関西学院大学リポジトリ
111
0
0
全文
(2) Table of Contents •. Abstract. ···Page 3-5. •. Introduction. ···Page 6-9. Chapter. 1 Structure of chemicals and their effects on PDI activity •. Introduction l···Page 10-11. •. Chapter. 1-1 Effects of bisphenol A derivatives on protein disulfide isomerase (PDT) activity. •. •. 1. Materials and Methods 1-1. ···Page 12-15. 11. Results 1-1. ···Page 15-17. III. Discussion 1-1. ···Page 17-19. IV. Figures 1-1. ···Page 20-25. Chapter. 1-2 Different biological effects of polybrominated diphenyl ethers (PBDEs) with different chemical structures 1. Materials and Methods 1-2. ···Page 26-29. 11. Results 1-2. ···Page 29-31. III. Discussion 1-2. ···Page 32-34. IV. Figures 1-2. ···Page 35-41. Chapter.2 Inhibition mechanism of chemicals on activity of PDI • •. ···Page 42-44 Introduction 2 Chapter 2-1 Identification of bisphenol A-binding site of protein disulfide isomerase (PD 1) 1. Materials and Methods 2-1. ···Page 45-47. 11. Results 2-1. ···Page 47-51. III. Discussion 2-1. ···Page 51-53. IV. Figures 2-1. ···Page 54-62. V. Tables 2-1. ···Page 63-66 1.
(3) •. Chapter.2-2 Preliminary crystallographic analysis of triiodothyronine-bb' fragment complex of protein disulfide isomerase (PDI) 1. Materials and Methods 2-2. ···Page 67-68. 11. Results and Discussion 2 -2. ···Page 68-69. 111. Figures 2 -2. ···Page 69. IV. Table 2-2. ···Page 69-70. Chapter. 3 Function of PDI in T3-induced gene expression • •. ···Page 71-72 Introduction 3 Chapter.3-l Function of protein disulfide isomerase (PDI) in T3-induced growth hormone (G H) expression in G H3 cells 1. Materials and Methods 3-1. ···Page 74-78. 11. Results 3-1. ···Page 78-83. 111. Discussion 3-1. ···Page 84-86. IV. Figures 3-1. ···Page 87-94. V. Tables 3-1. .. ·Page 95. • Conclusions • Abbreviations • Acknowledgements • References •. ···Page 96 ···Page 97-98 ···Page 99 .. ·Page 100-107. Accomplishments. ···Page 108-109. 2.
(4) Abstract Bisphenol A (BPA) is an endocrine disrupting chemical with several biological effects. Previously, protein disulfide isomerase (PDI) was isolated as a protein binding to BPA. PDI is a dithiolldisulfide oxidoreductase that facilitates the correct folding of proteins.. It consists of a, b, b', a' and c domains, with catalytic active sites in a and a'domain. PDI has not only dithiolldisulfide oxidoreductase activity but also chaperone activity and the capacity to bind to thyroid hormone (Ts).. Previous studies have found that. BPA binds to PDI competitively with Ts, and BPA and Ts inhibit the catalytic activity of PDI.. In the present study, to clarify effects of BPA and its analogs on the catalytic. activity and physiological functions ofPDI, three investigations were conducted.. First,. effects on PDI activity were examined focusing on chemical structure of chemicals. Second, the mechanism inhibiting the activity of PDI was investigated focusing on the structure of the PDI protein.. Finally, to clarify its physiological function, PDI's role in. Ts-induced gene expression was examined In the first chapter, I investigated the relationship between the chemical structure of BPA derivatives and polybrominated diphenyl ethers (PBDEs), and their effects on the catalytic activity of PDI.. To clarify the structure of BPA required to inhibit the. isomerase activity ofPDI, I investigated the effects ofBPA derivatives on the isomerase and chaperone activity of PDI.. Among six chemicals, only phenolic chemical, could. bind to PDI and these chemicals also had an inhibitory effect on PDI activity, suggesting that the hydroxyl group(s) of the benzene ring(s) is important to bind to PDI and inhibit the catalytic activity of PDI. activity of PDI.. I also tested the effects of PBDEs on the isomerase. PBDEs have been used in a variety of consumer products as flame. retardants and are known to be widespread environmental pollutants which probably affect biological functions of mammalian cells.. I tested the effects of 10 PBDE. compounds and their metabolites on PDI activity.. HO-PBDEs specifically inhibited the. isomerase activity of PDI, with 4'-OH-PBDE more effective than 2' (or 2)-OH-PBDEs suggesting that the hydroxyi group(s) of the benzene ring(s) is important to inhibit the catalytic activity of PDI and substitution of the 4' position is more effective to inhibit PDI activity.. 4'-OH-PBDE inhibited the isomerase activity ofb'a'c, but not that of ab. and a'c, suggesting that the b' domain of PDI is essential for the inhibition by 4'-OH -PBDE.. Also, the effects of these chemicals on the production of growth hormone. (GH) in GH3 cells were investigated.. Levels of mRNA and protein of GH stimulated by. Ts were reduced by 4'-OH-PBDE and 4'-MeO-PBDE. The reduction in GH expression caused by these compounds was not changed by the over-expression or knockdown of 3.
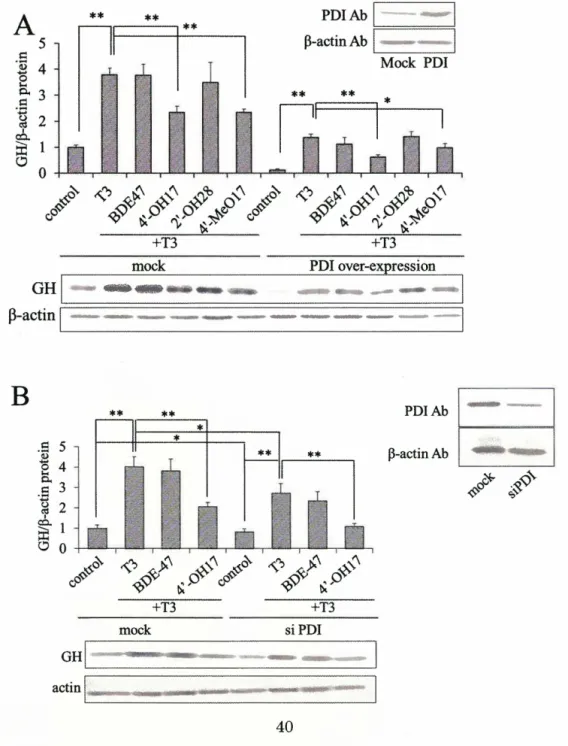
(5) PDI in GH3 cells, while these manipulations significantly suppressed the expression of GH.. These results suggest that the biological effects of PBDEs differed depending on. their brominated and hydroxylated positions. In the second chapter, I investigated the mechanism by which BPA inhibits the catalytic activity ofPDI.. I identified the BPA-binding domain ofPDI, and investigated. effects of BPA on the structure of PDI. Also, I tried to analyze the crystal structure of Ts and the Ts-binding region complex.. First, I identified the BPA-binding domain with. ab, b'a'c, a, b, b', and a'c fragment peptides of PDI by surface plasmon resonance spectroscopy.. BPA interacted with ab, b'a'c, a, and b', suggesting that a and b' domains. are important to the interaction.. Second, ab, b'a'c, a, b, b', a', abb'a', abb', b'a', db', and. a'c fragment peptides were used for their isomerase activity with RNase as a substrate. BPA could inhibit the activity of the peptide fragments including b', suggesting that the b' domain contributes to the inhibition of the catalytic activity of PDr by BPA. investigated the BPA-binding capacity of PDI by amino acid substitution. BPA-binding activity with the mutation ofH258. decreased its activity. activity of PDI.. Next, I PDI lost. The mutation ofQ245 and N300 also. Furthermore, acidic condition increased the BPA-binding. These results suggest that the charge of these amino acids, especially. H258, is important for BPA to bind to PDI.. To clarify the structural features of BPAITs. and the PDI complex, I tried to perform a crystallographic analysis.. I succeeded in. obtaining a crystal of Ts-bb' fragment and analyzed the 3D-structure of the Ts-bb' complex.. Both the native bb' crystals and Ts-bb' crystals were obtained by the hanging. drop vapor diffusion technique with 1.6 M tri -sodium citrate (pH 6.2) as a precipitant. The space group of the native bb' crystals was found to be C222 with the following unit-cell parameters: a = 94.8. A, b = 114.9 A, and c = 182.9 A; while the space group of the Ts-bb' complex crystals was P212121 with a = 99.9 A, b = 184.5 A, and c = 232.2 A. Diffraction data for both types were collected to a resolution of 3.06 and 3.00 A, respectively. In the third chapter, I investigated the function of PDI in Ts-mediated gene expression in rat pituitary tumor (GH3) cells.. Previous study found that. over-expression of PDI in GH3 cells suppressed Ts-induced growth hormone (GH) expression.. This finding suggests that PDI regulates the Ts and thyroid hormone. receptor (TR)-mediated gene expression in GH3 cells. the mechanism regulating the Ts response.. In this study, I tried to clarify. Over-expression of wild-type PDI and db',. which was deleted of the Ts-binding domain, suppressed the Ts-induced GH expression. Conversely, over-expression of CIA mutants of PDI, which have an alanine substituted for the catalytic active cysteine and has the capacity to bind BPA, did not suppress the 4.
(6) GH expression, suggesting the contribution of catalytic activity of PDI to GH expression. Treatment with an oxidizing agent also suppressed GH expression.. These results. indicated that regulation of redox state by PDI is important for GH expression.. I. focused on redox factor I (Ref-I) as a mediator of the redox regulation of PDI. Ref-I-overexpression facilitated the T3-induced GH expression.. On the other hand,. C64S Ref-I did not facilitate GH expression, suggesting that Ref-I, especially the 64th cysteine, contributes to the T3-response.. In a immunoprecipitation study, it was found. that Ref-I interacts with PDI and that cysteine64 is important for this interaction.. In. this study, the regulation of T3-response via the redox activity of PDI and the contribution of Ref-I to this response was demonstrated. Through these studies, I could clarify the relationship between chemical-binding and inhibition of PDI activity, and the physiological function of PDI in T3-mediated gene expression.. 5.
