報告
2014年5月に駿河湾で採集されたセンジュエビ類後期幼生の
DNA分析による種判別
Species identification of Polychelid postlarvae collected in Suruga Bay
on May, 2014 by DNA analysis
柳本 卓
1・小西光一
1・髙見宗広
2Takashi Yanagimoto, Kooichi Konishi, and Munehiro Takami
はじめに センジュエビ科は,第1から4歩脚,種によって は第5歩脚までその先端に鋏を持つことから,千手 観音にちなんで「センジュエビ」という名前が付け られている.センジュエビ科は世界中の中~漸深層 に分布し,6属40種以上存在している (Galil, 2000). 日本周辺には8種類が記録されている(山本・長澤, 2011).センジュエビ科の複眼は退化し,角膜など の痕跡もない.センジュエビ類は中生代から新生 代の初期にかけて浅海に生息していたエリオン科 (Eryonidae) に似た形態を持っていることから,この グループから深海へと適応拡散した(蒲生,1980) 生きた化石として考えられている.そのため,セン ジュエビ科は甲殻類の系統進化を調べる上で重要で あり,様々な先行研究が行われている(Schram & Dixon, 2004; Boisselier-Dubayle et al., 2010; Bracken-Grissom et al., 2014など).一方,センジュエビ科は 関東以南の太平洋沿岸の深海域に広く分布している が (馬場,1986),まとまった漁獲量がないので漁業 価値がなく,調査研究はほとんど行われていない. しかし,近年,主要漁獲対象種のみばかりでなく, 生態系管理という立場から,漁獲対処種以外につい ての正確な種判別と生態の把握が重要となってきて いる. センジュエビ科の幼生に関する情報はほとんどな く,地中海西部で採集された幼生について外部形態 とDNA分析からセンジュエビ(Polycheles typhlops) の幼生であると報告され(Torres et al., 2014),日本 周辺では福島沖と駿河湾で採集されたセンジュエビ 類の幼生についてDNA分析による種判別を行い, これらがセンジュエビ類のPentacheles laevisの幼生 であることが報告されている (柳本ら,2015).本研 究では,2014年5月に駿河湾で採集されたセンジュ エビ類の幼生 (Fig. 1) について,先行研究と同様に DNA分析により種判別を行い,何種であるか検討 した. 材料と方法 本研究で用いた後期幼生は東海大学海洋学部所属 の実習船北斗によって2014年5月に駿河湾から口径 1.6 mリ ン グ ネ ッ ト(目 合 い2 mm, 1 mm, 0.5 mm) (Takami & Fukui, 2010)) によって採集された.得ら れた標本は写真を撮影した後 (Fig. 1),70%エタノー ルで固定した.
幼生の腹節あるいは胸脚から筋肉片約10 mgを摘
1 国立研究法人 水産研究・教育機構 中央水産研究所
〒236–8648 神奈川県横浜市金沢区福浦2–12–4 National Research Institute of Fisheries Science, Fisheries
Research and Education Agency, 2–12–4 Fukuura, Kanazawa, Yokohama, Kanagawa 236–8648, Japan E-mail: [email protected]
2 東海大学海洋学部
〒424–8610 静岡県静岡市清水区折戸3–20–1 Tokai University, School of Marine Science and
Technol-ogy, 3–20–1 Orido, Shimizu, Shizuoka, Shizuoka 424– 8610, Japan
