培養赤潮ブランクトンの自己消滅
佐藤優行,吉野東吾
AUTOLYSISOFREDTIDEPLANKTONS
MasayukiSATO andT6gOYosHINO
Ce11s of red tide planktons,Chattonelhlantique,Heterosigma akashiwo and hoYOCentrum mihi−mum, disappearedreadilyundersuchcultureconditionsasanaerobic,dark,higher(pH9)orlower’(pH5,7)pHand highertemperature(250,300,350C).Intrace11ularacidphosphataseactivity ofPminimumincreased tempo−
rarilyduringthesecellextinctionphases
赤潮ブランクトンであるC血z抽乃βJ由α乃軸祝β,放ね和痩硯川紘崩御およびPれ晰鋸署吼加雛形チ拙乃よ桝〝椚を用い,細 胞が自己消滅を起こす条件を検討した。各細胞を増殖させた後,培養の通気,光,♪ガ,温度の各条件を変えることに ょって速やかに細胞が消滅した。Pminimumの場合について,自己消滅過程における残存細胞内のaCid phos− phatase活性の変動を追跡した。どの条件変化による消滅過程においても,d時活性が増大する現象が認められた。 緒 地球という有限の生態系で生物が生き続けていくためには,生命活動のために環境から侵食した空間と栄養物質を, あとで必ず環境へ再利用できる型で返還し,それによって生態系の原状回復がはかられねばならない。その観点に立 てば,従来細胞レベルで知られているフトートリシスという現象も,単に無秩序化へ向かう脱制御の過程と把えるので はなく,生態系の原状回復という目的を達成するために,あらかじめプログラムされた合目的的な制御過程と見るこ とができるのではなかろうか。そのような考えのもとに,大橋らは「プログラムされた自己解体モデル」という仮説 を提唱した(1)。我々はこの仮説にもとづき,各種の微生物を対象に自己解体現象に関する研究を行ってきた(2 ̄4)。 その一層として赤潮ブランクトンの細胞を研究材料として取り上げた。これらのブランクトンは異常発生して赤潮 となった後,急速に消滅してしまうことがある。その原因としては,風や潮流など物理的なことや,動物ブランクト ンの餌になったり細菌によって分解されたりすることが知られているが,その他に細胞自身が起こす自己消滅があ る(5)。この自己消滅も,細胞の“プログラムされた自己解体”の結果と見ることができるのではなかろうか。そのよう な観点からすると,自己消滅が進みつつある培養中の残存細胞内でほ自己解体が起こっているのではないかと考えら れる。 そこでまず,自己消波が起こる培養条件,しかも細胞内で起こる生化学的変化(解体酵素活性など)を追跡できる ような比較的穏やかな条件を検討することとした。今回は,3種のプランタトソを用い,それぞれ単独に増殖させた 後,培養条件(通気,光,pH,温度)を変えてみた。そしてどんな条件の場合に細胞数が減少していくかを観察した。 ついで,自己解体の荷負い手として予想しているリソゾーム酵素(その代表としてacidphosphatase)活性が,その 自己消滅過程でどう変化するかもしらべた。 実験材料および方法 1.供試ブランクトン Cゐαfわ乃β地α乃軸〟e,月壱′g和一Sな桝ααぬゐ室紺0および伽和C㈹加齢㍑刑薇劫㈹肌を本実験で使用した。これらは本学 部海水利用学研究室より分譲を受けた。いずれも赤潮の原因となる,いわゆる赤潮ブランクトンである。76 香川大学農学部学術報告 第39巻 第1号(1987) 2 培養法 培地にはつぎのような組成のErd−Schr・eiber型改変培地(6)を用いた。すなわち,NaNO,120mg,K2HPO45mg,
EDTA−FeO26mg,EDTA−MnO.33mg,ビタミソBl−HClO.1mg,ビタミソB121pg,ビオチソ1JLg,トリスアミノメ
