は じ め に
地球規模の急速な温暖化は1960 年代以降から進 み始めた(Hansen & Lebedeff 1987).これとは別 に,生態系を広範囲に変化させる主要因として, 北大西洋振動やエルニーニョ・南方振動に代表さ れる大気と海洋が連動して変動する現象がある
─ 原 著 論 文 ─
標識調査情報に基づいた 2000 年代と 1960 年代の
ツバメの渡り時期と繁殖状況の比較
出口智広 *・吉安京子・尾崎清明
公益財団法人山階鳥類研究所保全研究室 〒270–1145 千葉県我孫子市高野山 115 (2012 年 1 月 29 日受付;2012 年 6 月 6 日受理) キーワード:標識調査情報,ツバメ,渡り,繁殖,気候変動.Tomohiro DEGUCHI*, Keiko YOSHIYASU, Kiyoaki OZAKI. 2012. Comparison of Barn Swallow migration and breeding based on banding records from the 1960s and 2000s. Jpn J Ornithol 61: 273–282.
Abstract. There have been very few studies of the effects of global climate change on
migration and breeding of land birds in Asia (including Japan), compared with those in Europe and North America. In this study, we compared banding records of Barn Swallow
Hirundo rustica collected in Japan in 1961–1971 (the 1960s) with those in 2000–2010 (the
2000s) to consider climatic effects on their migration and breeding. From the frequency distributions of the numbers of adults, juveniles, and nestling birds released per day, the timing of rapid increase in adults was about half a month earlier in 2000s than 1960s, but the period of rapid decrease of adults did not vary between these two periods. The distribu-tion of juveniles 40 years ago was almost the same as at present. The timing of rapid increase in nestlings was about half a month earlier in the 2000s than in the 1960s. The ratio of juveniles to adults was slightly smaller in the 2000s than in the 1960s. These results indi-cate that both arrival and commencement of breeding in Barn Swallows were earlier in the 2000s than in the 1960s, but their departure did not vary between the two periods. Further-more, their annual fecundity was higher in the 1960s than in the 2000s. Changes in arrival date and commencement of breeding, were expected based on other available information, although it was anticipated that there would be effects of different habitats and study sea-sons between the two periods. Banding records were considered to be invaluable in assess-ing departure date and annual fecundity, because of a lack of other information suitable for the analysis of long-term change.
Key words: Banding records, Barn Swallow, Migration, Breeding, Climate Change. Division of Avian Conservation, Yamashina Institute for Ornithology, Konoyama 115, Abiko, Chiba 270–1145, Japan.
日 本 鳥 学 会 誌
Japanese Journal ofOrnithology © The Ornithological Society
of Japan 2012
* E-mail: [email protected] doi:10.3838/jjo.61.273
JOI:JST.JSTAGE/jjo/61.273
(Stenseth et al. 2002; Walther et al. 2002).この現象 には数十年周期の振動が見られ,近年では,1970 年代中期に世界的な影響が生じた位相反転が観測 された(山川 2005).このような地球規模の長期 的気候変動が陸生鳥類の渡りや繁殖に及ぼす影響 の有無は,欧米に生息する種について数多く報告 されてきたが(Cotton 2003; Hüppop & Hüppop 2003; Crick 2004; Marra et al. 2005; Gordo 2007),日本を 含むアジアにおける報告は著しく乏しい(Primack et al. 2009).
