プロイディゲーム 染色体セットの数と組み合わせの変化は魚介類に何をもたらすか
10
0
0
全文
(2) 198. イツにおいて両生類を材料に始まり,魚類では 1950 年. カワの三倍体では,雌は卵巣が未発達で,雄は少量の異. 代の旧ソ連, 1960 年代の英国において研究が行われ. 数体精子しか産生せず不妊と判断されたが,成長は二倍. た 。4,5). そ し て , 1970 ~ 1980 年 代 に ア メ リ カ の Thor-. 体に劣った。20) 一方,巻貝(腹足類)のフクトコブシで. gaard , フ ラ ン ス の Chourrout , 日 本 の 小 野 里 ら に よ. は三倍体の体重,殻長は二倍体と差がなかったが,可食. り,サケ科魚類における操作原理の解明と技術開発,さ. 部の比率が高かった。15)貝類の三倍体は魚類と異なり,. らに養殖性能評価が大きく進展した。15) これらの成果. 良好な養殖性能を示すと思われる。4,5). を受け,国内では全国自治体によりご当地養殖魚介類へ. 三倍体および雌性発生二倍体では第二極体放出阻止に. の応用研究が進み,三倍体マガキ(かき小町,広島県),. より 誘起 さ れる た め, 四分 子分 析 half-tetrad analysis. 全雌ヒラメ(兵庫県,鳥取県等),雑種サーモン(絹姫. による DNA マーカーあるいは遺伝子座と染色体動原体. サーモン,愛知県信州サーモン,長野県等)などの優. 間の遺伝地図作成の格好の材料となった。4,5,16,21,24,27) こ. れた成果が見られた。4). れらの分析により,染色体のテロメア(端部)近くに存 在するマーカー(遺伝子)座とセントロメア(動原体). 魚介類における染色体操作の応用. 近くに存在する座を明らかにすることができた。テロメ. 本稿のメイントピックスとなるサケ科とドジョウ科以. ア近くの座を利用することにより交差(組換え)の存否. 外の魚介類で著者らが共同研究により行った染色体操作. を確認できるので,雌性発生あるいは雄性発生倍加半数. 研究を表 1 に示す。1128) いずれの魚介類においても,初. 体が完全ホモ接合であることがマコガレイ,17) マツカ. 期は染色体倍加条件あるいは配偶子の遺伝的不活性化条. ワ,21) ゼブラフィッシュ28)で明らかにされた。さらに,. 件の検討であったが,1114,19,23,26). その後,研究の重点は. ニホンウナギ,24,25) ミカドチョウザメ27)では,マーカー. 養殖性能の評価に移った。成熟期以前の三倍体の成長は. (遺伝子)-動原体地図の情報を用いて,人工授精に由. 二倍体と同等あるいはそれに劣るが,成熟期では不妊性. 来する子孫に見られる自然三倍体が多精ではなく第二極. の発現により良好となるというのが魚類一般における評. 体放出の自然阻止により出現することが明らかにされ,. 価であった。4,5) しかし,成長は魚種により異なり,マツ. 卵の過熟(老化)との関係が示唆された。. 表1 種 エゾアワビ. フクトコブシ トラフグ. 魚介類の染色体操作による主要な成果. 染色体操作. 主. 要. な. 成. 果. 文献. 雌性発生半数体. 精子遺伝的不活性化の最適 UV 照射条件. 11. 三倍体. 極体放出阻止の最適低温高温圧力処理条件. 12. 雌性発生二倍体. 精子の最適 UV 照射条件,極体放出阻止の最適低温処理条件. 13. 三倍体. 極体放出阻止の最適低温処理条件. 14. 三倍体. 雌雄生殖腺発達不全,可食部. 3n>2n 精子遺伝的不活性化の最適 UV 照射条件. 15. 雌性発生二倍体. 16. 第二極体放出第一卵割阻止の最適温度処理条件 マコガレイ. 雌性発生二倍体倍加半数体. 遺伝子 動原体地図,XXXY 型推定 精子遺伝的不活性化の最適 UV 照射条件 第二極体放出第一卵割阻止の最適温度圧力処理条件. 17. 遺伝子 動原体地図,倍加半数体確認,XXXY 型推定 性転換雌性発生二倍体後代子孫 マツカワ. 雌性発生二倍体三倍体. XXXY 型決定,全雌集団作出 精子遺伝的不活性化の最適 UV 照射処理条件. 18 19. 第二極体放出第一卵割阻止の最適温度圧力処理条件 三倍体 雌性発生二倍体倍加半数体. XXXY 型推定,成長 ♀>♂ 第二極体放出阻止の最適高温処理条件. 22. 三倍体. マーカー 動原体地図. 24. 過熟による自然第二極体放出抑制の推定. 25. 精子遺伝的不活性化の最適 UV 照射処理条件. 26. (自然三倍体モザイク) ベステルチョウザメ. 20 21. 三倍体. 雌性発生二倍体 二ホンウナギ. 3n 不妊性,成長 2n>3n マーカー 動原体地図,倍加半数体確認. 雌性発生二倍体三倍体. 23. 第二極体放出阻止の最適高温処理条件,ZWZZ 型推定 ミカドチョウザメ ゼブラフィッシュ. (自然倍数体) 雄性発生倍加半数体クローン. マーカー 動原体地図,過熟による自然第二極体放出抑制の推定. 27. 低温処理による雄性発生,高温処理による倍加半数体作出. 28. 雄性発生による次世代クローン作出.
