総合研究題目
マウス腎臓における 2 つのマクロファージ集団の同定と その機能および糖尿病性腎症の新規治療法に関する研究
伊
い藤
とう誓
せい悟
ご(腎臓病学専攻)
防衛医科大学校
令和 2 年度
目次 頁
第
1章 背景 … 1
第
2章 研究
1腎臓における機能に着目した
2種類のマクロファージの同定と糖尿病性腎症への 関与
第
1節 背景および目的 …
3第
2節 材料と方法
(1)
使用実験動物 …
5(2) 尿アルブミン排泄量、血清クレアチニン値および随時血糖値の測定
… 5
(3)
腎臓および糸球体の細胞懸濁液の作成 …
5(4) フローサイトメトリー
… 6
(5) Mφs
の
TNF-α産生能の評価 …
7(6) Mφs
の
ROS産生の評価 … 8
(7) Mφs
の貪食能の評価 …
8(8) 腎ホモジネートTNF-α
の測定 … 8
(9)
腎臓の組織学的評価 …
9(10) 糸球体におけるWilms tumor 1
陽性細胞の評価 … 9
(11)
間質における
alpha smooth muscle actin発現の評価 …
10(12) 腎に限局した間歇的低線量放射線照射
… 11
(13)
統計解析 …
11第
3節 結果
(1)
腎における
2種類の
Mφs集団の同定 …
13(2) 糖尿病の有無と腎TNF-α
および各
Mφs機能の関連 … 13
(3) 糖尿病性腎症の進行に伴う尿アルブミン排泄とCD11bhigh Mφs
の増加 … 14
(4)腎症進行モデルにおける
CD11bhigh Mφsと
CD11blow Mφsの性質 …
14(5) 腎症進行モデルにおけるCD11bhigh Mφs
と
CD11blow Mφsの機能 … 15
(6)糖尿病性腎症の進行と腎
Mφsの機能変化 …
16(7) 糸球体におけるMφs
の分布と
TNF-α産生能 … 16
(8)
腎に限局した間歇的低線量放射線照射が腎症へ及ぼす効果 …
16(9) 腎に限局した間歇的低線量放射線照射が腎Mφs
へ及ぼす効果 … 17
第
3章 研究
2糖尿病性腎症マウスモデルにおける
C-Cケモカイン受容体タイプ
2アンタゴニスト の治療効果
第
1節 背景および目的 …
19第
2節 材料と方法
(1)
使用実験動物 …
21(2) 尿アルブミン排泄量、血清クレアチニン値および随時血糖値の測定
… 21
(3)
腎臓の細胞懸濁液の作成 …
21(4) フローサイトメトリー
… 21
(5) Mφs
の
TNF-α産生能の評価 …
21(6) Mφs
の
ROS産生の評価 … 21
(7) Mφs
の貪食能の評価 …
21(8) 腎臓の組織学的評価
… 21
(9)
糸球体における
Wilms tumor 1陽性細胞の評価 …
21(10) INCB3344
の投与 … 21
(11)
統計解析 …
21第
3節 結果
(1) INCB3344
が腎症および全身に及ぼす効果 … 23
(2) INCB3344
が腎臓の炎症や
Mφs分布に及ぼす効果 …
23(3) INCB3344
が腎
Mφsの
M1/M2フェノタイプや機能に及ぼす効果 … 23
第
4章 研究
3糖尿病性腎症マウスモデルにおける
L-カルニチンの治療効果ならびに腎
Mφsに与 える影響
第
1節 背景および目的 …
25第
2節 材料と方法
(1)
使用実験動物 …
26(2) 尿アルブミン排泄量、血清クレアチニン値および随時血糖値の測定
… 26
(3)
腎臓の細胞懸濁液の作成 …
26(4) フローサイトメトリー
… 26
(5) Mφs
の
TNF-α産生能の評価 …
26(6) Mφs
の
ROS産生の評価 … 26
(7) Mφs
の貪食能の評価 …
26(8) 腎ホモジネートTNF-α
の測定 … 26
(9)
腎臓の組織学的評価 …
26(10) 糸球体におけるWilms tumor 1