日本は,面積的には全世界の陸地の0.25%に過 ぎないが,南北に長く,起伏に富み,複雑な海岸 線を有することから,生物多様性が極めて豊かな 地域であり,鳥類では全世界の種の約6%にあた る542 種が利用する重要な生息地となっている (日本鳥学会 2000).国内では,1961 年から標識 調査が50 年間継続して行われており,計 409 種・ 470 万羽の標識放鳥が記録されている(山階鳥類 研究所 2002).そのうち,1972 年以降の記録は, 環境庁(省)が中心となり,委託事業として収集 されたものであるが,1961 年から 1971 年の記録約 15 万羽は,林野庁,米軍移動動物病理学調査所の 支援を受け,山階鳥類研究所が主体となって集め たものである(山階鳥類研究所標識研究室 1985). 日本を中心とする東アジアの鳥類を対象に,地 球規模での気候変動の影響を国土スケールで調べ るにあたり,国内における標識調査記録,特に大 規模気候変動の出現時期と近い 1961 年から 1971 年までの記録の学術的価値は高い.しかし,この 期間に集められた約15 万羽の記録は,紙媒体のま まの保管が利用を妨げ,その価値がほとんど発揮 されてこなかったが,近年その電子化が進み,解 析が容易になった. 本研究は,地球規模の気候変動が生じたこの50 年の間に,日本に訪れる鳥類の渡り時期と繁殖状 況の変化の有無を標識調査情報から記述すること を目的とし,その代表例としてツバメHirundo rustica を選んだ.欧米では,長距離を渡ること, および情報収集の容易さ等の理由から,気候変動 が渡り鳥に及ぼす影響を調べるのに適した材料と してツバメが選ばれ,飛来および繁殖時期の早期 化が多く報告されている(Møller 2002; Bulter 2003; Gordo & Sanz 2006; Rubolini et al. 2007; Sparks & Tryjanowski 2007; Sparks et al. 2007).そのため,日 本でも同様の状況が見られるかを調べることは興 味深い.本研究では,1961∼1971 年の 11 年間(以 下,1960 年代)の標識調査記録と 2000 ∼ 2010 年 の11 年間(以下,2000 年代)の標識調査記録を 用いて,ツバメの渡り時期と繁殖状況の変化の有 無を調べた. 方 法 1960 年代の標識調査は,再捕獲による標識個体 の移動分散の解明を主な目的としており,あらか じめ決められた場所で行われていた(山階鳥類研 究所標識研究室 1985).捕獲個体の記録内容は, 種名,性別,年齢(成鳥:1 歳以上の個体,幼 鳥:1 歳未満の個体,巣内雛,不明),標識情報, 放鳥日,放鳥場所名である.また,捕獲個体がす でに標識付きであった場合,半年未満の同一場所 での再捕獲,半年以後の同一場所での再捕獲,5 km 以上離れた場所での再捕獲の 3 カテゴリーに分 けて記録されている.その一方で,調査捕獲方法 に関する情報はなく,調査を行ったが捕獲放鳥さ れなかったことを示すデータも残されていない. 一方,2000 年代の標識調査は,移動分散の解明 だけでなく,個体群動態のモニタリングや,形 態・分類・生理学上の情報収集も調査目的に含ま れている(山階鳥類研究所 2008).記録内容には, 1960 年代と同じ項目に加え,調査捕獲方法と捕獲 できなかったことを示すデータも含まれている. ツバメを解析対象に選んだ理由は,1960 年代に 行われた標識調査の中で最もデータ数が多く(全 データ数の12%),放鳥場所が広く分散(20 都道 府県・64 箇所)していたためである.加えて,ツ バメの幼鳥は成鳥と比べて尾羽が短いという明瞭 な特徴があり(山階 1980),調査者が年齢を誤認 する可能性が小さいことも,本種を解析対象に選 んだ理由である. 本研究では,同一個体の再捕獲による解析結果 への影響を排除するため,再捕獲として記録され た個体(1960 年代:N=558 羽,2000 年代:N= 696 羽)を除いた新放鳥のデータ(1960 年代: N=16,821 羽,2000 年代:N=54,038 羽)を解析 に用いた. 日本に飛来するツバメは,主にフィリピン,イ ンドネシア,マレーシア,ベトナム南部で越冬し, 3 月中旬から本州で初飛来が確認されるようにな り,4 月中旬から 5 月上旬が春の渡り時期と言わ れている(清棲 1978;山階鳥類研究所 2002). 渡って来たツバメは,ヨシ原などの湿生草地に小 規模の春ねぐらを作り,4 月頃からは市街地や宅 地に集まって人家などの建築物で営巣を始める (環境庁 1994;須川 1999).そして,1∼2 回の繁 殖の後,6 月から大規模なねぐらを湿生草地に作 り,10 月中旬から 11 月中旬までに越冬地へと渡 去する.また,熊本,京都,静岡などでは,国内 で集団越冬する個体がいることも報告されている (清棲 1978).これらの情報に基づき,解析にあ たって,飛来期を4∼5 月,繁殖期を 6∼8 月,渡 去期を9∼10 月とした. 渡り時期については,成鳥(1960 年代:N= 2,049 羽,2000 年代:N=8,390 羽)と幼鳥(1960 年代:N=12,460 羽,2000年代:N=38,066 羽)の
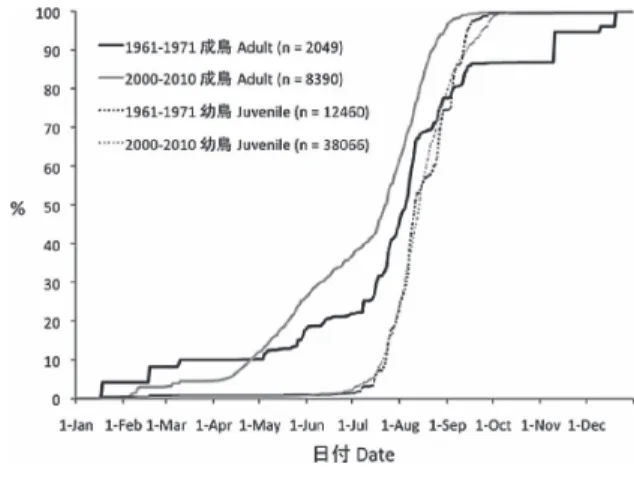
データを用いて作成した日別放鳥数の累積頻度分 布から,急増時期を飛来のピーク時期,急減時期 を渡去のピーク時期と判断し,1960 年代と 2000 年代で比較した.加えて,ツバメの調査場所が両 時期の間で地理的に変化した可能性を考慮し,飛 来期の4 月,5 月,渡去期の 9 月,10 月は月ごと に,6∼8 月は繁殖期としてまとめ,1960 年代と 2000年代で放鳥数の地域分布を県単位で比較した. ツバメの放鳥が無かった県については,調査が実 施されなかったために放鳥が無い場合と,調査が 実施されたにもかかわらず放鳥が無い場合を判別 するため,他の小型陸鳥の放鳥が無かった県とツ バメの放鳥だけが無かった県を区別して示した. ツバメの個体数密度が高い生息環境は季節に よって異なる可能性がある.