(7) Introduction Bisphenol A (BPA) is widely used as a monomer of polycarbonate and epoxy resins. For example, the coating on food cans, dental sealants, and plastic products such as food trays, and baby bottles, contain BPA(1-3).. BPA is known to leach out from these. products with acid or heat; therefore, it effects on the human body are a concern (4,5). In fact, BPA has been detected in the placentas of pregnant women and saliva after dental treatment at a high concentration (131p.M) (6). systems have been studied for a long time.. Effects of BPA on reproductive. BPA binds to steroid hormone receptors,. including estrogen receptor (ER) , and androgen receptor, and disrupts hormone reactions (3,7-9).. The binding of BPA to these receptors leads abnormalities of. reproductive functions, sexual differentiation and behavior (10-13). However, recent studies suggested that exposure to BPA at an early developmental stage also causes neural toxicity (14).. For example, prenatal and neonatal exposure to. BPA in mice leads to aggressive behavior, decreases in adaptability, and enhancement of methamphetamine sensitization (15,16).. Also, in a previous study in our laboratory,. Mori et. al. (17,18) found that exposure to BPA in Xenop us embryos induced malformation of the head region and a small-eye phenotype. in the central nervous system were not identified.. However, target proteins. In a previous study, to clarify the. mechanism of neural toxicity, a BPA-binding protein was isolated from the synaptosome fraction of rat brain using a BPA affinity column.. The purified. BPA-binding protein was identical to protein disulfide isomerase (PDI) (19). PDI was first identified in 1964 as an enzyme which facilitates the formation of correct disulfides, as monitored by the recovery of enzymatic activity of ribonuclease A by Anfinsen and his coworkers (20).. PDI is mainly located in the endoplasmic. reticulum (ER) , and catalyzes the formation, reduction and rearrangement of disulfide bonds of nascent or misfolded proteins (21-23). domains.. PDI is composed of a, b, b', a' and c. The a, b, b', and a' domains have a thioredoxin fold.. Catalytic active sites. containing a Cys-Gly-His-Cys (CGHC) sequence exist in a and a', while band b' are catalytically inactive (24,25).. The b' domain has a highly hydrophobic region which is. important for interactions with unfolded or misfolded proteins (26,27).. Following the b'. domain, there is an x-linker which provides flexibility and plays an important role in the recognition of substrates by the b' domain (27).. Only in yeast has the crystal. structure of a full-length PDI been solved (28,29). The structure of the yeast PDI showed that its four domains form an overall 'U' shape (28,29). 6. In human PDI, the.
(8) crystal structure ofb'-x and b-b'-x-a'-c was solved (27,30).. The crystal structure ofb'x. showed that the x-linker interacts with the hydrophobic region in the b' domain (27). Also, the structure of a reduced b-b'-x-a'-c revealed that the redox state of a' regulates the conformational change ofb-b'-x-a'-c and its catalytic activity(30). The redox state of PDI is regulated by ER-localized proteins.. As oxidases for PDI,. endoplasmic reticulum oxidoreductin 1 (Ero1) and peroxiredoxin IV (PrxIV) have been identified (31,32).. Ero1 is considered a main regulator of PDI activity (33).. It. possesses flavin adenine dinucleotide (FAD) as a coenzyme and serves as the primary oxidase of PDI (34,35).. Ero1 uses oxygen as an electron acceptor, and hydrogen. peroxide is produced as a byproduct (35).. Recent studies suggested that Ero1 more. effectively oxidizes the cysteine residues of the active site in the a' domain than a domain of PDI (36,37).. Crystal structural analysis of Ero1 suggested that Ero1. interacts with a hydrophobic pocket in the b' domain via its B-hairpin and this interaction leads to effective oxidation of the a' domain (38).. PrxIV is an ER -localized. peroxidase which metabolizes the hydrogen peroxide produced by Ero1(39).. PrxIV. contains cysteine residues in its catalytic active site, and interchain disulfide bonds are formed in the reaction cycle of the peroxidase activity (31,32).. The disulfides are. known to be reduced by the dithiol ofPDI which exists in the isomerase active site. In some PDI -interacting proteins, the redox state of their cysteine residues changed by PDI.. IS. For example, enzymes such as NADPH oxidase complex (40,41) and. vitamin K epoxide reductase (42,43) interact with, and have their catalytic activity regulated by, PDI.. Some transcription factors such as interleukin-4(44), E2A. helix-loop-helix proteins(45), and Pax5(46) are also regulated by reduction of their intramolecular disulfides by PDI.. Also, PDI activity has been reported to be required. for the maturation of MHC Class 1(47), infections of viruses such as HIV (48,49), the activation of tissue factor-mediated blood coagulation(50-52), and so on. PDI function is also important in the developmental stage.. Zhang et a1. (53). reported that ER stress occurs during development of the central nervous system in the mouse, and several ER-resident stress-regulated chaperones, including PDI family proteins and calreticulin, were expressed at higher levels in the embryonic brain and retina, than in adult tissues.. Hoshijima et a1. (54) found that PDI is expressed in the. embryonic midline and is required for left/right asymmetry. A dysfunctional PDI leads to protein misfolding.. In neurodegenerative diseases,. s-nitrosylation of cysteine residues in catalytic active sites of PDI is detected (55-57). The accumulation of misfolded proteins is seen in neurodegenerative disorders such as Parkinson's disease, amyotrophic lateral sclerosis, prion neurotoxicity, and Alzheimer's 7.
(9) disease, and causes the apoptosis of neural cells (57-60).. Inhibition of PDI activity by. s-nitrosylation is considered to be one of the causes of the accumulation of misfolded protein.. On the other hand, expression of PDI and other proteins involved in protein. quality control are induced by the accumulation of misfolded proteins through the unfolded protein response (UPR) or ER stress pathway (61). PDI is a multifunctional protein with roles other than as a thiol-disulfide catalyst. PDI also serves as a molecular chaperone, and prevents nonproductive folding (62). On the other hand, the chaperone activity of PDI is known to be affected by cyclophilin Band cyclosporine A in. vitro (63).. PDI also acts as the. B-subunit of. collagen-prolyl-4-hydroxylase, and microsomal triacylglycerol transfer protein, and contributes to the conformational stabilization of their a-subunit which possesses catalytic activity (64,65).. Moreover, PDI is also a thyroid hormone (T3) or estrogen. (E2)- binding protein (8).. The affinity of PDI for T3 is reported to be 4.3 pM and. considerably higher than that of thyroid hormone receptors(8).. Therefore, PDI is. considered to act as a buffer of hormones using high capacity, low affinity hormone-binding sites.. A previous study in our laboratory investigated the PDI. function in the T3 response using rat pituitary tumor (GH3) cells(66).. In GH3 cells,. expression of growth hormone (GH) mRNA is T3-dependently induced via thyroid hormone receptor (TR), therefore GH3 cells are often used in studies of the T3-response of cells.. The study found that over-expression of PDI in GH3 cells suppressed. T3-mediated growth hormone (GH) expression. T3-mediated gene expression of GH3 cells.. PDI is considered to regulate the. Effects of BPA on GH3 cells were also. investigated. It was found that BPA affects T3-induced GH release but not GH mRNA expression. There are 20 related mammalian proteins, which are mainly located and function in the ER, in the PDI family(67).. PDI is the first characterized protein in the PDI family.. Three other PDls (PDILT, PDlp, ERp57) have the same domain composition, a, b, b', a' (22).. ERp72 has an additional catalytic a domain at the N-termina1(22).. The other. PDls vary in domain composition and catalytic active site sequence, and some do not have a catalytic active domain(22).. In a previous study, BPA-binding capacity and. effects ofBPA on catalytic activity ofPDI family proteins were investigated, focusing on ERp57, ERp72, ERp29 and ERp46.. ERp57 and ERp72 are ubiquitous proteins with a. similar domain structure and active site sequence (CGHC) with PDI(22).. ERp57 is a. glycoprotein -specific disulfide isomerase acting with lectin -like chaperons, calnexin or calreticulin, to recruit substrates (68).. One of the physiological actions ofERp57 is the. processing of MHC class I molecules (69,70).. 8. ERp57 is involved with the folding of.
(10) MHC class I-heavy chains by catalyzing the formation of disulfide bond (71).. ERp72 is. also a disulfide isomerase in the ER which possesses three catalytic active domains. As substrates of ERp72, cholera toxin, matrilin-3, and mutant LDL receptors were identified (72,73). Additionally, ERp72 is also known as a member of a multi-chaperon complex(74) which regulates the folding of interferon-gamma in the ER.. ERp46 has. only catalytic active domains (a-a-a), and the same sequence as PDI, ERp57 and ERp72 in its catalytic active sites (75). thiol-reactive active sites.. On the other hand, ERp29 does not contain. The crystal structure of ERp29 has been clarified and. revealed that ERp29 has one thioredoxin-like domain and a D domain composed of five alpha helixes (76).. Dimerization of ERp29 is essential for its diverse functions as an. escort factor by binding to secretory proteins such as thyroglobulin (Tg) in the ER to facilitate secretion(77).. Previous study found that ERp57, ERp72 and ERp46 have. very little capacity to bind to BPA and T3, and that BPA and T3 do not inhibit their catalytic activity (78,79).. On the other hand, ERp29 has grater binding capacity for. BPA than PDI. Based on this background, to investigate the effects of "chemical binding" on PDI functions, I performed three experiments.. First, focusing on the ligands, I researched. the structure of ligands required to bind to PDI and inhibit its isomerase activity.. Also,. I investigated the effects of polybrominated diphenyl ether (PBDE) on PDI activity. PBDE has been used as a flame retardant and suspected of affecting the development of animals (80-82).. Second, focusing on the structure ofPDI, I identified the BPA-binding. site of PDI, and investigated the mechanism by which BPA inhibits the isomerase activity of PDI.. Third, I investigated the function of PDI in T3-mediated gene. expression using GH3 cells.. 9.
