博 士 学 位 論 文
反復寒冷ストレス負荷マウスにおける
体温調節能と炎症反応性の変化に関する研究
近 畿 大 学 大 学 院
薬 学 研 究 科 薬 科 学 専 攻
宮
本
朋
佳
反 復 寒 冷 ス ト レ ス 負 荷 マ ウ ス に お け る 体 温 調 節 能 と 炎 症 反 応 性 の 変 化 に 関 す る 研 究 宮 本 朋 佳博 士 学 位 論 文
反復寒冷ストレス負荷マウスにおける
体温調節能と炎症反応性の変化に関する研究
平 成 2 9 年 3 月近 畿 大 学 大 学 院
薬 学 研 究 科 薬 科 学 専 攻
宮
本
朋
佳
1
目次
緒論 ... 1 第 1 章 ストレス誘発体温上昇 に対する反復寒冷ストレスの影響 ... 3 緒言 ... 3 I. 実験方法 ... 4 II. 実験結果 ... 7 III. 考察 ... 12 第 2 章 炎症性刺激に対する反応性 に及ぼす反復寒冷ストレスの影響 ... 15 緒言 ... 15 第 1 節 Lipopolysaccharide 誘起発熱反応 に対する反復寒冷ストレスの影響 ... 16 I. 実験方法 ... 16 II. 実験結果 ... 19 III. 考察 ... 25 第 2 節 シクロホスファミド誘起膀胱炎・膀胱痛への 反復寒冷ストレスの影響 . 27 I. 実験方法 ... 27 II. 実験結果 ... 29 III. 考察 ... 32 総括 ... 34 引用文献 ... 37 略語一覧 ... 45 主論文 ... 46 謝辞1
緒論
現代社会は、ストレス社会と言われ、多くの人が様々なストレスに曝されている。 近年、仕事によるストレスから生じる精神障害による労災認定は増加しており、労働 者に対するメンタルヘルス対策が問題となっている。厚生労働省は平成 27 年 12 月よ り事業者によるストレスチェックを義務化し、これによってストレスが労働者の精神 に及ぼす影響についての社会的な認識が高まっている。ストレスは、精神状態だけで なく循環器系、泌尿器系、消化器系などにも影響を及ぼし、いわゆる心身症を引き起 こす1)。 体温調節能や炎症反応は恒常性を維持するための重要な生理反応である。両者には 密接な関係があり、体温調節の中枢である視床下部における炎症反応は発熱反応と深 く関係している 2)。体温調節能と炎症反応は共にストレスにより影響を受けることが 報告されており2, 3)、ストレスによる発熱反応は、既存の抗炎症薬による治療が有効で ない場合も多く、臨床上問題となっている 4)。また、ストレスは炎症反応に関わり、 うつ病を増悪させる要因となることもある3)。 ストレスには、一過性の刺激によって起こる急性ストレスと刺激の持続や反復によ って起こる慢性ストレスがある。急性ストレス刺激により、視床下部-下垂体-副腎 皮質系(hypothalamic-pituitary-adrenal axis: HPA 系)や交感神経系の活性化が起こるこ とで副腎皮質刺激ホルモンの分泌亢進5, 6)や血圧7)・心拍数の増加8-10)、消化管運動の 低下・亢進11)などが生じ、慢性ストレス時にはそれらがより複雑化する。さらに、慢 性ストレスはストレス関連疾患の発症機序に大きく関わっていることが知られている 12, 13)。 急激な気温変化や季節の変わり目に体調不良を訴える人が多いことから、環境温度 変化はストレッサーの 1 つとして考えられている。喜多らにより考案された反復寒冷 (repeated cold; RC) ストレスは14)は、春先や秋口に見られる気温変動、夏季あるいは冬 季に室内外を出入りする場合の急激な気温の変化をシミュレートしたもので、マウス やラットを特殊なスケジュールに従って約 1 週間、室温と低温に繰り返し曝露するこ とで作製される。本ストレスを負荷した動物は、自律神経失調症モデルとして認めら れており、痛覚過敏15)、持続的低血圧16, 17)や起立性低血圧18)、心拍数増加17, 18)などを2 含む多様な生理的および病態生理的変化を示す 19)。また、脳内ではアセチルコリン、 ノルアドレナリン、セロトニンなどモノアミン類の含量や代謝回転に異常が認められ ている20, 21)。 慢性的なストレスが生体反応に影響を与えることは知られているが、その詳細なメ カニズムはわかっていない。そこで本研究では、ヒトにおいて慢性ストレスにより生 じる体調変化や関連する疾患を探るため、1 週間の RC ストレスを負荷したマウスが、 新たな急性ストレスや炎症性刺激に対してどの様な反応を示すかについて検討した。 第 1 章では RC ストレスが急性拘束ストレスによって誘発される体温上昇に影響を 与えるか否かを検討した。第 2 章、第 1 節では、マウスの深部体温を、テレメトリー システムを用いて無麻酔、無拘束で測定し、lipopolysaccharide (LPS) 誘起発熱反応に 対する RC ストレスの影響を検討した。第 2 章、第 2 節では、ヒトの間質性膀胱炎に 似た症状を示すシクロホスファミド誘起マウス膀胱炎モデルを用いて、RC ストレス が膀胱炎症および膀胱痛に与える影響を検討した。以上について、ヒトのストレス関 連疾患・病態との関係を考察した。
3
第 1 章 ストレス誘発体温上昇
に対する反復寒冷ストレスの影響
緒言
発熱は、患者が医師に訴える最も一般的な症状である。多くは、感染に伴う免疫系 の活性化により生じるが、ストレスが発熱の原因となることも報告されており、スト レス性高体温症として最近注目されている22, 23)。心理的なストレスによる体温上昇は、 ラット22, 24)やマウス25-27)、ヒト28-32)など様々な哺乳動物で観察される。急性ストレス による体温上昇には交感神経興奮を介した褐色脂肪組織におけるアドレナリン β3受 容体を介する非ふるえ産生熱が重要な役割を果たすことが報告されている33-35)。急性 ストレス誘発高体温には、骨格筋のふるえ産生熱、褐色脂肪組織の熱産生および皮膚 血管収縮による熱放散の低下のいずれもが関与する 2)。しかし、慢性的なストレスが 心因性発熱に及ぼす影響についてはよくわかっていない。慢性ストレスは、精神疾患、 循環器疾患、消化器疾患など様々な病態の悪化に、急性ストレス以上に大きく関わっ ているため12, 13)、慢性的なストレスが生体反応にどのような影響を及ぼすのか調べる ことは極めて重要である。 RC ストレス動物14)は、マウスやラットの飼育環境温度を特定のスケジュールに従 って繰り返し急変させながら約一週間飼育することで作製される。RC ストレス動物 では、抑うつ、不安などの精神症状20, 36)、痛覚過敏37)、低血圧16)、血小板機能低下38) などの多様な生体機能異常のほか、血中ノルアドレナリン濃度の上昇39)も見られるた め、自律神経失調症モデル動物として認められているほか、最近では線維筋痛症モデ ルとしても利用されている40, 41)。一方、慢性ストレスに長期的に負荷すると、異なる 急性ストレスに対する感受性が変化することも報告されていることから42, 43)、本章で は、一週間の RC ストレスを負荷したマウスにおいて、急性拘束ストレスによって誘 発される体温上昇反応が変化するか否かを検討し、そのメカニズムを解析した。4
I. 実験方法
実験動物として体重 20-30 g(約 4-5 週齢)の ddY 系雄性マウス(日本 SLC、静岡、 紀和実験動物、和歌山)を使用した。マウスの通常飼育は室温 24±1 ℃、12 時間毎 の明暗サイクル(8 時から 20 時まで点灯)下で行い、6-8 匹/1 ゲージとした。飼料 (MF、オリエンタル酵母)及び水は自由に摂取させた。全ての実験は、近畿大学動物 実験委員会によって承認され、米国国立衛生研究所による実験動物の管理と使用のた めのガイドラインに従って行った。 RC ストレスの負荷は秦らの方法44)に準じて行った。即ち、室温 24℃の飼育室と庫 内温度 4℃の動物飼育用チャンバーの両方にマウス飼育用ケージを用意し、マウスを、 毎日 9 時から 16 時までの間は 1 時間ごとに両ケージ間に移し変え、16 時から翌朝 9 時までは 4℃のチャンバー内で飼育するという環境温度変化に 7 日間曝すことにより 負荷した。実験当日の朝 11 時まで RC ストレスの負荷方法に従って入れ替えを行い、 室温に戻して 30 分以上経た後、実験に使用した。 3. 急性拘束ストレス負荷 マウスを通常飼育温度である 24℃にコントロールされた飼育室において、直径 3 ㎝ のプラスチックチューブで 1 時間拘束した。 4. 直腸温測定の方法 直腸温は、サーミスター温度計(夏目製作所、東京)を用いて測定した。すなわち、 プローブを肛門から約 2 cm 挿入し、拘束ストレス負荷直前、負荷 20、40、60 分後に 直腸温の測定を行った。 1. 実験動物 2. RC ストレス負荷5
5. 血清コルチコステロン濃度の測定
血漿サンプルは、拘束ストレス負荷直後に採取した。イソフルラン麻酔下、マウス の右心房より約 0.5 mL 採血し、室温にて 30 分間放置した後、6000 rpm、10 分間遠心 分離し、上清を回収し、-80℃で保存した。血清コルチコステロン(CORT)濃度の測 定は Corticosterone EIA Kit(Cayman Chemical Company, Michigan, USA)を使用した。
6. 褐色脂肪組織重量の測定 イソフルラン麻酔下、マウス心臓から生理食塩水 20 mL で灌流を行い背部肩甲骨下 の褐色脂肪組織を摘出し、電子天秤により秤量した。 7. Western blot 法 ウレタン (1.5 g/kg) 麻酔下、心臓から生理食塩水 20 mL で灌流を行ったマウスより 背部肩甲骨下の褐色脂肪組織を摘出し、液体窒素にて凍結した。各サンプルは、0.1 % phenylmethylsufonylfluoride, 0.15 U/mL aprotinin および 1 mM sodium orthovanadate を含 む RIPA バッファー (1 % PBS, 0.5 % sodium deoxycholate Igepal CA-630 および 0.1 % SDS) を加えホモジナイズおよび超音波処理した後,10000 ×g、4 ℃で 10 分間遠心分離 し、上清を採取した。Brad ford 法にてタンパク量を測定した後、上清に 2-mercatoethanol と bromophenol blue を加え、95 ºC で 5 分間煮沸処理した。サンプル中のタンパク質は 12.5 %の SDS-polyacrylamide gel (和光純薬工業、大阪)を用いて 2 時間の電気泳動 により分離し、分離させたタンパクをpolyvinylidene difluoride (PVDF) 膜 (Immobilon-P; Millipore Corporation、 Billerica, MA, USA) へ転写し、anti-UCP1 goat antibody (1:200) (Santa Cruz Biotechnology, CA, USA)、および anti-GAPDH rabbit antibody (1:5000) (Santa Cruz Biotechnology) を、5% skim milk を含む Tris-based solution/HCl buffer、0.1% Tween-20 (TBS/T buffer) でそれぞれの倍率に希釈して 4ºC で一晩反応させた。次に、 二次抗体としてHRP-conjugated anti-rabbit IgG 抗体 (1:5000) (Chemicon International, CA, USA) と anti-goat IgG(1:1000)(Chemicon International)を 5% skim milk を含む TBS/T buffer で希釈し、室温で 90 分反応させた後、chemiluminescence (ナカライテスク、京 都)で発光させ、Image Quant 400 (GE healthcare, Buckinghamshire, UK)でバンドを検出