そこでまず,2000 年 代の放鳥場所について記録されている調査環境, および2000 年代の位置情報と 1960∼70 年代に国 土地理院が発行した5 万分 1 地形図から推測可能 な1960 年代の放鳥場所の調査環境を,川沿いや湖 沼のヨシ原などの「湿生草地」,建築物や集落など の「市街地・宅地」,農耕地や林などの「その他」 に分け,判断不可能だった「不明」を加えた計4 カテゴリーについて,4∼10 月における日別・場 所別の調査回数に占める各カテゴリーの割合を 1960 年 代(4 月:N=11 回,5 月:N=47 回,6 月:N =97 回,7 月:N=87 回,8 月:N=95 回, 9 月:N=101 回,10 月:N=15 回)と 2000 年代 (4 月:N=216 回,5 月:N=421 回,6 月:N=463 回,7 月:N=534 回,8 月:N=533 回,9 月:N= 373 回,10 月:N=78 回)について月ごとに調べ た. 繁殖時期については,巣内雛(1960 年代:N= 1,175 羽,2000 年代:N=889 羽)と幼鳥の日別放 鳥数の累積頻度分布を1960 年代と 2000 年代で比 較した.幼鳥については,調査環境によって個体 数密度が異なる可能性があるため,上記4 カテゴ リーからねぐらとして利用する「湿生草地」の デ ー タ(1960 年 代:N=10,505 羽,2000 年 代: N=30,372 羽)のみを用いた.なお,営巣地付近 と判断される「市街地・宅地」のカテゴリーにつ いては,1960 年代の標識調査では幼鳥が 53 羽し か放鳥されていなかったため,この比較を行わな かった. 繁殖成績については,6∼8 月の繁殖期に捕獲放 鳥された幼鳥と成鳥の割合から判断した(Peach et al. 1996).調査環境によって個体数密度が異なる 可能性を考慮し,最も放鳥数が多かった「湿生草 地」のデータのみについて1960 年代と 2000 年代 で比較した.「市街地・宅地」のカテゴリーについ ては,1960 年代の標識調査では成鳥がわずか 8 羽 しか放鳥されていなかったため,この比較を行わ なかった. 渡り時期,繁殖時期についての放鳥日の比較で は, 等 分 散 の 条 件 が 満 た さ れ な か っ た た め, Wilcoxon Rank Sum test を用いた.11∼3 月の放鳥 日データは越冬個体が捕獲された可能性が高いた め(静岡県林務部 1962),渡り時期の解析には用 いなかった.繁殖成績の比較ではχ2-test を用いた. なお,標識調査では捕獲個体への負担軽減を重 視し,捕獲個体はその場で速やかに放鳥すること になっているので(山階鳥類研究所 2008),放鳥 日,放鳥場所は,捕獲日,捕獲場所とみなした. 結 果 1) ツバメの飛来および渡去時期の比較 日別放鳥数の累積頻度分布について,成鳥の放 鳥数が急増した時期は,1960 年代では 5 月初旬で あったが,2000 年代ではそれより半月ほど早い 4 月中旬だった(図1).一方,成鳥の放鳥数が急減 する時期は,両期間ともに9 月中旬に見られた. また,1960 年代では,越冬中と思われる個体が捕 獲放鳥された11 月中旬にも急増時期が見られた. 冬期(11 月∼3 月)を除いた成鳥の放鳥日は 1960 年代(中央値:8 月 2 日,N=1,575)に比べ 2000 年 代(7 月 23 日,N=8,001)が有意に早かった(z= 13.76,P<0.01).一方,幼鳥の日別放鳥数の累積 図1.1960 年代と 2000 年代におけるツバメの成鳥と幼 鳥の日別放鳥数の累積頻度分布.
Fig. 1. Cumulative frequency distributions of the number of adult and juvenile Barn Swallows released per day in the 1960s and 2000s.
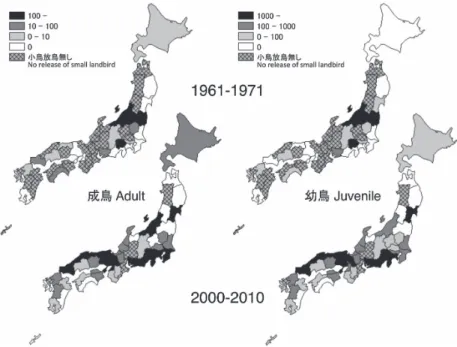
頻度分布は1960 年代と 2000 年代がほぼ一致し, 放鳥数はともに7 月中旬に急増し,1960 年代は 9 月中旬,2000 年代は 9 月下旬に急減した(図 1). 冬期を除いた幼鳥の放鳥日は,統計的には有意な 違いが認められたものの,その差は3 日と成鳥の 差(10 日)より小さかった(1960 年代:8 月 10 日, N=12,311,2000 年代:8 月13日,N=37,830,z= 2.27,P=0.02). 繁殖期の調査地は,1960 年代と 2000 年代の両 方でほぼ全国的に分布していたが,1960 年代には 西日本で調査が実施されなかった可能性の高い場 所が多く,逆に2000 年代では西日本に調査地が多 い傾向が見られた(図2).また,1960 年代では, 繁殖期に北海道で幼鳥が放鳥された記録は無かっ た. 飛来期の調査地に関しても,1960 年代には標識 が実施されていなかった可能性が高い場所が西日 本に多かった(図3).飛来期に成鳥が放鳥された 調査地は,1960 年代には山口でのわずかな例を除 いて4 月の記録は無かったが,5 月に入ってから は繁殖期に近い分布が見られた.一方,2000 年代 では,飛来期に成鳥が放鳥された調査地の分布は 4 月から繁殖期に近い分布が見られた. 渡去期の調査地も,1960 年代には調査の実施さ れなかった可能性が高い場所が西日本に多かった (図4).1960 年代と 2000 年代,ともに 9 月におけ る成鳥の放鳥地の分布は繁殖期と似た広がりであ り,10 月に入ると放鳥場所が大きく減少した. 各月の調査環境の割合を調べた結果,1960 年代 は「湿生草地」が8 月,9 月に高い割合を占めて おり,他の月は「その他」,「不明」の割合が高 かった(図5).いずれの月も「市街地・宅地」の 割合は低く,4 月の割合は 0 だった.一方,2000 年代は,4 月以降,それまで高かった「市街地・ 宅地」の割合が減少し,逆に低かった「湿生草地」 の割合が増加する傾向が見られた(図 5).また, 「その他」の割合は月間で大きな違いは見られず, 「不明」の割合はいずれの月も少なかった. 2) ツバメの繁殖時期の比較 巣内雛の日別放鳥数の累積頻度分布について, 1960 年代は 5 月下旬から放鳥が急増し,7 月下旬 に急減する傾向が見られた(図6).一方,2000 年 代は5 月上旬から放鳥が急増し,8 月初旬に急減 図2.1960 年代と 2000 年代における 6 ∼ 8 月のツバメの放鳥地の地域分布と日 別放鳥数.左上は1960 年代の成鳥,右上は 1960 年代の幼鳥,左下は 2000 年代 の成鳥,右下は2000 年代の幼鳥を示す.左の凡例は両期間の成鳥,右の凡例は 両期間の幼鳥に対応する.