タン05gおよび土壌抽出液50mlを濾過海水に溶かして1月とした。pHは78−80に調整した。この培地を試験管あ るいは三角フラスコに入れ,これにシリコ栓をほどこした後,加圧滅菌した。培養は通常200C,14時間明,10時間陪 の静置で行った。毎日1回以上培養液をゆるく撹絆した。 3培養条件の変換 培養の途中でつぎのようにして各条件を変えた。i)通気培養液に無菌濾過した窒素ガスを十分に吹き込み,酸 素を追い出した後,滅菌ゴム栓で培養器を密栓した。ii)光二培養器の表面をアルミホイ′しで完全に包み光を遮断し た。iii)pH滅菌した0.1NHClあるいほ01NNaOHを用い培養液のpHを5,7,9に変えた。iv)温度250,300, 350Cの各恒温槽へ移した。 4.細胞破砕および粗酵素液の調製\ 細胞を遠心分離(2,000×g,10分間)により集め,これを005MトリスーHCl緩衝液(pH75)で−・回洗浄した後,
適量の同緩衝液にけんだくした。このけんだく液を氷冷しつつ,超音波処理(1分間,5回)を行い,細胞を破砕し た。この破砕液を遠心分離(8,000×g,10分間)し,得られた上清液を粗酵素液とした。 5分 析 i)細胞数の測定:培養を前記トリスーHCl緩衝液で適当に希釈し,それを試料として025mlとり,経線入りスラ イドグラス上に載せ,原敬鏡下で計数することにより求めた。ii)タンパク貿の定量年血清アルブミンを標準物質 として,Lowryらの方法(7)により求めた。iii)acidphosphatase活性てP−nitrophenylphosphateを基質に用いて前 報(2)に記した方法で測定した。 実 験 結 果 1増殖曲線 Fig1に3種のブランクトンの典型的な増殖曲線を示した。培養開始後10∼14日で定常期に達し,20∼25日から死滅 期に入り,30日あるいはそれ以上でほとんど消滅した。定常期における細胞数は,C胡萌閲で3∼4万個/ml,ガ α鮎5ゐ去紺0で30−40万個/ml,P彿よ乃左研〝椚で10−15万個/mlであった。 S重層蔓喪栗山﹁ざ眉嘩竃霊∴取ら ︵竜\s01×︶S〓むU 箋昂雲昌∴︶睾︵琶\もlX︶S〓むU 1 2 3 4 20 Culturetime(days) 10Figl GrowthCurveSOfPlanktons
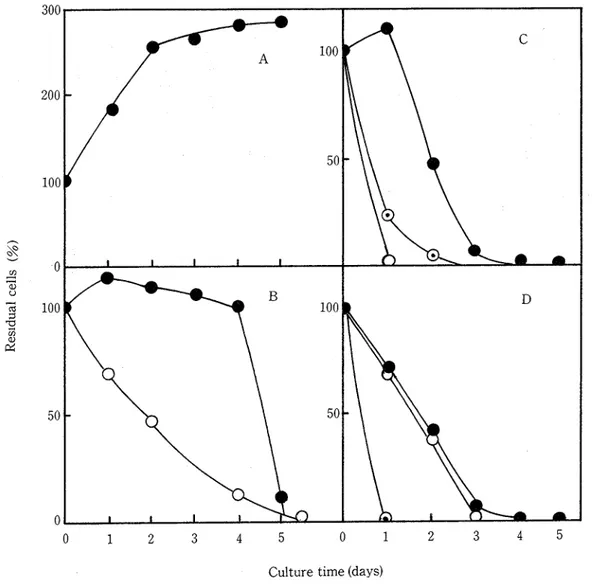
●:Cα光軸〟g0:〃α勉sゐ∠ぴ0◎:P椚去扁刑祝桝 2自己消滅条件の検討 自己解体は細胞が死んだ後に進むと考えている。したがって死滅期の細胞群が研究対象として適している。しかし, 供試のブランクトンの場合,通常の培養では死滅期に達するのに長時間を要する(Fig1)。しかもいつも同じ時間に達 するとは限らない。そこで,ある程度増殖させた段階で,人為的に培養条件を変えることにより,確実にしかも短時間に死滅期を迎えさせようと考えた。ここでほ,通気,光,pH,温度の各条件を変えて細胞数が減少するかどうかを 調べた。この自己消滅する時期を死滅期とした。本実験での培養ほ,10mlの培地を入れた試験管中で行った。 i)CゐαJわ邦eJおα乃軸〝βの場合 Fig2のAはコントロール,すなわち培養条件を変えずにその■まま続けた時のものであり,本実験では細胞数が増加 した。まだ定常期に達していなかった。B,C,Dはそれぞれ通気と光,pH,温度の各条件を変えた場合を示す。い ずれの場合もコソトローリレとほ対照的に細胞数は減少した。とりわけ温度とpHの影響が著しかった。pH5あるいは 350Cに変えた時には1日後すでに大きく減少し,細胞ほほとんど見られなくなった。 ︵訳︶sコむU−ヨpISU出 0 1 2 3 4 5 0 1 2 3 4 5 Culturetime(days) Fig2”ResidualCellsofC antiqueundertheVar’iousCultureConditions A:COntrOlB:anaerObic(○)anddark(●)C:pH5(○),7(●)and9(◎)D:25’(●),300(○)and 350(◎) ii)ガβね和Sな肌αα勉sゐ∠紺0の場合 Fig3に結果を示した。他の2種とちがう点ほ,嫌気状態にした時に細胞の消滅が著しいことであり,1日後で細胞 数ほ急激に減少した。またpHの変化にも弱く,どのpH条件でも1日目で大きく減少した。350Cでは1日目で全細 胞が消滅した。300,250Cでほ2日目までコントロールとほぼ同じだったが,その後減少した。