(11) Chapter. 1 Structure of chemicals and their effects on PDI activity Introduction 1 Bisphenol A (BPA) is a monomer of epoxy reSIn and is contained in industrial chemical products such as food cans, dental composites, baby-feeding bottles and plastic waste samples.. On the other hand, exposure to BPA at an early developmental stage. may cause neuronal toxicity during embryonic development.. To clarify the mechanism. of BPA effects on central nervous system, BPA- binding-protein was isolated from the synaptosome fraction of the rat brain, which was identical to protein disulfide isomerase (PDI) (19).. PDI has isomerase activity and facilitate the correct folding of proteins by. dithiolldisulfide exchange. Also PDI has chaperone activity and binding capacity of thyroid hormone (T3), and is considered as a multifunctional protein.. In previous. study in our laboratory, it was demonstrated that BPA binds to PDI competitively with T3, and BPA and T3 inhibit catalytic activity of PDI.. However, required structure of. BPA to bind to PDI and inhibit catalytic activity had not been clarified.. In the study of. chapter 1-1, in order to clarify the effects of BPA on PDI functions, I investigated the relationship. between chemical. structure. of BPA derivatives. PDI -mediated isomerase activity and chaperone activity.. and. effects on. I also investigated the effects. of changes in the PDI active center on the binding of chemicals to PDI, using a PDI mutated isomerase active site, and oxidized or reduced PDI. Polybrominated diphenyl ethers (PBDEs) are widely used in a variety of consumer products, for example, as flame retardants in plastics, textiles, coatings, and the electrical components found in computers, televisions, and appliances(83). also concerned about biological effects same as BPA.. PBDE is. In the past several years, PBDEs. have turned out to be widespread environmental pollutants found in sediment and fish from rivers, indoor air(84), and house dust(85).. PBDEs have also been detected in. human blood and breast milk, because of their bio-accumulative nature(86).. The. PBDEs most commonly detected in human samples are 2,2' ,4,4'-tetrabromodiphenyl ether. (BDE-47),. 2,2',4,4',5-pentabromodiphenyl. ether. (BDE-99),. 2,2',4,4',6-pentabromodiphenyl ether (BDE-I00), and 2,2',4,4',5,5'-hexabromodiphenyl ether (BDE-53).. In addition, recent studies have shown that oxidizing enzymes such. as cytochrome P450 convert PBDEs to hydroxylated PBDEs (OH-PBDEs), which are further methylated to form MeO-PBDEs (87-89).. Several recent reports have. described the effects of PBDEs on mammalian cells, for example, oxidative stresses in 10.
(12) vitro (90,91), increases in levels of lipid peroxidation and oxidized glutathione(SO,92), an increase in some phase II metabolizing enzymes such as uridine diphosphoglucuronosyl transferase (UGT) in rats(93), endocrine disrupting effects, and so on(93,94).. Moreover,. effects ofPBDEs on nuclear hormone receptor activity have been reported (S2,95,96). T3, and BPA have two benzene rings either or both of which are hydroxylated.. BPA. and T3 bind to PDI via a hydroxyl group of the benzene ring(s) and inhibit the isomerase activity ofPDI.. In the study of chapter 1-2, the BPA derivatives which have a hydroxyl. group can also bind to PDI, while those with methylated hydroxyl groups are not able to bind.. BPA derivatives, which possess methyl group(s) substitutions for hydrogen(s) at. the center of the compounds, are also able to bind to PDI and inhibit isomerase activity similarly to BPA (97,9S).. Moreover, it was demonstrated that hydroxylated. polychlorinated biphenyls (HO-PCBs) interact with PDI and inhibit its activity(99). Compounds with a halogenated substitution at the ortho position of phenol such as HO-PCB and T3 have stronger binding affinity to PDI which confers on these compounds stronger inhibitory effects on isomerase activity than that of BPA(9S,99). Moreover, previous study investigated the effects of BPA on the T3-evoked response of rat pituitary tumor (GH3) cells(66). (GH). In GH3 cells, production of growth hormone. is stimulated by T3 via the thyroid hormone receptor (TR)(100).. BPA can. disrupt the function of thyroid hormone in GH3 cells, and the production of GH induced by T3 is influenced by the expression ofPDI. In the study of Chapter 1-2, to investigate the relationship between the structure of PBDEs or metabolites and their toxicity, I tested effects of these chemicals on the isomerase activity of PDI and on the response of GH3 cells to T3.. 11.
(13) 1-1 Effects of bisphenol A derivatives on protein disulfide isomerase (PDI) activity I. Materials and Methods 1-1. Materials 2,2-bis(4-hydroxyphenyl)propane. (bisphenol A),. 1,I-bis(4-hydroxyphenyl)ethane. (bisphenol E), and 4,4-methylenebisphenol (bisphenol F) were purchased from Wako Pure Chemical Industries Ltd. (Osaka, Japan).. [G-3H]BPA ([3H]BPA; specific activity,. 5 Cilmmol) was obtained from Moravek Biochemicals, Inc. (Brea, CA).. Sucrose. monolaurate. Japan).. was. purchased. from. Dojindo. Laboratories. (Kumamoto,. Diphenylpropane, 3,3',5-triiodo-L-thyronine (T3), ribonuclease A (RNaseA) type III from bovine pancreas, monobromobimane and alcohol dehydrogenase were purchased from Sigma-Aldrich Corp. (St. Louis, MO). Chemical Industry (Tokyo Japan).. 4-a-Cumylphenol was purchased from Tokyo. 2,2-bis(4-methoxyphenyl)propane (DMBPA) was. prepared as described previously (101).. BPA and other chemicals were dissolved in. ethanol or dimethylsulfoxide (DMSO) at 100 mM to make stock solutions, which were stored at -80°C.. T3 was dissolved in 0.1 M NaOH at 10 mM to make stock solutions. and was stored at -20°C.. Appropriate vehicle controls were performed in all. experiments Cloning and expression of rat PDI mutants Rat PDI cDNA was cloned in previously study (19).. Rat PDI has two domains (a,a'). for isomerase activity. I prepared 3 mutants; by changing Cys residues (at Cys54 and Cys57 in a domain, and Cys398 and Cys401 in a' domain) to Ala residues: mutant of a and a' , designated aa'm; mutant of a ,am; mutant of a' ,a'm. PDI mutant (aa'm) was constructed from four fragments (Fig. 1-1-1). Fragment 1 was amplified with two primers, forward primer 1: 5'-GGGGGGATCCTCCGACATGCTGAGCCGTGC-3' and reverse. primer. 1:. 5'-TGGGGCCAGTGCTTTGGCGTGGCCAGCCCATGGGG-3'. (underline, complementary sequence to forward primer 2, and double underline, exchanged nucleotide for Cys to Ala). Fragment 2 was amplified with forward primer 2: 5'-AAAGCACTGGCCCCAGAGTATGCCAAAGC-3' (underline, complementary sequence to reverse primer 1) and reverse primer 2: 5'-TCAAGTATGCGCTGGTTGTCAGTG-3'. Fragment. 3. was. AGATCCTGTTCAC-3'. amplified and. with. forward reverse. prImer. 3:. prImer. 5'-CTTTAAGGGCA 3:. 5'-. CGGGGCTAGCTGCTTGGCGTGACCAGCCCAGGGAG -3' (underline, complementary sequence to forward primer 4, and double underline, exchanged nucleotide for Cys to 12.
(14) Ala).. Fragment. 4. was. amplified. with. forward. prImer. 4:. 5'-AAGCAGCTAGCCCCGATTTGGGATAAA-3' (underline, complementary sequence to reverse. primer. 3). and. reverse. 5'-TTGCGGCCGCGGCGCCCAGATCTGGCTTCTGCA-3'.. primer The. four. fragments were purified from an electrophoresis with an agarose gels.. 4: amplified. The second. PCRs were performed with two fragments (fragment 1 and 2, or fragment 3 and 4) and two primers (forward primer 1 and reverse primer 2, or forward primer 3 and reverse primer 4), and then, third PCR was performed with two fragments (fragment 1+2 and 3+4) and two primers (forward primer 1 and reverse primer 4).. The amplified. fragment was subcloned into pGEM-T Easy (Promega Corp., Madison, WI), and then the cDNA of mutant rPDI in pGEM-T Easy was digested with SphI and SalI, and subcloned into pQE-82L vector (QIAGEN, Valencia, CA). were prepared as follows.. PDI mutants am and, a'm. The C-terminal region of rPDI WT in pQE-80L was cut out. with ClaI and SalI, and subcloned into the pQE-82L plasmid, which includes aa'm, and digested with ClaI and SaIl. The digested C-terminal region ofrPDI aa'm was subcloned into the pQE-80L plasmid, which includes WT, and digested with ClaI and SaIl. The resulting plasmid was transfected into Escherichia coli (E.coil) DH5a (Toyobo Co. Ltd., Osaka, Japan). Purification of histidine-tagged rat fusion PDI Rosetta gami B (Novagen, Madison, WI) E.coli cells transformed with pQE-80L encoding histidine-tagged rat PDI were grown at 37°C in 2x yeast extract-tryptone-rich medium containing 0.1 mg/ml ampicillin, 0.034 mg/ml kanamycin, 0.02 mg/ml tetracycline, and 0.015 mg/ml chloramphenicol.. Protein expression was induced by. adding 1.0 mM isopropylthio-B-O-galactoside. After additional cultivation for 8 h, E. coli cells were harvested and lysed in a lysis buffer (50 mM NaH2P04 (pH 7.5), and 300 mM NaC!), containing 1.0 mg/mllysozyme and 20 mM imidazole for 60 min at 4°C.. E. coli. cell lysate was sonicated for 1 min, and then proteins were solubilized with 0.25 % sucrose monolaurate for 60 min at 4 °c with gentle stirring.. The lysate sample was. centrifuged at 50,000 x g for 30 min, and the supernatant was loaded onto a Ni-NTA agarose column (QIAGEN).. After the column was washed with washing buffer (lysis. buffer including 0.1 % sucrose monolaurate), the protein was eluted with lysis buffer containing 250 mM imidazole, and 0.1 % sucrose monolaurate.. The eluted fraction was. dialyzed against 50 mM Tris-HCI buffer (pH 7.5), and loaded onto an anion exchange column (MonoQ).. Mono Q HR 10/10 column (Amersham Bioscience, Tokyo, Japan) was. run with a linear gradient of 0.15 -0.3 M NaCI in a 10 column volume of20 mM Tris-HCI, pH 7.5) using an AKTA Explorer Systems (Amersham Bioscience). 13.
