生体膜環境でのアポリポタンパク質
アミロイド形成機構の解明
2018
徳島大学大学院 薬科学教育部
博士後期課程 製剤分子設計学分野
水口智晴
i
目 次
目次... i 略語表...ii 序論...1 本論...6 1 章 アミロイドーシス変異アポ A-I の脂質膜環境でのアミロイド線維形成機構...6 1. 1 水溶液中および脂質膜上でのアポ A-I アミロイド線維形成...6 1. 2 アポ A-I の脂質膜結合挙動...13 1. 3 アポ A-I 線維化領域ペプチドの線維形成性および脂質膜結合性...18 1. 4 部位特異的蛍光プローブ標識による脂質膜上でのアポ A-I 局所環境の解析...22 1. 5 脂質膜結合によるアポ A-I ヘリックス構造の安定化...26 1. 6 小括 ...27 2 章 アミロイドーシス変異アポA-I のアミロイド線維形成に及ぼすホスファチジルセ リンおよびコレステロールの影響...29 2. 1 脂質膜上でのアポ A-I アミロイド線維形成性の変化...29 2. 2 アポ A-I の脂質膜結合性の変化...32 2. 3 アポ A-I 線維化領域ペプチドを用いた線維形成性の評価...36 2. 4 部位特異的蛍光プローブ標識アポ A-I を用いた脂質結合局所構造の評価...40 2. 5 膜蛍光プローブを用いた脂質膜構造変化の評価 ...42 2. 6 小括 ...43 総括...44 実験の部...46 参考文献...55 謝辞...62 公刊論文...63略 語 表
Acrylodan(Ac):6-acryloyl-2-dimethylaminonaphthalene a.u.:arbitrary unit apoA-I:apolipoprotein A-I apoE:apolipoprotein E CD:circular dichroism Chol:cholesterolDLS:dynamic light scattering
DMPC:dimyristoyl phosphatidylcholine
E. coli:Escherichia coli
EDTA:ethylenediaminetetraacetic acid em.:emission
EPR:electron paramagnetic resonance ex.:excitation
Fmoc:9-fluorenylmethyloxycarbonyl GdnHCl:guanidine hydrochloride GP:generalized polarization HDL:high density lipoprotein IAPP:pro-islet amyloid polypeptide
IPTG:isopropyl-β-D(-)-thiogalactopyranoside ITC:isothermal titration calorimetry
Laurdan:6-dodecanoyl-2-dimethylaminonaphhtalene LDL:low density lipoprotein
LDLR:low density lipoprotein receptor NAC:non amyloid- component PC:phosphatidylcholine
PL:phospholipid
Prodan:6-propionyl-2-(dimethylamino)naphthalene PS:phosphatidylserine
SAA:serum amyloid A
SDS-PAGE:sodium dodecyl sulfate-polyacrylamide gel electrophoresis SUV:small unilamellar vesicle
T2D:type 2 diabetes
iii TEM:transmission electron microscopy
ThT:thioflavin T
TIRFM:total internal reflection fluorescence microscopy Tris:2-amino-2-hydroxymethyl-1,3-propanediol
Trx:thioredoxin TTR:transthyretin
VLDL:very low density lipoprotein
序 論
アミロイドーシス(アミロイド病)は、原因となる前駆タンパク質が凝集し、分解抵抗性のアミ ロイド線維として全身の様々な組織や臓器に沈着して機能不全を引き起こす疾患の総称である (Figure I)1。現在までに、アルツハイマー病や透析アミロイドーシス、パーキンソン病など多くの 疾患が報告されている 2-5。それぞれ原因となるタンパク質が異なり、アルツハイマー病ではアミロ イド、パーキンソン病では-シヌクレインなど、40 種類を超えるタンパク質がアミロイドーシスを 引き起こすことが知られている(Table I)。中には、疾患とは直接関係のないタンパク質やペプチド でも、条件によりアミロイド線維を形成し得ることが分かっている4,6,7。例えば、長期透析患者に見 られる透析アミロイドーシスは2 ミクログロブリンの蓄積によって発症し、これは血液透析という 医療がもたらした現代病とも言える。つまり、本来疾患とは関係がなかったはずのタンパク質が、 現代アミロイドーシスとして問題視されていると考えられ、アミロイドーシスは高齢化・高度医療 社会において、特に深刻な疾患である。 Figure I アミロイドーシスの概略図2
Table I 代表的なアミロイドーシスとその前駆タンパク質2-5
臨床病名 前駆タンパク質
Alzheimer's disease amyloid precursor protein (APP),
presenilin-1 (PS-1), presenilin-2 (PS-2)
Parkinson's disease -synuclein
Frontotemporal lobar degeneration (FTLD)-tau tau
Creutsfeldt-Jacob Disease prion protein
Transthyretin familial amyloidosis transthyretin (TTR)
Hereditary lysozyme amyloidosis lysozyme
Hereditary fibrinogen -chain amyloidosis fibrinogen -chain
Hereditary ApoAI/II amyloidoses apolipoprotein A-I, apolipoprotein A-II
Secondary systemic amyloidosis serum amyloid A (SAA)
Hemodislysis-related amyloidosis 2-microglobulin
Finnish hereditary amyloidosis gelsolin
Type-II diabetes islet amyloid polypeptide (IAPP)
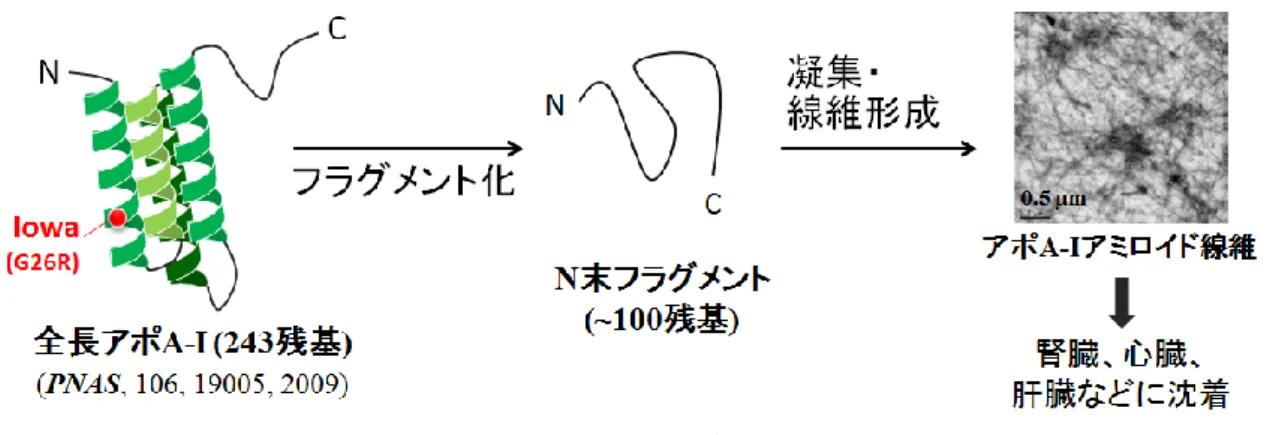
アポリポタンパク質A-I(アポ A-I)は、善玉コレステロールとして知られる高密度リポタンパク 質(HDL)の主要構成タンパク質として循環血中の HDL 粒子の産生、成熟、取込などを制御し、コ レステロール恒常性維持を担う 8,9。これらの機能は、アポ A-I が柔軟に構造変化をすることで制御 されており、アポ A-I は、その柔軟で不安定な構造ゆえ、遺伝子変異や酸化修飾などによりアミロ イド化を起こしやすいことが知られている10-12。アポA-I は全長 243 残基のアミノ酸からなり、両親 媒性ヘリックスがヘリックスバンドル構造をとるN 末端領域(1-187 残基)と、ランダムコイル構造 をとるC 末端領域の二ドメイン構造をとる13-15。現在までに40 種類を超える遺伝子変異が報告され ており、それらは低HDL 血症や家族性アミロイドーシスの発症に関与する16。
アポA-I のアミロイドーシス変異は N 末側領域(1-90 残基、154-178 残基)に集中し(Figure II)
17、組織沈着アミロイド線維からは80-100 残基からなる N 末端フラグメントが検出されている18-21。
アポA-I アミロイドーシスは、Figure III に示すように、全長アポ A-I が遺伝子変異などの影響によ
りN 末端側約 100 残基にフラグメント化され、形成された不安定な N 末フラグメントが凝集し、
シート構造に富んだアミロイド線維を形成するという機序で発症すると考えられている22。