(3) 199. 現在までに性決定遺伝子が,メダカ,トラフグ,ニジ. るか)により,子孫が正常に発生を進め孵化生育する. マス等で明らかにされてきたが,形態学的に識別可能な. 場合と,胚発生異常を示し死亡する場合が見られる。6,7). 性染色体を持つ種を除き,多くの魚類において遺伝的性. 1970 年代までの雑種研究では,異種間授精により生じ. 決定機構は不明であった。しかし,人為的な雌性発生二. る子孫が遺伝的に「雑種」となっているか否かについて. 倍体では XX-XY 型の動物では全雌が, ZW-ZZ 型の動. は検討されておらず,水産学的に無価値とされる致死性. 物では雌(超雌 WW ,組換え型雌 ZW )と雄( ZZ )両. 雑種については全く関心がもたれなかった。6) 著者は雑. 者の出現が期待される。25,8,10). また,雄性発生二倍体で. 種胚の生存死亡の機構に関心をもち,大学院修士,博. は XX-XY 型の動物では雌(XX)と超雄(YY)の出現. 士課程における研究課題とした。現在多用されている分. が,ZW-ZZ 型の動物では雄(ZZ)のみの出現が期待さ. 子生物学的手法は全くない時代であったので,酵素の遺. れる。25,8,10) 従って,雌性発生あるいは雄性発生二倍体. 伝多型(アイソザイム,アロザイム)の種間差をマーカー. における性比から性決定機構を推定できる。25,8,10). トラ. として,様々なサケ科魚類の種間,属間の組合せで作出. フグ,16) マコガレイ,18) マツカワ22)では雌性発生二倍体. した生存性,致死性の雑種子孫の胚発生過程における雑. における全雌に近い性比から XX-XY 型が,ベステル. 種型酵素の発現時期を検討した。その結果,胚発生初期. チョウザメ26) では 70 近くの雌出現率から ZW-ZZ 型. は母親型(母系種の酵素)のみが見られたが,生存性雑. が推定された。ゼブラフィッシュ雄性発生倍加半数体お. 種では胚体形成が進むステージ前後から雑種型が認めら. よびそれを用いたクローン系では雄しか出現しなかった. れた。6) このことは,母系,父系の遺伝子の産物が雑種. ことから,本種の遺伝的性決定は基本的に ZW-ZZ 型と. 型のアロザイムを作ること,すなわち,雑種が両親由来. 推定された。28). の染色体セットからなる遺伝子型を持つことを示す。6) これに対して,発生過程で死んでしまう致死性雑種では 雑種型酵素が見られないものが多かった。6) 致死性雑種. サケ科魚類雑種の発生能力と染色体セット構成 人為交雑も染色体セットの組み合わせを変える手法で. で雑種型が生じない原因として,胚の遺伝子自体が働か. あることから,染色体操作の一部と考えられる。魚類で. ない生理的異常と,両親種由来染色体のいずれかが脱落. 人為交雑をすると,種の組合せと方向(母系種を何にす. して遺伝的な雑種になっていない構造的異常の両方が考. 表2. サケ科魚類の種間属間雑種の生存性,倍数性,染色体数および核型 核. 雌×雄. 倍加処理. 生存性. 倍数性. M/SM 両親種. 種間雑種. 属間雑種. カラフトマス. -. +. 二倍体. サクラマス. -. +. 二倍体. マスノスケ. -. +. 二倍体. シロザケ. -. +. 二倍体. アメマス. -. +. 二倍体. カワマス. -. +. 二倍体. サクラマス×カラフトマス. -. +. 異質二倍体. サクラマス×マスノスケ. -. +. 異質二倍体. マスノスケ×サクラマス. -. +. 異質二倍体. シロザケ×サクラマス. -. -. 異質二倍体. 型. 染色体数. ST. A. 2n=52 2n=66. 48. 4. 0. 34. 24. 8. 2n=68 2n=74. 32. 28. 8. 26. 6. 42. 2n=84 2n=84. 16. 2. 66. 16. 2. 66. 2n=59 2n=67. 41. 14. 4. 33. 26. 8. 2n=67 2n=70. 33. 26. 8. 30. 15. 25. 2n=70 2n=84. 30. 15. 25. 16. 2. 66. 2n=75 2n=75. 25. 13. 37. 25. 13. 37. 2n=79 3n=116. 21. 4. 54. サクラマス×シロザケ. -. -. 異質二倍体. アメマス×カワマス. -. +. 異質二倍体. サクラマス×アメマス. -. +. 異質二倍体. アメマス ×サクラマス. -. +. 異質二倍体. シロザケ×アメマス. -. -. 異質二倍体. +. +. 異質三倍体. アメマス×シロザケ. -. -. 低二倍性異数体. 34 7 75 細胞の 75 が染色体数 50 65 に分布. +. -. 低三倍体異数体. 細胞の 71 が染色体数 75 100 に分布. -. -. 異質二倍体. +. +. 異質三倍体. -. -. 低二倍性異数体. シロザケ×カワマス カワマス×シロザケ. 原著ではイワナ Japanese charr と表記, M中部着糸型染色体,SM次中部着糸型染色体,ST次端部着糸型染色体,A端部着糸型染色体. 2n=79 3n=116. 21. 4. 54. 34 7 75 細胞の74 が染色体数 5065 に分布.