(15) Preparation of oxidized PDI (Oxi PDI) or reduced PDI (Red PDI) Reduced PDI was prepared by incubating purified PDI (2 mg) with 10 mM DTT in a final volume of 1 mL of 50 mM MES (pH 6.3) for 18 hours at room temperature. Separately, oxidized PDI was prepared by incubating 2 mg PDI with 2 mM GSSG in a final volume of 1 mL of 50 mM Tris-HCl (pH 7.5) for 18 hours at room temperature, and then passed through Bio-Gel P6 spin column (Bio-Rad Laboratories, Hercules, CA) to remove DTT or GSSG. Detection of thiol group by monobromobimane (mBBr) labeling and fluorescence detection Red PDI and Oxi PDI (100 pL of each) were incubated with 20 mM mBBr (10 pL) at room temperature for 30 min.. Excess mBBr was removed with a Bio-Gel P6 spin. column, and then PDI was subjected to SDS-PAGE and visualized with uv. Competitive binding assay to PDI. Purified histidine-tagged rat PDI was incubated with 100 nM. (125. I]T3 or 100 nM. [3H]BPA in a 50 mM Tris-HCl buffer (pH7.5), containing 150 mM NaCl for 120 min at 4 °C with varying concentration of test chemicals in a final volume of 0.5 mL. Unbound [3H]BPA was separated from bound [3H]BPA according to the method of Horiuchi et a1. (102) with some modifications.. Briefly, after incubation, proteins were. precipitated by adding equal volumes of 50 mM Tris-HCl buffer (pH7.5), containing 12 % polyethylene glycol 6,000, 150 mM NaCl, and 200 mM ZnCb to the reaction mixture, followed by centrifugation at 15,000 x g for 5 min at 4°C.. The supernatant. was aspirated, and then the pellet surfaces and insides of tubes were washed twice with 1.5 mL 50 mM Tris-HCl buffer (pH7.5 ), containing 6 % polyethylene glycol 6,000, 150 mM NaCl, and 200 mM ZnCb The pellets were dissolved with 0.1 N NaOH. Radioactivity was measured using a B counter (LS 6500, Beckman Coulter, Inc., Fullerton, CA) Isomerase activity of PDI Isomerase activity was assayed as described previously (19).. Briefly, reduced and. denatured RNase A (8 pM) was incubated with 1 pM PDI in 100 mM phosphate buffer (pH 8.0) containing 4.5 mM cytidine 2',3'-cyclic monophosphate (cCMP), 2 mM EDTA, 1 mM GSH, and 0.2 mM GSSG at 25°C in a final volume of 0.2 ml. started by adding reduced and denatured RNase A.. The reaction was. Changes in absorbance at 296 nm. were monitored with a spectrophotometer, Multiskan Spectrum instrument (Thermo Labsystems, Boston, MA). Chaperone activity of PDI Thermally induced protein aggregation of ADH was measured as the turbidity of the 14.
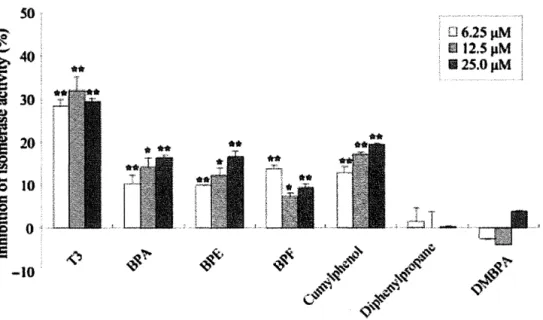
(16) solution monitored at 360 nm on a spectrophotometer at 40°C.. Aggregation was. initiated by the addition of 11 f.1M ADH to 50mM HEPES-NaOH buffer (pH7.0) containing 10 mM DTT, 1 p.M PDI, and test chemicals in a final volume of 200 p.1. All aggregation experiments were normalized by control experiments, in which aggregation with the substrate alone was defined as being 100 % of the total substrate aggregation.. II. Results 1-1. The required structure ofBPA derivative binding to PDI To understand the relationships between the chemical structure of BPA derivatives and binding activity to PDI, competitive binding assay were performed using [3H]BPA in the presence of BPA derivatives (Fig. 1-1-2).. Cumylphenol inhibited [3H] BPA. binding to PDI as well as BPA, but diphenylpropane have low inhibitory effect (Fig. 1-1-3A), suggesting that the hydroxyl group of phenyl ring(s) is required for binding activity.. Inhibition of [3H] BPA binding to PDI by BPE and BPF is almost identical to. BPA (Fig. 1-1-3B), suggesting that substitution of the methyl group at the center ofBPA did not contribute to the binding affinity to PDI.. On the other hand, DMBPA did not. inhibit [3H] BPA binding to PDI, so the hydroxyl group of phenyl ring(s) is required for binding affinity. Effects of changes In the structure of PDI Isomerase domain on binding of BPA derivatives The binding capacity of BPA to oxidized (Oxi) and reduced (Red) PDI was investigated.. Using Walker's method (103), I prepared Oxi or Red PDI.. Formation of. a free thiol group was confirmed from bands which reacted with thiol-specific fluorescent probe mBBr.. Values were normalized by the intensity of bands which. detected by CBB staining (Fig. 1-1-4A). As a result, BPA binding activity ofOxi PDI or Red PDI was almost identical to WT PDI (Fig. 1-1-4B), and Kdvalues ofBPA binding to Oxi and Red PDI were 8.49 and 16.87 p.M, respectively (Fig. 1-1-4C).. These results. indicated that BPA binding activity was not change by the redox state of the isomerase active site.. I next examined whether the Cys residue of the isomerase active site. contributed to BPA binding activity. measured isomerase activity.. I purified three PDI mutants (Fig. 1-1-4D), and. As a result, BPA binding activity of all PDI mutants. (aa'm, am, and a'm) was almost identical to WT PDI (Fig. 1-1-4D).. These result. indicated that the Cys residue of the isomerase active site did not contribute to BPA binding activity. Inhibition of isomerase activity ofPDI by BPA derivatives To understand the relationships between the chemical structure of BPA derivatives 15.
(17) and the inhibitory effect on PDI -mediated isomerase activity, isomerization of reduced RNase A to the active form was spectrophotometrically detected using cCMP as RNase substrate in the presence ofBPA derivatives (Fig. 1-1-5).. Each chemical was added to. reaction buffer at a final concentration of 6.25, 12.5, and 25.0 p.M. Both BPA and Ts effectively inhibited isomerase activity as previously reported (19).. BPE and BPF also. inhibited isomerase activity, suggesting that substitution of the methyl group at the center of BPA did not contribute to the inhibitory effect.. Cumylphenol inhibited. isomerase activity, but diphenylpropane did not, suggesting that the hydroxyl group of the benzene ring is essential for the inhibitory effect of isomerase activity.. These. results indicated that inhibitory effect correlate with binding affinity to PDI. Isomerase activity of Oxi or Red PDI and PDI mutants . . I next performed an isomerase assay of Oxi PDI and Red PDI.. GSSG promoted. protein disulfide formation with PDI, and GSH promoted protein reduction to dithiol with PDI(104), so, to maintain the redox state of PDI, I performed an assay in the presence or absence of GSSG and GSH. Oxi PDI, and Red PDI have similar activity for isomerization of reduced RNase Ain spite of the absence ofGSH and GSSG (Fig. 1-1-6A), suggesting that the redox state is not important for isomerase activity. contribution of each domain to isomerase activity.. I confirmed the. I found that am and aa'm have. defects in their of RNase A oxidation ability compared with WT PDI (Fig. 1-1-6B).. In. contrast, the activity of a'm was almost equal to WT PDI. Inhibition of chaperone activity ofPDI by BPA derivatives PDI also acts as a molecular chaperone. I examined the effect of BPA derivatives on chaperone activity of PDI.. Thermal aggregation of ADH by PDI was detected by. measuring the time-dependent change in turbidity (scattering) at 360 nm in the presence of test chemicals. Aggregation of ADH was suppressed by PDI (data not shown).. The chemicals used in this study have low effects on chaperone activity of PDI. (Fig. 1-1-7). Contribution of PDI isomerase domain to chaperone activity Next, to assess the contribution of the redox state of the active site to the chaperone activity, I determined chaperone activity of Oxi PDI and Red PDI.. To eliminate the. reduction ofPDI by DTT, chaperone assay was performed in the absence ofDTT, which was used as a promoter for aggregation of ADH.. There were no significant differences. between Oxi PDI and Red PDI in chaperone activity (Fig. 1-1-8A).. These results. suggest that the redox state of Cys residues of the PDI isomerase domain is not important for its chaperone activity.. Moreover, to assess the contribution of Cys. residues of active site to chaperone activity, I determined chaperone activities of PDI 16.
(18) mutants.. The mutant, am, revealed lower chaperone activity than WT PDI, although. chaperone activity of a'm and aa'm were lower than that by WT (Fig. 1-1-8B). These results strongly suggest that the PDI a' domain plays an important role in the chaperone activity. Effects of endogenous tyrosine or tryptophan derivatives on isomerase activity of PDI Monoamine derivatives are mainly produced from tyrosine phenylalanine, or tryptophan, and act as neurotransmitter or hormone. produced from tyrosine.. T3 and T 4 are also. Some monoamine derivatives have phenolic structure;. therefore, effects of them on isomerase activity were tested.. Isomerase activity of PDI. was detected in the presence of 20 p.M monoamine derivatives (Fig. 1-1-9). Slight inhibitory effects of tyrosine or tryptophan on isomerase activity were seen, but some derivatives such as Ts,. T4,. 3,4-dihydroxyphenylacetic acid,. deoxyepinephrine,. octop amine , 6-hydroxymelatonin effectively inhibited the PDI activity.. From these. results, not only phenol structure, but also hydrophobicity is considered to be required for the inhibitory effects on isomerase activity of PDI.. ill. Discussion 1-1 Previously, PDI was isolated from the rat brain as a BPA binding protein (19).. In. this study, to identify the basic PDI function involving the BPA binding activity, I performed experiments using six BPA derivatives, and oxidized or reduced PDI and three kinds of PDI mutants. From the result of competitive binding assay, I found that the hydroxyl group of benzene ring(s) is required for binding affinity and substitution of center the methyl group did not contribute to binding affinity.. These results suggest that BPA. derivatives bind to PDI via a hydroxyl group, and inhibit isomerase activity.. BPA has. been reported to interact with estrogen -related receptor gamma (ERRy) via an important hydrogen bond between a phenol group of BPA and Arg316 of ERRy, and a collaborative hydrogen bond between phenol group and Glu 275.. I considered that BPA. interacts with PDI via a hydrogen bond between the phenol group of BPA and hydrophilic amino acid ofPDI as well as ERRy (105,106). ERRy is identified as a high affinity nuclear receptor for BPA and its Kd value is 5.5 nM (105).. Biological function ofERRy is not well understood and effects ofBPA on this. receptor are not known.. On the other hand, the Kd value of rat PDI for BPA was 22.6. p.M (19) and much higher than that ofERRy.. However, PDI has been reported to be a. high capacity and low affinity hormone reservoir of the endoplasmic reticulum (8).. So,. it is considered that BPA disrupts PDI functions in the cytosol or endoplasmic reticulum, 17.