出してQuickGene (Fuji Co.) を用いてDNAを抽出し た.抽出したDNAをテンプレートにして,PCR法に よ りmtDNAのCOI領 域 と 核DNAのITS1を 含 む 領 域(以下,ITS1領域と略記する)の増幅をおこなっ た.COI領域の増幅にはユニバーサルプライマーの LCO1490 (5′-GGTCAACAAATCATAAAGATATTGG- 3′) とHCO2198 (5′-TAAACTTCAGGGTGACCAAAA AATCA-3′)を用いた(Folmer et al., 1994).ITS1領域
の増幅にはユニバーサルプライマーのMD-1(5′-
cttgactatctagaggagt-3′) と5.8 SH (5′-agcttgcgttcttcatcga- 3′)を用いた(Sajdax & Phillips, 1997).サーマルサ イクラーとしてABI9700(Applied Biosystems)を用 いた.PCR反応は,最初に94℃2分加熱し,その後, 94℃30秒,55℃30秒,72℃2分を30サイクル行い, 最後に72℃で7分加熱した.PCR反応溶液は,粗 DNA溶 液1 μl, 2.5 mM dNTP溶 液2.5 μl, 10×Buffer (TaKaRa) 2.5 μl, 50 μMの各プライマー0.5 μl, TaKaRa
Ex Taq Polymerase 0.125U,総量が25 μlになるように
超純水を加えたものである.PCR産物を1.5%アガ
ロースゲル (NuSieve3:1, TaKaRa) で電気泳動し,エ チジウムブロマイド染色によって増幅産物の確認を した.増幅産物をQuick PCR Purification Kit (Qiagen)
により精製した.COI領域について,精製した増幅
産物をテンプレートとして,BigDye Terminator Kit v3.1(Applied Biosystems)を用いてシーケンス反応
を行った.シーケンス反応には,PCRで用いたのと
同じプライマーを用いた.シーケンス反応物をエタ ノールで精製した後,ABI PRISM 3730XL Genetic Analyzer(Applied Biosystems)で電気泳動して塩基
配列を決定した.核DNAのITS1領域の増幅産物を
TOPO TA Cloning kit (Invitrogen) によりTAクローニ ングした.形質転換した大腸菌からM13 (5′-gtaaaa cgacggccag-3′) とM13R (5′-caggaaacagctatagac-3′) の プライマーを用いて,コロニーPCRを行った.PCR の反応条件等は上記と同様である.精製したPCR産 物について,上記したCOI領域と同様にシーケンス 反応を行い,塩基配列を決定した. 決定した塩基配列を用いて,インターネット上の 遺伝子相同性検索ソフトBlast (Altschul et al., 1990) により,本サンプルの塩基配列がどのような種と近 いか調べた.また,データベース上から近縁種の塩 基配列を集め,Clustal W (Thompson et al., 1994) によ
りアライメントを行った.また,MEGA5.05ソフト
ウ エ ア (Tamura et al., 2011) により,Kimura’s two- parameter model (Kimura, 1980)の遺伝距離を算出し, 近隣結合法(Saitoh & Nei, 1987)により系統樹を作成
した.系統樹の信頼性は1,000回のブーストラップ 検 定(Felsenstein, 1985)に よ り 評 価 し た. ま た, MEGA5.05ソフトウエアにより,種間の塩基置換数 や 塩 基 置 換 率 を 調 べた.幼 生 の 塩 基 配 列はDNA データベースにLC076645–LC076650で登録した. 結 果 幼生のITS1領域の塩基配列を5つの配列を得るこ
Fig. 1. Photograph of eryoneicus postlarvae collected from Suruga Bay on July 2014. A) Dorsal view, B) lateral view, C) frontal view.
Table 1. Frequencies (%) of variable sites on ITSI region among species of Polychelidae.