Fig. 2. Areal distribution and abundance of Barn Swallows released per day from June to August in the 1960s and 2000s. Adults and juveniles released in the 1960s are shown at the upper left and right, respectively. Adults and juveniles released in the 2000s are shown at the lower left and right, respectively. The left abundance legends correspond to adults for the two periods and the right abundance legends correspond to juveniles for the two periods.
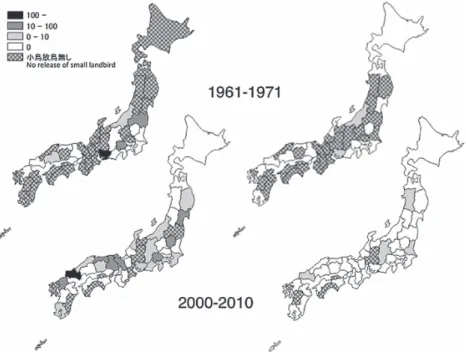
図3.1960 年代と 2000 年代における飛来期のツバメ成鳥の放鳥地の地域分布と放 鳥数.左上は1960 年代の 4 月,右上は 1960 年代の 5 月,左下は 2000 年代の 4 月, 右下は2000 年代の 5 月を示す.
Fig. 3. Areal distribution and abundance of adult Barn Swallows released per day in the 1960s and 2000s spring migration period. Birds released in April and May 1960s are shown at the upper left and right, respectively. Birds released in April and May 2000s are shown at the lower left and right, respectively.
図4.1960 年代と 2000 年代における渡去期のツバメ成鳥の放鳥地の地域分布と放
鳥数.左上は1960 年代の 9 月,右上は 1960 年代の 10 月,左下は 2000 年代の 9 月, 右下は2000 年代の 10 月を示す.
Fig. 4. Areal distribution and abundance of adult Barn Swallows released per day in the 1960s and 2000s autumn migration period. Birds released in September and October 1960s are shown at the upper left and right, respectively. Birds released in September and October 2000s are shown at the lower left and right, respectively.
する傾向が見られた(図9).巣内雛の放鳥日は, 1960 年代(中央値:6 月 15 日)に比べ 2000 年代 (6 月 8 日 ) が 有 意 に 早 か っ た(z=10.37,P< 0.01). 「湿生草地」における幼鳥の日別放鳥数の累積頻 度分布は,幼鳥の全放鳥数(図1)とほぼ同じ傾 向となり,両期間ともに放鳥は7 月中旬に急増し, 1960 年代は 9 月中旬に,2000 年代は 9 月下旬に急 減した(図8).「湿生草地」における幼鳥の冬期 を除いた放鳥日は,1960 年代(中央値:8 月 9 日) に比べ2000 年代(8 月 15 日)が有意に遅かった (z=12.74,P<0.01). 3) ツバメの繁殖成績の比較 6∼8 月の「湿生草地」において捕獲放鳥された 幼鳥と成鳥は,1960 年代では 8,389 羽と 1,029 羽, 2000 年代では 23,342 羽と 4,045 羽で,割合の差は わずかであったが(1960 年代:幼鳥 89%・成鳥 11%,2000 年代:幼鳥 85%・成鳥 15%),統計的 には有意な違いが認められた(χ2=87.12,P< 0.01). 考 察 気象庁の観測統計によれば,2000 年代の日本の 年平均気温は1960 年代に比べて約 0.8°C 上昇して おり,この変化の大部分は1980 年代末に短期間で 生じたことがわかる(気象庁 2011a).一般的に, 北太平洋全体の気候は,海面水温変動が関わる太 平洋数10 年振動の影響を強く受けているが(山川 2005),1980 年代末に見られた日本の急激な気温 変化については,北極圏の気圧が関わる北極振動 の位相反転が北太平洋西部の気象に強く影響した 結果として解釈されている(見延 2001).また, 日本における降水量の長期変化として,日本海側 地方では梅雨期(6∼7 月)の降水が相対的にこの 時期の後半に集中する傾向が近年見られている (Endo 2011).このような気候変化が見られる中, 1960 年代と 2000 年代の間では,ウメ Prunus mume やサクラPrunus spp. など多くの植物が開花や開葉 時 期 を1∼2 週 間 早 め( 樋 口 ら 2009; 気 象 庁 2011b), キ ア ゲ ハPapilio machaon, ホ タ ル 類 Luciola spp.,アブラゼミ Graptopsaltria nigrofuscata,
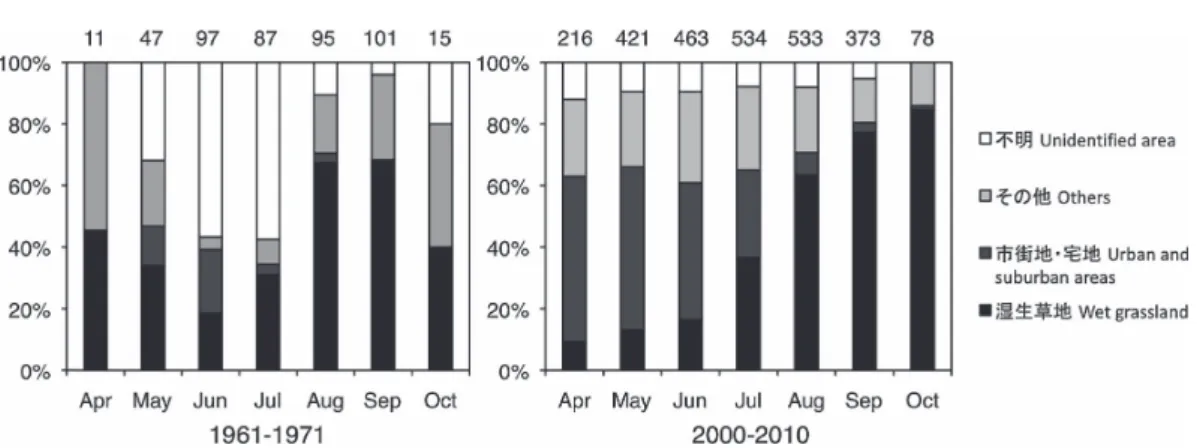