アポ A-I アミロイドーシスに関しては、組織沈着アミロイド線維として検出されたアポ A-I 1-93 フラグメントについてタンパク質分子レベルでの線維形成メカニズムに関する研究がなされていた 23。2011 年には Bellotti らのグループにより、アポ A-I 1-93 フラグメントは生理的中性環境下で線維 形成性を示さないが、酸性環境下においては線維形成性を示し、さらに L90P や70-72 など特定の アミロイドーシス変異の存在によりその線維形成が促進されることが報告された24。 一方で我々のグループは、同じく組織沈着アミロイド線維として検出されたアポ A-I 1-83 フラグ メントに着目し25,26、アポA-I 1-83 フラグメントが生理的中性環境下においても線維形成性を示すこ とを報告した27。 アポA-I 1-83 フラグメントは、N 末領域に存在する代表的な遺伝子変異の 1 つであり、26 番目の グリシン(G)残基が塩基性アミノ酸であるアルギニン(R)残基に置換された Iowa(G26R)変異 体としても検出されている25,26,28。そこで、Iowa(G26R)変異がアポ A-I 1-83 フラグメントの構造 および線維形成性に与える影響について評価を行った。その結果、Iowa(G26R)変異により、全長 アポA-I の N 末ヘリックスバンドル構造が不安定化され両親媒性ヘリックスの疎水面が水層へ露出 されることが明らかとなった27。これが生体内での N 末フラグメントの形成に関与すると考えられ る。 さらにIowa(G26R)変異は、アポ A-I 1-83 フラグメントの生理的中性環境下でのアミロイド線維 化を促進した。26 番目のグリシン残基を、アルギニン残基と同じ塩基性アミノ酸であるリジン(K) 残基に置換しても大きな線維化促進効果が見られなかったこと、また、反対に酸性アミノ酸である グルタミン酸(E)残基に置換すると線維形成が抑制されたことから、アポ A-I 1-83 フラグメントの 線維化促進効果はIowa(G26R)変異に特異的であることが示された27。 Iowa(G26R)変異による線維化促進効果の要因として、変異によるシート構造の伸長があげら れる。グリシン残基はストランド形成性が低いため29,30、26 番目がグリシン残基であるアポ A-I 1-83 フラグメントは、線維形成性(シート形成性)の高い領域が N 末端側の 14-22 残基領域、および中 央部の 46-59 残基領域までであると考えられている 24,31。しかし、Iowa(G26R)変異体では、実際
に電子常磁性共鳴(electron paramagnetic resonance;EPR)実験により、2 ヶ所の線維形成領域が 14-31
残基領域、41-59 残基領域までと、それぞれ伸長することが示されている(Figure IV)32。
4 一方で、生体内環境においてどのようにアポ A-I アミロイド線維が形成されるかについては未解 明である。アポ A-I アミロイドーシスに限らず、アミロイド沈着物からは細胞膜脂質成分やアポリ ポタンパク質E などの共沈着が報告されていることから33,34、細胞膜上において線維が形成されるこ とが予想される。実際に、他のアミロイドタンパク質において、細胞膜などの脂質膜環境がアミロ イド線維形成を促進するとの報告が多数なされている35-37。
例えば、islet amyloid polypeptide(IAPP、アミリン)は、負電荷リン脂質を含有する脂質膜上にお
いて線維形成が促進される38-40。IAPP は膵細胞からインスリンと共分泌される 37 残基のペプチド ホルモンであり、摂食抑制作用や胃内容物排出遅延作用、グルカゴン分泌抑制作用を担うが、Ⅱ型 糖尿病(T2D)患者では肝臓にアミロイドとして蓄積し、肝障害を起こすことがある。負電荷リン 脂質による線維化促進は、N 末端側に存在する正電荷アミノ酸が、負電荷を帯びた脂質膜に結合し て局所的に濃縮され、露出したC 末側領域どうしが相互作用することで凝集・線維形成が促進され るためであると言われている(Figure V)41。高グルコース状態により膵島細胞膜中の負電荷リン脂 質含有量が増大することが示されているため 42、負電荷リン脂質による IAPP の線維化促進は T2D と関連して起こり得る。 Figure IV アポ A-I 1-83 フラグメントの線維形成領域 14 22 46 59 14 31 41 59
アポA-I 1-83/G26R
アポA-I 1-83Iowa (G26R)
Figure V アミロイドタンパク質の脂質膜上での線維形成機序 (参考文献35 より転載)また、-シヌクレインも負電荷リン脂質を含有する脂質膜上において線維形成が促進される 43-45。 -シヌクレインは神経細胞に局在しており、正確な機能は分かっていないが、シナプスの可塑性や 神経伝達物質の調整などを担うことが示唆されている。その異常蓄積によりパーキンソン病や多系 統萎縮症、レビー小体型認知症を発症することが知られている。-シヌクレインは 140 残基からな るタンパク質で、N 末端側(7-87 残基領域)には両親媒性ヘリックス形成領域、中央部(61-95 残基 領域、non amyloid- component;NAC)は凝集性の高い領域、C 末端側(104-140 残基領域)には 14
個の負電荷アミノ酸を含む酸性度の高い領域が存在する。C 末端側の 120-140 残基領域は、負電荷リ ン脂質を含有する脂質膜よりも、中性リン脂質のみからなる脂質膜に対して親和性が高いことが分 かっている45。そのため、中性脂質膜上では脂質膜に結合したC 末端側領域同士の静電的反発(タ ンパク質-タンパク質間の反発)により、NAC 領域の凝集が阻害され、線維化を起こしにくいと考 えられる。一方、負電荷リン脂質を含有する脂質膜上においては、脂質-タンパク質間の静電的反 発によりC 末端側領域の脂質結合が阻害され、脂質膜上におけるタンパク質-タンパク質間の反発 が抑えられるため、NAC 領域の凝集と、それに続く線維化が促進されると考えられる。パーキンソ ン病患者の脳では、負電荷リン脂質含有量が増大しているとの報告もあることから 46、負電荷リン 脂質による-シヌクレインの線維化促進は、パーキンソン病などの病態解明のために重要な知見で あると言える。 本研究では、これらの知見に基づき、アポ A-I N 末フラグメントのアミロイド線維形成における 脂質膜環境の影響について(1 章)47、さらに、構成脂質組成の影響について(2 章)48、主に物理化 学的手法により評価を行った。これにより、実際の生体内環境に近い不均一系でのアポ A-I アミロ イド線維形成機構の解明と、新たなアミロイドーシス制御・治療法開発の基盤となる科学的知見の 構築を目的としている。
6
本 論
1 章 アミロイドーシス変異アポ A-I の脂質膜環境でのアミロイド線維形成機構
1. 1 水溶液中および脂質膜上でのアポ A-I アミロイド線維形成
本研究において使用したアポA-I は大腸菌発現系により作製した(実験の部参照)27。
アポA-I N 末 1-83 フラグメント(アポ A-I 1-83)および Iowa(G26R)変異型 N 末フラグメント(ア
ポA-I 1-83/G26R)を水溶液中(pH 7.4、37℃)でインキュベートし、アミロイド線維に特異的に結
合する蛍光色素チオフラビンT(ThT)の蛍光強度を経時的に測定することにより、線維形成性の評
価を行った49,50。Figure 1-1A は、アポ A-I 1-83/G26R の ThT 蛍光スペクトルの経時変化を示しており、 時間依存的な蛍光強度の増大からアミロイド線維の形成を示す。Figure 1-1B には水溶液中における アポA-I 1-83 およびアポ A-I 1-83/G26R の 485 nm における ThT 蛍光強度の経時変化を示す。このグ ラフから、水溶液中においてはアポA-I 1-83、アポ A-I 1-83/G26R のいずれも強い線維形成性を有す ることが分かる。さらに、アポA-I 1-83 と比較してアポ A-I 1-83/G26R の ThT 蛍光の上昇が早いこ とから、Iowa 変異はアポ A-I 1-83 フラグメントの線維形成を促進することが示された。 アミロイド線維形成機構を説明するモデルとして、重合核依存性重合モデルが提唱されている51,52。 このモデルは、前駆タンパク質が重合してアミロイド核(オリゴマー)を形成した後、重合核ある
Figure 1-1 (A) ThT 蛍光スペクトルと (B) 蛍光強度の経時変化. Fibril formation was monitored
by ThT fluorescence for apoA-I 1–83 and 1–83/G26R incubated at pH 7.4 in solution. The protein concentration was 0.1 mg/ml.(参考文献 47 より一部変更して転載. © the American Society for Biochemistry and Molecular Biology.)