(4) 200. えられた。そこで,胚の染色体を徹底的に観察すること. る。例えば,コイやキンギョは,他のコイ科魚類の 2. で,雑種が両親種の染色体を持つかどうか,すなわち構. 倍の染色体数 100 を持つが,核型としては二倍体( 2n. 造的異常の有無を確かめた。結果を表 2 にまとめる. = 100 )であり, 2 セットの染色体をもつ。従って,人. が,生存性雑種は両親種の中間の染色体数と核型をもつ. 為的に紫外線照射精子を授精して雌性発生を誘起した場. ことから,両親種由来ゲノムを持つ異質二倍体であっ. 合,子孫は卵に由来する 1 セットの染色体( 1n = 50 ). た。6,7) 一方,致死性雑種には異質二倍体と低二倍性異数. しか持たず,半数体症候群を示し死亡する。ところが,. 体の両方が見られた。6,7). 前者は遺伝子発現の異常によ. 染色体数 100 のドジョウの精子を用いて,人為雌性発. り,後者は胚発生過程における染色体削減により遺伝子. 生を誘起すると,子孫は半数体症候群を示すことなく,. 自体が欠落し,異常が生じるものと思われた。6,7). 正常な形態を持つドジョウとして発生した(表 3)。810). 雑種では,母系と父系のゲノム間の不適合性のみなら. 人為雄性発生を行った場合も同様の結果が得られた(表. ず,母系細胞質と父系核ゲノム間の不適合性も発生能力. 3)。810) このことは染色体数 100 のドジョウは 4 セット. に影響する。そこで,染色体操作を用いて,雑種におけ. の染色体をもつ四倍体(4n=100)であり,産生する配. 2)。6,7). 偶 子 は 2 セ ッ ト の 染 色 体 を も つ 2n で あ る こ と を 示. る母系ゲノムと父系ゲノムの比を変えてみた(表. その結果,異数体とはならない致死性雑種の場合,第二. す。810). 極体放出を阻止して母系ゲノム 2 セットと父系ゲノム 1. 遺伝的に四倍体であることが確認できたので,これら. セットをもつ異質三倍体とすると,生存性が回復する例. の雌雄の産生する配偶子は 2n である。そこで,2n 配偶. が見られた。一方,交雑により染色体異常が生じて異数. 子の交配により四倍体家系(4n=100)を維持するとと. 体となる雑種では母系染色体セットの倍加による生存性. もに,野生型二倍体と四倍体間の正逆交雑とその後の染. 回復効果は見られなかった。6,7) 現在まで,異質三倍体化. 色体操作(第二極体放出阻止)により三倍体(3n=75),. による生存性回復効果の分子機構は明らかにされていな. 五倍体( 5n = 125)を作出した(表 3)。810) 六倍体(6n. いが,それまで雑種が作れなかった交雑の組合せにおい. =150)は四倍体同士の交配の後,第二極体放出を阻止. ても,子孫を作り,養殖品種作成に供せるようになった. して誘起した。これら六倍体雌雄は成熟して,減数分裂. ことは応用上の大きな成果であった。. により受精能力を持つ 3n 配偶子を産生した。そこで, 六倍体と四倍体あるいは二倍体との交配により,五倍. 自然四倍体とそれに派生する高次倍数体の特性―ドジョ. 体,四倍体をそれぞれ作出した。810) 以上の様に,倍数. ウをモデルとして. 体間の交配と染色体操作を駆使することで,様々な染色. もし, 2n 配偶子を産生する同質四倍体が通常授精の. 体セットを持つ子孫をゲームのように作り出すことがで. 後の卵割阻止で作出できれば,異質四倍体や父系染色体. きる。810) 以上の成果を発表した国際学会では,まるで. を 2 セット持つ異質三倍体などを作出して,染色体. 倍 数 体 を 作 り 出 す ゲ ー ム ( プ ロ イ デ ィ ゲ ー ム Ploidy. セットと生存能力の関係解明に供することができる。し. Game)のようだとしばしば揶揄された。. かしながら,サケ科では同質四倍体の作出が最難関で あった。4,5). 作出した各種の倍数体の特性を表 3 に要約する。810). かなり詳細に処理条件を検討し,処理群の発. 二倍体と四倍体の性比はほぼ 11 であった。野生型二. 生胚の中に四倍体を確認しても,孵化仔魚は多く浮腫な. 倍体ドジョウでは XX-XY 型の性決定システムが雌性発. どを持ち,生存率は極めて低かった。4,5). また,生残魚の. 生二倍体の性比から推定されている。四倍体が雌. な か に は 二 倍 体 ― 四 倍 体 モ ザ イ ク が 高 頻 度 で出 現 し. XXXX-雄 XXYY とすると,XX, XY, YY 精子が産生さ. た。4,5). 生殖細胞が四倍体となったモザイク魚が 2n 卵を. れ,次世代では雄(XXXY, XXYY)が多くなってしま. 産生する事例もわずかに認められたが,血液中に四倍体. うことが考えられる。しかし,四倍体の性比は 1 1. 細胞が少ないモザイク魚は二倍体と判定され廃棄される. で,四倍体から作出した六倍体の性比も 1 1 であっ. 場合も多かった。4,5) フランスの INRA,本邦の長野県,. た。さらに,人為的に誘起した雌性発生四倍体( 2n 卵. 静岡県では四倍体作出に成功したが,著者の人為四倍体. を雌性発生させた後に,第二極体放出阻止)が全雌であ. を用いた染色体操作研究は思うように進まなかった。. ることをみると(表 3),二倍体と同様に安定した性決. しかし,市販品ドジョウ(後日,中国起源が判明)の. 定をしていることが考えられる。しかしながら,ドジョ. 中に,偶々自然四倍体を見出したことから,この四倍体. ウ倍数体の性決定の詳細は他の倍数性動物と同様に目下. 個体を材料として,課題解決にチャレンジした。810). のところ不明である。. 日. 本 の ド ジ ョ ウ Misgurnus anguillicaudatus は 2n = 50 の. 偶数の倍数体である四倍体および六倍体はいずれも減. 野生型二倍体がほとんどであるが,市販品に見出した四. 数分裂により 2n および 3n 配偶子を各々産生し,雌雄. 倍体(と思われる個体)は染色体数 100 を示した。四. ともに妊性を示した。四倍体の産生する 2n 卵,六倍体. 倍体かどうかは 4 セットの染色体をもつことが鍵にな. の産生する 3n 卵のサイズは野生型二倍体の産生する 1n.