クマゼミCryptotympana facialis の初見や初鳴日も 1 ∼ 2 週間程度早まるなど(気象庁 2011b),日本 の陸域生物の生物季節には大きな変化が認められ る. 一般に,数100 ∼数 1,000 km の渡りを行う鳥類 は,到着地の気象変化を予測して渡りのタイミン グを選ぶわけではなく,到着地の気象と強く関わ 図5.1960 年代と 2000 年代におけるツバメの月別放鳥環境.上の数字は日別・場所別の調査回数を示す
Fig. 5. Monthly habitats where barn swallows were released in the 1960s and 2000s. The upper numbers indicate study numbers per day per site.
図6. 1960 年代と 2000 年代の国内におけるツバメの巣 内雛と湿生草地における幼鳥の日別放鳥数の累積頻度分 布.
Fig. 6. Cumulative frequency distribution of the number of Barn Swallow nestlings and the juveniles at wet grassland sites released per day in the 1960s and 2000s.
る大気の状態が広範囲に影響を及ぼすことで,出 発地での栄養状態,移動速度,中継地での滞在時 間などが変わり,その結果として,渡りの到着日 と到着地の気象変化との間に相関関係が現れると 言われている(Gordo 2007).本研究対象であるツ バメについて,気象庁の調査結果によれば,1960 年代と2000 年代の全ての年でデータが得られてい る東北から九州までの計29 地点において,その 7 割以上にあたる21 地点の初見日は 2000 年代の方 が早まっており(1960 年代の各地点の平均初見 日:3 月 21 日∼4 月 21 日,2000 年代の各地点の平 均初見日:3 月 11 日∼4 月 16 日),その差は平均 で6 日(レンジ:1∼12 日) であった(気象庁 2011b).また,アフリカ南部で越冬し,繁殖のた めに飛来するヨーロッパのツバメの初見日も, 1970 年代始めから急速な早期化が報告されている (Gordo & Sanz 2006; Sparks & Tryjanowski 2007).
これらの知見は,1960 年代のツバメ放鳥数の急増 時期が,2000 年代に比べて半月ほど遅かったこと を示す本研究結果と一致する(図1).日本,韓国, およびイギリスの6 都市では,ツバメの初見日と 気温の間に負の相関が認められており(Sparks et al. 2007; Primack et al. 2009),本研究の結果も近年 の温暖化の影響が関わっているのかもしれない. ただし,1960 年代の記録には,繁殖のために分 布が集中すると考えられる4∼5 月の市街地・宅地 の情報がほとんど含まれておらず(図5),本研究 結果を解釈する際にはこの点が問題となる.また, 1960 年代の記録には,調査を行ったが捕獲放鳥さ れなかったことを示すデータが存在しないため, 正確な判断はできないが,ツバメの放鳥記録があ る全調査回数に占める飛来期(4 月,5 月),繁殖 期(6∼8 月),渡去期(9 月,10 月)の調査回数 の割合は,1960 年代(飛来期:12.8%,繁殖期: 61.6%,渡去期:25.6%)に比べ 2000 年代(飛来 期:24.3%,繁殖期:58.5%,渡去期:17.3%)の 方がより早い時期に集中していた(図5).そのた め,両期間の急増時期の違いには調査時期の影響 も関わっているのかもしれない. なお,1960 年代には,ツバメの飛来期にあたる 4∼5 月に,調査が実施されなかった可能性の高い 地域が西日本に多く見られ(図3),データの有無 の地理的な偏りが飛来時期の解釈に影響を及ぼす 可能性が考えられた.そこで,1960 年代と 2000 年代の両年代で地理的な偏りなくデータが存在す る気象庁の初見日調査情報(気象庁 2011b)を用 いて,標識調査でデータが得られた1960 年代の地 域(9 地点・160 回)と 2000 年代の地域(25 地 点・1,814 回)における平均日を比較した.その結 果,2000 年代の方が 2 日早いだけだったため,両 期間で調査地域が異なることによる影響は小さい と判断された. 一方,ツバメの渡去日の長期的変化を調べた結 果は,日本を含むアジアではこれまで報告されてい ない.ヨーロッパの場合,スペイン国内で60 年分 のデータを集めた報告では,1980 年代中旬から緩 やかな早期化が見られたが(Gordo & Sanz 2006), イギリスで30 年以上継続的に調べられた報告では 早晩の傾向は認められていない(Sparks et al. 2007). 一般的に,鳥類の繁殖終了後の渡り時期には,繁 殖開始前のものと違って,一定傾向の長期的変化 が認められないことが知られており,その理由と して,渡り距離の長さや年間の繁殖回数の違いな どが関わると考えられている(Gordo 2007).本研 究で解析した標識調査記録では,1960 年代と 2000 年代の成鳥および幼鳥の放鳥数の急減時期は9 月 中∼下旬であった(図 1).本研究の結果だけでは 判断できないが,日本におけるツバメの秋の渡り 時期は,1960 年代と 2000 年代の間で大きく変わっ ていないのかもしれない. ツバメの繁殖時期の長期的変化を調べた結果は, 日本を含むアジアではこれまで報告されていない. イタリアにおいて,ツバメと越冬地のアフリカ南 部から繁殖のために渡ってくるヨーロッパアマツ バメApus apus の初見日と初卵日を 1980 年代から 25 年間継続的に調べた報告では,2 種ともに初見 日と初卵日の両方で早期化が認められている (Rubolini et al. 2007).