spectra 460 485 510 535 560 585 0 50 100 150 200 0 3 6 9 12 24 48 72 96 120 1-83/G26R: 0.1 mg/ml Wavelength (nm) F lu o re s c e n c e i n te n s it y ( a .u .) solution 0 20 40 60 80 100 120 0 100 200 300 400 1-83 1-83/G26R 1-83 1-83/G26R Time (h) F lu o re s c e n c e i n te n s it y a t 4 8 5 n m ( a .u .) 0 - 120 h
A B
いは線維端に前駆タンパク質が次々に結合し、線維が伸長するというものである(Figure 1-2)53。 ThT 蛍光測定より得られたアミロイド線維形成の経時変化はシグモイド型の曲線を示しており、重 合核依存性重合モデルに従うとき、ThT 蛍光が上昇するまでの保持時間(lag time)は核形成過程の 起こりやすさを反映する54。Figure 1-1B より、アポ A-I N 末フラグメントについても、ThT 蛍光強 度の経時変化がシグモイド型の曲線を示しており、重合核依存性重合モデルに従う線維形成挙動を とると言える。そこで、アポA-I 1-83 とアポ A-I 1-83/G26R の ThT 蛍光経時変化を式[2](実験の部
参照)に従ってフィッティングし、lag time を算出した(Table 1-1)55。アポA-I 1-83/G26R はアポ A-I 1-83 と比較して lag time が短縮したことから、Iowa(G26R)変異による核形成過程の促進が示
唆された27。また、線維の伸長過程を反映する伸長速度定数 k について、アポ A-I 1-83/G26R ではア ポA-I 1-83 と比較して k の増大を示すことから、Iowa 変異は核形成過程だけでなく、伸長過程も促 進することが分かる。 ところで、26 番目のグリシン残基をアルギニン残基と同じ塩基性アミノ酸であるリジン残基に置 換したところ、Iowa(G26R)変異ほどの大きな線維化促進効果は見られず、また、反対に酸性アミ ノ酸であるグルタミン酸(E)残基に置換すると線維形成が抑制されるという結果が得られた27。ア
ポA-I の 26 残基目周辺のアミノ酸配列は、KDSGRDY(アポ A-I 23-29 残基領域)であり、この領域
がシート構造を形成する時、シートの片側の面にて 26 番目のグリシン(G)残基と隣接するアミ ノ酸は、酸性アミノ酸であるアスパラギン酸(D)残基となる。このことから、26 番目を塩基性ア ミノ酸に置換したアポA-I 1-83/G26R およびアポ A-I 1-83/G26K では隣接するアスパラギン酸残基と の静電的相互作用によりシート構造形成が促進され、一方、酸性アミノ酸に置換したアポ A-I 1-83/G26E では静電的反発によりシート構造形成が阻害されるのではないかと考えられる。さらに、 リジン残基よりアルギニン残基への置換の方が大きい線維化促進効果を示すことから、アポA-I 1-83 フラグメントの線維化促進効果はIowa(G26R)変異に特異的なものであると言える。
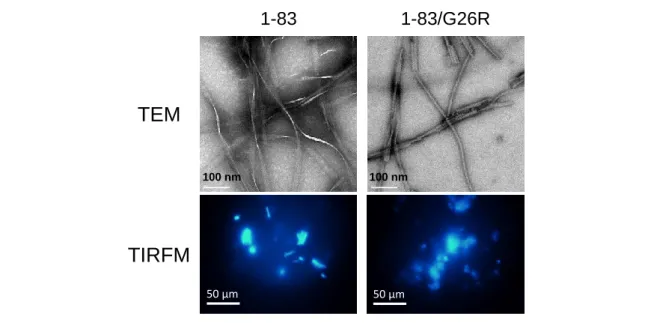
8 Table 1-1 アポ A-I N 末フラグメントの ThT パラメータ k (h-1) lag time (h) 1-83 0.070 ± 0.02 30.7 ± 11.7 1-83/G26R 0.357 ± 0.18 5.5 ± 3.2 Figure 1-3 に示すように、インキュベート後について透過型電子顕微鏡(TEM)および ThT 蛍光を 利用した全反射蛍光顕微鏡(TIRFM)観察を行ったところ、アポ A-I 1-83、アポ A-I 1-83/G26R のい ずれも線維状の凝集物の形成が確認できた。TEM 画像から、アポ A-I N 末フラグメントにより形成 されたアミロイド線維は、太さ約10 nm、長さ数百 nm であり、一般的に報告されるアミロイド線維 の形態と一致する。しかし、形成された線維を詳細に観察すると、アポA-I 1-83 ではねじれの多い 形態が、アポA-I 1-83/G26R ではねじれが少なく真っ直ぐな形態が観察され、Iowa(G26R)変異が 線維形態に影響を及ぼしている可能性も考えられる。 Figure 1-2 アミロイド線維形成の経時変化モデル (参考論文53 より一部変更して転載)
続いて、脂質膜上での線維形成性について評価した。脂質膜として卵黄由来ホスファチジルコリ ン(PC)により作製した small unilamellar vesicle(SUV;粒子径~ 30 nm)を用い55,56、脂質膜上にお けるアポA-I 1-83 およびアポ A-I 1-83/G26R の線維形成性について評価した。Figure 1-4 は SUV 存在
下におけるThT 蛍光強度の経時変化を示す。このグラフから、アポ A-I 1-83 は ThT 蛍光の上昇が見
られず、SUV 存在下では著しく線維形成が阻害されることが示された。一方、アポ A-I 1-83/G26R
は水溶液中と同様にThT 蛍光が上昇し、Iowa(G26R)変異体は SUV 上での線維形成性を有するこ
とが示された。わずか一残基の違いにより、一方ではSUV 上での線維形成が阻害され、もう一方で
は線維形成性を示すという、非常に興味深い結果となった。
Figure 1-3 水溶液中で形成されたアミロイド線維の形態観察. TEM and TIRFM
images of apoA-I 1–83 variants after 120 h incubation at pH 7.4 in solution. Scale bars of TEM and TIRFM represent 100 nm and 50 m, respectively.(参考論文 47 より転載. © the American Society for Biochemistry and Molecular Biology.)
TIRFM
TEM
1-83
100 nm 100 nm1-83/G26R
50 µm 50 µm10
インキュベート後についてTEM および TIRFM 観察を行ったところ、SUV 上において ThT 蛍光が
上昇したアポA-I 1-83/G26R のみ線維状の凝集物が確認できた(Figure 1-5)。SUV 存在下でインキュ
ベートしたアポA-I 1-83/G26R の TEM 画像において見られる球状のものは SUV の凝集物と考えられ、
粒子径が小さく不安定なSUV がインキュベート中に融合しているものと見られる。しかし、同様に
SUV の融合が見られるアポ A-I 1-83 では線維形成に至らないことから、SUV の融合は線維形成性に は影響を与えないと考えられる。
Figure 1-4 脂質膜上におけるアポ A-I 1-83 フラグメントの ThT 蛍光経時変化. Fibril
formation was monitored by ThT fluorescence for apoA-I 1–83 and 1–83/G26R incubated at pH 7.4 in the presence of egg PC SUV. The protein concentration was 0.1 mg/ml. The PC/apoA-I weight ratio was 30.(参考文献 47 より一部変更して転載. © the American Society for Biochemistry and Molecular Biology.)
EPC SUV 0 20 40 60 80 100 120 0 100 200 300 400 1-83 on EPC SUV 1-83/G26R on EPC SUV 1-83 1-83/G26R Time (h) F lu o re s c e n c e i n te n s it y a t 4 8 5 n m ( a .u .)
Figure 1-5 脂質膜上で形成されたアミロイド線維の形態観察. TEM and TIRFM images of apoA-I 1–
83 variants after 120 h incubation at pH 7.4 in the presence of egg PC SUV. The protein concentration was 0.1 mg/ml. The PC/apoA-I weight ratio was 30. Scale bars of TEM and TIRFM represent 100 nm and 50 m, respectively.(参考論文 47 より転載. © the American Society for Biochemistry and Molecular Biology. )
続いて、アポA-I N 末フラグメントと脂質の濃度比の影響について評価した(Figure 1-6A、B)。 アポA-I N 末フラグメント 100 g/mL に対し、SUV PC 濃度を 1.0 mg/mL(PC/アポ A-I = 10(w/w)) もしくは3.0 mg/mL(PC/アポ A-I = 30(w/w))として ThT 蛍光測定を行ったところ、アポ A-I 1-83
は脂質濃度依存的に線維形成が阻害された。これはSUV の存在がアポ A-I 1-83 の線維形成を阻害し
ていることを示す。一方でアポA-I 1-83/G26R は PC/アポ A-I = 10(w/w)の場合において ThT 蛍光
強度が高く、PC/アポ A-I = 30(w/w)の場合よりも線維化しやすいことが分かった。しかし lag time については脂質濃度依存的に延長しており、核形成過程が起こりにくくなっていることが分かる。 この点については、脂質膜上での二次構造が影響していると考えられるため、1. 4 にて詳細に述べる。 アミロイドタンパク質は脂質膜に結合すると、水溶液中(三次元)から脂質膜上(二次元)に濃 縮されることにより、タンパク質間の相互作用が起こりやすくなるため、線維形成が促進されると 言われている(Figure 1-7)58。しかし、アミロイドタンパク質の1 つである-シヌクレインについて 報告があるように 59、脂質膜上での線維形成には最適な脂質/タンパク質濃度比が存在することが示 唆される。つまり、Figure 1-6 に見られるようなアポ A-I 1-83/G26R の線維形成性の違いは、PC/アポ A-I 濃度比が低いとき(= 10(w/w))、SUV 粒子上のアポ A-I の局所濃度が高くなるため、アポ A-I の凝集と線維化が促進されるが、一方、PC/アポ A-I 濃度比が高いとき(= 30(w/w))は、SUV 粒子
上のアポA-I の局所濃度が低くなり、また、線維伸長に必要な水溶液中からのアポ A-I モノマーの供
給が低下するため、線維化が起こりにくくなっていると考えられる。この考察が妥当であるかを判
断するためには、アポA-I 1-83/G26R が水溶液中で線維形成を示さないような条件において、SUV PC
濃度比を変えて線維形成性を評価する必要があると考えた。
Figure 1-6 アポ A-I N 末フラグメントの線維形成性に対する脂質濃度の影響. Fibril
formation was monitored by ThT fluorescence for (A) apoA-I 1-83 or (B) apoA-I 1–83/G26R incubated at pH 7.4 in the absence or presence of egg PC SUV. The protein concentration was 0.1 mg/ml. The PC/apoA-I weight ratio was 10 or 30.(参考論文 47 より一部変更して転載. © the American Society for Biochemistry and Molecular Biology. )
1-83 (0,10,30) 0 20 40 60 80 100 120 0 100 200 300 400 10 30 PC / apoA-I (w/w) = 0 Time (h) F lu o re s c e n c e i n te n s it y a t 4 8 5 n m ( a .u .)