Sample and
species name Abbreviation
Sample and speices name Larvae PT HA PL polychelid postlarvae Larvae
Polycheles typhlops PT 19.6
Homeryon armarium HA 23.0 22.2
とができた.5つの平均塩基置換数は0.4塩基で,平
均塩基置換率は0.1%であった.この塩基配列につ
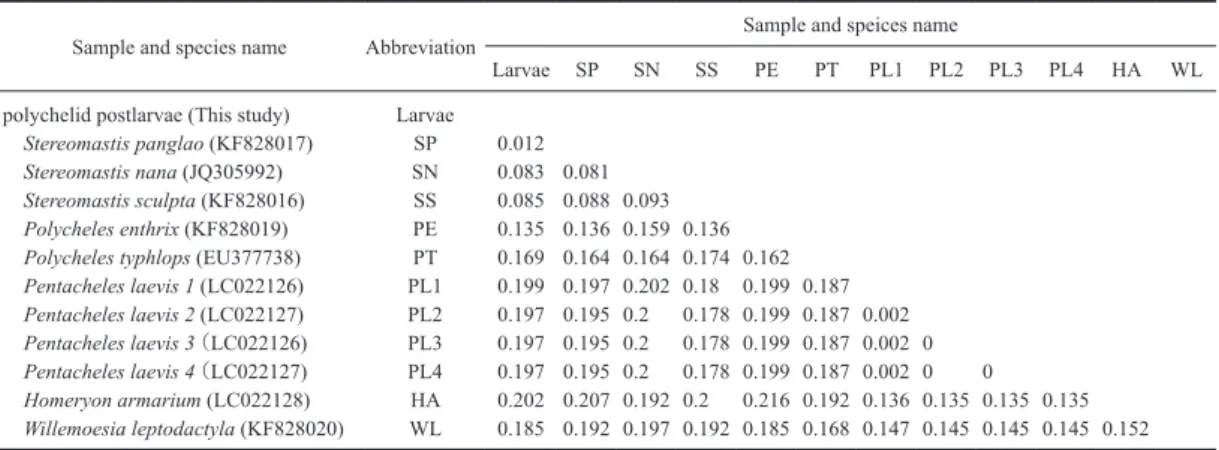
いて,Blast分析を行ったところ,Pentacheles laevis, Homeryon armarium, センジュエビPolycheles typhlops の塩基配列と相同性が高かった (柳本ら,2015).そ れら以外では,ジャノメガザミのITS1の塩基配列 (AM410544)と相同性が高かったが,一致率は非 常に低かった.得られたITS1の塩基配列と相同性 の高かった上記3種のデータを比較すると,平均塩 基置換数は88~104塩基で,平均塩基置換率は19.6 ~23%であった.塩基置換率が高く,本幼生の親種 のITS1の塩基配列は,既存のデータにはなかった. Pentacheles laevis(LC022102–LC022113, LC022126– LC022127),Homeryon armarium (LC022114–LC022122), Polycheles typhlops (LC026058–LC026068)とそれら 以外で一番相同性が高かったジャノメガザミの塩基 配列 (AM410544) を外群として,系統樹を作成した (Fig. 2).3種はそれぞれクレードを形成したが,幼 生の配列は幼生だけでクレードを形成し,他種と塩 基配列が明らかに異なっていることが分かった.し かし,幼生の塩基配列はPentacheles laevisやHomeryon armariumよりもセンジュエビPolycheles typhlopsの クレードに近かった. 幼生のCOI領域の塩基配列について,Blast分析を 行ったところ,一致率の高かった上位種はStereomastis panglao (98%) (KF828017),Stereomastis sculpta (92%) (KF828016),Stereomastis nana(92%)(JQ305992) などのStereomastis種であった.相同性の高かった 上位種と外群としてセンジュエビ類以外で一番相同 性の高かったコシオリエビの一種(AY288292)(Lee & Kim, 2013)の塩基配列を用いて,系統樹を作成し たところ(Fig. 3),幼生の塩基配列はStereomastis panglao(KF828017)の塩基配列と高い信頼度で一 つのクレードを形成した.続いて,ヒメセンジュエ ビStereomastis nana (JQ305992),Stereomastis sculpta (KF828016) が順にクレードに加わるという形であっ た.塩基置換率はStereomastis panglaoと1.2%の違 いで,他種とは8.3%以上異なっていた(Table 2). Fig. 2. Neighbor-joining tree of the sequences of
ITSI region constructed using Kimura two-parameter distances for polychelid species and the postlarvae. Numbers beside internal branches indicates bootstrap probabilities based on 1,000 pseudoreplicates. Bootstrap probabilities higher than 50% are shown.