(19) but not in the nuclear.. PDI expression is induced by unfolded protein response, and. inhibits aggregation of protein by its isomerase or chaperone activity (59,61). In order to investigate the relationship between the redox state of the isomerase active site of PDI and BPA binding activity, I performed a BPA binding assay using oxidized or reduced PDI. BPA binding activity was not changed by the redox state of PDI, suggesting that reduction or oxidation of isomerase active site does not change the structure of the chemical-binding site of PDI.. Using a series of PDI mutants, I found. that BPA binding activity was also not changed by substitution of Cys residues of isomerase active site, suggesting the Cys residues of the isomerase active site are not required for BPA binding activity.. These results indicated that BPA doesn't bind to the. isomerase active site of PDI. In an isomerase assay with BPA derivatives, I found that the structural requirement for an inhibitory effect on PDI mediated isomerase activity is identical to the binding affinity to PDI. These results suggest that the inhibitory effect correlate with binding affinity to PDI.. A previous report indicated that BPA degraded hypoxia inducible. factor 1 u (HIF-lu) via an unknown pathway and inhibited the hypoxia response, and that this phenomenon required two central methyl groups but not two phenol groups (101).. However, in this study I found that central methyl groups in BPA did not. contribute to the binding affinity to PDI and inhibitory effect on PDI -mediated isomerase activity, so the biological effect ofBPA is not a simple pathway. Next, to indentify the contribution of the redox state of isomerase active sites or each Cys residue, I examined the isomerase activity of oxidized or reduced, and PDI mutants. I found that the redox state is not important for isomerase activity, while, from the experiments using three PDI mutants, I found that am and aa'm have defective RNase A oxidation ability compared with WT PDI.. Lyles et a1. (107) reported that the. isomerase active site of a' domain contributes more to apparent steady-state substrate binding, and the active site of a domain contributes more to catalysis at saturating concentration of substrate using human PDI.. Because I used a lower concentration of. PDI than Lyles's report, RNase A might saturate and the activity of a domain was more detectable than that of a' domain. PDI is also known as a molecular chaperone so I examined the effect of BPA derivatives on chaperone activity of PDI. As a result, any chemicals used in this study did not affect chaperone activity ofPDI.. I considered that ADH recognition by PDI was. not affected by BPA and was different from RNase.. I next investigated the. participation of the redox state or mutation of isomerase active sites in chaperone activity.. First, I found that redox state of isomerase active sites is not required for. 18.
(20) chaperone activity.. On the other hand, chaperone activity was inhibited by the. substitution of at least a domain.. Whiteley et a1 (108) indicated that the capacity for. immunoglobulin solubility of a'm was lower than am in the study using Spodoptera. frugiperda (Sf-9) cells.. My result was reflecting in vivo situation.. Puig et a1. (62). reported that a 21 kDa C-terminal fragment ofPDI also has chaperone activity; however, the chaperone activity of this fragment was lower than full-length of PDI.. Taken. together, the activity of a domain was not essential for chaperone activity, but was required for the facilitation of chaperone activity; therefore, chaperone activity is not dependent on isomerase activity. Several reports indicate that accumulation of misfolded protein by disfunction of PDI induce neurodegenerative diseases (56,57).. S-Nitrosylation of PDI inhibits. catalytic activity of PDI and links protein misfolding to neurodegeneration in Parkinson's or Alzheimer's disease (56,57).. In addition, PDI is expressed in the. embryonic midline and is required for left/right asymmetry (54).. These findings. suggest that PDI play an important role in the function of brain and central nervous systems, and I can considered that BPA affects these functions of PDI and disrupts the formation of neuronal systems by accumulation of misfolded proteins. The present study indicates that phenolic compounds specifically bind to PDI and inhibit isomerase activity.. Moreover, the inhibitory mechanism of isomerase activity. was not by direct binding to isomerase active sites, so I must identify a BPA binding site in PDI.. In fact, cumylphenol, BPE and BPF are also widely used as flame retardants. in numerous products (109), so I am exposed to these chemicals in daily life.. This. study will provide useful information to predict the biological effects of chemicals. Moreover, this study also provides useful information to predict the biological effects of chemicals and structural studies of PDI with the function of BPA or T3 binding.. 19.
(21) Figures 1-1. IV. \VT. ~a. ~a 7. Hb'. "Hb. / CGHC. aa'111. Hb. 7. Hb'. " Hb ~a 7 ". Ha' 7. ~a. 7. ". i\GH...~. Hb'. Ha' 7. ~. "lB". CGHC. . J.\. . GH..J.\.. a'111. "~. CGHC:. 4~GH;\. am. ~. Ha'. Hb. Hb'. ". Ha' 7. oLL\.GHA. C~OHC. Fig. 1-1-1 Domain structures of wild and mutant PDls. HO. -< ) em< lID. )-OH. IDCO. -< ). Bisphello1 . ~ . (BPA). j <fID< ~ \\ ~ >. t-. OH. Bisphenol E (BPE). HO. -<. ) - OCID. CID Dimetllylbisphenol A (D~,mpA). ;). HO ' \. lID(. Cluuylphenol. )-! <. )-OH. Bisphenol F (BPF) Fig.I-I-2 Structures ofBPAderivatives. Diphenylpropane. 20.
(22) (A). (B). ...... BPA. ...... BPA 4-BPE. -G-eumylphenol 120. 120. --ts- Diphenylpropane. 110. 110. -100 ~. -100. 90 .S -a .5 .c 80. OIl .5 90. ~ ~. e.., co. ]. .c 80 Q). Q). > 'p. .S2. ~. -lrBPF. *DMBPA. .~. 70. ~ ~. 60. 70 60. 50. 50. 40. 40 -6. -7. -8. -5. -4. -3. -7. -8. -3. -4. Chemical (log M). Chemical (log M). Fig. 1-1-3. -5. -6. Competitive inhibition of [3H]BPA binding by BPA derivatives.. 0.1 mg/mL. of purified PDI was incubated with 100 nM [3H]BPA in the presence or absetice of test chemicals at various concentrations for 120 min at 4°C.. Relative binding (%) is plotted. against log concentration (M) of unlabeled test chemicals. Values with [3H]BPA only were set at 100 %. A. -. 1.2 1.0. go §'. ~i 0.8 ::t: OI:! 0.6. -. .5 .J:J. -. ~ ~ 0.4. -. -.f. B 120 '$. "Q. tfj ....... ~. Data shown are the means of duplicate determining.. 60 40 20. ~. ;S 0.2. > '.Q .!! <U. n.d.. 0.0 \VT. '". Red. Oxi. 100 80. 0. wr. Red. Oxi. mBBr. D. caB. C. 3.0. OJ). 2.5 2.0. c. :.a-. ]i. +-Oxi -G- Red. •. -. «I )(. WT. ......... ~. aa'm. am. ~. a'm. ~ 120 bO. <'0 1.5. Q..-. -. 62 -47.5 -~ (kDa). ~. 1.0. :.c. ~- 0.5. 100 80 60. ~ 40 ':;3. '.:!..... 0.0 0. 5. 10. 15. 20. ..5 20. ~. 25. PH]BPA(~). 21. o. ~~~~~~~~~~~--~. WT. &afm. am. aIm.
(23) Fig. 1-1-4. Binding assay for Oxi or Red PDI and PDI mutants.. The redox state of Oxi PDI and Red PDI were detected usind mBBr (A). and oxidized PDI were treated with 2 mM mBBr.. Reduced PDI. Labeling was carried out for 30 min. at room temperature, excess mBBr was removed with Bio-Gel P6 spin column, and then samples were separated by SDS-PAGE and visualized with UV. subsequently stained with eBB.. The gel was. Binding activity of Oxi and Red PDI (B).. 2 nM of. Oxi and Red PDI were incubated with 100 nM [3H]BPA for 120 min at 4 °e. Values are the mean ±SD of three replicates.. Kinetic analysis of Oxi and Red PDI with BPA (e).. PDI was incubated with six concentrations of [3H] BPA. Nonspecific binding was determined in the presence of 1 mM unlabeled BPA, and specific binding was calculated by subtracting the nonspecific binding from total binding.. Binding activity of PDI. mutants (D).. Samples were separated. PDI mutants were constructed as described.. by SDS-PAGE and detected by eBB staining. Refolding assay of reduced RNase A was performed in the presence of PDI mutants. 100 nM [3H]BPA for 120 min at 4°e.. 5 nM of PDI mutants were incubated with. Values are the mean ± SD of three replicates.. so 6.25J1M 12.SpM. 'It.. 112S.0J1M. -10. Fig. 1-1-5. Effect of test chemicals on PDI -mediated isomerase activity. Refolding assay of reduced RNase A was performed in the presence of test chemicals at various concentrations.. The inhibition is relative shown to that observed in the. absence of test chemicals as 0%.. Values are the mean ±SD of three replicates.. *p<0.05, **p<O.Ol compared with control.. 22.
(24) -e. B 120. A 180 l60 .~ 140 > "ij 120 : 100 G 80 60. "0'. ~100. b :~ 80 u. ~ ~. 60. i. -~ 40. 40. j. 20. 20. 0. OSH OSSG. Fig. 1-1-6. t:,,~:t. + +. 0. L. +. WT. aa/m. am. am. +. Isomerase activity of Oxi or Red PDI, and PDI mutants.. Refolding assay of reduced RNase A was performed in the presence of Oxi PDI or Red PDI (A), and PDI mutants (B).. Isomerase activity of oxidized or reduced PDI was. measured in the presence or absence ofGSH or GSSG.. 06.25 JIM pM 1125.0 JIM II 12.S. -10. Fig. 1-1-7. Effect of test chemicals on PDI -mediated chaperone activity.. Effect of test chemicals on chaperone activity of PDI.. Native ADH was incubated in a. spectrophotometer at 40°C in 50 mM HEPES, pH 7.0, with or without PDI and various concentration of chemicals.. Thermal aggregation was monitored by measuring. absorbance at 360 nm every minute.. The rate of ADH aggregation (LlAImin) was. normalized to the rate observed in the absence of test chemicals as 0 %. mean ± SD of three replicates.. 23. Values are the.