(5) 201 表3. ドジョウにおける多様な倍数体の誘起とその特性 性比. 雌. 雌雄. 妊性. 倍数性. 卵径(mm). 1614. +. 1n. 1.01.2. n.d.. n.d.. +. n.d. 561. n.d.. n.d.. -. n.d.. n.d.. n.d.. 2n (50). +. 2n×2n/CS, PS 4n×4n. 3n (75). +. n.d. 6327. n.d. -. n.d. +/. ~1.5n. 4n (100). +. +. 2n. 1.21.4. +. 2n. 2n (50). +. +. 1n. 1.1. n.d.. 雌性発生四倍体. 4n×UV 4n×UV/PS. 7369 1 0. 4n (100). +. 250. n.d.. 雄性発生二倍体. UV×4n. 2n (50). +. n.d.. 2n×4n, 4n×2n. 3n (75). +. n.d.. n.d. +. 3n (2n×4n)×UV. 3n (75). +. 7 0. +. 倍数体 二倍体 雌性発生半数体 雌性発生二倍体 雄性発生半数体 雄性発生二倍体 三倍体 四倍体 雌性発生二倍体. 三倍体 雌性発生三倍体. 五倍体 六倍体. 交雑/ 染色体操作. 倍数性 (染色体数). 生存性. 2n×2n. 2n (50). +. 2n×UV 2n×UV/CS. 1n (25). -. 2n (50) 1n (25). UV×2n UV×2n/PS. 4n×2n/HS 4n×4n/PS. 卵. 雄. 精子. 妊性. 倍数性. +. 1n. n.d. 1n. 1.1. 3n. 1.4. 1n. 1.1. 3n 1.4n1.5n. 1.4 1.31.4. 5n (125). +. n.d.. +. 2n. 1.2. 6n (150). +. 1110. +. 3n. 1.4. n.d. - -. n.d. +. 3n. UV配偶子の紫外線照射処理,CS第二極体放出阻止のための低温処理,HS同 高温処理,PS同 圧力処理. 卵よりも大きかった。一方,四倍体と二倍体の交配に由. (1)の場合は 4 セッ ト 2 組)の二つの道筋が考えられる。. 来する三倍体について,生殖腺を観察したところ,三倍. トの染色体が減数分裂時に四価染色体としてグループを. 体雄は不妊であり,雌は大型卵と小型卵のいずれか,あ. 作るが,(2)の場合は両親種に由来する相同染色体が二. るいは両方を産生した。交配子孫の倍数性判定と遺伝型. 価染色体を作ると考えられる。四倍体ドジョウの精母細. 分析から前者は 3n ,後者は 1n と推定された(表 3 )。. 胞,卵母細胞の減数分裂像を見ると,少数の四価染色体. 以上の結果は,不妊性を示す人為的な同質三倍体と大き. と多数の二価染色体により構成されていた。810) 四価染. く異なり,三倍体であっても染色体セットの構成により. 色体が見られたことは,四倍体が一旦はゲノム全体の重. 妊性を示す場合があることがわかった。810) その後の細. 複により作られたことを示唆する。おそらく,四価染色. 胞学的,遺伝学的研究から三倍体の 3n 卵はほぼ遺伝的. 体ばかりの減数分裂は不安定であり,短時間で再二倍体. に均一であり,「減数分裂前核内分裂」の機構(後述). 化することで安定した対合から成る二価染色体が増加し. で,また,1n 卵は「減数分裂雑種発生」の機構(後述). たものと推察された。810) ゲノム重複により二倍体から. により形成されることが推定された。810). 四倍体が生じたことは,中国長江流域の二倍体と四倍体. 二倍体,三倍体と四倍体の成長を約 3 年間の混合飼. ドジョウが一つのクレードに纏められ,両者がミトコン. 育試験により比較したところ,四倍体の成長は二倍体,. ドリア DNA のハプロタイプを共有すること29)からも支. 三倍体に劣った。同様に 3 年 2 月齢まで,六倍体,五. 持される。. 倍体,四倍体の間で比較したところ,六倍体の成長は著 しく劣り,体重は他の半分程度であった。同様にクロー. 自然クローンドジョウの発見. ン二倍体と四倍体(後述)の間で 6 月齢魚の成長を比. 日本国内各地の野生集団で自然倍数体の出現を調べた. 較すると,両者はゲノム量以外遺伝的に同一なはずであ. ところ,四倍体は全く見られなかったが,いくつかの地. るが,四倍体の成長は有意に劣った。810) 以上の結果. 域で自然三倍体が見られた。810) 特に顕著な出現率を示. は,ある範囲を超えたゲノム量の上昇は生物に有害な効. したのが北海道東部網走管内である。同様な三倍体出現. 果を与えることを示した。. は新潟県の一部でも見られ,その原因として 2n 卵を産. ところで,自然四倍体ドジョウはどのように形成され. む二倍体の存在が示された。そこで,道東で採集した二. たのだろうか。四倍体の成因として,(1)ゲノム全体の. 倍体ドジョウから得た成熟卵を人為的に雌性発生させた. 重複(同質四倍体相同な染色体セット 4 組),( 2 )交. と こ ろ , 6 尾 中 4 尾 は 1n 卵 を 産 み 致 死 的 な 半 数 体 と. 雑を介したゲノム倍加(異質四倍体母系種からの相同. なったが,2 尾からは正常なドジョウ仔魚が出現した。. な染色体セット 2 組父系種からの相同な染色体セッ. このことは,これら 2 尾は 2n 卵を産むことを示してい.
(6) 202. る。そこで,これらの 2n 卵に対して正常二倍体ドジョ. たところ,アロザイムの結果と同様に,道東(A グルー. ウの精子( 1n )を授精し,三倍体子孫が生じるか否か. プ)と他の地域(B グループ)に二分され,他の地域も. を確かめたところ,予想に反して三倍体よりも二倍体の. 大まかに二つのグループ( B1 と B2 グループ)に分岐. 仔魚が多数出現した。多数の二倍体仔魚の出現は自然の. した。810) そして, A グループのなかにクローン特異的. 単為発生の可能性を強く示したことから,これらの 2n. なハプロタイプが含まれ,これらは有性生殖をおこなう. 卵にキンギョの精子を授精した。その結果,多数の正常. 同じ A グループの野生型二倍体のハプロタイプとは異. なドジョウ仔魚と少数の致死的な発生異常をもつドジョ. なった。810) アロザイムで識別した 3 グループの分布は. ウとキンギョの雑種が出現した。受精卵の切片を見ると,. mtDNA で識別した 3 グループとよく一致した。また,. 2n 卵に侵入した精子は前核化できず,卵は雌性前核の. B2 グループは前述の自然四倍体を含む中国産ドジョウ. みで卵割を開始した。また,一部の卵では精子由来の. に近いグループであった。最近のミトコンドリアゲノム. 1n 雄性前核が 2n 雌性前核と融合する像も低い率で見ら. 全体のシーケンスから,A グループは Misgurnus nikol-. れた。従って,多くの 2n 卵が精子依存性の単為発生,. skyi, M. mohoity(黒竜江ドジョウ)に近く,B1 グルー. すなわち雌性発生を自然に起こし,一部の 2n 卵が 1n. プは M. bipartitus(北方ドジョウ)に近縁なドジョウと. 精子を取り込んで自然三倍体となっていた。810) 二倍体. 判明した。30) M. anguillicaudatus を記載した標本は中国. ドジョウが 2n 卵を産み,これらの卵が雌性発生して二. 舟山群島産であるが,中国産ドジョウは上記とは別のク. 倍体として発生することは,子孫は母親と同一の遺伝子. レードにあった。30) 以上の結果は日本産ドジョウの分類. 型をもつ公算が高い。そこで,反復配列をプローブとし. 学的再検討の必要性を示している。. た DNA フィンガープリント法と複数のマイクロサテラ. 