6
した。これをパーソナルコンピューターに取り込んだ後、ImageJ45)を用いてバンドの 濃さを数値化した。
8. 使用薬物および投与スケジュール
実験にはジアゼパム(和光純薬工業、大阪)、インドメタシン、SR59230A (Sigma-Aldrich , St Louis, MO, USA)、ミフェプリストン(東京化成工業、東京)を用い た。ジアゼパムとミフェプリストンは 0.5%カルボキシメチルセルロースナトリウム (CMC・Na)に懸濁した。インドメタシンは炭酸水素ナトリウムで溶解した。SR59230A は生理食塩液に溶解した。ジアゼパム 4 mg/kg およびミフェプリストン 50 mg/kg は、 拘束ストレス開始 60 分前に経口投与した。インドメタシン 5 mg/kg は拘束ストレス開 始 30 分前に皮下投与した。SR59230A 5 mg/kg は拘束ストレス開始 30 分前に腹腔内投 与した。 9. 統計処理 得られた実験値は、平均値 ± 標準誤差で示した。統計学的有意性の判定には 2 群 間の場合はStudent’s-t検定を用い、3 群以上の比較にはTukey法を用いて検討し、P<0.05 で有意差ありとした。
7
II. 実験結果
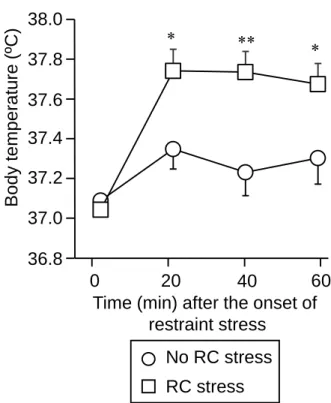
Fig. 1 はマウスに対して 60 分間の拘束ストレスを付加した時の体温を経時的に測定 した結果である。RC ストレスにより明らかな急性拘束ストレス負荷により非 RC スト レス群および RC ストレス群で体温上昇が認められたが RC ストレス群における体温 上昇がより顕著であった。(Fig. 1) 。Fig. 1. Effect of RC stress on acute restraint stress-induced hyperthermia in mice. Data show the mean with
S.E.M. for 11-16 mice. *P<0.05, **P<0.01, vs. the non-RC group.
36.8 37.0 37.2 37.4 37.6 37.8 38.0 B o d y te m p e ratu re (ºC)
Time (min) after the onset of restraint stress 0 20 40 60 ** * * RC stress No RC stress 1. 急性拘束ストレス誘発体温上昇に対する RC ストレスの影響
8 交感神経の興奮により誘起される熱産生には褐色脂肪組織のβ3受容体が関与する33) ことから、RC ストレスによる急性拘束ストレス誘発体温上昇の亢進への β3受容体の 関与を検討した。β3受容体阻害薬である SR59230A 5 mg/kg は急性拘束ストレス負荷 1 時間前の腹腔内投与により RC ストレスによる急性拘束ストレス誘発体温上昇の亢進 を有意に抑制した (Fig. 2A) 。また、マウス肩甲骨にある褐色脂肪組織および褐色脂 肪組織のマーカーである UCP1 タンパク発現量は RC ストレスにより増加傾向を示し た (Fig. 2B, C) 。 2. RC ストレスによる急性拘束ストレス誘発体温上昇の亢進に対する β3受容体阻害 薬 SR59230A 投与の効果
9
Fig. 2. Effect of SR59230A, a β3 adrenoceptor antagonist, on the enhancement of acute restraint stress-induced
hyperthermia by RC stress, and protein levels of UCP1 in the brown adipose tissue (BAT) of RC-stressed mice. (A) The mice received a single i.p. administration of SR59230A at 5 mg/kg or vehicle 60 min before the onset of restraint stress. The acute restraint stress-induced hyperthermia (ΔT) is presented as the difference of the body temperature before and after restraint stress for 0, 20, 40 or 60 min. (B) and (C) BAT weight, the one relative to the body weight (B) and protein levels of UCP1 in the BAT tissue, as assessed by Western blotting (C) in RC-stressed mice. Typical photographs for Western blotting and quantified data by densitometry are shown by densitometry (C). Data show the mean with S.E.M. for 10-14 or 4-5 (B, C) mice. **P<0.01 vs. the vehicle-treated, non-RC group; †P<0.05, ††P<0.01 v.s. the vehicle-treated, RC group.
Time (min) after the onset of restraint stress No RC / Vehicle RC / Vehicle RC / SR59230A No RC / SR59230A 0 20 40 60 -0.4 0 0.4 0.8 1.2 † †† † ** ** ⊿ T (º C ) 0 50 100 150 200 250 300 B A T w e ig h t (m g ) 0 2 4 6 8 10 12 BA T w eig ht /b o d y w e ig h t (m g /g ) No RC RC No RC RC 0 0.1 0.2 0.3 0.4 0.5 0.6 No RC RC UCP1 GAPDH UC P 1 /G A P DH (Arbitrary u n it) (A) (B) (C)
10
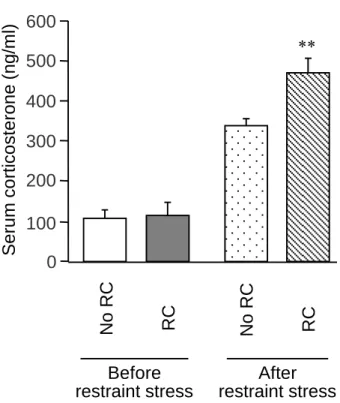
Fig. 3 は非ストレス群および RC ストレス群における急性拘束ストレス負荷後の血 清コルチコステロンを測定した結果である。拘束ストレス負荷前の血清コルチコステ ロン濃度に変化は認められなかったが、RC ストレスにより明らかな拘束ストレス誘 発コルチコステロン分泌の亢進が認められた (Fig. 3) 。
Fig. 3. Effect of RC stress on the secretion of corticosterone in response to acute restraint stress in mice. Blood
was collected before and after 60-min restraint stress in mice. Data show the mean with S.E.M. for 12-22 mice. **P<0.01 vs. the non-RC group.
Fig. 4A はRC ストレスマウスにおける拘束ストレス誘発体温上昇亢進にコルチコス テロン産生増加が関与するか否かを検討するため、グルココルチコイド受容体阻害薬 ミフェプリストンの効果を調べた結果である。拘束ストレス負荷 1 時間前におけるミ フェプリストン 50 mg/kg の経口投与は急性拘束ストレス誘発体温上昇の亢進には影 0 100 200 300 400 500 600 Before restraint stress N o RC RC S e ru m co rtico st e ro n e (n g /m l) ** After restraint stress N o RC RC 3. 急性拘束ストレス誘発コルチコステロン分泌に対する RC ストレスの影響 4. RC ストレスによる急性拘束ストレス誘発体温上昇の亢進に対するミフェプリス トンおよびジアゼパム投与の効果
11
響しなかった。また Fig. 4B は RC ストレスの不安様行動に対して効果を示すベンゾ ジアゼピン系抗不安薬ジアゼパムの効果を調べた結果で、ジアゼパム 4 mg/kg の経口 投与によっても RC ストレスによる急性拘束ストレス誘発体温上昇の亢進に変化は認 められなかった。
Fig.. 4 Influence of mifepristone, a glucocorticoid receptor antagonist, and diazepam, anxiolytic, on the
enhancement of acute restraint stress-induced hyperthermia in RC-stressed mice. The mice received a single oral administration of mifepristone at 50 mg/kg (A) or diazepam at 4 mg/kg (B) 1 h before 20-min restraint stress. The acute restraint stress-induced hyperthermia (ΔT) is presented as the difference of the body temperature before and after 20-min restraint stress. Data show the mean with S.E.M. for 6-9 mice. *P<0.05, vs. the vehicle-treated, non-RC group.