加えて,オランダにおいて 同じく長距離を渡るヨーロッパマダラヒタキ Fice-dula hypoleuca について,最初の 5∼10 羽が標識放 鳥された日と初卵日を1960 年代から 30 年以上調 べた報告でも,その両方において早期化が認めら れている(Both et al. 2005).また,日本に飛来す るツバメと同じくフィリピン,ボルネオ島北部か ら繁殖のために渡るコムクドリSturnus philippensis の初卵日について,新潟で1970 年代から 30 年近 く調べた報告でも早期化が認められている(Koike et al. 2006).これらの知見は,1960 年代の巣内雛 の放鳥時期が,2000 年代に比べて 1 週間ほど早 かったことを示す本研究結果と一致する(図6). 一方,「湿生草地」における幼鳥の放鳥時期では, 2000 年代の方が遅い傾向が見られた(図 6).ツバ メの育雛期間には大きな幅が見られないことから (19∼21 日,Bryant & Hails 1983),幼鳥が「湿生
草地」にねぐら入りする時期には,例えば照度な ど(Kiis 1986),気候変動とは別の決定要因がある のかもしれない.また,デンマークでは,1970 年 代以降,ツバメの各年における第1 回目と第 2 回 目の産卵時期の間隔が年々広がっている結果が報 告されている(Møller 2007).本研究の結果でも, 巣内雛放鳥数の頻度分布の両裾は,1960 年代に比 べ2000 年代の方が広がっており(図 6),両期間 の幼鳥がねぐら入りする時期には,産卵時期の間 隔の変化が関わっているのかもしれない. 気候変動が鳥類の繁殖成績に影響を及ぼす主な メカニズムとして,繁殖時期と餌資源の出現時期 の一致の程度があげられる(Visser et al. 1998; Both & Visser 2001).また,気候変動は,鳥類の年内の 繁殖可能期間にも影響を及ぼすため,年に複数回 繁殖できる種では,2 回目以降の繁殖を行う頻度 と関わる(Visser et al. 2003; Gordo 2007).例えば, 地中海地域に渡ってくるヨーロッパマダラヒタキ では,ここ20 年の間に餌の出現時期が早まったの に対し,繁殖開始時期には変化が見られないため, 繁殖成績が悪化している(Sanz et al. 2003).しか し,デンマークに渡ってくるツバメでは,第1 回 目と第2 回目の産卵数には気候変動の影響が認め られるが,孵化および育雛成功率にはこのような 影響は認められず,顕著な年変化も見られていな い(Møller 2002).日本を含むアジアでは,ツバメ の繁殖成績の長期的変化に関する情報がほとんど 報告されていないため,1960 年代の全成鳥数/全 幼鳥数の比率(12.3%)に比べて 2000 年代の比率 (17.3%)がわずかに上がっている本研究の結果の 妥当性を判断することはできないが,1960 年代と 2000 年代では,日本に渡ってくるツバメの繁殖成 績はやや減少しているのかもしれない.ただし, 他の結果についても言える事だが,本研究で扱っ たサンプルサイズは非常に大きいため,比較結果 は統計学的に有意な違いとして認められやすい傾 向にある.そのため,繁殖成績などに見られたわ ずかな違いに関しては,必ずしも生態的な変化を 表していない可能性がある. また,本研究では,幼鳥と成鳥の比率を繁殖成 績の指標としたが,全ての成鳥が繁殖したという 仮定における1 番い当たりの年間繁殖成績[全幼 鳥放鳥数/(全成鳥放鳥数×0.5)]は 1960 年代で は16.2 羽,2000 年代では 11.3 羽となり,これら の値は実際にツバメ1 番いが巣立たせる年間の雛 数(5∼10 羽,Møller 2007)より多い.一般に, かすみ網で鳥類を捕獲した場合,幼鳥の捕獲数は 捕獲努力量に関わらず,環境中の生息密度の良い 指標となるが,成鳥の捕獲数は捕獲努力量を増し た場合,実際の生息密度を過小評価しやすくなる という結果が出ている(Burton & DeSante 2004; de Feu & McMeeking 2004).そのため,本研究の結 果には捕獲努力量,例えば1960 年代の方が少ない 場所でより高頻度に調査されたり,多くの枚数の かすみ網が用いられた可能性などの影響も表れて いる可能性がある. 本研究では,標識調査情報の活用例として,気 候変動との関係性に注目し,1960 年代と 2000 年 代のツバメの渡りと繁殖状況を比較した.1960 年 代の標識調査は,再捕獲による標識個体の移動分 散の解明を主な目的としており,1971 年以降の データとは一部違う目的で実施されていた.その ため,渡り時期や繁殖状況を解析するためには, 情報に不十分な点が見られた.しかしながら,飛 来時期と繁殖開始時期については,多くの既有の 知見と一致することから,結果にはある程度の妥 当性があると判断された.今後,他の渡り鳥につ いても同様の解析を進めて行く価値があると考え られる.一方,渡去時期や繁殖成績については, 国土スケールの長期変動解析に有効な情報が他に 乏しく,標識調査情報が重要なデータになると言 える.特に,国内の飛来数が近年増加したアオジ Emberiza spodocephala や減少したカシラダカ E. rustica(米田・上木 2002)は,1960 年代の標識調 査においても比較的多くのデータが広範囲に得ら れており,今後解析を進めると良い材料と言える. なお,渡去時期については目視観察など,繁殖成 績については巣ごとの繁殖状況(産卵数や巣立ち 雛数)の情報収集を補足的に行えば,結果の解釈 に対する信頼性がより高まると考えられる.また, 本研究テーマとは別に,ツバメを材料として, 1961 年から 2000 年代にいたる連続的な経年変化 パターンを記述することや,長期変動の地域差, およびそれらと気象条件の関係などを調べること も興味深いテーマである. 本研究は,非常に多くの鳥類標識協力調査員による継 続的な調査努力と情報提供無くしてはありえなかった. また,山階鳥類研究所保全研究室の室員には,調査情報 のデータベース登録および本原稿の内容確認にご協力い ただき,佐藤文男氏,富田直樹氏,2 人の査読者には多 くの有益なご意見を頂戴した.これらの方々に深く感謝 申し上げる.なお,本研究は,文部科学省の科学研究費 補助金特定奨励費事業「山階鳥類研究所データベースシ ステムの構築と公開」の一環として実施した.