1-83/G26R (0,10,30)
0 20 40 60 80 100 120 0 100 200 300 400 500 10 30 PC / apoA-I (w/w) = 0 Time (h) F lu o re s c e n c e i n te n s it y a t 4 8 5 n m ( a .u .)A
1-83B
1-83/G26R12
そこで、アポA-I 1-83/G26R 濃度 50 g/mL において、様々な SUV PC 濃度で ThT 蛍光測定を行っ た際の結果をFigure 1-8 に示す。アポ A-I 1-83/G26R 濃度 50 g/mL は、水溶液中(PC/アポ A-I = 0 (w/w))、つまり三次元では、ThT 蛍光強度が低く、線維形成を示しにくい濃度であると言える。し かし、PC/アポ A-I 濃度比を上げると(PC/アポ A-I≦10(w/w))、ThT 蛍光強度が PC 濃度依存的に 上昇した。これは、水溶液中という三次元ではアポ A-I 分子間の衝突とそれに続く線維の核形成が 起こりにくいが、脂質膜上という二次元にアポA-I が濃縮されることにより、アポ A-I 分子同士の衝 突頻度が上がり、線維化が促進されたものと考えられる。しかし、さらにPC/アポ A-I 濃度比を上げ ると(PC/アポ A-I>10(w/w))、ThT 蛍光強度が低下していき、脂質膜上においても線維を形成し にくくなることが確認された。これは、上記 Figure 1-6B の結果と同様に、次の通りに説明される。
SUV PC 濃度を上昇させ SUV 粒子数が増大することで、各 SUV にアポ A-I が分散して結合、つまり、 SUV 1 粒子あたりのアポ A-I の局所濃度が SUV PC 濃度依存的に低下していく。また、アポ A-I の SUV への結合量が増大するため、水溶液中に存在するアポ A-I モノマー量が低下し、線維伸長に必
要な水溶液中からのアポA-I モノマーの供給が低下する。これらにより、PC/アポ A-I 濃度比が高い
ときは、アポA-I 1-83/G26R であっても脂質膜上での線維化が起こりにくくなるものと考えられる59。
Figure 1-7 脂質膜上におけるアミロイドタンパク質の二次元濃縮のモデル.
(A) Distribution of molecules in a 3D space. (B) Distribution of molecules in the 2D space of the membrane surface.(参考論文 58 より一部変更して転載. © EBSA 2007)
1. 2 アポ A-I の脂質膜結合挙動
SUV 上でアポ A-I 1-83 フラグメントの線維形成性の違いをもたらす要因について考察するにあた り、全長アポ A-I は脂質膜上で-ヘリックス構造に富んだ構造へと変化することが知られているた め、SUV 結合時のアポ A-I の二次構造変化および結合性の評価を行った。二次構造変化を円偏光二 色性(CD)測定より、SUV への結合性を等温滴定型カロリメトリー(ITC)測定により評価した。 まず、アポA-I の結合が SUV の形態に影響を与えないことを動的光散乱(DLS)測定により確認し た(Figure 1-9)。Figure 1-8 PC/アポ A-I 濃度比と線維形成性の関係. Effects of the PC to apoA-I
ratio on ThT fluorescence intensities for apoA-I 1–83/G26R incubated in the presence of egg PC SUV. The protein concentration was 0.05 mg/ml.(参考論文 47 より一部変 更して転載. © the American Society for Biochemistry and Molecular Biology.)
Data 1 0 10 20 30 40 50 60 70 80 90 100 0 50 100 150 200 250 SUV PC/apoA-I (w/w) M a x im a l T h T f lu o re s c e n c e ( a .u .)
Figure 1-9 SUV の粒子径測定. Particle size distributions of PC SUV determined by dynamic
light scattering measurements on a Zetasizer Nano ZS (Malvern). The data were represented as volume-based distributions.(参考論文 58 より転載) EPC 0.1 1 10 100 1000 10000 0 5 10 15 20 PC SUV PC SUV + apoA-I 1-83/G26R Hydrodynamic diameter (nm) D is tr ib u ti o n ( % )
14
Figure 1-10A にはアポ A-I 1-83/G26R に対し SUV PC 濃度を上昇させた際の CD スペクトルの変化
を示す。アポA-I N 末 1-83 フラグメントは水溶液中では 198 nm に負の極大を持つランダムコイル構
造に特徴的なスペクトルを与え、SUV に結合すると 208 および 222 nm に負の極大を持つスペクトル
を示し、-ヘリックス構造へと構造変化することが分かる。222 nm の平均残基モル楕円率からヘ
リックス含量を算出し(実験の部参照)、SUV 結合時のアポ A-I 1-83 とアポ A-I 1-83/G26R のヘリッ
クス含量を比較した(Figure 1-10B)。その結果、アポ A-I 1-83 と比較して、アポ A-I 1-83/G26R は SUV
結合時のヘリックス含量が低下することが示された。なぜIowa(G26R)変異が、Figure 1-10B に示
す大きなヘリックス形成性の低下をもたらすのかについて考察するため、Iowa 変異周辺がヘリック ス構造を形成した際の各残基の位置関係と、Iowa 変異による影響について、ヘリカルホイール図を 描くことにより調べた。
CD spectra of apoA-I 1-83/G26R + SUV
190 200 210 220 230 240 250 260 -15000 -10000 -5000 0 5000 10000 15000 SUV PC/apoA-I 0 (w/w) 5 10 20 30 SUV PC/apoA-I = 0-30 w/w Wavelength (nm) M o la r e ll ip ti c it y ( d e g c m 2 /d m o le )
A B
Data 9 0 5 10 15 20 25 30 10 20 30 40 1-83 1-83/G26R 1-83 1-83/G26R SUV PC/apoA-I (w/w) -h e li x c o n te n t (% )Figure 1-10 (A) CD スペクトル、(B) ヘリックス含量. A, far-UV CD spectra of apoA-I 1–83/G26R
bound to egg PC SUV. The protein concentration was 50 g/ml. B, increases in -helix content of apoA-I 1–83 (●) and 1–83/G26R (△) as a function of the weight ratio of PC to apoA-I.(参考文献 47 より転載. © the American Society for Biochemistry and Molecular Biology.)
Figure 1-11 はアポ A-I 16-33 残基領域のヘリカルホイール図である。このようにアポ A-I 16-33 残 基領域は、塩基性アミノ酸であるリジン(K)残基およびアルギニン(R)残基、酸性アミノ酸であ るアスパラギン酸(D)残基といった電荷を有するアミノ酸に富んだ親水面と、疎水性アミノ酸に富 んだ疎水面を有する、両親媒性のヘリックス構造を形成することが分かる。アポ A-I の脂質膜への 結合は、両親媒性ヘリックスの疎水面を脂質膜の疎水性アシル鎖領域に埋め込むことによって起こ る13,60。26 番目のグリシン(G)残基はヘリックスの疎水面に位置しているため、Figure 1-11 の通り、 ここが電荷を有するアルギニン(R)残基に置換されることで疎水面が縮小して両親媒性が低下し、 これがFigure 1-10B に示すような Iowa(G26R)変異によるヘリックス形成性の低下につながると考 えられる27,61。
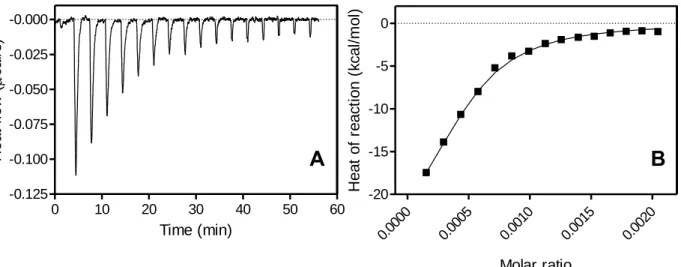
続いて、アポA-I と SUV との相互作用様式について詳細に調べるため、ITC 測定を行った。Figure
1-12A には、ITC 測定で SUV に対しアポ A-I 1-83/G26R を滴下した際のサーモグラムを示す。SUV
が過剰に存在する場合にはアポ A-I 1-83/G26R が SUV に結合して発熱反応が起こるが、アポ A-I
1-83/G26R が飽和すると結合熱の発生が見られなくなる。このサーモグラムから Figure 1-12B に示す 結合等温線を得、フィッティングを行うことにより熱力学的パラメータを得た。SUV 結合時の ITC
測定をアポA-I 1-83 についても行い、得られた熱力学的パラメータを Table 1-2 にまとめた。その結
果、アポA-I 1-83 と比較してアポ A-I 1-83/G26R では SUV に対する結合親和性が増大し、最大結合
量(Bmax)および負の結合エンタルピー(H)値が低下することが分かった。負のエンタルピー変
化は、主にヘリックス中の水素結合形成によってもたらされると考えられている 56。そのため Iowa
変異による負のH 値の低下は、CD 測定において Iowa 変異により SUV 結合時のヘリックス含量が 低下していた結果と一致する。
16
1-83/G26R (0.02 mM) + SUV (0.3 mM)
0.00 00 0.00 05 0.00 10 0.00 15 0.00 20 -20 -15 -10 -5 0 Molar ratio He a t o f re a c ti o n ( k c a l/ m o l) Kd (μg/ml) Bmax (amino acids/mol PC) Ga (kcal/mol) H (kcal/mol) TSb (kcal/mol) 1-83 3.8 ± 0.3 0.34 ± 0.01 -11.1 ± 0.1 -23.7 ± 1.9 -12.6 ± 1.9 1-83/G26R 1.7 ± 0.1 0.15 ± 0.01 -11.6 ± 0.1 -20.3 ± 0.8 -8.7 ± 0.8Figure 1-6 において、アポ A-I 1-83 は SUV PC 濃度依存的に線維形成が阻害され、一方アポ A-I 1-83/G26R は SUV PC 濃度を増大させても線維形成性を維持するという結果が得られていた。ここで、
ITC 測定の結果から Iowa(G26R)変異による Bmaxの低下が示されたことにより、アポA-I 1-83/G26R
における脂質膜上での線維形成性の維持は、アポA-I 1-83 と比較して脂質膜への結合量が低下し、
水溶液中に多くのアポA-I 1-83/G26R モノマーが残存するために起こる可能性が考えられた。そこで、
線維形成を評価した濃度条件における、アポA-I の SUV への結合率の算出を試みた。
Figure 1-12 (A) ITC サーモグラム、(B) 結合等温線. A, isothermal titration thermogram for
binding of apoA-I 1–83/G26R to egg PC SUV. B, binding isotherm calculated from A.