プロスタグランジン E2 (PGE2) は、感染性の発熱において関与が示唆されているこ とから、Fig. 5 では RC ストレスにより誘起される拘束ストレス誘発体温上昇亢進への シクロオキシゲナーゼ (COX) の関与を検討するため、COX 合成阻害薬であるインド メタシンの効果を検討した。拘束ストレス負荷 30 分前におけるインドメタシン 5 mg/kg の皮下投与は、RC ストレスによる拘束ストレス誘発体温上昇の亢進に対し効果 を示さなかった (Fig. 5) 。 M if e p ri sto n e V e h icl e No RC RC 0 0.2 0.4 0.6 0.8 1.0 1.2 0 0.2 0.4 0.6 0.8 1.0 ⊿ T ( ºC ) 1.2 * M if e p ri sto n e V e h icl e Di a z e p a m V eh ic le No RC RC Di a z e p a m V e h icl e * 5. RC ストレス状態における拘束ストレス誘発体温上昇に対するインドメタシ投与 の効果 (A) (B)
12
Fig. 5. Influence of indomethacin, a cyclooxygenase inhibitor, on the enhancement of acute restraint
stress-induced hyperthermia in RC-stressed mice. The mice received a single s.c. administration of indomethacin at 5 mg/kg or vehicle before the onset of restraint stress. The acute restraint stress-induced hyperthermia (ΔT) is presented as the difference of the body temperature before and after restraint stress for 0, 20, 40 or 60 min. Data show the mean with S.E.M. for 7-9 mice. *P<0.05, **P<0.01 vs. the vehicle-treated, non-RC group.
III. 考察
本章において、RC ストレスマウスでは、拘束ストレスによる体温上昇および急性 ストレス反応の指標であるコルチコステロン分泌が増加することが明らかとなった。 さらに、RC ストレスによる拘束ストレス誘発体温上昇の亢進がアドレナリン β3受容 体阻害薬により抑制されたことにより、ストレス誘発体温上昇の亢進に交感神経、褐 色脂肪組織を介した熱産生が重要な役割を果たしていることが示唆された。 生体における体温調節の中枢は脳の視索前野であり、感染性発熱時は視索前野 PGE2/EP3 受容体系を介して視床下部背内側核-延髄縫線核系神経回路が活性化する ことで交感神経が興奮し、骨格筋におけるふるえ産生熱、褐色脂肪組織における非ふ るえ産生熱、皮膚血管収縮による熱放散抑制の結果、体温上昇がおこる46-48)。心理的Time (min) after the onset of restraint stress No RC / Vehicle RC / Vehicle RC / Indomethacin No RC / Indomethacin ** * -0.4 0 0.4 0.8 1.2 1.6 0 20 40 60 ** ⊿ T (º C )
13 ストレスにより誘発される体温上昇にも、視床下部背内側核-延髄縫線核系の神経回 路を介する熱産生が関与することが報告されている 49)。一方、EP3 受容体ノックア ウトマウスでは LPS による発熱は起こらないが、急性ストレスによる体温上昇は認 められる26)。このようにストレス誘発高体温にプロスタノイドが関与しないことは、 RCストレスマウスにおける拘束ストレス誘発体温上昇の亢進にインドメタシンが影 響しなかった本章の結果に矛盾しない (Fig. 5) 。RC ストレス動物は不安状態にある ことが報告されており50, 51)、ストレス誘発高体温に不安が関与すること52)が知られ ている。しかし、抗不安薬ジアゼパムが RC ストレスマウスにおける拘束ストレス誘 発体温上昇の亢進に影響しなかったことから (Fig. 4B) 、この体温上昇反応亢進に不 安は関与しないことが示唆される。 社会敗北ストレスにより誘発される体温上昇がアドレナリン β3受容体阻害薬によ って抑制されることが報告されている33)。褐色脂肪組織ではノルアドレナリン刺激に より熱産生が生じ、反復拘束ストレスはノルアドレナリンによる熱産生を増強させる 53)。本章において RC ストレスによる拘束ストレス誘発体温上昇の亢進が β 3受容体阻 害薬で抑制されたことから (Fig. 2A) 、交感神経-β3受容体を介した褐色脂肪組織にお ける非ふるえ産生熱が RC ストレスにより増強された可能性が示唆される。このこと は、RC ストレス動物では血漿ノルアドレナリン濃度が顕著に増加している39)との報 告や、今回、褐色脂肪組織の重量および UCP1 タンパク質発現量に増加傾向がみられ たこと (Fig.2B, C) により支持される。 グルココルチコイドは COX-2 誘導に伴うPGE2産生を抑制することが知られており 54)、感染性発熱にも影響を与える46, 47, 55)。しかし、グルココルチコイドがストレス誘 発高体温に関与するか否かはまだよく分かっていない。そこでグルココルチコイド受 容体がストレス誘発高体温に影響を与えるかを検討するため、グルココルチコイド受 容体阻害薬であるミフェプリストンを投与したところ、RC ストレスによるストレス 誘発高体温の亢進に影響を与えなかった (Fig. 4A) 。このことより、RC ストレスマウ スにおける拘束ストレス誘発コルチコステロンの増加は、体温上昇反応の亢進には寄 与しないと考えられる。 RC ストレスを 1 週間負荷したマウスでは血中コルチコステロン濃度の恒常的な増 加は見られない (Fig. 3) 。しかし、RC ストレスとは異なるストレスである拘束スト
14 レスの急性負荷により誘起されるコルチコステロン分泌は、RC ストレスマウスにお いて亢進している (Fig. 3) 。これはストレスに繰り返しさらされることで同種のスト レスに対しては HPA 系に「慣れ」が生じるが56, 57)、異なるストレスに対してはむし ろ反応が増大するという以前の報告58, 59)と矛盾しない。この慢性ストレス負荷後の異 なる急性ストレスに対する HPA 系の反応性上昇には、グルココルチコイドのネガテ ィブフィードバック機能の低下60)や海馬の ERK 活性の変化61)が関与することが示唆 されている。 以上より RC ストレスマウスにおける拘束ストレス誘発体温上昇の亢進には、内因 性プロスタノイドやグルココルチコイド、不安症状は関与せず、交感神経系-アドレ ナリン β3受容体系の過剰興奮による褐色脂肪組織における熱産生の増強が寄与する ことが示唆された。
15
第 2 章 炎症性刺激に対する反応性
に及ぼす反復寒冷ストレスの影響
緒言
急性ストレス刺激は、視床下部-下垂体-副腎皮質系を活性化し、グルココルチコイ ドの分泌を促進する62)。グルココルチコイドは、炎症を抑制するが、免疫反応を抑制 するため、易感染状態を誘起する可能性がある。実験動物において、急性ストレス負 荷により交感神経系活性化を介した炎症性サイトカインの産生増加がおこる 63)一方、 慢性的なストレスがグルココルチコイド受容体を介した炎症抑制作用を減弱するとの 知見も報告されており64)、ストレスは炎症反応に対して抑制および促進の両面的な作 用を有することが考えられる。そこで本章では、炎症性刺激に対する反応性に及ぼす 慢性ストレスの影響を明らかにすることを目的として、第 1 節では LPS による発熱反 応、第 2 節では、抗がん薬シクロホスファミドによって誘起される膀胱炎・膀胱痛に RC ストレスがどのような影響を及ぼすかを検討した。16
第 1 節 Lipopolysaccharide 誘起発熱反応
に対する反復寒冷ストレスの影響
発熱は、感染症や全身性炎症に対する生理反応の 1 つで、免疫系を活性化し、ウイ ルスや細菌の増殖を抑制する 65)。感染性発熱反応には、炎症により発現増加する COX-2 によって産生される PGE2が関与することが明らかにされている66)。実験動物 における LPS の全身性投与は、炎症反応の研究によく用いられるモデルである。マウ スやラットに LPS を投与すると、2 峰性の発熱反応がみられることが報告されている が、どちらの発熱反応にも COX-2/PGE2系が関与する66)。第 1 章で示したように、急 性ストレスによって発熱がおこることがあるが、炎症性の発熱反応とは、機序が異な っているようである 2)。一方、慢性ストレスを負荷した動物において、炎症性の発熱 反応がどのように変化するはよくわかっていない。 慢性的ストレスは、様々な疾患を誘起したり悪化させたりすることが知られており、 RC ストレスを負荷した動物も多様な病的症状を示すが、致死量の LPS 投与に対して はむしろ耐性を示すことが報告されている67)。この RC ストレスマウスにおける LPS 耐性発現の機序はまだよく分かっていない。そこで第 2 章、第 1 節ではテレメトリー システムを用いてマウスの体温を無麻酔・無拘束下で測定し、LPS 誘起発熱反応に対 する RC ストレスの影響を検討した。I. 実験方法
