relation between resolution of molecular structure and size of the crystals surrounded by the solution in the capillaries. However, the r.m.s. deviation of least square fit of all atom positions of both glucose isomerase folds was about 0.078Å. This deviation was little. The distributions of water sites in both glucose isomerase structures were almost identical.
Keywords:capillary, X-ray crystallography, Three-dimensional structure, hydration, crystallization control