(25) --. B. A 120. -. ~. 100. ~ c-. 80. «. 60. a. 0. -. = .9. Ie WT I I I. I·II~ I. :c 0 c-. !~~.·~·aml. L...~ ..~~I!!J. o. .~. I, 40. j. r~"··'''-~-- -----"-~~- ~'1. ~ 120~ ~. «~. 20. o. -POI. WT. Oxi. Red. 100. 200. 300. 400. 500. 600. POI (nM). Fig. 1-1-8. Chaperone activity of PDI mutants, and Oxi or Red PDI.. Native ADH was incubated in 50 mM HEPES, pH 7.0, with various concentrations of PDI at 40 DC. To determine the involvement of the redox state ofPDI, chaperone activity of Oxi and Red was assessed in the presence or absence of DTT (A).. The rate of ADH. aggregation (L1AImin) was normalized to the rate observed in the absence of PDI as 100 %.. Values are the mean ± SD of three replicates.. 24.
(26) --. ';f.. "-". .c. 45. :~ 40 u. *. -Cd. 35 ~ 30 § 25 .~ ~ 20 0 ~ 15 10 C 0 .t: 5 ~ 0 (!) til. **. (!). -. ~,..". Fig. 1-1-9. **. *. ~. Effect of amine derivatives on PDI -mediated isomerase activity. Refolding assay of reduced RNase A was performed in the presence of 20 pM amine derivatives.. The inhibition is relative shown to that observed in the absence of test. chemicals as 0%.. Values are the mean ±SD of three replicates.. compared with controL. 25. *p<0.05, **p<0.01.
(27) 1-2 Different biological effects of polybrominated diphenyl ethers (PBDEs) with different chemical structures 1. Materials and methods 1-2. Materials PBDEs were provided from Shigeyuki Kitamura (Department of Health and Phamaceutical Sciences, Nihon Pharmaceutical University) which were synthesized by following methods.. BDE47 was synthesized according to the method of Teclechiel et al.. (110), and BDE85 and BDE99 following the method of Orn et al. (111).. 4'-HO-BDE17,. 4'-MeO-BDE17, 4'-HO-BDE42, and 4'-HO-BDE49 were synthesized according to the method of Marsh et al. (112).. Synthesized PBDEs were >95 % pure.. purchased from Wako Pure Chemical Industries Ltd. (Osaka, Japan).. BPA was. Ribonuclease A. (RNase A) type III from bovine pancreas was purchased from Sigma-Aldrich Corp (St. Louis, MO).. BPA and PBDEs were dissolved in DMSO (Wako) at 10 mM to make stock. solutions and stored at -20 DC. All other chemicals used were purchased from Wako. In this experiment, 0.1 pM- 20 pM PBDEs in DMSO solution (final concentration of DMSO at 0.1%- 0.2%) were added to assay system or cell culture medium. samples, the same amount of DMSO were also added as vehicle.. In control. Ts was dissolved in. 0.1 M NaOH at 10 mM to make stock solutions and stored at -20 DC.. 10 pM or 10 nM. Ts in NaOH solution were added to assay system or cell culture medium. concentrations of NaOH were 0.1 mM or 0.1 pM.. Final. In control samples, the same. amounts of NaOH were also added as vehicle. Purification of rat histidine-tagged PDI Rosetta gami™ B E.coli cells (Novagen, Madison, WI) transformed with pQE-80L encoding. a. rat. histidine-tagged. PDI. were. grown. at. 37. DC. in. 2x. yeast. extract-tryptone-rich medium containing 0.1 mg/ml ampicillin, 0.034 mg/ml kanamycin, 0.02 mg/ml tetracycline, and 0.015 mg/ml chloramphenicol, and protein expression was induced by adding 1.0 mM isopropylthio-B-O-galactoside.. After an additional. cultivation for 8 h, E. coli cells were harvested and lysed in a lysis buffer (50 mM NaH2P04, (pH 7.5), containing 300 mM NaCI, 1.0 mg/ml lysozyme and 20 mM imidazole) for 60 min at 4 DC.. The cell lysate was sonicated for 1 min.. The lysate. sample was centrifuged at 50,000 x g for 30 min, and the supernatant was loaded onto a Ni-NTA Agarose column (Qiagen, Hilden, Germany). After the column was washed with the lysis buffer, the protein was eluted with lysis buffer containing 250 mM imidazole. 26.
(28) Isomerase activity of PDI The assay of isomerase activity was performed as described previously(19). Reduced and denatured RNase A (1.25-200 pM) was incubated with 1, 2, 10, or 100 /lM PDI in 100 mM potassium phosphate buffer, (pH 8.0), containing 4.5 mM cytidine 2',3'-cyclic monophosphate, 2 mM EDTA, 1 mM glutathione, and 0.2 mM glutathione disulfide with 5, 10 or 20 pM of PBDE at 25°C. denatured RNase A.. The reaction was started by adding reduced and. The changes in absorbance of cytidine monophosphate at 296 nm. were monitored with a spectrophotometer, the Multiskan Spectrum Instrument (Thermo Labsystems, Boston, MA).. To draw Lineweaver-Burk plots (supplemental. data), the amount of active RNase was determined by the following formula, Et(active RNase) = 8Abs296/8t {Km(cCMP)(1+[CMP]t lKi (CMP»+[cCMP]t}. kcat[cCMP]t where Et is the concentration of total active RNase; Km(cCMP) is the half saturation constant for cCMP (8 mM); m(CMP) is the inhibitor constant by CMP (2.1mM);. kcat. is. the apparent unimolecular rate constant of active RNase (14.3 min-I), Abs296 is the absorption coefficient of cCMP and CMP at 296 nm (0.38 (CMP)/mMI cm cell) (113). Pull down assay A purified histidine-tagged bb' (2 pM) and denatured RNase A (4 pM) were incubated for 30 min at 25°C, in 50 mM sodium phosphate buffer (pH7.5) in the presence or absence of 5, 10, and 20 pM of 4'-OH-BDE17, and pull down was performed with Ni-NTA agarose. silver-staining.. Precipitated proteins were analyzed by SDS-PAGE and detected by CNBr-activated Sepharose 4B (1 m!) (GE-Healthcare) was reacted. with reduced and denatured RNase A (0.5 mg) according to the manufacturer's instructions.. Histidine-tagged bb' (1 pM) and RNase A-binding Sepharose were. incubated in 50 mM Tris-HCI (pH 7.5) containing 0.3 M NaCI and 0.1 % NP40 in the presence or absence of 10 or 20 pM of 4'-OH-BDE17 at 25°C for 30 min.. Proteins. precipitated with RNase A-binding Sepharose were analyzed by SDS-PAGE and detected by CBB staining.. CNBr-activated Sepharose which was reacted with ethanol. amine was used as a control. Cell culture and transfection The rat pituitary tumor cell line GH3 was provided by Health Science Research Resources Bank (cell number JCRB9047, Osaka, Japan).. Cells were maintained in. Ham's F-10 medium containing 15% horse serum and 2.5% fetal bovine serum, and incubated at 37°C in a humidified atmosphere of 5% C02. PDI were cloned as described previously(66).. Cells which over-expressed. The mammalian expression vector. pcDNA3.1 (Invitrogen, San Diego, CA) containing PDI cDNA or an unmanipulated 27.
(29) vector was transfected by a standard lipofection method using GenePORTER2 Reagent (Genlantis, San Diego, CA).. Forty-eight hours later, stably transfected cells were. selected in the medium containing 300 pg/ml of G418, and cloned. was performed as follows. (Tokyo, Japan).. RNA interference. siRNA for rat PDI was purchased from Funakoshi Co Ltd. It was a cocktail of double strand RNAs (5'-cugcaaaacugaaggcagatt-3',. 5'-ggaaaaacuuugaggaggutt-3', and 5'-ggauaaacugggagagacatt-3'). The cocktail (10 nM each) or control siRNA was mixed with 5 pL of siLentfect (Bio-Rad Laboratories) in a final volume of 2.5 mL ofT3-depleted (Td) medium (114) and transfected into GH3 cells in 3.5-cm dishes (2X10 5 cells).. After 6 h, the medium was changed to new Td medium.. Two days later, GH3 cells were re-transfected by the same method, and after an additional 2 days, a third transfection was carried out. Induction of GH GH3 cells were cultured in 3.5-cm dishes at 2 x 10 5 cells/dish in Td medium for 24 hours to eliminate endogenous T3 in cells.. The culture medium was then replaced with. fresh Td medium containing test chemicals, and 24 hours later, cells were collected. GH protein in the cells was detected by Western blotting using anti-GH monoclonal antibody (Chemicon International, Temecula, CA). Isolation of RNA and reverse transcription -PCR GH3 cells were cultured in Td medium for 24 h, the culture medium was replaced with fresh Td medium containing test chemicals, and incubation was continued for 24 h. Total RNA was extracted from GH3 cells with Isogen (Nippon gene, Toyama, Japan). A reaction mixture containing 1 pg of RNA and 200 U of MMLV reverse transcriptase (Takara) was reacted according to the manufacturer's direction as follows: 10 min at 25°C and 60 min at 42°C, then 10 min at 70°C to stop the reaction.. The quantitative. real-time polymerase chain reaction (qRT-PCR) was performed using a Thermal Cycler Dice Real Time System Single TP850 (Takara, Shiga, Japan).. SYBR Primer Ex TagH,. 10 pmol of forward and reverse primer, and 1 pg of cDNA were mixed and RT-PCR was performed according to the manufacturer's instructions.. PCR was conducted at; 95°C. for 10 sec, and then 40 cycles of 95°C for 5 sec and 60°C for 20 sec.. Primers for rat GH. (Gene Bank Acc.No: GQ890681) were 5'-CTGCTGACACCTACAAAGA-3' (sense) and 5'-CAGTGTGTGCCTAGAAAGCA-3' (antisense). CAA24528). were. Primers for rat B-actin (Acc.No:. 5'-CTACAATGAGCTGCGTGTGG-3'. 5'-TGAGGTAGTCTGTCAGGTCC-3' (antisense).. (sense). and. Data obtained by RT-PCR were. analyzed by the second Derivative Maximum method according to the manufacturer's directions. Luciferase reporter gene assay 28.