実験動物として体重 20-30 g(約 4-5 週齢)の ddY 系雄性マウス(日本 SLC、静岡、 紀和実験動物、和歌山)を使用した。マウスの飼育は第 1 章と同様に行った。 反復寒冷ストレス負荷は第 1 章と同様に行った。 1. 実験動物 2. RC ストレス負荷17
マウスをペントバルビタール (10 mg/kg) (大日本住友製薬、東京)、メデトミジン (0.3 mg/kg) (Meiji Seika ファルマ、東京) およびミダゾラム (4 mg/kg) (Sandoz Inc、東京) の 腹腔内投与による全身麻酔下、正中切開し、体温計測用のテレメトリー送信機 (TA-F10, Data Sciences International, Saint Paul, MN, USA) をマウス腹腔に埋め込み、2 日間の回復期間の後、実験に供した。送信シグナルはデータ解析ソフト (Dataquest A.R.T Silver, Data Sciences International, Saint Paul, MN, USA) により解析し、10 秒ごとに 測定した体温の平均値を算出した。
深部体温の日内変動の測定は、体温を 40 分おきに 24 時間測定し、ストレス負荷前 と、ストレス負荷最終日に測定を行った。
深部体温は少なくとも LPS (1 mg/kg) (Escherichia coli 055:B5, Sigma) 投与 30 分前か ら測定し、投与直前の体温のベース体温として使用した。LPS の投与は11:30 から 15:00 までの間で行い、投与後 140 分間深部体温を測定した。 血液サンプルは、LPS 投与 20、90、150 分後に採取し、第 1 章と同様の方法で血清 コルチコステロン濃度を測定した。 ウレタン (1.5 g/kg) 麻酔下、マウス心臓から生理食塩水 20 mL で灌流を行い右肺の 中葉、および視床下部を摘出し、液体窒素にて凍結した。ウェスタンブロット法は第 1 章と同様に行った。すなわち各サンプルは、第 1 章と同様の方法で調整し、サンプ ル中のタンパク量を 12.5%の SDS-polyacrylamide gel(和光純薬工業)を用いて 2 時間 の電気泳動により分離させた。分離させたタンパクを PVDF 膜へ転写し、anti-COX-2 3. テレメトリーシステムによる深部体温の測定 4. 概日リズムの測定 5. LPS 誘起発熱の測定 6. 血清コルチコステロン濃度の測定 7. Western blot 法
18
goat polyclonal antibody (1:1000) (Santa Cruz Biotechnology, CA, USA)、およびanti-GAPDH rabbit antibody (1:5000) (Santa Cruz) を、5% skim milk を含む Tris-based solution/HCl buffer、 0.1% Tween-20 (TBS/T buffer) でそれぞれの倍率に希釈して 4ºC で一晩反応させた。次 に、二次抗体として HRP-conjugated anti-rabbit IgG 抗体 (1:5000) (Chemicon International, CA, USA) と anti-goat IgG (1:3000) (Chemicon International) を 5% skim milk を含む TBS/T buffer で希釈し、室温で 90 分反応させ、chemiluminescence(ナカライテスク) で発光させ、Image Quant 400 で陽性バンドを検出した。これをパーソナルコンピュー ターに取り込んだ後、ImageJ を用いてバンドの濃さを数値化した。 ジクロフェナク(和光純薬工業)は生理食塩水に溶解して LPS 投与 1 時間前に経口 投与した。 統計処理は第 1 章と同様の検定方法を用いて行った。P<0.05 で有意差ありとした。 8. 使用薬物および投与スケジュール 9. 統計処理
19
II. 実験結果
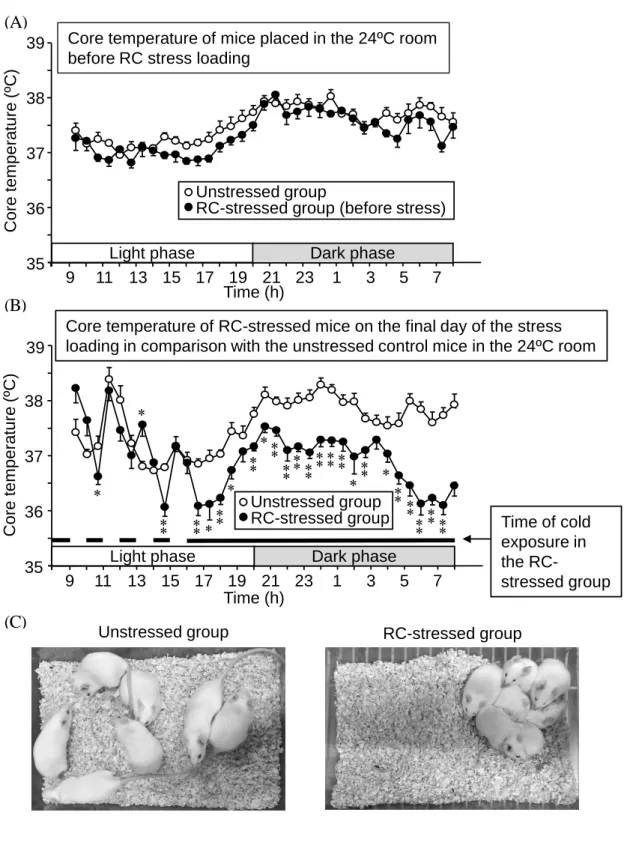
始めにテレメトリー装置を用いて RC ストレス負荷前および負荷最終日の深部体温 を測定した。非ストレスマウスおよび RC ストレス負荷前のマウスの深部体温は日中 (8:00-20:00)と比較して夜間(20:00-8:00)に上昇する傾向が認められた (Fig. 6A) 。 一方、RC ストレス負荷最終日、低温環境にある夜間帯の深部体温は、非ストレスマ ウスと比較して有意に低下していた (Fig. 6B) 。この夜間帯 (21:00) において、低温環 境にいる RC ストレスマウスの動きは少なく、室温コントロールマウスと比較して明 らかに活動性が低下していた (Fig. 6C) 。 1. 日内体温変動に対する RC ストレスの影響20
Fig. 6. Circadian fluctuations of core temperature in RC-stressed and unstressed mice. (A) Core temperature
for 24 h before exposure to RC stress. (B) Core temperature for 24 h on the final day of RC stress loading. (C) Typical photographs of the unstressed mice in the 24ºC room and RC-stressed mice in the 4ºC room at 21:00 (dark phase) on the final night of RC stress loading. Data show the mean ± S.E.M. from 7 mice. *P<0.05, **P<0.01 vs. the unstressed group.
Core te m p e ra tu re ( ºC ) 35 36 37 38 39 9 11 13 15 17 19 21 23 1 3 5 7 Time (h) * * * * * * * Core te m p e ra tu re (ºC)
Core temperature of RC-stressed mice on the final day of the stress loading in comparison with the unstressed control mice in the 24ºC room
Time of cold exposure in the RC-stressed group 35 36 37 38 39 9 11 13 15 17 19 21 23 1 3 5 7
RC-stressed group (before stress) Unstressed group
Core temperature of mice placed in the 24ºC room before RC stress loading
Time (h) ** ** ** ** **** **** ** ** ** ** ** ****** ** RC-stressed group Unstressed group
Light phase Dark phase
Light phase Dark phase
RC-stressed group Unstressed group
(A)
(B)
21
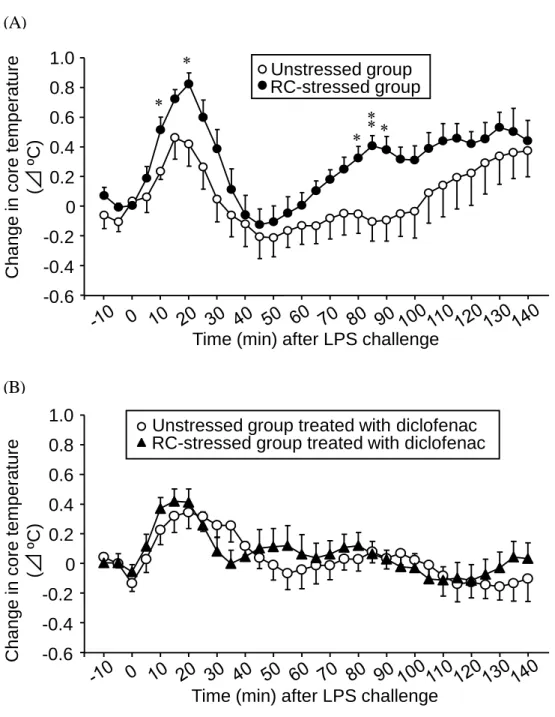
マウスに LPS (1 mg/kg) を腹腔内投与することにより、投与 10-30 分後および 70-140 分後に体温が上昇し、2 峰性の発熱反応が認められた (Fig. 7A) 。この投与 10、20 お よび 80-90 分後の発熱反応は非ストレス群と比較して RC ストレスマウスにおいて有 意に増加していた (Fig. 7A) 。LPS 誘起発熱反応に関与することが知られる COX-2 66) の阻害薬であるジクロフェナク(30 mg/kg)を投与したところ、RC ストレスマウスに おける LPS 誘起発熱反応の増強は消失した (Fig. 7B) 。なお、LPS 投与直前の体温は、 ジクロフェナクを投与していない非ストレスマウスと RC ストレスマウスでは、それ ぞれ 37.0 ± 0.1 (n = 7) と RC ストレス群は 37.0 ± 0.1 (n = 8) 、また、ジクロフェナク を投与した非ストレスマウスと RC ストレスマウスでは、それぞれ 36.9 ± 0.1 (n = 6) と 37.0 ±0.1 (n = 8) であり、ジクロフェナク単独投与は、LPS 投与前の基礎体温に影 響しなかった。 2. LPS 誘起発熱反応に対する RC ストレスの影響およびジクロフェナク経口投与の 効果
22
Fig. 7. LPS-induced hyperthermia in RC-stressed and unstressed mice. (A) Core temperature was monitored
following i.p. administration of LPS at 1 mg/kg in RC-stressed and unstressed mice. (B) The mice received oral administration of diclofenac at 30 mg/kg, a cyclooxygenase inhibitor, 60 min before LPS injection. Data show the mean ± S.E.M. from 7-8 (A) or 6-8 (B) mice. *P<0.05, **P<0.01 vs. the unstressed group.