A B
1-83/G26R (0.02 mM) + SUV (0.3 mM)
0 10 20 30 40 50 60 -0.125 -0.100 -0.075 -0.050 -0.025 -0.000 Time (min) He a t fl o w ( c a l/ s )a Free energy was calculated according to G = -RT ln 55.5(1/K d). b The entropy of binding was calculated from G = H - TS
Table 1-2 アポ A-I N 末フラグメントの SUV 結合における熱力学的パラメータ. Thermodynamic
parameters of binding of apoA-I variants to egg PC SUV at 25℃.(参考文献 47 より転載. © the American Society for Biochemistry and Molecular Biology.)
Table 1-2 に記載した解離定数(Kd)およびBmaxを用い、異なるPC/アポ A-I 濃度比において SUV
に結合したアポA-I の割合、および、SUV 粒子上に結合したアポ A-I 分子数を算出し Figure 1-13 に
示した(実験の部参照)62。
Figure 1-13 にて算出した SUV への結合率をもとに、Figure 1-6 に示すアポ A-I 1-83 とアポ A-I 1-83/G26R の脂質濃度による線維形成性の違いについて考察する。
アポ A-I 1-83 は、PC/アポ A-I 濃度比が高いとき(>30(w/w))線維形成性を示さず、このとき
Figure 1-13(●)から、100 %近くのアポ A-I 1-83 が SUV に結合した状態であったことが分かる。そ
してPC/アポ A-I 濃度比が低いとき(≦10(w/w))には線維形成を示したが、SUV 表面はアポ A-I 1-83
で飽和しており(Figure 1-13(○))、水溶液中には SUV に結合していないアポ A-I 1-83 が 40 %以上 存在することが分かる(Figure 1-13(●))。つまり、PC/アポ A-I 濃度比が低いとき(≦10(w/w)) の線維化は、SUV に結合していないアポ A-I 1-83 によるものであり、SUV 結合状態のアポ A-I 1-83 に線維形成能はないと考えられる。
一方アポA-I 1-83/G26R は、PC/アポ A-I 濃度比が高いとき(>30(w/w))にも線維形成性を維持
した。このとき、100 %近くのアポ A-I 1-83/G26R が SUV に結合していることから(Figure 1-13(▲))、 SUV 結合状態のアポ A-I 1-83/G26R も線維形成能を有することが分かる。すなわち、Iowa 変異体の
みが脂質膜上で線維を形成したことは、脂質結合性の違いによるものではなく、アポ A-I の脂質結
合状態の違いによるものであると言える。
Figure 1-13 アポ A-I N 末フラグメントの SUV への結合性の比較. Changes in the fraction % of
apoA-I bound to SUV (●and▲) and molar ratio of bound apoA-I to PC on the SUV surface (○and△) for apoA-I 1–83 (dashed line) and 1–83/G26R (dotted line) with increasing weight ratio of PC to apoA-I. Fraction % of apoA-I bound to SUV were derived from Kd and Bmax values. Molar ratios of bound apoA-I to PC on the SUV surface were derived assuming that surface PC is located on the outer leaflet of SUV available for apoA-I binding is 67% of total PC.(参考文献 47 より転載. © the American Society for Biochemistry and Molecular Biology.)
10 20 30 40 50 60 0 2 4 6 8 10 12 0 20 40 60 80 100 100 1-83 1-83/G26R 1 SUV PC/apoA-I (w/w) A p o A -I b o u n d /s u rf a c e P C (m m o l/m o l) ( , ) A p o A -I b o u n d ( % ) ( , )
18
1. 3 アポ A-I 線維化領域ペプチドの線維形成性および脂質膜結合性
これまでの結果から、アポA-I 1-83/G26R による SUV 上での線維形成性の維持は、Iowa(G26R)
変異周辺のヘリックス形成性の低下によるSUV 結合構造の違いが原因で生じることが示唆された。
そこで、脂質膜上での線維化における Iowa(G26R)変異の局所的な影響を評価するため、アポ
A-I 8-33 ペプチドおよびアポ A-I 8-33/G26R へプチドを Fmoc 固相合成法により作製した27,47。アポ A-I N 末フラグメント中には線維形成性の高い領域が N 末端側 14-22 残基領域、および中央部 46-59 残基領域の2 ヶ所存在しており(Figure 1-14)24,31、アポA-I 8-33 残基領域はこの N 末端側の線維化
領域を含む。実際にこれらのペプチドを用いて、水溶液中における線維形成性をThT 蛍光測定(Figure
1-15A)および ThT を利用した TIRFM 観察(Figure 1-15B)により評価したところ、経時的な ThT
蛍光強度の上昇とThT 蛍光を発する凝集物が観察され、これらのペプチドが線維形成性を有するこ
とを確認した63。
Figure 1-14 アポ A-I N 末フラグメントの凝集性予測64,65
Amyloid aggregation propensity of
human apoA-I 1-83
0 10 20 30 40 50 60 70 80 0 5 10 -4 -3 -2 -1 0 1 2 3 14-22 AmylPred2 Zagg (pH 7) 46-59Amino acid residue
A m y lP re d 2 # h it s Za g g a g g re g a tio n p ro p e n s ity
アポA-I 8-33 残基領域は脂質親和性が高く、脂質結合に伴いヘリックス構造へと構造変化する領
域である66。そこでまず、アポA-I 8-33 ペプチドおよびアポ A-I 8-33/G26R ペプチドについて、SUV
に対するITC 測定を行い(Figure 1-16)、得られた熱力学的パラメータを Table 1-3 に示した。アポ
A-I 8-33 ペプチドでは対応するアポ A-I 1-83 フラグメント(Table 1-2)と比較して、SUV への結合親
和性は変わらないが、最大結合量が低下した。また、アポA-I 8-33 ペプチドと比較して、8-33/G26R ペプチドではSUV 結合に伴う発熱が小さく、Iowa 変異によりヘリックス形成性が低下していること が示唆された。実際にCD 測定により脂質結合に伴う二次構造変化を評価したところ、アポ A-I 8-33 残基領域がSUV 結合に伴ってランダムコイル構造からヘリックス構造へ転移すること、および Iowa (G26R)変異によるヘリックス形成性の低下が確認された(Figure 1-17)。このことから、Iowa 変 異が線維化領域のヘリックス形成を阻害することで線維形成性の違いを引き起こす、つまり、ヘリ ックス形成の阻害による不安定な線維化領域の露出が脂質膜上での線維形成を引き起こすのではな いかと考えた。
Figure 1-15 水溶液中におけるアポ A-I 8-33 ペプチドの(A) ThT 経時変化、(B) TIRFM 画像. A,
fibril formation was monitored by ThT fluorescence for apoA-I 8–33 (●) and 8–33/G26R (△) incubated at pH 7.4 in solution. Peptide concentrations were 0.1 mg/ml. B, TIRFM images of apoA-I 8–33 peptides after 120 h incubation at pH 7.4 in solution. Scale bars of TIRFM represent 50 m.(参 考文献 47 より一部変更して転載. © the American Society for Biochemistry and Molecular Biology.)