核 遺 伝 子 RAG 1 ( recombination activating gene 1 ). イトマーカーを用いて調べたところ, 2n 卵から生じた. の配列を調べると,配列自体は A と B のクレードにき. 二倍体子孫はすべて母親と遺伝的に同一のクローン二倍. れいに別れ,それぞれの配列は A と B グループ野生型. 体であった。一方,三倍体子孫は母親の遺伝子型と精子. 二倍体に見られた。ところが,クローンはすべての個体. 由来のマーカーを示した。以上の結果は,道東には雌雄. が A と B の配列の両者を持つヘテロ接合体であった。. の有性生殖をしている野生型二倍体ドジョウに加えて,. 同 様 の 結 果 は 核 遺 伝 子 IRBP2 ( interphotoreceptor. これらの野生型ドジョウ雄の精子を用いて雌性発生によ. retinoid-binding protein 2)においても得られた。これら. り繁殖しているクローンドジョウが生息し,精子核を偶. の結果はクローンが A 及び B グループに由来する遺伝. 然取り込んだ時は自然三倍体が出現することを示してい. 子を有する雑種起源であることを示す。810) ドジョウの. る。810) 同様のクローンは石川県の能登島の一部に認め. 遺伝的に分岐したグループは RAG-1 の RFLP,反復配. られた。810). 列 ManDra および ManBgl の電気泳動像により簡単に 識別され,いずれの手法においても,クローンは A と. 日本産ドジョウにおける遺伝的分岐とクローンの交雑起. B1 グ ル ー プ の 染 色 体 セ ッ ト を 有 す る こ と が 示 さ れ. 源. た。31) 三倍体やクローンが出現する地域と他の地域のドジョ. さらに,ManDra 配列をプローブとして A および B1. ウは同じドジョウか。また,先の四倍体ドジョウと日本. グループの野生型二倍体から得た染色体分裂像( 2n =. に生息するドジョウ( 2n = 50 )との関係はどうか。ド. 50)について FISH(‰uorescence in situ hybridization). ジョウを材料とする以上,日本全国に生息するドジョウ. 分析を行った。32) その結果,A グループのドジョウの染. についての考察が必要であることに気づき,遺伝マー. 色体には全くシグナルが見られなかったのに対し, B1. カーを用いた集団遺伝学的研究に着手した。先ず,複数. グループのドジョウでは,すべての染色体のセントロメ. のアロザイム座を用いて調べたところ,日本産ドジョウ. アに強いシグナルが見られた。次にクローンの染色体分. は,道東のドジョウ(A グループ)とそれ以外(B グルー. 裂像に対して FISH をおこなったところ,全体の半分の. プ)に分けられ,さらに,それ以外のドジョウは二つの. 25 本の染色体にのみシグナルが見られた。このことか. グループ(B1 と B2)に分岐した。810) B1 は北海道から. ら,クローンはシグナルを示さない A グループ由来の. 本州,四国,九州のほぼ全国各地に分布し, B2 は関. 25 本の染色体と強いシグナルを示す B1 グループ由来. 東甲信越東海に多く見られ,さらに本州西部日本海. の 25 本の染色体をもつ雑種であることが示された。32). 側でも高い頻度で見られた。ドジョウのように馴染み深. 以上の結果は,クローンは両グループの染色体を有す. く,どこにでもいるありふれた魚は,日本中同じだろう. る こ と を 示 す 。 し か し , 現 生 の A グ ル ー プ と B1 グ. と思っていたが,異質なグループにより構成されていた. ループ間の人為交雑に由来する雑種には 2n の卵を形成. ことに驚いた。. する子孫も見られたが,それらの 2n 卵は自然雌性発生. 次に,ミトコンドリア( mt ) DNA 調節領域を解析し. を起こさなかった。また,道東には B1 グループ野生型.
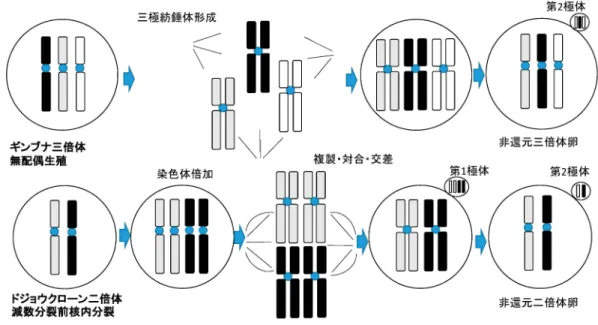
(7) 203. 二倍体は分布しない。従って,クローンは現生の雑種で. し,精母細胞の減数分裂像には 50 本の二価染色体がみ. はなく,過去の交雑に起源すると考えられる。810). られたことから,雌と同様の機構により 2n 精子を形成 していた。810). クローンの生殖機構減数分裂前核内分裂と姉妹染色体 間対合. 前に述べたようにクローンは雑種起源であり,A と B グループに由来する染色体により構成される。そこで,. クローンの 2n 卵形成機構解明のため,卵母細胞を採. 精 母 細 胞 の 減 数 分 裂 像 で FISH を お こ な う と , A グ. 取し,人為的に最終成熟を誘起した。移動を開始した卵. ループ由来の染色体はそれらの間で,B グループ由来の. 核胞を崩壊前に単離して染色体観察を行うとともに,そ. 染色体はそれらの間で対合を行い,二価染色体を形成し. の後の過程を観察した。その結果,クローン二倍体にお. ていた。32) また,核小体形成部位をもつ第一染色体で. いても野生型二倍体と同様に,卵母細胞は減数分裂再開. は,同じグループに由来する姉妹染色体が対合している. 後,第一極体を放出して第一分裂を完了し,第二減数分. ことが明らかになった。32) 従って,異なるグループに由. 裂中期で排卵を待った。この結果は,クローン生殖をす. 来する染色体は対合することができないので(やむな. るギンブナ三倍体のように減数分裂第一分裂をスキップ. く),減数分裂に入る前にすべての染色体が倍加す. して,第二分裂のみで成熟卵を形成する無配偶生殖(ア. る。そして,これらの姉妹染色体があたかも相同染色体. ポミキシス)(図 1 )とは異なる機構が関与しているこ. のように対合して,疑似的な減数分裂を行い, 2n の配. とを示す。810) そこで,卵核胞の中の染色体を観察した. 偶子を形成することが分かった。32) この場合,染色体の. ところ,野生型二倍体では染色体が対合して形成される. 乗換え(組換え)があっても同じエレメントの交換とな. 二価染色体が 25 本見られた。これに対して,クローン. るので遺伝的変異は起こらず,生じる配偶子は体細胞と. では二価染色体が 50 本見られた。体細胞は野生型もク. 遺伝的に同一のクローンとなる。32). ローンも 2n=50 であるので,クローンの卵母細胞では. それでは,減数分裂前に生じる全染色体の倍加は何. 野生型の二倍の染色体をもつことになる。このことは減. 時,どの細胞で起こるのであろうか。生殖腺の組織観察. 数分裂に入る前に,すべての染色体が倍加したこと(減. ではクローンドジョウの精原細胞は野生型に比べて大型. 1)。810). クローンは雌性. であった。そこで,パーコールの密度勾配遠心分離で精. 発生で繁殖することから自然界では全雌であるが,テス. 原細胞を濃縮して,これらの核を染色するとともに,提. トステロン処理により性転換ができる。このような性転. 供いただいた初期の精原細胞を特異的に染める抗体によ. 換クローン雄は遺伝的に均一なクローン 2n 精子を形成. りラベルし,フローサイトメトリーに供した。その結. 数分裂前核内分裂)を示す(図. 図1. 非還元卵形成機構 上段ギンブナ三倍体に見られる「無配偶生殖」,三極紡錘体形成により減数分裂の第一分裂がスキップされ,第二分裂で非 還元三倍体卵が形成される。下段ドジョウクローン二倍体に見られる「減数分裂前核内分裂」,すべての染色体が倍加し, 姉妹染色体があたかも相同染色体のように複製,対合,交差(同じエレメントの交換なので変異は生じない)を行い,二回の 減数分裂を経て,非還元二倍体卵が形成される。.