摘 要 地球規模の気候変動が鳥類の渡り・繁殖に及ぼ す影響の有無は,ヨーロッパや北米に生息する種 について数多く報告されているが,アジアにおけ る報告は著しく乏しい.本研究では,大規模な気 候変動が生じたこの50 年の間に,日本に訪れるツ バメの渡り時期,繁殖時期,繁殖成績が変化した かどうかを検討するため,1961∼1971 年の 11 年 間(以下,1960 年代)と,2000∼2010 年の 11 年 間(以下,2000 年代)の標識調査データを比較解 析した.日別放鳥数の頻度分布において,成鳥の 急増時期は2000 年代の方が半月ほど早まったが, 急減時期は変わっていなかった.幼鳥については, 2000 年代と 1960 年代の頻度分布はほぼ一致した. 巣内雛については,急増時期が2000 年代の方が半 月ほど早まっていた.成鳥に対する幼鳥の放鳥数 の比率は2000 年代の方がやや減少していた.これ らの結果は,ツバメの飛来時期・繁殖開始時期は 2000 年代の方が早まったが,渡去時期は両期間で 変わっておらず,また繁殖成績は2000 年代の方が 低下したことを示唆している.飛来時期と繁殖開 始時期については,調査環境や調査時期の影響も 考えられるが,多くの既有の知見と一致すること から,比較結果にはある程度の妥当性が判断され た.一方,渡去時期や繁殖成績については,国土 スケールの長期変動解析に有効な情報が他に乏し く,標識調査情報が重要なデータになると言える. 引 用 文 献
Both C & Visser ME (2001) Adjustment to climate change is constrained by arrival date in a long-distance migrant bird. Nature 411: 296–298.
Both C, Bijlsma RG & Visser ME (2005) Climate effects on timing of spring migration and breeding in a long-distance migrant, the pied flycatcher Ficedula hypoleuca. J Avian Biol 36: 368–373.
Bryant DM & Hails CJ (1983) Energetics and growth patterns of three tropical bird species. Auk 100: 425–439.
Bulter CJ (2003) The disproportionable effect of global warm-ing on the arrival dates of short-distance migratory birds in North America. Ibis 145: 484–495.
Burton KM & DeSante DF (2004) Effects of mist-netting frequency on capture rates at monitoring avian productivity and survivorship (MAPS) stations. Stud Avian Biol 29: 7–11.
Cotton PA (2003) Avian migration phenology and global climate change. Proc Natl Acad Sci USA 100: 12219– 12222.
Crick HQP (2004) The impact of climate change on birds. Ibis 146: 48–56.
de Feu CR & McMeeking JM (2004) Relationship of juveniles captured in constant-effort netting and local abundance. Stud Avian Biol 29: 57–62.
Endo H (2011) Long-term changes progress in Baiu rainfall using 109 years (1901-2009) daily station data. Sora 9: 5–8. Hansen J & Lebedeff S (1987) Global trends of measured
surface air temperature. J Geophys Res 92: 13345–13372. Gordo O (2007) Why are bird migration dates shifting? A
review of weather and climate effects on avian migratory phenology. Clim Res 35: 37–58.