Fig. 3. Neighbor-joining tree of the sequences of COI region constructed using Kimura two-parameter distances for polychelid species and the postlarvae. First and second numbers after species name indicate individual and clone number, respectively. Numbers beside internal branches indicates bootstrap probabilities based on 1,000 pseudoreplicates. Bootstrap probabilities higher than 50% are shown.
このことからも,本幼生はStereomastis panglaoであ ると推測された. 考 察 本分析で2014年5月に採集されたセンジュエビ類 の幼生は核DNAのITS1の塩基配列からセンジュエ ビ科であることが分かった(Fig. 2).mtDNAのCOI の塩基配列からStereomastis属に属し,Stereomastis panglaoであると考えられた(Fig. 3, Table 2).ただ し,本研究ではStereomastis panglaoの成体サンプル を用いたのではなく,データベースに登録している 塩基配列を用いている.そのため,今後,成体を入 手して再検討する必要がある.Ahyong & Chan (2008) によると,Stereomastis panglaoはボホール海とスル 海の水深784–1773 mに限定的に分布していると報 告しているが,本幼生が駿河湾で採集されたことか ら,本種の成体の駿河湾において分布している可能 性が考えられた.一方,Stereomastis panglaoの幼生 期の生態について明らかになっていないが,南の海 域に生息するStereomastis panglaoの幼生がウナギの シラス幼生のように,北赤道海流から黒潮に乗って 駿河湾まで移送された可能性も否定できない. 本報告で使用した駿河湾で採集されたセンジュエ ビ幼生は,柳本ら(2015)で用いている駿河湾のセ ンジュエビ科の幼生と同時に採集されていた.当 初,柳本ら(2015)が用いている駿河湾と福島沖で 採集されたセンジュエビ幼生はPentacheles laevisで あったことから,本報告で用いた幼生も同一種と考 えられていた.しかし,本解析で明らかになったよ うに,本幼生はStereomastis panglaoであると考えら れた.センジュエビ類の幼生の分布が単純でないこ とが推測される.今後,よりサンプル数を増やし て,網羅的に検討していく必要がある. 謝 辞 駿河湾で採集されたセンジュエビ科幼生の採集に は,東海大学海洋学部水産学科教授福井 篤氏,東 海大学海洋学部所属小型舟艇北斗の乗組員各位にご 協力して頂いた.この場を借りて厚くお礼申し上げ る.本研究の一部は,平成27年度東京大学大気海 洋研究所学際連携研究費で実施された. 文 献
Ahyong, S. T., & Chan, T-Y., 2008. Polychelidae from the Bo-hol and Sulu Seas collected by Panglao 2005 (Crusta-cea: Decapoda: Polychelidae). The Raffles Bulletin of Zoology, Supplement, 19: 63–70.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipan, D. J., 1990. Basic local alignment search tool. Journal of Molecular Biology, 215: 403–410.
馬場敬次,1986. センジュエビ科.日本陸棚周辺の十脚
甲殻類(馬場敬次・林健一・通山正弘編),社団
法人日本水産資源保護協会編,東京.pp. 156–159. Boisselier-Dubayle, M. C., Bonillo, C., Cruaud, C., Couloux,
A., Richer de Forges, B., & Vidal, N., 2010. The phylo-genetic position of the ‘living fossils’ Neoglyphea and Laurentaeglyphea (Decapoda: Glypheidea). Comptes Rendus Biologies, 333: 755–759.
Table 2. Frequencies (%) of variable sites on COI region among species of Polychelidae.