78 香川大学農学部学術報告 第39巻 第1号(1987)
︵訳︶s〓むUldコp倭還
0 1 2 3 4 5 0 1 2 3 4 5
Culturetime(days)
Fig3ResidualCellsofH akashiu)O under theVariousCultureConditions SymboIsarethesameasFig.2 iii)Pγ0γPCβ乃わⅦ肌用∠乃∠刑〟偶の場合 Fig4に示したように,他の2種と比べると各条件とも最終的な減少率がやや小さかった。嫌気的条件にした場合1 日目で細胞ほほは半減したが,5日後でも約10%残存していた。pH7の場合はコントロールと同じく細胞は減らな かった。しかしpH5,9でほ1日目で約30%しか残っていなかった。 3ProYOCentYu玩minimum(D消滅期におけるacidphosphatase惰性 P∽i扁研祝∽の場合について,前項の実験結果を参考にして人為的に消滅期を起こさせ,その過巷における残存細 胞のacid phosphatase活性を追跡してみた。この実験での培養ほ,300mlの培地を入れた500ml容三角フラスコ中で 行った。なお,C〈Z乃軸〝βおよび〃αゐαSゐま抑0の場合は,細胞がこわれやすく,遠心分離による細胞の回収が困難であっ たので,本実験には用いなかった。 接種後通常培養を10日間行い,その後各条件を変えた。Fig5は,条件を変えずにそのまま続けたコントロール(A), 光を遮断し暗くした場合(B),窒素置換により嫌気的にした場合(C),およびpHを9に上げた場合(D),それぞ れにおけるacidphosphatase活性を示す。いずれも細胞106個当たりの活性および比活性として表した。また同時に 細胞数の変化も示した。
︵訳︶sコむU↓再コp講じ出 0 1 2 3 4 5 0 1 2 3 4 5 Culturetime(days)
Fig4ResidualCellsofPminimumundertheVariousCultureConditions
SymboIsarethesameasFig2 コントロールでほ細胞数がほとんど変化せず,培養ほ定常期に達していることがわかる。この時acidphosphatase 活性もほとんど変化なかった。それに対して条件を変えた場合いずれも細胞数が減少した。しかし前項の実験の場合 に比べ減少が極端ではなく,細胞の回収には.好都合であった。aCidphosphatase活性の方ほいずれの場合も細胞の減 少する途中で,一旦増大することが認められた。 考 察 供試のブランクトンは酵母やかびなどに比べ細胞表層がぜい弱である。とくにCの紙料鋸と〟α勉sゐよ紺0は細胞壁 がなく破裂しやすい(8)。したがって本実験での細胞の自己消滅ほ,必ずしも我々の期待するような自己解体の結果とは 限らないだろう。単純な物理的破裂によるものも多いだろうと考えられる。我々の研究目的からすると,死んで自己 解体が進みつつあるがまだ破裂にまでは至っていない状態の細胞を得るのが望ましい。その点から見ると,全細胞数 の減少だけをしらべるのではなく,細胞の生死を判別して,死細胞の増加をもメルクマールとするべきである。そこ でメテレソブルーなどを用いて染色による識別法を検討した。しかし今回は成功しなかった。 P川扇扇■研〟雛の細胞は表面に殻板を持っており(8),他の2種と比べて丈夫であるため細胞を集めて酵素活性を測る80 香川大学農学部学術報告 第39巻 第1号(1987) 二て二こ こ≡=三二こ:−・−′. ︵ぎ\S苫コN.〇TX︶音A一も再じ叫J竃ds ︵竜\s01×︶hぷ已nuコむU O 2 1 0 5 0 24 48 72 0 Cultur・etime(hr)
Fig」5 AcidPhosphataseActivityinP minimum CellsundertheVariousCultureConditions A:COntrOlB:darkC:anaerObicD:pH9 ●:Ce11number0:aCtivity perlO6cells