陽性細胞の評価 … 26
(11)
腎実質細胞および
Mφsにおけるミトコンドリア由来
ROS産生の評価 …
26(12) 近位尿細管細胞におけるミトコンドリア由来ROS
産生の評価 … 27
(13)
腎組織における
Superoxide dismutase 2発現の評価 …
27(14) L-カルニチンの投与
… 28
(15)
統計解析 …
28第
3節 結果
(1) 糖尿病モデルマウスの腎実質細胞および腎 Mφs
におけるミトコンド リア由来
ROSの解析
… 29
(2) L-カルニチンが腎実質細胞および Mφs
のミトコンドリア由来
ROSに及ぼす効果
… 29
(3) L-カルニチンが近位尿細管細胞のミトコンドリアに及ぼす効果
… 29
(4) L-
カルニチンが腎
SOD2発現に及ぼす効果 …
30(5) L-カルニチンが腎症へ及ぼす効果
… 30
(6) L-
カルニチンが腎
Mφsの分布や機能に及ぼす効果 …
30第
5章 考察 …
32第
6章 結論 …
48謝辞 …
50略語一覧 … 51
引用文献 …
53図 … 72
1
第
1章 背景
糖尿病の重大な合併症のひとつである糖尿病性腎症は、先進国および発展途 上国の両方を含む全世界において、末期腎不全の主要な原因疾患であり
1、特に 本邦においては、血液透析に至る患者の
40%を占め、透析導入後の 5年生存率 は
50%程度とその予後は非常に不良である
2。したがって、その病態の解明と治 療法の確立は、これらの患者の予後の改善において重要である。
糖尿病、アテローム性動脈硬化症、非アルコール性脂肪性肝炎(
non-alcoholic steatohepatitis:
NASH) に 至 る 基 礎 病 態 と し て メ タ ボ リ ッ ク シ ン ド ロ ー ム
(
metabolic syndrome:
MetS)が存在し
3、近年、
MetSに関連したマウス実験モ デルにおいては、マクロファージ(Mφs)およびそれが産生する
tumor necrosisfactor-α
(
TNF-α)が慢性炎症を介し病態の進行に関連しており、特に接着因子で
あり補体
C3b受容体でもある
CD11bを高発現した
Mφs(CD11b
high Mφs)が関与していることが報告されている
4-7。
CD11bhigh Mφsは、自己免疫性腎炎の病態へ の関与が報告されており、近年、腎臓病学の領域においても注目を集めている
Mφs集団である
8-10。さらに、糖尿病性腎症については活性酸素種(
reactive oxygenspecies:ROS)もまた、病態に関与していることが示唆されている11
。
これまでに、関らは、肝臓において、
Mφsをその表現型と機能に応じて、骨 髄由来(bone marrow-derived:BM-)Mφs と、組織固有(tissue resident:Res-)
Mφs
という
2つの集団に分類した
12-16。さらに、
BM-Mφsおよびそれらの
TNF- α産生が細菌感染に関連し、
NASH病態の誘発に重要であることを示した
16。ま
た、最近
10年程の間に、肝臓の
Kupffer細胞、脳のミクログリア、皮膚のラン
ゲルハンス細胞などの
Res-Mφsは、胎児の卵黄嚢に由来し臓器固有の前駆細胞
より分化した細胞であることが示され、
BM-Mφsと発生学的にも機能的にも異
なる集団であると捉えられるようになってきた
17-20。実際に、マウスの肝臓
Mφs2
においては、
Res-Mφs(
Kupffer細胞)が
ROSを生成し、細菌に対して強力な貪 食・殺菌作用を示す一方で、炎症性サイトカインの産生能は弱く、放射線抵抗性 の性質を持ち、対照的に、
BM-Mφsは、貪食作用は弱いものの、強い炎症性サイ トカインの産生能を有し、放射線感受性の性質を持つことが明らかにされた
12,13,15
。