Sample and species name Abbreviation Sample and speices name
Larvae SP SN SS PE PT PL1 PL2 PL3 PL4 HA WL
polychelid postlarvae (This study) Larvae
Stereomastis panglao (KF828017) SP 0.012
Stereomastis nana (JQ305992) SN 0.083 0.081
Stereomastis sculpta (KF828016) SS 0.085 0.088 0.093
Polycheles enthrix (KF828019) PE 0.135 0.136 0.159 0.136
Polycheles typhlops (EU377738) PT 0.169 0.164 0.164 0.174 0.162
Pentacheles laevis 1 (LC022126) PL1 0.199 0.197 0.202 0.18 0.199 0.187 Pentacheles laevis 2 (LC022127) PL2 0.197 0.195 0.2 0.178 0.199 0.187 0.002 Pentacheles laevis 3 (LC022126) PL3 0.197 0.195 0.2 0.178 0.199 0.187 0.002 0 Pentacheles laevis 4 (LC022127) PL4 0.197 0.195 0.2 0.178 0.199 0.187 0.002 0 0 Homeryon armarium (LC022128) HA 0.202 0.207 0.192 0.2 0.216 0.192 0.136 0.135 0.135 0.135 Willemoesia leptodactyla (KF828020) WL 0.185 0.192 0.197 0.192 0.185 0.168 0.147 0.145 0.145 0.145 0.152
Bracken-Grissom, H. D., Ahyong, S. T., Wilkinton, R. D., Feldmann, R. M., Schweitzer, C. E., Breinholt, J. W., Bendall, M., Palero, F., Chan, T. Y., Felder, D. L., Ro-bles, R., Chu, K.-H., Tsang, L.-M., Kim, D., Martin, J. W., & Crandall, K., 2014. The emergence of Lobsters: Phylogenetic relationships, morphological evolution and divergence time comparisons of an ancient group (Decapoda: Achelata, Astacidea, Glypheidea, Polyche-lida). Systematic Biology, 63: 457–479.
Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783–791. Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrigenhoek,
R., 1994. DNA primers for amplification of mitochon-drial cytochrome c oxidase subunit I from diverse meta-zoan invertebrates. Molecular Marine Biology and Bio-technology, 3: 294–299.
Galil, B. S., 2000. Crustacea Decapoda: Review of the gen-era and species of the family Polychelidae Wood-Ma-son, 1874. Memoires du Museum national d’Histoire naturelle, 184: 285–387.
蒲生重男,1980.深海の甲殻類.海洋と生物,2: 88–98. Kimura, M., 1980. A simple method for estimating evolu-tionary rate of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, 16: 111–120.
Lee, S., & Kim, W., 2013. First reports of two species of Ge-nus Raymunida (Crustacea: Decapoda: Anomura) from Korea. Animal Systematics Evolution and Diversity, 29(4): 279–287.
Saitou, N., & Nei, M., 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, 4: 406–425. Sajdax, S. L., & Phillips, R. B., 1997. Phylogenetic
relation-ships among Coregonus species inferred from the DNA sequences of the first internal transcribed spacer (ITS1) of ribosomal DNA. Canadian Journal of Fisheries Aquatic Science, 54: 1494–1503.
Schram, F. R., & Dixon, C. J., 2004. Decapoda phylogeny: addition of fossil evidence to a robust morphological cladistic data set. Bulletin of the Mizunami Fossil Mu-seum, 31: 1–19.
Takami, M., & Fukui, A., 2010. Larvae and juveniles of Lep-toderma lubricum and L. retropinnum (Argentiformes: Alepocephalidae) collected from Suruga Bay, Japan. Ichthyological Research, 57: 406–415.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., & Kumar, S., 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolution-ary distance, and maximum parsimony methods. Mo-lecular Biology and Evolution, 28: 2731–2739. Thompson, J. D., Higgins, D. G. & Gibson, T. J., 1994.
CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weight-ing, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22: 4673–4680. Torres, A. P., Palero, F., Dos Santos, A., Blanco, P. A.-E., &
Guerao, A. B.-G., 2014. Laraval stages of the deep-sea lobster Polycheles typhlops (Decapoda, Polychelida) identified by DNA analysis: morphology, systematic, distribution and ecology. Helgoland Marine Research, 68: 379–397. 山本圭介・長澤和也,2011. 東シナ海大陸斜面域で採集 されたエビ類.日本生物地理学会会報,66: 27–39. 柳本 卓・小西光一・髙見宗広・猿渡敏郎,2015. 本 州太平洋沿岸で採集されたセンジュエビ科の後期 幼生のDNA種判別.Cancer, 24: 7–13.