(30) Luciferase reporter gene assay was performed using the pGL3 vector which includes rat GH 5'-fianking region (GenBank accession no. X12967) containing thyroid hormone response element (TRE) (from -1803 to +5) as previously reported(66). Statistical analysis The statistical significance was analysis using student's t-test.. 11. Results 1-2. Inhibitory effects of PBDEs on isomerase activity of PDI In this study, I used BDE47, 4'-OH-BDE17, 2'-OH-BDE28, 4'-MeO-BDE17, BDE85, 4'-OH-BDE42, 2-0H-BDE15, 2-MeO-BDE15, BDE99, and 4'-OH-BDE49 as test chemicals (Fig.1-2-1).. Isomerase activity was quantified as RNase A activity which. was resumed by an oxidative folding of RNase A from its reduced state.. 10 and 20 pM. of 4'-OH-BDE17, 4'-OH-BDE42, 4'-OH-BDE49, 2'-OH-BDE28, and 2-0H-BDE15 effectively inhibited the isomerase activity of PDI (Fig. 1-2-2A). Moreover, inhibitory effects of 4'-OH-PBDEs were stronger than those of 2' (or 2)-OH-PBDEs (Fig. 1-2-2A), suggesting an importance of the location of a hydroxyl group.. Apparently, a. substitution of the 4'-position is more vital in inhibitory effects compared to that of the 2' (or 2)-position. Endogenous PDI in the lumen of the endoplasmic reticulum could reach to considerable levels as high as 200 pM (115).. This prompted us to compare inhibitory. effects of 20 pM 4'-OH-PBDE17 on PDI at concentrations of different orders of magnitude, 2, 10, and 100 pM. As a result, there was little difference in an inhibitory effect of 4'-OH-PBDE17 on three different concentrations of PDI, and in all cases, 20-30% of PDI activity was inhibited (Fig. 1-2-2B).. Of particular, a significant. inhibition to 100 pM PDI revealed that PBDEs could be a functional inhibitor of PDI under physiological conditions. Inhibition mechanism of PBDEs on isomerase activity of PDI PDI consists of a, b, b', a' domains and c region.. I constructed an N-terminal. fragment (ab), two C-terminal fragments (b'a'c and a'c), and investigated their isomerase activity in the presence ofBDE47, 4'-OH-BDE17, or T3 (Fig. 1-2-3).. BDE47. did not inhibit the isomerase activity of any of four fragments. In contrast, 4'-OH-BDE17 inhibited activities of a full-length PDI and a C-terminal fragment b'a'c, while this compound did not inhibit isomerase activity of fragments ab and a'c (Fig. 1-2-3). These results strongly suggest that the target of 4'-OH-BDE17 to inhibit the isomerase activity is the b' domain ofPDI.. To assess the effects of 4'-OH-BDE17 on the. interaction between the b' domain and RNase A, I performed a pull down assay with the 29.
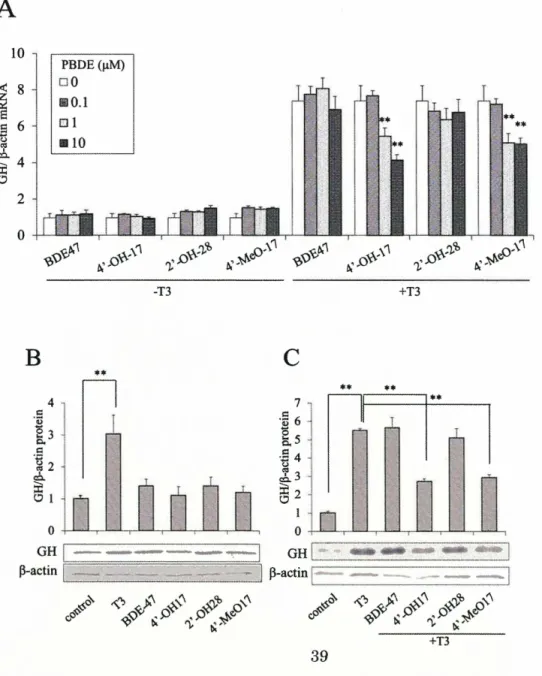
(31) bb' fragment of PDI and the reduced RNase A (Fig. 1-2-4A). His-tag was expressed in E coli. and purified.. The bb' fragment with a. The bb' fragment and denatured RNase. A were mixed in the presence or absence of 4'-OH-BDE17, and pull down was performed with Ni-NTA Agarose.. As a result, the amount of RNase A precipitated by Ni-NTA. Agarose with bb' fragment was reduced as concentration of 4'-OH -BDE 17 was increased (Fig. 1-2-4A).. In contrast, when the bb' fragment was mixed with reduced RNase A. immobilized on CNBr-activated Sepharose, the bb' fragment was precipitated with RNase A-immobilized Sepharose resin (Fig. 1-2-4B). Amount of bb' was decreased in the presence of 4'-OH-BDE17 suggesting that 4'-OH-BDE17 inhibited the interaction of the bb' with RNase A.. Kinetic parameters for isomerase activity of the full-length PDI. was measured in the presence of two different concentrations (5 and 20 p.M) of 4'-OH -BDE 17 using the various concentration of reduced RNase A as a substrate. Lineweaver-Burk plots indicated that 4'-OH-BDE17 changed the values of Km but. Vmax,. value was maintained relatively stable, strongly suggesting that the relation between 4'-OH-BDE17 and reduced RNase A is competitive on the substrate binding site of PDI (Fig. 1-2-4C). The effects ofPBDEs on GH production I previously found that BPA disrupt GH production of GH3 cells (66).. This finding. have prompted us to investigated the effects of BDE47 (parent compound), 4'-OH-BDE17, 2'-OH-BDE28 and 4'-MeO-BDE17 on the levels of GH mRNA accumulation in GH3 cells which produce GH by stimulation ofT3 (Fig. 1-2-5A). of the PBDEs changed GH mRNA levels in the absence of T3. have an agonistic effect on GH expression.. None. Thus, PBDEs may not. In sharp contrast, GH mRNA levels were. significantly stimulated in the presence of T3 and this stimulated level was reduced by 4'-OH-BDE17 and 4'-MeO-BDE17 in a concentration-dependent manner (Fig. 1-2-5A). I next tested the protein levels of GH protein in GH3 cells in the presence or absence of BDE47 and its derivatives, 4'-OH-BDE17, 2'-OH-BDE28 and 4'-MeO-BDE17 or T3 (Figs. 1-2-5B and 1-2-5C). The level of the GH protein was only stimulated in the presence of T3 but any ofBDE derivatives were inert in the absence ofT3 (Fig. 1-2-5B).. In contrast,. in the presence ofT3, the stimulated level ofGH protein was reduced by 4'-OH-BDE17 and 4'-MeO-BDE17 (Fig. 1-2-5C). GH release of PDI -over-expressing and PDI -knockdown cells PDI binds to T3 and is thought to be a reservoir of thyroid hormone in cells(8).. If. 4'-OH-BDE17 bind to PDI and competes with T3 on PDI, 4'-OH-BDEs may affect the level of free T3 and thus may change GH expression.. Also, previous study. demonstrated that over-expression ofPDI reduced the level ofT3-stimulated expression 30.
(32) of GH, suggesting a function of PDI as a modulator of free Ts levels and hence the levels of GH protein in GH3 cells (66).. To assess the toxicity of PBDEs on PDI function in. Ts-mediated GH production, I investigated the effects of PBDEs on GH expression in PDI-over-expressing and PDI-knockdown-GH3 cells (Fig. 1-2-6).. Over-expression of. PDI, which was confirmed by western-blot analysis (Fig. 1-2-6A), significantly reduced Ts-stimulated level of GH expression as described previously(66) (Fig. 1-2-6A).. On the. other hand, knockdown ofPDI by RNAi (which was confirmed by western blot analysis in Fig 1-2-6B) did not cause a significant increase in GH production (Fig. 1-2-6B). both. over-expressing. and. knockdown. cells,. two. PBDEs,. 4'-OH-BDE17. In and. 4'-MeO-BDE 17, efficiently inhibited GH expression independently of PDI levels, showing the same profile to that in intact cells (Figs. 1-2-6A and 1-2-6B). These results suggest that inhibitory effect of 4'-OH-BDE17 and 4'-MeO-BDE17 on GH expression is not directly mediated by PDI. To clarify whether or not the inhibitory effect of 4'-OH-BDEs on GH expression is related directly to TR and the TR -mediated transcription control, GH promoter activity was measured with luciferase reporter assay in the presence or absence of BDE47 or 4'-OH-BDE17 (Fig. 1-2-6C).. Both BDE47 and 4'-OH-BDE17 was completely inert in. modulating the activity of the GH promoter (Fig. 1-2-6C), suggesting that suppression ofGH by 4'-OH-BDE17 is not mediated by TR and TRE.. These data are in agreement. with the report by Kojima et.al.(82) that 4'-OH-BDE17 and 4'-MeO-BDE17 did not affect TR transcription via TRE.. On the other hand, GH promoter activity and its. stimulation by Ts were suppressed by over-expression of PDI, suggesting that PDI may directly act on TR regulated promoter to repress it (Fig. 1-2-6D). These results suggest that suppressive effect of 4'-OH -BDE 17 on Ts-stimulated levels of GH expression stimulated by Ts may occur independently of either PDI or TR. Contribution of ER to inhibition of GH release by PBDEs I also investigated the contribution of estrogen receptor (ER) to suppression of GH expression because 4'-OH-BDE17 and 4'-MeO-BDE17 have agonist activity of ER. To investigate the contribution of ER in T3-mediated gene expression, GH mRNA levels were determined in the presence of T3 and E2 or ICI 182,780 which is one of antagonistic inhibitor of ER. As a result, E2 suppressed the GH expression in the presence of T3 (Fig. 1-2-7A); on the other hand, ICI 182,780 enhanced the GH expression (Fig. 1-2-7B). From these results, I considered that agonist activity of ER suppress the T3-mediated GH expression.. ill. Discussion 1-2 31.