しかしながら、腎臓においては、様々な
Mφsの分類が試みられているものの
21
、
BM-Mφsと
Res-Mφsという異なる
2種類の
Mφsという観点からの腎臓にお ける分布とその機能の解析はこれまでなされていない。
本研究は、腎臓における、これらの
2種類の
Mφsの分布と機能、およびそれ
らが糖尿病性腎症の病態に対し、いかに関わるかを明らかにし、またその結果を
踏まえ、新規の糖尿病性腎症に対する治療法を探索することを目的とする。
3
第
2章 研究
1腎臓における機能に着目した
2種類のマクロファージの同定と糖尿病性腎症への関 与
第
1節 背景および目的
近年、糖尿病性腎症の病態の根底には慢性炎症が存在し、マウスモデルを使 用した研究においては、
CD11b陽性
Mφsの関与が報告されている
4,5。
Mφsを活 性化する因子として、
Mφsに発現する
Toll様受容体(toll-like receptor:TLR)が 知られており、これらは感染初期における病原体のパターン認識受容体として 作動し、
TLR4はグラム陰性桿菌の菌体抗原であるリポ多糖(lipopolysaccharide :
LPS)を認識し、
TLR9は細菌が普遍的にもつ非メチル化
CpGモチーフを認識す る
22。TLR9 はマウスにおいては主に
Mφsと
Bリンパ球に存在し、非メチル化
CpGモチーフをもつ細菌
DNAにより刺激され活性化することで、
TNF-α産生 に関与し、細菌感染時の初動免疫として機能する
23,24。なお、TLR9 活性化には 負の側面も存在し、敗血症性急性腎障害の病態に関与するとの報告もあり
25、私 どもの過去の研究においても、CpG 刺激により、TLR9 を介し、Mφs と
naturalkiller T
(
NKT)細胞が互いに協同して、急性の肝障害および腎障害を起こしたこ
とを示した
26-28。最近、これらの受容体は病原体の認識装置として感染防御に関わるのみなら
ず、代謝性疾患に生ずる全身性の慢性炎症の端緒としても機能している可能性
が指摘されており、特に、
TLR4については、糖尿病性腎症の病態への関与が示
唆されている
4,5,29。他方、
TLR9については、
MetSおよび肥満症における、自然
免疫活性化より始まる慢性炎症病態の契機となりうることが示唆されているも
のの
30、糖尿病性腎症における関与については不明である。
4
以上を踏まえ、研究
1においては、まず
2型糖尿病モデルマウスと非糖尿病
正常マウスの腎臓における
Mφsの存在と分布、それらの様々な機能とその違い
を解析した。その上で、
Mφsにおける
TLR9発現と糖尿病性腎症の病態との関
連について解析した。さらに、糖尿病性腎症の新規治療として腎臓への低線量放
射線照射を行い、その効果を検討した。
5
第
2節 材料と方法
(1) 使用実験動物
雄
5-20週齢の
2型糖尿病モデル
db/dbマウス(
C57BLKS/J lar- +Leprdb/+Leprdbマウス)と、その非糖尿病正常対照群として
mistyマウス(C57BLKS/J lar-m+/m+
マウス)を日本クレア(東京)から購入し使用した。全てのマウスは
12時間置 きの明暗サイクル下かつ自由に飲水および摂餌可能な環境で飼育された。
本研究で用いた動物実験の手順は防衛医科大学校動物実験倫理委員会により 承認されており(承認番号:19004) 、全ての実験においては、動物の苦痛を最小 限化する最大限の努力を払うとともに、倫理委員会が定めたガイドラインと規 則を遵守して行った。
(2) 尿アルブミン排泄量、血清クレアチニン値および随時血糖値の測定
db/dbマウスは
5、
8、
12、
16、
20週齢において、
mistyマウスは
8、
12、
16、
20週齢において、代謝ケージ(CL-0305、日本クレア)を用いて