Cha n g e in co re te m p e ratu re (⊿ ºC) -0.6 -0.2 0 0.2 0.4 0.6 0.8 1.0 -0.4
Time (min) after LPS challenge
RC-stressed group treated with diclofenac Unstressed group treated with diclofenac
Cha n g e in co re te m p e ratu re (⊿ ºC) -0.6 -0.2 0 0.2 0.4 0.6 0.8 1.0
Time (min) after LPS challenge -0.4
* *
* * RC-stressed groupUnstressed group
**
(A)
23 LPS 誘起発熱には肺および視床下部における COX-2 発現が重要な役割を果たすと の報告がある55)。そこで LPS 投与 40 および 90 分後の肺および視床下部の COX-2 発 現量をウェスタンブロット法により検討した。その結果、肺および視床下部の COX-2 発現は LPS による影響は認められず、さらに RC ストレスによっても変化しなかった (Fig. 8A, B, C) 。 第 1 章と同様に RC ストレスは通常のコルチコステロン濃度に影響を与えなかった (Fig. 8D) 。LPS 誘起発熱にコルチコステロンが関与するかを検討するため、LPS 投与 20、90、150 分後のコルチコステロン濃度を測定した。その結果 LPS 投与 90 分後には コルチコステロン濃度の上昇が見られたが、RC ストレスによる影響は認められなか った (Fig. 8A, D) 。一方、LPS 投与 150 分後のコルチコステロン濃度は非ストレス群 と比較して RC ストレス群において高値となった (Fig. 8D) 。 3. RC ストレスマウスにおける肺および視床下部の COX-2 発現量に対する LPS の 効果 4. LPS 誘起コルチコステロン分泌に対する RC ストレスの影響
24
Fig. 8. Expression of COX-2 protein in the lung and hypothalamus, and serum corticosterone levels after LPS
injection in RC-stressed and unstressed mice. (A) Schedule for blood sampling and tissue isolation after i.p. administration of LPS at 1 mg/kg. (B) and (C) Western blot analysis of COX-2 protein levels in the lung (B) and hypothalamus (C) 40 min and 90 min after i.p. LPS. Top and bottom panels show typical photographs of Western blotting and the quantified data by densitometry, respectively. (D) Serum corticosterone levels 20, 90 and 150 min after i.p. LPS. Data show the mean ± S.E.M. for 4 (B), 4-5 (C) or 4-9 (D) mice. ††P<0.01 vs. vehicle in the unstressed or RC-stressed mice; *P<0.05 vs. LPS in the unstressed mice.
Unstress + LPS Unstress + vehicle RC + LPS RC + vehicle 0 200 400 600 S e rum co rti co ste ron e (ng /m l) 20 90 150 * 0
The time after i.p. LPS (min) ††
††
†† ††
0 20 40 90 150
LPS (i.p.) The collection of blood
The isolation of tissues
(min) Hypothalamus 800 Unst ress RC st ress 0.4 0.6 0.8 1.0 1.2 1.4 0.2 0 V e h icl e L P S V e h icl e L P S COX -2 / GA P DH (Arbitrary u n it) COX-2 GAPDH 0.4 0.6 0.2 0 Lung 40 min Unst ress RC st ress V e h icl e LP S V e h icl e L P S 0 0.2 0.4 0.6 0.8 1.0 0 0.1 0.2 0.3 COX -2 / GA P DH (Arbitrary u n it) COX-2 GAPDH 90 min Unst ress RC st ress V e h icl e L P S V e h icl e L P S Unst ress RC st ress V e h icl e L P S V e h icl e L P S Lung Hypothalamus (A) (B) (C) (D)
25
III. 考察
本節では、RC ストレスマウスでは LPS により誘起される COX 依存性発熱反応が 亢進していることが明らかになった。RC ストレスマウスは致死量の LPS 投与に対し て耐性を示すことが報告されており67)、この耐性獲得に、今回認められた LPS 誘起発 熱反応の増強が寄与している可能性が考えられる。 今回、テレメトリー装置により RC ストレス負荷中の体温測定が可能となった。RC ストレス負荷最終日における低温環境下での深部体温の低下は、自発運動の低下と関 係している可能性があり、低温下での過剰なエネルギー消費を防ぐための適応現象の 1つではないかと考えられる。LPS によって誘起される発熱反応に免疫細胞から分泌 された炎症性サイトカインにより肺や肝臓、脳血管内皮で誘導される PGE2合成酵素 COX-2 が関与することが知られている55, 68, 69)。産生された PGE2は視床下部視索前野 から視床下部背内側核、延髄縫線核へと投影される GABA 作動性神経を抑制すること で発熱反応を引き起こす46-48)。LPS によって肺や肝臓での COX-2 の誘導が起こること 報告されている55)が、本実験においては有意な変化が認められなかった (Fig. 8B, C) 。 この矛盾の原因はまだ分からないが、ホスホリパーゼ A2、COX-1、PGE 合成酵素など PGE2産生に関与する COX-2 以外の酵素の発現量変化も今後調べていく必要があると 考える。 RC ストレスマウスでは、LPS 腹腔内投与 10-20 分後および 80-90 分後の発熱反応が 増強されていたが、コルチコステロン濃度上昇の亢進は、150 分後にしか認められな かった (Fig. 8D) 。コルチコステロンの前処置により LPS 誘起発熱が増強されること が報告されているが70)、本節の結果は RC ストレスによる LPS 誘起発熱の増強に内因 性のグルココルチコイドは関与しないことを示唆している。LPS は Toll-like receptor 4 (TLR4) を介して炎症を引き起こすことから、RC ストレスによる LPS 誘起発熱の増強 に TLR4 の発現増加が関与する可能性も考えられるが、RC ストレスマウスは、致死 量の LPS 投与に対して耐性を示すことが報告されている67)ことから、その可能性も考 えにくく、今後さらなる検討が必要である。26
以上、本節の結果より、RC ストレスによって LPS 誘起 COX 依存性発熱反応が増 強されることが明らかとなり、これは LPS に対する耐性獲得に寄与している可能性も 考えられる。
27
第 2 節 シクロホスファミド誘起膀胱炎・膀胱痛への
反復寒冷ストレスの影響
間質性膀胱炎は強い膀胱痛を伴う難治性疾患でその病因は未解明な部分が多い。ヒ トの間質性膀胱炎に似た症状を示す実験動物モデルであるシクロホスファミド (CPA) 誘起膀胱炎マウス71)では、膀胱痛の発現に知覚神経上に豊富に発現する Cav3.2T 型カルシウムチャネル72)および壊死細胞や活性化マクロファージなどから分泌される 核内タンパク high mobility group box 1 が関与することが報告されている73)。一方、間 質性膀胱炎患者の膀胱痛はストレスにより悪化することが報告されている74)。そこで、 RC ストレスマウスにおいて、シクロホスファミドを投与し膀胱炎・膀胱痛の程度を、 非ストレスマウスと比較検討した。また、シクロホスファミド処置マウスの膀胱組織 にマクロファージの浸潤が見られるとの報告75)もあることより、マクロファージ機能 の変化との関連性について検討した。I. 実験方法