8-33 (0.1 mg/mL) in solution
0 10 20 30 40 50 60 70 0 10000 20000 30000 8-33/G26R 8-33 Time (h) F lu o re s c e n c e i n te n s it y a t 4 8 5 n m ( a .u .)A B
20 Kd (μg/ml) Bmax (amino acids/mol PC) Ga (kcal/mol) H (kcal/mol) TSb (kcal/mol) 8-33 2.1 ± 0.1 0.059 ± 0.001 -10.8 ± 0.1 -20.2 ± 0.4 -9.4 ± 0.4 8-33/G26R 2.7 ± 0.2 0.041 ± 0.001 -10.7 ± 0.1 -13.7 ± 0.1 -3.0 ± 0.1
Figure 1-16 アポ A-I 8-33 ペプチドの SUV 結合における ITC サーモグラム.
Isothermal titration thermogram for binding of apoA-I 8–33/G26R to egg PC SUV.(参考 文献47 より転載. © the American Society for Biochemistry and Molecular Biology.)
8-33/G26R (0.03 mM)+SUV (PC 1 mM)
0 20 40 60 80 100 -0.14 -0.12 -0.10 -0.08 -0.06 -0.04 -0.02 -0.00Time (min)
He
a
t
fl
o
w
(
c
a
l/
s
)
a Free energy was calculated according to G = -RT ln 55.5(1/K d). b The entropy of binding was calculated from G = H - TS
Table 1-3 アポ A-I 8-33 ペプチドの SUV 結合における熱力学的パラメータ. Thermodynamic
parameters of binding of apoA-I peptides to egg PC SUV at 25℃.(参考文献 47 より一部変更して転載. © the American Society for Biochemistry and Molecular Biology.)
上記、脂質膜上でのヘリックス形成の阻害による不安定な線維化領域の露出が線維形成を引き起
こすのであれば、アポA-I 8-33 ペプチドおよびアポ A-I 8-33/G26R ペプチドを用いることにより、ア
ポA-I 1-83 フラグメントで見られていた脂質膜上での線維形成性の違いが再現可能であると考えら
れる。
そこで、アポA-I 8-33 ペプチドを用いて脂質膜上での線維形成性を評価した(Figure 1-18A)。水
溶液中ではいずれのペプチドも線維を形成していたが(Figure 1-15)、脂質膜上ではアポ A-I 8-33 ペ
プチドの線維形成が阻害され、一方、アポA-I 8-33/G26R ペプチドは線維形成性を維持するという、
アポA-I 1-83 フラグメントと同様の結果が得られた。ThT を利用した TIRFM 観察からも、アポ A-I
8-33/G26R ペプチドのみ脂質膜上で ThT 蛍光を発する凝集物が観察された(Figure 1-18B)。 以上の結果から、Iowa(G26R)変異による脂質膜上での線維化促進は、N 末端側の線維化領域が 局所的にヘリックス構造を形成せず、ランダムコイル構造のままであるための構造のゆらぎが原因 となっている可能性が示唆された。 序論および1. 1 にて述べたように、他のアミロイドタンパク質においては、脂質膜の存在は線維 形成を促進するための二次元濃縮の足場となると言われている35-37, 58。本研究において、アポA-I 1-83 は脂質膜上での線維形成が阻害され、一方アポA-I 1-83/G26R は線維形成性を維持するという興味深 い結果を示した。この結果は、他のアミロイドタンパク質のように、脂質膜による二次元濃縮効果 や最適な脂質/タンパク質濃度比の存在だけでは説明できない。そのため、1. 3 において示唆された、 Iowa 変異による局所的なヘリックス形成性の低下が線維形成領域の露出を引き起こすという新規機 序について、より詳細に検証する必要があると考えた。
Figure 1-17 (A) CD スペクトル、(B) ヘリックス含量. A, far-UV CD spectra of apoA-I 8–33 and 8–
33/G26R peptides in solution (a) or bound to egg PC SUV (PC = 3.0 mg/ml) (b and c). The peptide concentration was 50 g/ml. B, -helix contents of apoA-I 8–33 and 8–33/G26R bound to egg PC SUV at various weight ratios of PC to peptide.(参考文献 47 より転載. © the American Society for Biochemistry and Molecular Biology.)
CD spectra of
apoA-I 8-33 + Egg PC SUV
200 210 220 230 240 250 260 -30000 -20000 -10000 0 10000 20000 (a) 8-33 or 8-33/G26R (b) 8-33 + SUV (c) 8-33/G26R + SUV Wavelength (nm) M o la r e ll ip ti c it y (d e g c m 2 /d m o le )
-helix content
0 10 30 60 0 20 40 60 8-33 8-33/G26R SUV PC/peptide (w/w) -h e li x c o n te n t (% )A B
22
1. 4 部位特異的蛍光プローブ標識による脂質膜上でのアポ A-I 局所環境の解析
SUV 上における、アポ A-I 1-83 は線維形成が抑制されるが、アポ A-I 1-83/G26R は線維形成性を
維持するという違いについて、Iowa(G26R)変異が N 末端側の線維形成領域のヘリックス形成性を 低下させることが原因となっているのではないか、と述べてきた。1. 4 ではさらに詳細に SUV 結合 時の局所的な構造の違いを評価するため、内在性のトリプトファン(W、Trp)蛍光および環境感受 性蛍光プローブであるアクリロダン(Ac)を用いた蛍光測定を行った。 アポA-I 1-83 フラグメント中には、8、50、72 番目の 3 ヶ所に Trp 残基が存在する。この Trp 蛍光 を利用し、各位置のTrp 残基をそれぞれ独立して評価することができれば、アポ A-I 1-83 フラグメ ント中のN 末端側、中央部、C 末端側それぞれの局所的な評価が可能となる。そこで、各 Trp 残基 を1 つずつ残し、他 2 ヶ所の Trp 残基をフェニルアラニン(F)残基に置換したアポ A-I single Trp 変 異体を作製した27。8 番目の Trp 残基のみを残した変異体(アポ A-I 1-83/W50F/W72F)を W@8 と表
記し、他同様にW@50、W@72 と表記する。Figure 1-19A にはアポ A-I 1-83 W@8 の水溶液中および
SUV 結合時の Trp スペクトルを示す。このように、SUV 結合に伴って Trp 最大蛍光波長(wavelength of maximum fluorescence;WMF)の短波長側へのシフト、および、蛍光強度の増大を示した。1. 3 の Figure 1-17A に示すアポ A-I 8-33 ペプチドの CD 測定から、8 残基目周辺は脂質膜に結合し構造変化 する領域であることが分かっている。そのため、Figure 1-19A の Trp スペクトル変化は、8 残基目周
辺の環境がSUV に結合して疎水的な環境に移行したことを示す。同様の測定をアポ A-I 1-83/G26R
の各single Trp 変異体について行い、Figure 1-19B に水溶液中および SUV 結合時の WMF の比較を行
った。アポA-I 1-83/G26R W@8 および W@50 は SUV 結合に伴い WMF がシフトし、これらの領域
8-33 (0.1 mg/mL)
on SUV
at 250rpm 37
oC
0 10 20 30 40 50 0 10000 20000 30000 8-33/G26R + SUV 8-33 + SUV Time (h) F lu o re s c e n c e i n te n s it y a t 4 8 5 n m ( a .u .)A B
Figure 1-18 脂質膜上におけるアポ A-I 8-33 ペプチドの (A) ThT 経時変化、(B) TIRFM 画像. A,
fibril formation was monitored by ThT fluorescence for apoA-I 8–33 (●) and 8–33/G26R (△) incubated at pH 7.4 in the presence of egg PC SUV. Peptide and PC concentrations were 0.1 and 6.0 mg/ml, respectively. B, TIRFM images of apoA-I 8–33 peptides after 120 h incubation at pH 7.4 in the presence of egg PC SUV. Scale bars of TIRFM represent 50 m.(参考文献 47 より転載. © the American Society for Biochemistry and Molecular Biology.)
は脂質膜に結合して構造変化する領域であるが、アポA-I 1-83/G26R W@72 は水溶液中と SUV 結合 時にWMF の差が見られず、SUV 結合時の構造変化には関与しない領域であることを示した。 著者は以前、アポA-I 1-43/G26R、44-65、66-83 ペプチドを作製し、それぞれのペプチドの SUV 結 合時の二次構造変化を評価しており、Figure 1-20 にはそのヘリックス含量の変化を示した。このよ うに、アポA-I 1-43/G26R および 44-65 ペプチドは SUV 結合に伴いヘリックス構造を形成するが、 アポA-I 66-83 ペプチドはヘリックス含量に変化が見られないことが分かる。今回得られた single Trp 変異体の結果は、各 Trp 残基を含む対応するアポ A-I ペプチドの結果と一致しており、アポ A-I 1-83/G26R フラグメント中においても 72 番目周辺の C 末端領域は SUV 結合時のヘリックス構造形 成に関与しないことが示された。 300 320 340 360 380 400 420 0 500 1000 1500 2000 on SUV in solution Wavelength (nm) F lu o re s c e n c e in te n s ity (a .u .)