(33) PBDEs have two benzene rings linked by an ether bond, and several hydrogen atoms substituted with bromine atoms.. In the living cells, oxidizing enzymes such as P450. convert PBDEs to OH -PBDEs, and furthermore, methoxylated products (MeO-PBDEs) are also found(87,88).. In vitro experiment using human hepatocytes indicates that. 4'-OH-BDE49 is formed by CYP1A1, CYP2B1, and CYP3A1, and 4'-OH-BDE42 is formed solely by CYP2A2(116). The PBDE metabolites used in this study,. 4'-OH-BDE42 and 4'-OH-BDE49, were detected in glaucous gulls and polar bears as PBDE. metabolites(87).. 4'-OH-BDE17. metabolites of BDE47 in rats(117).. and. 2'-OH-BDE28 were. identified. as. Athanasiadou et al. (88) reported that. 4'-OH-BDE17, 4'-OH-BDE42 and 4'-OH-BDE49 were mainly detected as PBDE metabolites from sera of the children and hydroxylated PBDEs levels in human sera were 120 pg/g of lipid at the maximum concentration.. PBDEs were also detected in. human serum and adipose tissues, and have been found to be as high as 200 ng/g of lipid (118).. The concentration ofPBDEs in human milk was reported to be 956 ng/g of. lipid at maximum concentration (81).. In this study I used PBDEs at 1- 20 pM and. these concentrations are higher than those described above.. On the other hand, in a. case of BPA, accumulation in placenta is reported (4), suggesting that PBDE may also be concentrated in particular tissues such as placenta. Previous study found that BPA and hydroxylated PCB inhibit the activity of PDI (97-99).. In this study, the effects ofPBDEs on the functions ofPDI was investigated in. comparison with several metabolites of PBDEs, OH-PBDEs, which possess phenolic group (s), a presumable requisite for an interaction of these compounds to PDI (21,22). As I expected, both 4'-OH-PBDEs and 2' (or 2)-OH-PBDEs effectively inhibited the isomerase activity of PDI but non-hydroxylated PBDEs did not.. As has been. previously observed in the interaction between BPA and PDI (97,98), the present study showed that a hydroxyl group of PBDEs is likely relevant to the inhibitory effect of these compounds against isomerase activity of PDI.. Of particular interest is that 4'-. OH -PBDEs revealed stronger inhibitory effects on the isomerase activity of PDI than that of 2' (or 2)-OH-PBDE.. These results indicate the significance of a para-hydroxyl. group rather than ortho-hydroxyl group for inhibitory effect of PBDE metabolites on PDI activity.. Stronger inhibitory effects of 4'-OH -BDE 17 and 4'-OH -BDE42 compared. to 4'-OH-BDE49 suggest the importance of the number and the position of bromine substitutions for the function of OH -PBDEs. Apparently bromine substitution at 2'and 3'-position are more effective than that at 5'-position. Each of the a and a' domains contains one catalytically active Cys-Gly-His-Cys motif, and thiol groups of two cysteines playa role in thiol-disulfide exchange (24). 32. In.
(34) contrast, the band b' domains are redox inactive and provide the principal substrate-binding site (26).. I previously demonstrated that BPA bound to the a and b'. domains of PDI, and BPA-binding to the b' domain causes the inhibition of isomerase activity (119).. Similarly to this, the present study clearly showed that OH-PBDEs. interact with PDI via b' domain and inhibit the isomerase activity ofPDI, revealing the shared interaction mechanism between OH-PBDEs and BPA to PDI.. The pull down. efficiency of RNase A by bb' fragment was significantly reduced by 4'-OH-BDE17 in a concentration-dependent manner, suggesting the competition of this compound to RNase on PDI.. This is further supported by the results of kinetics analysis ofPDI with. 4'-OH-BDE17 as an inhibitor (Fig. 1-2-4C).. Inhibitory effect of 4'-OH-BDE17 was. stably expressed for a wide range ofPDI concentrations which reached to 100 pM in this study, strongly suggesting that tested concentrations of PBDEs (20 pM) is a physiologically functional.. It was shown in the present study that PBDEs also have some inhibitory effect on GH synthesis in GH3 cells although the mechanistic bases behind this are yet clear.. It. is noteworthy that the chemical compounds known to modulate the GH synthesis possess some structural similarity but function often totally opposite.. For instances,. hydroxylated PBDEs are similar to T3 in respect of having an ether structure and a halogenated phenol structure, whilst PBDEs had no ability to induce GH expression unlikely to T3.. Also in contrast to the fact that 4'-OH-BDE17 and 4'-MeO-BDE17. inhibited T3 mediated stimulation of GH expression, tetrabromobisphenol A, which has bromo groups in BPA structure, induces GH(95). PDI is known to be a T3 binding protein.. Our previous studies have shown that. over-expression of PDI reduces GH expression stimulated with T3, suggesting that excess PDI worked to absorb free T3.. However interestingly in this study, knockdown. of PDI also reduced the GH expression.. These data suggest that PDI may require. relatively narrow range of optimal concentrations to function properly in the GH production system in response to T3 in GH3 cells(66).. It is probable that the optimal. levels of PDI may be critical to maintain proper redox state and thus modulations of PDI levels beyond the preferable levels may cause the suppression of GH induction in T3 treated GH3 cells.. In fact, in pancreatic islets cells, PDI changes the insulin synthesis. by regulation of redox state of poly A binding protein (PABP), which regulates transcription of insulin (120). Notwithstanding, such GH production which may be supported by proper PDI concentrations is unlikely related to activity modulations of PDI by PBDEs since the 4'-OH-BDE17 treatment influenced little to the ratio ofGH production over those of 33.
(35) control cells.. Luciferase reporter assay of GH promoter further negated the. participation of TR in suppression of GH under 4'-OH-BDE17 treatment, which is in agreement with the data reported by Kojima et al.(82).. These results suggest that. PBDEs operate negatively for the GH expression independently with TR or PDI, although a detailed mechanism of PBDEs-mediated inhibition of GH induction is the issue waits to be studied.. Kojima et al (40) found that 4'-OH-BDE17 and. 4'-MeO-BDE17 behave as agonists for estrogen receptor (ER), which is also a potential modulator of GH expression.. Functional relationship between PBDEs and ER in GH. induction would also be an interesting issue for future study. In summary, effects of PBDEs on GH production have been suggested by some reports, but those of PBDEs metabolites have not been identified.. Here, I showed the. potential risk of PBDEs metabolites by demonstrating the potential functions of PBDEs metabolites as inhibitors of PDI and modulators of GH production, not only as hormone disruptants. It should be noted that the effect significantly differed depending on the brominated position or substituted group, strongly suggesting a hazardous nature specifically expressed due to their structure of PBDEs metabolites.. 34.
(36) Figures 1-2. IV. Br. Br. I~. I~. Br. Br. I~. I~. ~. ~. Br. OH. Br. Br. I~. I~. 0°'6 0°'6 0°'6 0°'6 ~. Br. ~Br. Br. BDE47. I~. OH. 4'-OH-BDE17. Br. I~. ~. ~Br. 2'-OH-BDE28. 0°,&Br o°,&Br B.o0'6Br Br. Br. Br. Br. OH. I~. Br. ~. ~Br. Br. BDE85. Br. Br. I~. I~. Br. oO'¢. #'-Br. Br. #. 2-0H-BDE15. ~. ~ OCH3. 4'-MeO-BDE17. 0'6Br OCH3. I~ Br~. Br. I~ I~ 0°~. OH. 4'-OH-BDE49. Structure ofPBDEs. BDE47,. 2,2',4,4'-tetrabromodiphenyl. 4'-OH-BDEI7,. ether;. 4'-hydroxy-2,2' ,4-tribromodiphenyl. ether;. 2'-OH-BDE28,. 2'-hydroxy-2,4,4'-tribromodiphenyl. ether;. 4'-MeO-BDEI7,. 4'-methoxy-2,2',4-tribromodiphenyl ether; BDE85, 2,2',3,4,4'-pentabromodiphenyl ether; 4'-OH-BDE42,. 4'-hydroxy-2,2',3,4'-tetrabromodiphenyl. ether;. 2-0H-BDEI5,. 2-hydroxy-4,4'-dibromodiphenyl ether; 2-MeO-BDE 15, 2-methoxy-4,4'-dibromodiphenyl ether;. BDE99,. 2,2',4,4',5-pentabromodiphenyl. 4'-hydroxy-2,2',4,5'-tetrabromodiphenyl ether. 35. ether;. I~ #. 2-MeO-BDE15. Br. BDE99. Fig. 1-2-1. Br. I~. ~OH. 4'-OH-BDE42. Br. ~. ~. Br. 4'-OH-BDE49,.
(37) A ~. ~. "-'. 40. C 35 .:>-. .-. **. o 10JlM WJ20JlM. **. ~. ~ 30. (1). !Zl. 25. C!:$. ~. .-§. **. 20. !Zl. ~. **. **. **. 15. 0. (1). **. **. ~. ~ 10. ~. **. *. * **. .-'-. 5. ~. . .0 ..d. 0. ~. ~. ~'b ~~. ~~. 'Ii ~'. 'V. ~~. SJ ~~O ~~O. B - 4'-OH-BDE17+4'-OH-BDE17 ----. 100 JlM. 0.08. 0.08. 0.08. ".,-',. ... _- ... §. 0.06. I. 00. 0\. N. 0.04 0.02. §. I. f/l. <Il. -<. I. -<. 0.04 0.02 -. 0. :~. t> C'\I <1). **. O. ~ 60. 60. 0. **. 120 .-6' 100. ·sg <1). 00. 00. ~. g. 60 40. C'\I. 20. o. 0. 40. 60. a>. **. 80 60. E 40 0. ~. 20. 20. Time (min). <1). <1). 40. 20 pM4'-OH-BDE17. 40. g 80. 00. ~. 0 20. Time (min). 120 .0 ·;;100 •.;::3. 100 80. 0.04 0.02. o -I. Time (min). 120. 0.06. 00. gj. o +----,-----,------, o 20 40 60. .0. §. 0.06 -. 00 0\ N. , ,,, , I. f/l. -<. I. I. ~. 20 0. +. + 36. +.
図
![Fig. 1-1-3 Competitive inhibition of [3H]BPA binding by BPA derivatives. 0.1 mg/mL of purified PDI was incubated with 100 nM [3H]BPA in the presence or absetice of test chemicals at various concentrations for 120 min at 4°C](https://thumb-ap.123doks.com/thumbv2/123deta/8221401.1281080/22.896.108.686.173.534/competitive-inhibition-derivatives-purified-incubated-presence-chemicals-concentrations.webp)


+7




関連したドキュメント
Then it follows immediately from a suitable version of “Hensel’s Lemma” [cf., e.g., the argument of [4], Lemma 2.1] that S may be obtained, as the notation suggests, as the m A
We have introduced this section in order to suggest how the rather sophis- ticated stability conditions from the linear cases with delay could be used in interaction with
While conducting an experiment regarding fetal move- ments as a result of Pulsed Wave Doppler (PWD) ultrasound, [8] we encountered the severe artifacts in the acquired image2.
[r]
Amount of Remuneration, etc. The Company does not pay to Directors who concurrently serve as Executive Officer the remuneration paid to Directors. Therefore, “Number of Persons”
6.大雪、地震、津波、台風、洪水等の自然 災害、火災、停電、新型インフルエンザを
[r]
[r]