(8) 204. 果,野生型二倍体の精原細胞では 2C の DNA 量にピー. の間では対合が起き,相同性が低い染色体は排除され. クが見られるのに対し,クローンの精原細胞では 4C の. る。従って,三倍体( AAB )では B グループ由来染色. DNA 量にピークがみられ,クローンの初期精原細胞で. 体は排除 され, 残った A グ ループ由 来染色体 間で複. 染色体倍加が起こっていることが明らかになった。810). 製,対合,乗換えが生じ,減数分裂が進行する。810) 一. これらの結果は,初期の精原細胞(おそらく,雌では卵. 方,三倍体( ABB )では A グループ由来の染色体が排. 原細胞)で染色体倍加が起こり,その結果,姉妹染色体. 除され,減数分裂が進む。810) クローン由来三倍体の卵. 同士での対合が生じ,クローン配偶子が形成されること. 核胞を見ると二価染色体が 25 本観察できるので,卵母. を示す。810) このことは初期の生殖細胞が,減数分裂過. 細胞核にはすでに 2n 分の染色体しか存在していない。. 程で対合ができるかどうかをチェックし,できそうもな. 染色体排除の機構に興味がもたれるが,現在のところ,. い場合は全染色体を倍加させる機構が存在することを示. 何時,どの細胞で染色体がどのようなプロセスを経て排. 唆している。. 除されるのかは全く不明である。以上のように,ドジョ ウではゲノム構成の異なる二倍体クローン( AB )と四. クローン四倍体における減数分裂とクローン由来三倍体. 倍 体 ク ロ ー ン ( AABB ), ク ロ ー ン 由 来 の 三 倍 体 (AAB, ABB)では,染色体セット(ゲノム)構成に応. における雑種発生 クローンドジョウの 2n 卵を確実に雌性発生させるた め,紫外線照射したキンギョ精子を授精して,その後,. じて特殊な生殖が生じる。 以上,ドジョウをモデルとした染色体セットの構成と. 第二極体放出を阻止すると,四倍体クローン(4n=100). 特殊な生殖様式について述べたが,これらはドジョウに. が 生 じ る 。810) ま た , 性 転 換 ク ロ ー ン の 2n 精 子 で ク. 特異的な現象ではなく,他の動物においても成立すると. ローンの 2n 卵を授精した時,精子核が偶々取り込まれ. 考える。何故なら,非還元的 2n 卵の形成はドジョウ. た場合も四倍体クローンが生じる。810) これら四倍体ク. 科,サケ科,メダカ科,コイ科などの人為雑種で報告さ. ローンは DNA 量が 2 倍あるが,遺伝的には二倍体のク. れ て お り , 2n 卵 を 介 し て 倍 数 体 が 生 じ る か ら で あ. ローンと同一である。従って,もし,二倍体クローンに. る。610) また,シマドジョウ類では雑種の産する 2n 卵. 「染色体倍加遺伝子」があれば,四倍体クローンも非還. が雌性発生をした希な例が報告されている。33) 自然界全. 元的に 4n 配偶子を作ると考えられる。しかし,四倍体. 体をみると雌性発生(クローン生殖)あるいは雑種発生. 配 偶 子 を 形 成 し た 。810). 四倍体の場合. (ヘミクローン生殖)により繁殖する魚種もアマゾンモ. は,既にその生殖細胞に体細胞同様に A グループ由来. リー,ギンブナ,アイナメ類34) など多岐にわたり,い. 染色体が 50 本, B グループ由来染色体が 50 本あるの. ずれも雑種起源とされている。610) このように,雑種―. で(染色体セットの構成 AABB 異質四倍体あるいは複. 倍数体―特殊な生殖は互いに強く関係する。今後,交雑. 二倍体),二倍体クローン(染色体セットの構成 AB ). 等によりもたらされる染色体セット(ゲノム)構成の変. の生殖細胞において染色体が倍加した場合(AABB)と. 化が染色体倍加や削減,さらに特殊な生殖を起こす分子. 同様の構成となっている。従って,このまま減数分裂を. 機構が解明できれば,従来の染色体操作とは異なる原理. ク ロ ー ン は 2n. 行うことにより 2n 配偶子が産生される。この結果は,. でプロイディゲームを行う新たな技術が開発できる。著. 特定の倍加遺伝子がクローンに存在するのではなく,生. 者は既に時間切れであるが,この問題は優れた後進に委. 殖細胞自体が染色体セットの構成をチェックして,減数. ねたい。. 分裂において染色体が対合できるかどうかを判断してい ることを示唆する。すなわち,四倍体のように対合でき る場合は,生殖細胞はそのまま減数分裂に入り,できな い場合は減数分裂前に染色体倍加を行う。. 結びと将来展望 魚介類の異種間交雑研究は 1970 年代に盛んになった が,雑種は作ってみないとその性能がわからず実用上の. クローンの 2n 卵(AB)が同所的に生息する A グルー. 確実性に乏しいこと,さらに放流等による遺伝資源保全. プ野生型二倍体( AA )の 1n 精子を取り込むと異質三. への影響が問題点とされ,6) その後研究は下火になっ. 倍体( AAB )が生じる。また, B グループ野生型二倍. た。染色体操作は 1980 年代にブームとなったが,人為. 体( BB )の 1n 精子を取り込むと異質三倍体( ABB ). 三倍体において期待通りの成長改善が見られない魚種が. が生じる。野生型二倍体( AA, BB)から第二極体放出. 多く見られたことから,養殖利用は停滞した。4,5) 結果と. 阻止法で作出する同質三倍体( AAA, BBB )において. して,交雑育種7) も染色体操作育種4) も傍流となり,表. は,雌は卵巣形成が不全となり,雄は異数性精子を形成. 現型, DNA マーカーおよびゲノム情報に基づく選抜育. する。しかし,クローン由来三倍体では雄が不妊とな. 種が主流となった。35,36) しかし近年,染色体操作と交雑. り,雌は 1n 卵を形成する。810) この卵形成は「減数分. は主要な不妊化手法と評価され,遺伝子組換え魚,ゲノ. 裂雑種発生」という様式で行われ,相同性が高い染色体. ム編集魚のみならず養殖品種自体の生物学的封じ込め,.