Figure 1-19 (A) アポ A-I 1-83 W@8 の Trp 蛍光スペクトル、(B) アポ A-I 1-83/G26R single Trp
変異体の最大蛍光波長. A, Trp fluorescence spectra of apoA-I 1–83 W@8 (50 g/ml) in the absence (solid line) or presence (dashed line) of SUV (1.5 mg/ml of PC). The excitation wavelength was 290 nm. B, change in WMF of apoA-I 1–83/G26R single Trp variants by binding to SUV.(参考文献 47 よ り転載. © the American Society for Biochemistry and Molecular Biology.)
A B
W@8 W@50 W@72 335 340 345 350 355 in solution on SUV W M F ( n m )24
SUV 結合時に疎水的環境への移行が見られたアポ A-I 1-83/G26R W@8、W@50 について、Iowa (G26R)変異が与える影響を評価した。Figure 1-21 には SUV 結合時のアポ A-I 1-83 およびアポ A-I 1-83/G26R それぞれの W@8 と W@50 の Trp 蛍光 WMF を示す。このように、いずれの位置について もアポA-I 1-83 と比較してアポ A-I 1-83/G26R では WMF が長波長側にシフトしており、Iowa(G26R) 変異の影響で親水的環境にあることが示された。
Figure 1-21 アポ A-I single Trp 変異体の WMF の比較.(参考文献 47 より
転載. © the American Society for Biochemistry and Molecular Biology.)
W@8 W@50 335 340 345 350 1-83 1-83/G26R W M F ( n m )
Figure 1-20 アポ A-I ペプチドの SUV 結合におけるヘリックス含量の変化
helix content
1-43/G26R
44-65
66-83
0 10 20 30 40 in solution SUV PC/peptide = 10 (w/w) SUV PC/peptide = 30 (w/w) SUV PC/peptide = 60 (w/w)
-h
e
li
x
c
o
n
te
n
t
(%
)
著者は以前、脳内でのコレステロール代謝を担うアポリポタンパク質E(アポ E)について、部位 特異的な蛍光標識の手法を用いて、それぞれの領域の特異的な自己会合性および脂質結合性の評価 を行った57。これにより、アポE の脂質膜への結合が経時的に 2 段階の結合挙動を示すことを明ら かとした。今回、この手法を適用することにより、Iowa(G26R)変異により伸長された N 末端側の 線維化領域(14-31 残基領域)中にある Iowa(G26R)変異周辺の環境の評価を試みた。Iowa 変異近 傍である22 番目のロイシン(L)をシステイン(C、Cys)に置換した変異体アポ A-I 1-83/L22C お よびアポA-I 1-83/L22C/G26R を作製し、導入した Cys 残基にチオール基特異的な蛍光色素アクリロ ダン(Ac)を標識した63,67,68。
Figure 1-22A にはアポ A-I 1-83/L22C-Ac の水溶液中および SUV 結合時の Ac 蛍光スペクトルを示
す。SUV 結合に伴い Ac 蛍光 WMF の短波長側へのシフト、および、蛍光強度の増大が見られたこと
から、標識部位周辺が疎水的環境へ移行したことが分かる69,70。Figure 1-22B に示すように、アポ A-I
1-83/L22C-Ac およびアポ A-I 1-83/L22C-Ac/G26R の WMF を比較したところ、水溶液中では変異体間 の差異は見られないが、SUV 結合時にアポ A-I 1-83/L22C-Ac/G26R では WMF がより長波長側にあ り、Iowa(G26R)変異により親水的環境に移行したことを示す。
これらのsingle Trp 変異体および Cys 置換変異体を用いた蛍光測定から、Iowa(G26R)変異はア
ポA-I 1-83 フラグメントの N 末端側の線維化領域(14-31 残基領域)をより水層に露出させる効果を
有することが示された。
ApoA-I 1-83/L22C-Ac spectra
400 450 500 550 600 0 100 200 300 400 500 in solution on liposome protein conc. 25 ug/ml
PC 750 ug/ml (PC/protein = 30) Ac labeled : 10 % (2.5 ug/ml) on SUV in solution Wavelength (nm) F lu o re s c e n c e i n te n s it y ( a .u .)
Figure 1-22 (A) アポ A-I 1-83/L22C-Ac の蛍光スペクトル変化、(B) 最大蛍光波長の比較. A,
acrylodan fluorescence spectra of apoA-I 1–83/L22C-Ac (acrylodan) (50 g/ml) in the absence (solid line) or presence (dashed line) of SUV (1.5 mg/ml PC). B, changes in WMF of apoA-I 1–83/L22C-Ac and 1–83/L22C-Ac/G26R by binding to SUV.(参考文献 47 より転載. © the American Society for Biochemistry and Molecular Biology.)
A B
WMF
1-83/L22C-Ac 1-83/L22C-Ac/G26R 450 460 470 480 490 500 510 in solution on SUV W M F ( n m )26
ここで、1. 1 の Figure 1-6B で見られた、アポ A-I 1-83/G26R の脂質膜上での線維化における脂質濃
度依存的なlag time の延長について考察する。Figure 1-14 に示した通り、アポ A-I 1-83 フラグメント
中には線維形成性の高い領域がN 末端側 14-22 残基領域、および中央部 46-59 残基領域の 2 ヶ所存 在する。それぞれの領域の線維形成における役割について、アポ A-I 1-43 ペプチドおよびアポ A-I 44-65 ペプチドを用いて調べたところ、中央部 46-59 残基領域が線維形成の核となり、N 末端側 14-22 残基領域は線維核からの伸長過程において重要な領域であることが分かっている 63。ところが、 Figure 1-20 より、中央部 46-59 残基領域を含むアポ A-I 44-65 ペプチドが脂質膜上で高いヘリックス 形成性を示すことが分かった。このことから、線維核形成に必要な 46-59 残基領域が脂質膜上にお いてヘリックス構造を形成し、線維形成領域が露出していないために核形成過程の延長、つまり、lag time の延長につながったものとみられる。すなわち、アポ A-I 1-83/G26R の脂質膜上での線維形成に おいて、他のアミロイドタンパク質で見られるような、脂質膜による線維化のlag time を促進する効 果が見られないことは、アポA-I 1-83 フラグメント中の線維形成領域とヘリックス形成領域の重複 がその一因となっているためであると考えられる。
1. 5 脂質膜結合によるアポ A-I ヘリックス構造の安定化
ここまで、SUV 上でのへリックス形成性に着目し、Iowa(G26R)変異により脂質膜上でへリック ス形成性が低下することが線維形成につながると考察してきた。1. 5 では形成されたへリックス構造 の安定性に着目し、アポA-I 1-83 とアポ A-I 1-83/G26R 間での比較を行った。 SUV 結合時のアポ A-I 1-83 変異体に、変性剤である塩酸グアニジンを添加した際のへリックス構 造の安定性をCD 測定により評価した。変性剤を添加した際のアポ A-I 1-83 の CD スペクトルの変化 をFigure 1-23A に示す。222 nm の平均残基モル楕円率から、各グアニジン塩酸塩濃度における変性 率を算出したグラフをFigure 1-23B に、この変性曲線から得られたパラメータを Table 1-4 に示す71。 GD〇は変性ギブズ自由エネルギーを、D1/2は変性中点を、m は変性の協同性を表すパラメータである。 パラメータを比較すると、アポA-I 1-83/G26R はアポ A-I 1-83 より低濃度のグアニジン塩酸塩濃度で 変性中点(D1/2)を示し、GD〇が低下していることから、形成されたへリックス構造がより不安定で あることが分かる。 すなわち、Iowa 変異は、アポ A-I 1-83 フラグメントの N 末端側のへリックス形成性およびへリッ クス構造の安定性を低下させることで脂質膜上での線維形成を促進していることが示された。Table 1-4 SUV 結合アポ A-I N 末フラグメントの変性パラメータ GD 〇 (kcal/mol) D1/2 (M) m (kcal/mol apoA-I/mol GdnHCl) 1-83 3.5 ± 0.2 2.2 ± 0.4 1.6 ± 0.1 1-83/G26R 1.9 ± 0.3 1.4 ± 0.4 1.4 ± 0.1
1. 6 小括
脂質膜環境でのアミロイド線維形成性を、アミロイド線維に特異的な蛍光色素チオフラビン T (ThT)測定、および、透過型電子顕微鏡(TEM)観察や ThT を利用した全反射蛍光顕微鏡(TIRFM) 観察により評価したところ、脂質膜への結合によりアポA-I 1-83 フラグメントのアミロイド線維形成は著しく阻害される一方、Iowa(G26R)変異を有するアポ A-I 1-83 フラグメント(アポ A-I 1-83/G26R)では脂質膜上においても線維形成性を有することが明らかとなった。 アポA-I 1-83 フラグメント中には 2 ヶ所の線維形成性の高い領域が存在する。円偏光二色性(CD) 測定より、水溶液中で主にランダムコイル構造であるアポA-I 1-83 フラグメントは、脂質膜に結合 するとヘリックス構造を形成することが示されたことから、脂質膜上での安定なヘリックス構造形 成によって線維形成が阻害されたと考えられた。一方、Iowa 変異はアポ A-I 1-83 フラグメントのヘ リックス形成性を低下させることが示され、へリックス構造を形成せずに露出した不安定な線維化 領域が相互作用することにより、アポA-I 1-83/G26R は脂質膜上においても線維形成性を維持すると 考えられた(Figure 1-24)。 1-83 on EPC CD spectra 200 210 220 230 240 250 260 -20000 -10000 0 10000 20000 30000 GdnHCl 0 - 4 (M) Wavelength (nm) M o la r e ll ip ti c it y (d e g c m 2 / d m o le )
A B
Figure 1-23 (A) グアニジン塩酸塩添加時の CD スペクトル変化、(B) 変異体間での変性曲線の比較. A, changes in CD spectra of apoA-I 1‒83 bound to egg PC SUV upon increasing concentrations of GdnHCl. B, GdnHCl denaturation curves of apoA-I 1‒83 (●) and 1‒83/G26R (△) bound to egg PC SUV.