実験動物として体重 20-30 g(約 4-5 週齢)の ddY 系雄性マウス(紀和実験動物、和歌 山)を使用した。マウスの飼育は第 1 章と同様に行った。 反復寒冷ストレス負荷は第 1 章と同様に行った。 既報71, 72)に従い、マウスに、シクロホスファミド (Sigma-Aldrich) 200 - 400 mg/kg を 腹腔内に投与することにより膀胱炎を誘起した。 1. 実験動物 2. RC ストレス負荷 3. シクロホスファミド誘起マウス膀胱炎モデルの作製28 既報の方法71, 72)に従い、シクロホスファミド腹腔内投与 3 時間半から 4 時間後、マ ウスを網目が 5 mm 四方の金網に置き、透明なプラスチックケース (23.5×16.6×12.4 cm) を被せ、尿道口周囲皮膚表面に対するなめおよび下腹部のひきずり行動を指標と し、この反応回数を 30 分間測定した。 既報の方法72)に準じて、シクロホスファミド投与 4 時間後に、0.008、0.07、0.4、お よび 1.0 g の強度の異なる 4 種類の von Frey filament により尿道口から肛門付近の皮膚 表面を刺激した。フィラメントによる刺激は弱いものから強いものへと順に行い、各 フィラメントあたり 5-10 秒の間隔で 10 回刺激を行った。刺激に伴う侵害受容反応は 以下に示した通りにスコア化し、結果は 10 回の刺激によって得られたスコアの合計値 で示した。 0 点:反応なし 1 点:外尿道口周囲への舐めあるいは咬みつき、位置を変える、体幹を前屈させる、 上体を起こす、肢をばたつかせる 2 点:ジャンプする 膀胱痛関連痛覚過敏の測定後にマウスを安楽死させた後、膀胱を摘出し、膀胱湿重 量を測定した。 マウスを頸椎脱臼した後、40 ml のリン酸緩衝液を用いて腹腔マクロファージを採 取し、1000 rpm、4ºC で 5 分間遠心分離し、35 mm プラスチック dish に 1×105 cells/dish となるように細胞を播種し、10%FCS を含む RPMI-1640 培地で 37℃、5%CO2の条件 下で 24 時間培養後、培養液を交換した後 lipopolysaccharide (LPS) (Escherichia coli
4. シクロホスファミド誘起膀胱痛様侵害受容行動の測定
5. シクロホスファミド誘起膀胱関連痛覚過敏の評価
6. シクロホスファミド誘起膀胱炎の重症度の評価
29
026:B6) (Sigma-Aldrich) で 24 時間刺激した。上清を回収し TNF-α、IL-1β 量を ELISA キット (eBioscience, San Diego, CA, USA) を用いて測定した。
シクロホスファミドは、生理食塩水で溶解し、200 - 400 mg/kg の用量で腹腔内投与 した。ミノサイクリン (Sigma-Aldrich) は、生理食塩水で溶解しシクロホスファミド 投与 1 および 24 時間前に 30 mg/kg の用量で腹腔内投与した。
得られた実験値は、平均値 ± 標準誤差で示した。統計学的有意性の判定には 2 群 間の場合はstudent’s-t 検定を用い、3 群以上の比較には Tukey 法を用いて検討し、P<0.05 で有意差ありとした。von Frey test の統計学的有意性の判定には、Kruskal-Wallis H 検 定の後に、LSD 型多重比較検定によるノンパラメトリック解析を行った。
II. 実験結果
マウスにシクロホスファミド 400 mg/kg の腹腔内投与することにより投与 3 時間 半から4 時間後に膀胱痛様侵害受容行動および、また尿道口から肛門付近の皮膚表面 における関連痛覚過敏が認められ (Fig. 9A, B) 、膀胱相対湿重量が増加していた (Fig. 9C) 。しかし、これらの反応の程度に非ストレスマウスと RC ストレスマウスで 差は見られなかった (Fig. 9A, B, C) 。 マクロファージ/ミクログリア阻害薬ミノサイクリンは、30 mg/kg の腹腔内投与に より、非ストレスマウスおよびRC ストレスマウスにおけるシクロホスファミド誘起 侵害受容反応および関連痛覚過敏を有意に抑制した (Fig. 9A, B) 。一方、ミノサイク リンは、膀胱湿重量増加には影響しなかった (Fig. 9C) 。 8. 使用薬物および投与スケジュール 9. 統計処理 1. 高用量シクロホスファミドにより誘起される膀胱炎・膀胱痛へのマクロファージ の関与と RC ストレスの影響30
Fig. 9 Effect of minocycline (Mino), a macrophage/microglia inhibitor, on the bladder pain-like nociceptive
behavior, referred hyperalgesia and increased bladder weight after systemic administration of CPA at 400 mg/kg in unstressed (US) and RC-stressed mice. Minocycline at 30 mg/kg was administered i.p. twice 1 h and 24 h before i.p. administration of CPA at 400 mg/kg. Data show the mean ± SEM for 7-9 unstressed or 4-5 RC-stressed mice. *P<0.05, **P<0.01 vs. Vehicle (V) + V; †P<0.05, ††P<0.01 vs. V + CPA.
0 0.5 1.0 1.5 2.0 2.5 3.0 0 5 10 15 20 25 30 (A) Num b e r o f n o cicep tiv e b e h a v ior fo r 3 0 m in 0 5 10 15 20 0 5 10 15 20 N ocicep tiv e score V CPA 400 V M in o V V V Min o V US RC CPA 400 (i.p.) (i.p.) (mg/kg) 0.008 g Filament 0.07 g Filament 0.4 g Filament 1.0 g Filament US V V M in o V CPA 400 (i.p.) (i.p.) (mg/kg) V V M in o V CPA 400 V V M in o V CPA 400 V V M in o V CPA 400 RC V V M in o V CPA 400 (i.p.) (i.p.) (mg/kg) V V M in o V CPA 400 V V M in o V CPA 400 V V M in o V CPA 400 Nocice p tiv e sco re (B) B lad d e r w e ig h t / b o d y w e ig h t (m g /g ) (C) V CPA 400 V M in o V V V M in o V CPA 400 (i.p.) (i.p.) (mg/kg) US RC ** † ** † ** ** ** ** ** ** * * * †† †† † † † **
31
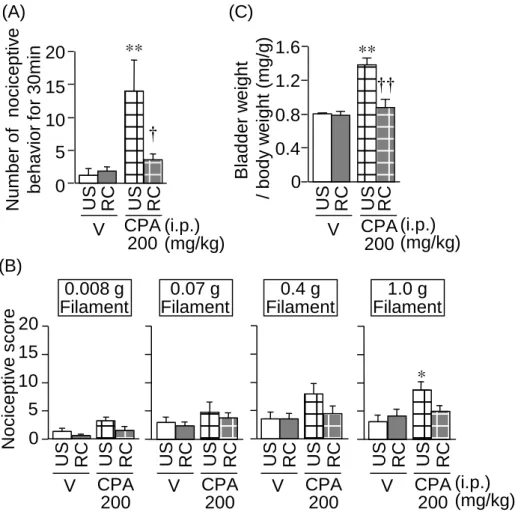
シクロホスファミドの用量を 200 mg/kg に下げて腹腔内投与し、誘起される膀胱 炎・膀胱痛に及ぼすRC ストレスの影響を調べた。その結果、RC ストレスマウスで は、低用量シクロホスファミド誘起侵害受容反応、関連痛覚過敏、および膀胱湿重量 が、非ストレスマウスに比べて、有意に減少していた (Fig. 10) 。
Fig. 10 Effect of RC stress on the bladder pain-like nociceptive behavior, referred hyperalgesia and increased
bladder weight caused by a low dose, 200 mg/kg, of CPA in mice. CPA at 200 mg/kg was administered i.p. to unstressed (US) and RC-stressed mice. Data show the mean ± SEM for 5-6 mice. *P<0.05, **P<0.01 vs. US treated with vehicle (V). †P<0.05, ††P<0.01 vs. US treated with CPA.
(i.p.) (mg/kg) 0 5 10 15 20 ** † * V US RC CPA 200 US RC B lad d e r w e ig h t / b o d y w e ig h t (m g /g ) (A) Num b e r o f n o cicep tiv e be ha v ior f or 30 m in Nocice p tiv e sco re 0.008 g Filament 0.07 g Filament 0 5 10 15 20 0.4 g Filament 1.0 g Filament (C) (B) V US RC US RC 0 0.4 0.8 1.2 1.6 ** †† V US RC CPA 200 US RC (i.p.) (mg/kg) CPA 200 (i.p.) (mg/kg) V US RC CPA 200 US RC V US RC CPA 200 US RC V US RC CPA 200 US RC 2. 低用量シクロホスファミドにより誘起される膀胱炎・膀胱痛に及ぼす RC ストレ スの影響
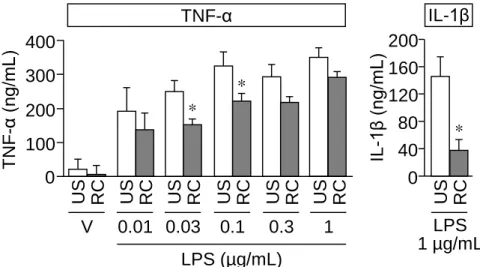
32 RC ストレスによるシクロホスファミド誘起膀胱痛および膀胱炎症状の低下にマク ロファージの機能変化が関与する可能性を検討するため、腹腔マクロファージを採取 し、LPS 刺激後による炎症性サイトカイン TNF-α および IL-1β の遊離量を調べた。そ の結果、RC ストレスマウスの腹腔マクロファージでは、LPS 刺激により分泌される TNF-α および IL-1β 量が、非ストレスマウスに比べて有意に低下していた (Fig. 11) 。
Fig. 11 LPS-induced production of TNF-α and IL-1β in peritoneal macrophages isolated from unstressed (US)
and RC-stressed mice. Peritoneal macrophages were stimulated with LPS at 0.01 , 0.03 , 0.1 , 0.3 or 1 µg/ml or vehicle (V) for 24 h. Data show mean ± SEM for 4-13 (TNF-α) or 4 (IL-1β). *P<0.05 vs US.