(9) 205. 知財保護の方法として注目されるとともに,生殖系列キ メラ作成の際の宿主として利用されている。4,5,7) また,. 11). 交雑は生活史の長い魚種では,短期間で優良品種を樹立 するための手法として見直される。7) さらに,異種間交. 12). 雑は同属内の近縁種を新規形質導入のための育種素材 (遺伝資源)として利用するための重要な方法であるこ. 13). とが指摘されている。36) 本稿では,交雑によりゲノム倍 加や特殊な生殖を生じる例を見たが,雑種に偶々生じた 異質倍数体を育種素材とした品種(フナ×コイ,フナ×. 14). ダントウボウ)が中国では大規模に養殖されている。7) このような交雑から生じる「自然倍数体」は植物育種で は重要な育種素材である。37) また,育種支援技術となる. 15). 生殖系列キメラは植物の「接ぎ木」の発想を魚類で発展 させたものである。38) 水産育種は家畜育種を手本として. 16 ). 発展してきたが,陸上動物ではできない育種を魚介類で 開発する出発点に我々はたどり着いたのかもしれない。 水産生物の特性を活かした今後益々の研究の発展を願い. 17). たい。 謝. 辞. すべての共同研究者に厚くお礼申し上げるとともに,. 18). 研究者として著者を育ててくれた日本水産学会に深く感 19 ). 謝する。 文 1). 2). 3). 4). 5). 6) 7). 8) 9) 10). 献. 荒井克俊.染色体操作.「魚類の DNA 分子遺伝学的ア プローチ」(青木宙,隆島史夫,平野哲也編).恒星社厚 生閣,東京.1997 : 3262. Arai K. Chromosome manipulation in aquaculture: recent progress and perspective. Aquaculture Science 2000; 48: 295303. Arai K. Genetic improvement of aquaculture ˆnˆsh species by chromosome manipulation techniques in Japan. Aquaculture 2001; 197: 205228. 荒井克俊,藤本貴史,山羽悦郎.第 6 章 染色体操作と 育種.「水産遺伝育種学」(中嶋正道,荒井克俊,岡本信 明,谷口順彦編).東北大学出版会,仙台. 2017: 99 118. Arai K, Fujimoto T. Chapter 6. Chromosome manipulation techniques and applications to aquaculture. In: Wang HP, Piferrer F, Chen SL, Shen ZG (eds). Sex Control in Aquaculture, Volume I. John Wiley & Sons, Ltd, NJ, USA. 2019: 137162. 荒井克俊.異質倍数体.「水産増養殖と染色体操作」(鈴 木亮編)恒星社厚生閣,東京.1989: 8294. 荒井克俊,藤本貴史,山羽悦郎.第 10 章 交雑と育種. 「水産遺伝育種学」(中嶋正道,荒井克俊,岡本信明,谷 口順彦編) .東北大学出版会,仙台.2017: 171 186. 荒井克俊.ドジョウの倍数性とクローン,それらの特殊 な生殖様式.動物遺伝育種研究 2009; 37: 5980. 荒井克俊.魚介類における雑種,倍数体とクローンの研 究 40 年をふりかえって.水産育種 2018; 48: 150. Arai K, Fujimoto T. Genomic constitution and atypical reproduction in polyploid and unisexual lineages of the Misgurnus loach, a teleost ˆsh. Cytogenetic Genome Res.. 20). 21). 22). 23). 24). 25). 26). 27). 2013; 140: 226240. Arai K, Naito F, Sasaki H, Fujino K. Gynogenesis with ultraviolet ray irradiated sperm in the Paciˆc abalone. Bull. Jpn. Soc. Sci. Fish. 1984; 50: 20192023. Arai K, Naito F, Fujimo K. Triploidization of the Paciˆc ablone with temperature and pressure treatments. Bull. Jpn. Soc. Sci. Fish. 1986; 52: 417422. Fujino K, Arai K, Iwadare K, Yoshida T, Nakajima S. Induction of gynogenetic diploids by inhibiting second meiosis in the Paciˆc abalone. Nippon Suisan Gakkaishi 1990; 56: 17551763. Kudo M, Arai K, Fujino K. Triploidization of Haliotis diversicolor diversicolor by cold shock. Nippon Suisan Gakkaishi 1991; 57: 12631267. 工藤真弘,荒井克俊,木本 功,皆川 恵,藤野和男. フクトコブシ人為三倍体の生残,成長および成熟.水産 増殖 1994; 42: 605613. 柿本芳久,相田 聡,荒井克俊,鈴木 亮.トラフグ Takifugu rubripes における低温,高温処理による雌性発 生二倍体の誘起.広島大学生物生産学部紀要 1994; 33: 103112. 柿本芳久,相田 聡,荒井克俊,鈴木 亮.マコガレイ Limanda yokohamae における温度および圧力処理による 雌性発生二倍体の作出とメチルテストステロン浸漬処理 に よ る 性 転 換 . 広 島 大 学 生 物 生 産 学 部 紀 要 1994; 33: 103112. Aida S, Arai K. Sex ratio in the progeny of gynogenetic diploid marbled sole Limanda yokohamae males. Fish. Sci. 1998; 64: 989990. 森 立成,斎藤節雄,岸岡稚青,荒井克俊.マツカワ Verasper moseri における三倍体および雌性発生二倍体の 誘起.日本水産学会誌 2004; 70: 145 151. Mori T, Saito S, Kishioka C, Arai K. Aquaculture performance of triploid barˆn ‰ounder Verasper moseri. Fish. Sci. 2006; 72: 270277. Lahresh Z, Kishioka C, Morishima K, Mori T, Saito S, Arai K. Genetic veriˆcation of induced gynogenesis and microsatellite-centromere mapping in the barˆn ‰ounder Verasper moseri. Aquaculture 2007; 272: S115S124. Mori T, Saito S, Matsuda T, Kayaba T, Kishioka C, Lahresh Z, Arai K. Sex ratio and growth performance of gynogenetic diploid barˆn ‰ounder Verasper moseri. Aquacult. Sci. 