1-83 vs 1-83/G26R on EPCSUV Fraction unfolded 0 1 2 3 4 0.00 0.25 0.50 0.75 1.00 1-83 1-83/G26R GdnHCl (M) F ra c to n u n fo ld e d
28 上記の仮説を検証するため、環境感受性蛍光プローブを利用したアポ A-I タンパク質の局所環境 の評価を試みた。Iowa 変異近傍にシステイン残基を導入した変異体(L22C)を新たに設計・作製し、 チオール基特異的な蛍光プローブであるアクリロダンを標識した。アクリロダンは周囲環境により 最大蛍光波長がシフトする特性を有するため、脂質膜結合時の局所的な環境評価が可能である。結 果、アポA-I 1-83 フラグメントでは標識部位周辺が脂質膜結合によりへリックス構造を形成する領 域であることを示した。一方、アポA-I 1-83/G26R では、アポ A-I 1-83 フラグメントよりも標識部位 周辺が親水的な環境にあることが示された。これは、Iowa 変異は脂質膜上において不安定な線維化 領域の水層への露出を引き起こすことを示す。 本研究から、アポA-I の代表的なアミロイドーシス変異である Iowa(G26R)変異は、脂質膜上で のヘリックス形成性の低下を引き起こすことにより、細胞膜などの生体膜脂質を足場としたアミロ イド線維形成に重要であることが示され、Iowa 変異がアポ A-I アミロイドーシスを引き起こす分子 メカニズムの解明につながる知見が得られた。 Figure 1-24 アポ A-I 1-83/G26R の脂質膜上での線維形成のモデル図
2 章 アミロイドーシス変異アポ A-I のアミロイド線維形成に及ぼすホスファチジルセリ
ンおよびコレステロールの影響
2. 1 脂質膜上でのアポ A-I アミロイド線維形成性の変化
1 章ではアポ A-I 1-83 フラグメントの線維形成における脂質膜の影響について評価を行った。その 結果、Iowa(G26R)変異により脂質膜上での線維形成が促進されることが明らかとなった。
2 章では Iowa 変異型アポ A-I 1-83 フラグメント(アポ A-I 1-83/G26R)の脂質膜上での線維形成に おける脂質組成の影響を評価するため、中性のホスファチジルコリン(PC)に加え、酸性リン脂質 であるホスファチジルセリン(PS)もしくはコレステロール(Chol)を用いて SUV を作製し、各 SUV 上での線維形成性について ThT 測定を行った。SUV の脂質組成は、PC のみ、PC/PS = 17/3 mol/mol (PS 15 %)もしくは PC/PS = 7/3 mol/mol(PS 30 %)、および PC/Chol = 2/1 mol/mol(Chol 33 %)の 4 種類である。Figure 2-1A はアポ A-I 1-83/G26R 50 g/ml に対し、SUV PL 濃度を 0.5 mg/ml(PL/ア ポA-I = 10(w/w))、Figure 2-1B は SUV PL 濃度を 1.5 mg/ml(PL/アポ A-I = 30(w/w))として ThT
蛍光経時変化を測定したグラフを示す。いずれのPL 濃度においても PS 含量を 15 %、30 %と増大さ
せたところ、PS 濃度依存的な線維形成の抑制が見られ、特に PS 30 %においては SUV PL 濃度 1.5 mg/ml(PL/アポ A-I = 30(w/w))の場合に線維化が完全に抑制された。一方、コレステロールの存
在はThT 蛍光上昇を促進しており、線維化を促進することが分かった。ThT 蛍光経時変化をフィッ
ティングすることで得られた線維化のlag time および線維伸長速度定数 k を、それぞれ Figure 2-2A
およびFigure 2-2B に示す55。これらのパラメータを比較すると、PS の存在は濃度依存的に lag time
を延長するが線維伸長速度には影響を与えておらず、脂質膜上での線維化における核形成過程を遅 らせることが分かった。一方、コレステロールはlag time を短縮しており、線維化の核形成過程を促 進していることが分かった。アポA-I 1-83 フラグメントについても同様に、組成の異なる各 SUV 上 での線維形成を評価したところ、いずれのSUV 上においても線維化は見られなかった(Figure 2-3)。 すなわち、1 章で述べたように Iowa 変異には脂質膜上でのアポ A-I 1-83 フラグメントの線維形成 性を維持する効果があるが、PS の存在は、この Iowa 変異による線維形成促進効果を阻害すること が分かった。また、PS による線維形成阻害効果は、線維伸長過程ではなく、核形成過程に対して大 きく影響を及ぼしている。既に述べた通り、アポA-I 1-83 フラグメント中に 2 ヶ所存在する線維形 成領域のうち、中央部46-59 残基領域が線維形成の核となる領域であり、N 末端側 14-22 残基領域は 線維核からの伸長過程において重要な領域である63。そのため、水溶液中とPC SUV 上を比較した 際のlag time の延長は、中央部 46-59 残基領域が安定なヘリックス構造を形成することによるものと
考えられた。しかし、PS の存在はさらに大きな lag time の延長につながっている。Iowa 変異は線維 化の核形成および伸長過程のいずれも促進する効果を有するため(Table 1-1)、PS とアポ A-I 1-83/G26R との相互作用には、Iowa 変異により延長された N 末端側 14-31 残基の線維形成領域が大 きく関与しているのではないかと考えられる。
30 0 60 120 180 240 0.0 0.2 0.4 0.6 0.8 1.0 Chol33% PC in solution PS15% PS30% 30 SUV PL Time (h) No rm a liz e d T h T flu o re s c e n c e in te n s ity ( a .u .) 0 50 100 150 N.D. in solution PC PS 15% PS 30% x10 SUV PL x30 SUV PL Chol 33%
***
***
***
****
****
**
L a g t im e ( h ) 0.000 0.025 0.050 0.075 0.100 0.125 N.D. in solution PC x10 SUV PL x30 SUV PL PS 15% PS 30% Chol 33%***
****
**
*
****
****
k ( h -1 )Figure 2-2 線維化の速度論的パラメータの比較. Comparison of lag time (A) and apparent rate
constant (B) for the fibril growth of apoA-I 1‒83/G26R bound to SUV. N.D., not determined. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001 versus“PC”.(参考論文 48 より転載)
A B
Figure 2-1 アポ A-I 1-83/G26R の脂質膜上での線維形成性. PL/apoA-I weight ratios were 10 (A) and
30 (B). Dotted line, in solution; ○, PC SUV; ▼, PS 15 % SUV; □, PS 30 % SUV; ▲, Chol 33 % SUV. ApoA-I 1‒83 variants were incubated at 37 °C with agitation on an orbital rotator in the presence of 10 μM ThT. Protein concentration was 0.05 mg/ml.(参考論文 48 より転載)
A B
0 60 120 180 240 0.0 0.2 0.4 0.6 0.8 1.0 Chol33% PC in solution PS15% PS30% 10 SUV PL Time (h) No rm a liz e d T h T flu o re s c e n c e in te n s ity ( a .u .)Figure 2-4 には各 SUV 上でインキュベートしたアポ A-I 1-83/G26R の TEM および TIRFM 画像を示
す。ThT 蛍光測定により線維化が確認された Chol 33 % SUV 上のサンプルは、PC SUV の場合と同様
に線維状の凝集物が確認された。一方PS 30 % SUV の場合は ThT 蛍光の上昇も見られず、線維状の
凝集物も確認できなかった。
Figure 2-3 アポ A-I 1-83 の脂質膜上での線維形成性. Fibril formation was monitored by ThT
fluorescence for apoA-I 1‒83 in the presence of SUV. PL/apoA-I weight ratio was 30. Dotted line, in solution; □, PC SUV; ▽, PS 30 % SUV; ▲, Chol 33 % SUV. Protein concentration was 0.05 mg/ml.(参考文献 48 より転載)
ApoA-I 1-83 + SUV
ThT time course (normalize)
0 60 120 180 240 0.0 0.2 0.4 0.6 0.8 1.0 in solution SUVs Time (h) No rm a liz e d T h T flu o re s c e n c e in te n s ity ( a .u .)
Figure 2-4 脂質膜上で形成されたアミロイド線維の形態観察. TEM and TIRFM images of
apoA-I 1‒83/G26R after incubation for 120 h in the presence of SUV at PL/apoA-I weight ratio of 10. Scale bars indicate 200 nm and 20 m, respectively.(参考文献 48 より転載)
PC SUV PS 30 % SUV Chol 33 % SUV
200 nm 200 nm 200 nm
20 µm 20 µm 20 µm