III. 考察
本節において、マクロファージ/ミクログリア阻害薬であるミノサイクリンはシクロ ホスファミド誘起膀胱相対湿重量に影響を与えることなく、膀胱痛と関連痛覚過敏を 阻止したことから、本モデルにおける膀胱痛の発現にはマクロファージが重要な役割 を果たすことが明らかとなった。これは、本モデルの膀胱組織でマクロファージの集 積が認められたとの報告75)と一致する。また RC ストレスマウスの腹腔マクロファー ジでは、LPS 刺激に対する TNF-α および IL-1β の分泌反応が低下していたことより、 RC ストレスマウスでは、マクロファージ機能の低下が原因で、シクロホスファミド 0 40 80 120 160 200 LPS 1 µg/mL IL-1β * 0 100 200 300 400 V US RC LPS (µg/mL) TNF-α * * T NF -α (ng/m L ) IL -1 β (ng /m L ) 0.01 US RC 0.03 US RC 0.1 US RC 0.3 US RC 1 US RC US RC 3. 腹腔マクロファージ機能に対する RC ストレスの影響33
により誘起される膀胱痛が、非ストレスマウスほど起こらなかった可能性が示唆され た。今回、ミノサイクリンが、シクロホスファミドにより誘起される膀胱痛を抑制し た一方、湿重量増加を抑制しなかったことは、マクロファージなどから分泌される high mobility group box 1 (HMGB1) やその標的分子である終末糖化産物受容体 (RAGE) が、 同モデルの膀胱痛に関与するが、炎症症状には関与しないとの報告73)と一致する。一 方、RC ストレスによって膀胱痛や膀胱炎症が軽減していたことは、間質性膀胱炎の 患者の膀胱痛がストレスによって増悪するとの臨床報告74)と矛盾する。この RC スト レスマウスにおいてマクロファージ機能の低下が見られたことより、このマウスは易 感染状態にある可能性が考えられ、感染性膀胱炎はむしろ促進される可能性もある。 以上、本節の結果から、RC ストレスマウスでは、シクロホスファミド誘起膀胱炎・ 膀胱痛が、非ストレスマウスよりも軽度であることが判明し、この膀胱痛の減弱には マクロファージの機能低下が関与する可能性が示唆された。
34
総括
本研究では、ヒトにおいて慢性ストレスにより生じる体調変化や関連する疾患を探 るため、1 週間の RC ストレスを負荷したマウスが、新たな急性ストレスや炎症性刺 激に対してどの様な反応を示すかについて検討した。 第 1 章では急性拘束ストレスによって誘発される体温上昇に対する RC ストレスの 影響について検討した。始めに、1 時間の急性拘束ストレスにより誘発される体温上 昇を調べたところ、RC ストレスマウスでは非ストレスマウスに比べて、体温上昇反 応が有意に亢進していた。そこで、この変化に熱産生組織である褐色脂肪組織に発現 するアドレナリンβ3受容体が関与するかを選択的拮抗薬であるSR59230A を用いて検 討した。その結果、RC ストレスマウスにおける拘束ストレス誘発体温上昇の亢進は SR59230A により有意に抑制された。さらに、褐色脂肪組織重量および褐色脂肪組織 のマーカーである uncoupling protein-1 のタンパク発現量を調べたところ、どちらにも わずかな増加傾向が認められた。一方、RC ストレスマウスでは非ストレスマウスと 比較して拘束ストレスによるコルチコステロン分泌も増加していることが明らかとな った。そこで、グルココルチコイド受容体拮抗薬であるミフェプリストンの効果を検 討したところ、RC ストレスマウスにおける拘束ストレス誘発体温上昇反応の亢進へ の影響は認められなかった。また抗不安薬ジアゼパムも RC ストレスマウスにおける 体温上昇反応亢進に影響しなかった。これらの知見より、RC ストレスマウスにおけ る拘束ストレス誘発体温上昇の亢進には、褐色脂肪組織に発現するアドレナリンβ3受 容体の活性上昇が関与し、副腎皮質ホルモンや不安状態の関与は否定的であることが 示唆された。 第 2 章、第 1 節では、マウスの深部体温を、テレメトリーシステムを用いて無麻酔、 無拘束で測定し、LPS 誘起発熱反応に対する RC ストレスの影響を検討した。始めに、 RC ストレス負荷最終日の深部体温を確認したところ、RC ストレスマウスの低温環境 にある夜間の深部体温は非ストレスマウスと比較して有意に低下しており、活動量も 低下傾向にあった。次に LPS を腹腔内投与したところ RC ストレスマウスは 10-30 分 後および 70-140 分後をピークとする 2 峰性の発熱反応が認められ、これは非ストレス マウスと比較して有意に増加していた。この RC ストレスによる LPS 誘起発熱反応の35 増強は COX 阻害薬ジクロフェナクにより抑制された。しかし、肺および視床下部の COX-2 タンパク発現量は、RC ストレスおよび LPS 投与により変化しなかった。また、 RC ストレスマウスおよび非ストレスマウスともに LPS 投与により 90 および 150 分後 に血清コルチコステロン濃度上昇が認められたが、それに対する RC ストレスの影響 は明確ではなかった。これらの知見から、RC ストレスは、LPS 刺激によって COX 依 存性に産生されたプロスタグランジンによる発熱反応を増強することが明らかとなっ た。RCストレスマウスは致死量のLPSに対して抵抗性を示すことが報告されており、 RC ストレスによる発熱反応の増強がこの LPS 耐性獲得に寄与している可能性が考え られる。 第 2 章、第 2 節では、ヒトの間質性膀胱炎に似た症状を示すシクロホスファミド誘 起マウス膀胱炎モデルを用いて、RC ストレスが膀胱炎症および膀胱痛に与える影響 を検討した。RC および非ストレスマウスの腹腔内にシクロホスファミドを大量 (400 mg/kg) 投与すると 3 時間半から 4 時間後に膀胱痛様行動、関連痛覚過敏および膀胱湿 重量増加が認められ、これらの症状の程度に両群間で差は認められなかった。マクロ ファージ/ミクログリア阻害薬ミノサイクリンは、RC および非ストレス群どちらにお いても膀胱湿重量増加に影響することなく、膀胱痛様行動および関連痛覚過敏を強く 抑制した。次に、低用量 (200 mg/kg) のシクロホスファミドを投与したところ、膀胱 痛様行動、膀胱関連痛覚過敏および膀胱湿重量増加は、RC ストレスマウスにおける 方が非ストレスマウスに比較して軽微であった。最後に、腹腔マクロファージにおい て LPS 刺激による TNF-α および IL-1β の分泌量を測定したところ、RC ストレスマウ スのマクロファージでは、非ストレスマウスに比べて、これらサイトカイン分泌量が 低下していた。このことより、RC ストレスマウスでは、マクロファージの機能の低 下が原因で、シクロホスファミドにより誘起される膀胱痛が非ストレスマウスと比較 して軽微であった可能性が示唆された。しかし、マクロファージ機能の低下は感染に 対する防御能低下に繋がる危険性を秘めており、今後感染性膀胱炎に及ぼす RC スト レスの影響についても調べる必要がある。 以上より、RC ストレスマウスではアドレナリン β3受容体が関与する急性拘束スト レス誘起体温上昇反応と、内因性プロスタグランジンに依存する LPS 誘起発熱反応が 上昇していることが明らかとなった。一方、RC ストレスによりマクロファージ機能
36
は低下するため、シクロホスファミド誘起膀胱痛はむしろ減弱することが示された。 本研究により、RC ストレスが体温調節能や内臓知覚に影響を与えることが明らかと なり、臨床において、ストレスが関係する様々な体調変化・異常を予防する上で、ス トレスコントロールを行うことの重要性が改めて裏付けられた。
37
引用文献
1) Swaab DF, Bao AM, Lucassen PJ: The stress system in the human brain in depression and neurodegeneration. Ageing Res Rev, 4, 141-194 (2005).
2) Nakamura K: Central circuitries for body temperature regulation and fever. Am J Physiol Regul Integr Comp Physiol, 301, R1207-1228 (2011).
3) Miller AH, Maletic V, Raison CL: Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol Psychiatry, 65, 732-741 (2009).
4) Oka T, Kanemitsu Y, Sudo N, Hayashi H, Oka K: Psychological stress contributed to the development of low-grade fever in a patient with chronic fatigue syndrome: a case report. Biopsychosoc Med, 7, 7 (2013).
5) Masatoshi T, Yasuko K, Ryoichi N, Yoshishige I, Shigeko T, Nobuyuki N: Time-related differences in noradrenaline turnover in rat brain regions by stress. Pharmacol Biochem Behav, 16, 315-319 (1982).
6) Anisman H, Lacosta S, Kent P, McIntyre DC, Merali Z: Stressor-Induced
Corticotropin-Releasing Hormone, Bombesin, ACTH and Corticosterone Variations in Strains of Mice Differentially Responsive to Stressors. Stress, 2, 209-220 (1998). 7) Henry JP, Stephens PM, Santisteban GA: A model of psychosocial hypertension
showing reversibility and progression of cardiovascular complications. Circ. Res., 36, 156-164 (1975).
8) Zimmerman RS, Frohlich ED: Stress and hypertension. J Hypertens Suppl, 8, S103-107 (1990).
9) Adams N, Lins MD, Blizard Da: Contrasting effects of social stress and foot-shock on acute cardiovascular response in salt-sensitive rats. Behav. Neural Biol., 48, 368-381 (1987).
10) Budd GM, Warhaft N: Body temperature, shivering, blood pressure and heart rate during a standard cold stress in Australia and Antarctica. J Physiol, 186, 216-232 (1966).
38
11) Rao SS, Hatfield RA, Suls JM, Chamberlain MJ: Psychological and physical stress induce differential effects on human colonic motility. Am J Gastroenterol, 93, 985-990 (1998).
12) Esch T, Stefano GB, Fricchione GL, Benson H: The role of stress in neurodegenerative diseases and mental disorders. Neuro Endocrinol Lett, 23, 199-208 (2002).