Gordo O & Sanz J (2006) Climate change and bird phenology: a long-term study in the Iberian Peninsula. Global Change Biol 12: 1993–2004.
樋口広芳・小池重人・繁田真由美 (2009) 温暖化が生物 季節,分布,個体数に与える影響.地球環境 14: 189– 198.
Hüppop O & Hüppop K (2003) North Atlantic Oscillation and timing of spring migration in birds. Proc Roy Soc Lond B 270: 233–240.
環境庁 (1994) 第 4 回基礎調査動植物分布調査報告書 鳥
類の集団繁殖地及び集団ねぐら.(オンライン) http://
www.biodic.go.jp/reports/ 4-03/e113.html,参照 2012-06-05.
Kiis A (1986) The timing of roosting in Greenfinches Cardu-elis chloris. Ornis Scand 17: 80–83.
気象庁 (2011a) 日本の年平均気温.(オンライン)http:// www.data.kishou.go.jp/climate/cpdinfo/temp/an_jpn.html, 参照2012-06-05. 気象庁 (2011b) 生物気象観測値 収録期間:1953∼2008.12. (財)気象業務支援センター,東京. 清棲幸保 (1978) 日本鳥類大図鑑 I(増補改訂版).講談 社,東京.
Koike S, Fugita G, Higuchi H (2006) Climate change and the phenology of sympatric birds, insects, and plants in Japan. Global Environ Res 10: 167–174.
米田重玄・上木泰男 (2002) 環境庁織田山一級ステー
ションにおける標識調査─1973 年から 1996 年におけ
る定量的モニタリング結果─.山階鳥研報 34: 96–111. Marra PP, Francis CM, Mulvihill RS & Morre FR (2005) The
influence on the timing and rate of spring bird migration. Oecologia 142: 307–315.
見延庄士郎 (2001) 同期した 50 年変動と 20 年変動がも たらす気候レジームシフト.号外海洋 24: 42–49. Møller AP (2002) North Atlantic Oscillation (NAO) effects of
climate on the relative importance of first and second clutches in a migratory passerine bird. J Anim Ecol 71: 201–210.
Møller AP (2007) Interval between clutches, fitness and climate changes. Behav Ecol 18: 62–70.
日本鳥学会 (2000) 日本産鳥類目録.日本鳥学会,東京. Peach WJ, Buckland ST & Baillie SR (1996) The use of
constant effort mist-netting to measure between-year changes in the abundance and productivity of common passerines. Bird Study 43: 142–156.
Primack RB, Ibáñez I, Higuchi H, Lee SD, Miller-Rushing AJ, Wilson AM & Silander Jr JA (2009) Spatial and inter-specific variability in phonological responses to warming
temperature. Biol Conserv 142: 2569–2577.
Rubolini D, Ambrosini R, Caffi M, Brichetti P, Armiraglio S & Saino N (2007) Long-term trends in first arrival and first egg laying dates of some migrant and resident bird species in northern Italy. Int J Biometeorol 51: 553–563.
Sanz JJ, Potti J, Moreno J, Merino S & Frias O (2003) Cli-mate change and fitness components of a migratory bird breeding in the Mediterranean region. Global Change Biol 9: 461–472.
静岡県林務部 (1962) 浜名湖畔の越冬ツバメ.静岡県林 務部,静岡.
Sparks T & Trjanowski P (2007) Patterns of spring arrival dates differ in two hirundines. Clim Res 35: 159–164. Sparks TH, Huber K, Bland RL, Bland RL, Crick HQP,
Croxton PJ, Flood J, Loxton RG, Mason CF, Newnham JA & Tryjanowski P (2007) How consistent are trends in arrival (and departure) dates of migrant birds in the UK? J Ornithol 148: 503–511.
Stenseth NC, Mysterud A, Ottersen G, Hurrell JW, Chan K-S & Lima M (2002) Ecological effects of climate fluctuations. Science 297: 1292–1296.
須川 恒 (1999) ツバメの集団塒地となるヨシ原の重要 性.関西自然保護機構会報 21: 187–200.
Visser ME, Adriaensen F, van Balen JH, Blondel J, Dhondt AA, van Dongen S, du Feu C, Ivankina EV, Kerimov AB, de Laet J, Matthysen E, McCleery R, Orell M & Thomson DL (2003) Variable response to large-scale climate change in European Parus populations. Proc Roy Soc Lond B 270: 367–372.
Visser ME, Noordwijk AJV, Tinbergen JM & Lessells CM (1998) Warmer springs lead to mistimed reproductionin great tits (Parus major). Proc Roy Soc Lond B 265: 1867–1870. Walther G–R, Post E, Convey P, Menzel A, Parmesan C,
Beebee TJC, Fromentin J-M, Hoegh-Goldberg O & Bairlein F (2002) Ecological responses to recent climate change. Nature 416: 389–395. 山川修治 (2005) 季節∼数十年スケールからみた気候シ ステム変動.地学雑誌 114: 460–484. 山階鳥類研究所 (2002) 鳥類アトラス.山階鳥類研究所, 我孫子. 山階鳥類研究所 (2008) 鳥類標識マニュアル(改訂第 11 版).山階鳥類研究所,我孫子. 山階鳥類研究所標識研究室 (1985) 日本の鳥類標識調査 (昭和36 年∼昭和 58 年).山階鳥類研究所,我孫子. 山階芳麿 (1980) 日本の鳥類と其の生態(復刻版).出版 科学総合研究所,東京.