◎:SpeCiacactivity のには適していた。ただし酵母などと比べ培養液当たりの細胞畳が少ない点で不利である。本実験では得られた細胞 抽出液が少量であったため,aCidphosphatase活性だけを測定した。この酵素ほ真核細胞におけるリソゾームのメル クマー・ル酵素とされている(9)。大橋らの仮説では,リソゾームが自己解体ロボットに相当するのでほないかと考えられ ており,我々もその仮説に立脚してこの酵素に注目したわけである。 Fig5に示すように,−・部の細胞が消滅していきつつある条件で,残存した細胞内ではこの酵素活性が上昇する事実
が認められた。我々ほすでにA*eYgillus nなerおよびSacchaYO鱒γCeS CeYeVisideKおいても同様に,死滅期にacid phosphatase括性やproteinase活性が増大する現象を観察してきた(23)。このような現象がどういうメカニズムで起 こるかほ今後の研究に得たねばならない。一L方,すでに大橋らはテトラヒメナを・用いて,細胞の死滅後,aCid phos− phataseが新たにde novo合成されることを示唆するデータを得ている(110)。SayareらもStr@bcoccus.カeαlisの オートリシス時に蛋白合成や核酸合成が活発化する事実を認めている(11)。このようなデータからも我々は,細胞内に あらかじめプログラムされたある種のメカニズムによってacid phosphataseを含む多くの解体酵素活性が増強され るのではないか,という考えを強くしている。本実験で得られた現象も,この仮説を実証していく上の−つの研究材 料となりうるであろう。 一方,赤潮に関する研究ほ活発に行われており,その発生磯構も生化学的生態学的研究により徐々に解明されつつ ある=2〉 。しかし赤潮の消滅機構についてほ研究が遅れているそうである。もしその解明が進めば,赤潮対策の叫助に
なるかもしれない。今回の実験は,海とはかけ離れた実験室レベルの条件で行ったものであり,赤潮との関係ほきわ めて薄い。しかし赤潮により近い条件の下でブランクトン細胞の自己解体メカニズムを追求することも,赤潮の消滅 機構解明の−・方向ではないかと考えられる。 謝 辞 プランクトンを分譲していただき,培養の仕方や赤潮ブランクトンに関する知見をご指導ご教示いただいた岡市友 利教授に感謝いたします。貴重な討論と助言をいただいた岩原章二郎教授および筑波大学の大橋 力博士に感謝いた します。 引 用 文 献 (1)大橋 力,中田大介,菊田 隆,村上和雄 科学 (1951) 基礎論研究,18,79(1987). (8)福代康夫 赤潮の科学,岡市友利編,恒星社眉生 (2)SATO,Mand M.SHIGEMURA:7セchhBullhFbc 閣,pp52−53,42−43,(1987). Agr彪酢Wathliv,37,7(1985) (9)CooNEEandPKSTUMPF:OutlinesofBio・ (3)SATO,Mand YOHTA‥7bckBullFbc Agr Chemistry,Fourth Ed,・John Wiley&Sons,
Inc”,NewYork,pp267−269(1976) (10)大橋 九 中田大介,菊田 隆 生化学,55,667 (1983). (11)SAYARE,M,LDANEO−MooREandGDSHOCK MAN:ノ助(fβわoJ,112,337(1972) (12)岡市友利編 赤潮の科学,恒星社厚生圃,pp91 −285(1987). (1987年5月25日受理) 物抑αこ玩去れ37,11(1985)
(4)SArO,M,Y OxADA and K TAOXA:Tbck
β“JJ鞄cAgγ物紺αこ侮れ37,17(1985) (5)柳田友道赤潮,講談社,p128,(1976). (6)岩崎英雄:藻類研究法,西沢一俊,千原光雄編, 共立出版札 pp.283−284,(1979). (7)LowRY,0H,NJRosEBROUGH,A.L FARR and RJ”RANDALL:JBioIChemリ193.265