13) Purdy J: Chronic physical illness: a psychophysiological approach for chronic physical illness. Yale J Biol Med, 86, 15-28 (2013).
14) Kita T, Hata T, Yoneda R, Okage T: [Stress state caused by alteration of rhythms in environmental temperature, and the functional changes in mice and rats]. Nihon Yakurigaku Zasshi, 71, 195-210 (1975).
15) Hata T, Kita T, Oyama, R.Itoh, E.Kawabata, A.Nishimura, Y: On the hyperalgesia in SART-stressed mice with vagotonic-type dysautonomia. Neurosciences., 12, 166-167 (1986).
16) Hata T, Kita T, Namimatsu A, Itoh E, Oda Y: [Changes of blood pressure and regional blood flow in SART rats and drug actions on these changes (author's transl)]. Nihon Yakurigaku Zasshi, 79, 335-342 (1982).
17) Hata T, Itoh E, Funakami Y, Ishida K, Uchida S: Blood pressure and heart rate are increased by AF-DX 116, a selective M2 antagonist, in autonomic imbalanced and hypotensive rats caused by repeated cold stress. Jpn J Pharmacol, 85, 313-321 (2001). 18) Hata T, Funakami Y, Itoh E: An Experimental Animal Model for Orthostatic
Hypotension: Repeatedly Cold-Stressed Rats with Autonomic Imbalance. Jpn J Psychosom Med., 607-706 (2005).
19) Kita T, Hata T, Ito E, Namimatsu A: Testing methods for vegetative syndrome in the rat and effects of Neurotropin and other drugs. Jpn J Psychosom Med., 23, 61-68 (1983). 20) Hata T, Nishikawa H, Itoh E, Watanabe A: Depressive state with anxiety in repeated
cold-stressed mice in forced swimming tests. Jpn. J. Pharmacol., 79, 243-249 (1999). 21) Hata T, Kita T, Kamanaka Y, Honda S, Kakehi K, Kawabata A, Itoh E: Catecholamine
levels in the brain of SART (repeated cold)-stressed rats. J Auton Pharmacol, 7, 257-266 (1987).
39
22) Vinkers CH, Groenink L, van Bogaert MJV, Westphal KGC, Kalkman CJ, van Oorschot R, Oosting RS, Olivier B, Korte SM: Stress-induced hyperthermia and infection-induced fever: Two of a kind? Physiol. Behav., 98, 37-43 (2009).
23) Rey S, Huntingford FA, Boltaña S, Vargas R, Knowles TG, Mackenzie S: Fish can show emotional fever: stress-induced hyperthermia in zebrafish. Proc Biol Sci, 282, 20152266-20152266 (2015).
24) Briese E, De Quijada MG: Colonic temperature of rats during handling. Acta Physiol Lat Am, 20, 97-102 (1970).
25) Olivier B, Zethof T, Pattij T, Van Boogaert M, Van Oorschot R, Leahy C, Oosting R, Bouwknecht A, Veening J, Van Der Gugten J, Groenink L: Stress-induced
hyperthermia and anxiety: Pharmacological validation. Eur. J. Pharmacol., 463, 117-132 (2003).
26) Oka T, Oka K, Kobayashi T, Sugimoto Y, Ichikawa A, Ushikubi F, Narumiya S, Saper CB: Characteristics of thermoregulatory and febrile responses in mice deficient in prostaglandin EP1 and EP3 receptors. J Physiol, 551, 945-954 (2003).
27) Zethof TJJ, Van Der Heyden JAM, Tolboom JTBM, Olivier B: Stress-induced hyperthermia as a putative anxiety model. Eur. J. Pharmacol., 294, 125-135 (1995). 28) Hiramoto T, Oka T, Yoshihara K, Kubo C: Pyrogenic cytokines did not mediate a stress
interview-induced hyperthermic response in a patient with psychogenic fever: a case report. Psychosom Med, 71, 932-936 (2009).
29) Kaneda Y, Tsuji S, Oka T: Age distribution and gender differences in psychogenic fever patients. Biopsychosoc Med, 3, 6-6 (2009).
30) Oka T, Oka K: Age and gender differences of psychogenic fever: a review of the Japanese literature. Biopsychosoc Med, 1, 11-11 (2007).
31) Timmerman RJ, Thompson J, Noordzij HM, van der Meer JW: Psychogenic periodic fever. Neth J Med, 41, 158-160 (1992).
32) McNeil GN, Leighton LH, Elkins AM: Possible psychogenic fever of 103.5 degrees F in a patient with borderline personality disorder. Am J Psychiatry, 141, 896-897 (1984).
40
33) Lkhagvasuren B, Nakamura Y, Oka T, Sudo N, Nakamura K: Social defeat stress induces hyperthermia through activation of thermoregulatory sympathetic premotor neurons in the medullary raphe region. Eur. J. Neurosci., 34, 1442-1452 (2011). 34) Ootsuka Y, Blessing WW, Nalivaiko E: Selective blockade of 5-HT2A receptors
attenuates the increased temperature response in brown adipose tissue to restraint stress in rats. Stress, 11, 125-133 (2008).
35) Gao B, Kikuchi-Utsumi K, Ohinata H, Hashimoto M, Kuroshima A: Repeated immobilization stress increases uncoupling protein 1 expression and activity in Wistar rats. Jpn J Physiol, 53, 205-213 (2003).
36) Hata T, Itoh E, Nishikawa H: Behavioral characteristics of SART-Stressed mice in the forced swim test and drug action. Pharmacol Biochem Behav, 51, 849-853 (1995). 37) Hata T, Kita T, Itoh E, Kawabata A: The relationship of hyperalgesia in SART
(repeated cold)-stressed animals to the autonomic nervous system. J. Auton. Pharmacol.,
8, 45-52 (1988).
38) Kawabata A, Hata T: Attenuation by prolonged nitric oxide synthase inhibition of the enhancement of fibrinolysis caused by environmental stress in the rat. Br J Pharmacol.,
119, 346-350. (1996).
39) Hata T, Itoh E, Kamanaka Y, Kawabata A, Honda S: Plasma catecholamine levels in SART-stressed rats and effects of drugs on stress-induced alteration in plasma and brain catecholamine levels. J. Auton. Pharmacol., 11, 15-25 (1991).
40) Mukae T, Uchida H, Ueda H: Donepezil Reverses Intermittent Stress-Induced
Generalized Chronic Pain Syndrome in Mice. J. Pharmacol. Exp. Ther., 353, 471-479 (2015).
41) Nishiyori M, Ueda H: Prolonged gabapentin analgesia in an experimental mouse model of fibromyalgia. Mol Pain, 4, 52-52 (2008).
42) Zlatković J, Todorović N, Bošković M, Pajović SB, Demajo M, Filipović D: Different susceptibility of prefrontal cortex and hippocampus to oxidative stress following chronic social isolation stress. Mol. Cell. Biochem., 393, 43-57 (2014).
41
43) Morilak DA, Barrera G, Echevarria DJ, Garcia AS, Hernandez A, Ma S, Petre CO: Role of brain norepinephrine in the behavioral response to stress. Prog
Neuropsychopharmacol Biol Psychiatry, 29, 1214-1224 (2005).
44) Kita T, Hata T, Itoh E, Harada N: Methacholine- and adrenaline-induced arrhythmias in repeatedly cold-stressed mice. Jpn J Pharmacol, 35, 327-329 (1984).
45) Schneider Ca, Rasband WS, Eliceiri KW: NIH Image to ImageJ: 25 years of image analysis. Nat Methods, 9, 671-675 (2012).
46) Lazarus M, Yoshida K, Coppari R, Bass CE, Mochizuki T, Lowell BB, Saper CB: EP3 prostaglandin receptors in the median preoptic nucleus are critical for fever responses. Nat. Neurosci., 10, 1131-1133 (2007).
47) Nakamura Y, Nakamura K, Matsumura K, Kobayashi S, Kaneko T, Morrison SF: Direct pyrogenic input from prostaglandin EP3 receptor-expressing preoptic neurons to the dorsomedial hypothalamus. Eur. J. Neurosci., 22, 3137-3146 (2005).
48) Nakamura K, Matsumura K, Kaneko T, Kobayashi S, Katoh H, Negishi M: The rostral raphe pallidus nucleus mediates pyrogenic transmission from the preoptic area. J Neurosci, 22, 4600-4610 (2002).
49) Kataoka N, Hioki H, Kaneko T, Nakamura K: Psychological stress activates a dorsomedial hypothalamus-medullary raphe circuit driving brown adipose tissue thermogenesis and hyperthermia. Cell Metab, 20, 346-358 (2014).
50) Nishikawa H, Hata T, Itoh E, Funakami Y: A role for corticotropin-releasing factor in repeated cold stress-induced anxiety-like behavior during forced swimming and elevated plus-maze tests in mice. Biol Pharm Bull, 27, 352-356 (2004).
51) Hata T, Nishikawa H, Itoh E, Funakami Y: Anxiety-like behavior in elevated plus-maze tests in repeatedly cold-stressed mice. Jpn. J. Pharmacol., 85, 189-196 (2001).
52) Vinkers CH, van Bogaert MJV, Klanker M, Korte SM, Oosting R, Hanania T, Hopkins SC, Olivier B, Groenink L: Translational aspects of pharmacological research into anxiety disorders: The stress-induced hyperthermia (SIH) paradigm. Eur. J. Pharmacol.,