2011; 59: 375382. Nomura K, Nakajima J, Ohta H, Kagawa H, Tanaka H, Unuma T, Yamauchi K, Arai K. Induction of triploidy by heat shock in the Japanese eel, Anguilla japonica. Fish. Sci. 2004; 70: 247255. Nomura K, Morishima K, Tanaka H, Unuma T, Okuzawa K, Ohta H, Arai K. Microsatellite-centromere mapping in the Japanese eel (Anguilla japonica) by half-tetrad analysis using induced triploid families. Aquaculture 2006; 257: 5367. Nomura K, Takeda Y, Unuma T, Morishima K, Tanaka H, Arai K, Ohta H. Post-ovulatory oocyte aging induces spontaneous occurrence of polyploids and mosaics in artiˆcial fertilization of Japanese eel, Anguilla japonica. Aquaculture 2013; 404405: 1521. Omoto N, Maebayashi M, Adachi S, Arai K, Yamauchi K. Sex ratios of triploids and gynogenetic diploids induced in the hybrid sturgeon, the bester (Huso huso female x Acipenser ruthenus male). Aquaculture 2005; 245: 3947. Havelka M, Zhou H, Hagihara S, Ichimura M, Fujimoto T, Yamaha E, Adachi S, Arai K. Spontaneous polyploidi-.
(10) 206 zation in critically endangered Acipenser mikadoi. Fish. Sci. 2017; 83: 587595. 28) Hou J, Fujimoto T, Saito T, Yamaha E, Arai K. Generation of clonal zebraˆsh line by androgenesis without egg irradiation. Sci. Rep. 2015; 5: 13346. 29 ) 李 雅娟,李 霞,周 賀.中国自然多倍体泥鰍的遺伝 学研究.中国農業科学技術出版社,北京.186.(中文) 30) Shibata K, Yen DT, Fujimoto T, Arai K. Comparative analysis of mitochondorial genomes in genetically distinct groups of the dojo loach Misgurnus anguillicaudatus. Mitochondorial DNA Part B 2020; 5: 38103812. 31) Fujimoto T, Yamada A, Kodo Y, Nakaya K, OkuboMurato M, Saito T, Ninomiya K, Inaba M, Kuroda M, Arai K, Murakami M. Development of nuclear DNA markers to characterize genetically diverse groups of Misgurnus anguillicaudatus and its closely related species. Fish. Sci. 2017; 83: 743456. 32) Kuroda M, Fujimoto T, Murakami M, Yamaha E, Arai K. Clonal reproduction assured by sister chromosome pairing in dojo loach, a teleost ˆsh. Chromosome Res. 2018; 26: 243253.. 33). 34). 35 ). 36 ). 37) 38). Choleva L, Janko K, Gelas KD, Bohlen J, ½Slechtov áa V, Rabova M, R áab P. Synthesis of clonality and polyploidy in vertebrate animals by hybridization between two sexual species. Evolution 2012; 667: 21912203. Kimura-Kawaguchi MR, Horita M, Abe S, Arai K, Kawata M, Munehara H. Identiˆcation of hemiclonal reproduction in three species of Hexagrammos marine reef ˆshes. J. Fish Biol. 2014; 85: 189209. 谷口順彦,坂本 崇,中嶋正道.第 9 章 水産養殖にお ける選抜育種. 「水産遺伝育種学」 (中嶋正道,荒井克俊, 岡本信明,谷口順彦編).東北大学出版会,仙台. 2017: 155170. 菊池 潔.第 11 章 水産育種におけるゲノム情報の利 用. 「水産遺伝育種学」 (中嶋正道,荒井克俊,岡本信明, 谷口順彦編).東北大学出版会,仙台.2017: 187211. 福井希一,辻本 壽. 「改訂版育種における細胞遺伝学」 (渡辺好郎監修).養賢堂,東京.2010.243. 荒井克俊,山羽悦郎.始原生殖細胞移植による魚の“接 ぎ木”生産.~低コストで病気のない種苗生産の可能性 ~.月刊アクアネット,湊文社,東京. 2008 11 巻 10 号5256..
(11)
図



関連したドキュメント
Masami Matsumura 1) , Mika Mori 2) , Toshimi Shimada 3) , Masaaki Kawashiri 2) and Masakazu Yamagishi 2) : 1) Division of Gen- eral Medicine, Center for
c加振振動数を変化させた実験 地震動の振動数の変化が,ろ過水濁度上昇に与え る影響を明らかにするため,入力加速度 150gal,継 続時間
本研究は,地震時の構造物被害と良い対応のある震害指標を,構造物の疲労破壊の
色で陰性化した菌体の中に核様体だけが塩基性色素に
腐植含量と土壌図や地形図を組み合わせた大縮尺土壌 図の作成 8) も試みられている。また,作土の情報に限 らず,ランドサット TM
Mochizuki, On the combinatorial anabelian geometry of nodally nondegenerate outer representations, RIMS Preprint 1677 (August 2009); see http://www.kurims.kyoto‐u.ac.jp/
サビーヌはアストンがレオンとの日課の訓練に注意を払うとは思わなかったし,アストンが何か技を身に
実際, クラス C の多様体については, ここでは 詳細には述べないが, 代数 reduction をはじめ類似のいくつかの方法を 組み合わせてその構造を組織的に研究することができる