24時間、自由 飲水および摂餌可能な環境で蓄尿を行った。蓄尿中の飲水量ならびに摂餌量も 同時に測定された。得られた尿を
4℃・1500 rpmで
5分間遠心後、その上清を採 取し、マウス尿中アルブミン酵素結合免疫吸着測定法(
enzyme-linked immuno- sorbent assay:ELISA)キット(Albuwell M; Exocell, Philadelphia, PA)を用い定量した。
血清クレアチニン(Cr)値と随時血糖値は、自由に摂餌可能な環境下で血液 サンプルを採取し、それぞれ酵素法により測定した(
SRL、東京)。
(
3) 腎臓および糸球体の細胞懸濁液の作成
マウスを深麻酔下で安楽死させ、生理食塩水による灌流後に腎臓を摘出した。
6
摘出した左腎臓を細切後に
37℃の温浴槽で
40分間コラゲナーゼ処理し(コラゲ ナーゼ;富士フイルム和光純薬、大阪) 、ステンレススチール製メッシュに濾過 して溶解した。その細胞溶解液を等張性
30%パーコール溶液(
Sigma, St. Louis,MO)に再懸濁し、等張性60%パーコール溶液に重層した後、15℃・1500 rpm
で
30
分間遠心分離した。遠心分離後に、
60%と
30%のパーコール溶液の間に生成 する単核球(mononuclear cell:MNC)が豊富に含まれる層を収集し、赤血球溶解 溶液に再懸濁した後、
40 μmのポリプロピレン製セルストレーナーで濾過した。
本法は、肝臓
MNCを採取するために確立された手法を改変したものである
27。 また、糸球体の細胞懸濁液については、
100 μmおよび
70 μmのセルストレー ナーを用いたシービング法による糸球体単離の後に、腎臓全体の細胞懸濁液と 同様に作成した。なお、その際は右腎を糸球体の解析に供し、左腎を腎全体の解 析に供することで、同一マウスにおいて糸球体と腎全体の細胞を比較出来るよ うにした。
(
4) フローサイトメトリー
使用抗体と免疫細胞が持つ
Fc受容体との非特異的結合を防止する目的で、腎 臓
MNCを豊富に含む細胞懸濁液へ
Fcブロッカー(
Clone: 2.4 G2; BD Biosciences,Franklin Lakes, NJ)を加え、4℃・15
分間インキュベートした。続いて、Mφs を
同定する目的で、
APC標識抗
CD45抗体(
Clone: 30-F11; eBioscience, San Diego, CA)、FITC 標識または
PE標識抗
F4/80抗体(Clone: BM8; eBioscience) 、および
PE-Cyanine5標識抗
CD11b抗体(
Clone: M1/70; eBioscience)を加え、
4℃・
15分 間インキュベートした。 腎に浸潤した
Mφsの数を解析する検体に対しては、
Ly6Gを高発現する好中球を除外して解析を行う目的で、
PE標識抗
Ly6G抗体(
Clone:1A8-Ly6g; eBioscience)を加え、Mφs
の表現型を解析する検体に対しては、各実
7
験目的に応じ、
PE標識抗
Ly6C抗体(
Clone: HK1.4; eBioscience) 、
PE標識
CD80抗体 (Clone: B7-1; eBioscience) 、
PE標識
CD206抗体 (Clone: MR6F3; eBioscience) 、 および
PE標識
TLR9抗体(
Clone: J15A7; BD Biosciences)をそれぞれ追加し、
同様にインキュベートした。なお、
CD206抗体および
TLR9抗体は、それぞれ細
胞内の
CD206および
TLR9発現の評価に供するため、これらの抗体のインキュ
ベ ー ト 直 前 に 、 細 胞 懸 濁 液 に 対 し 、
BD Cytofix/CytopermTM Fixation/Permeabilization Kit(
BD Biosciences)を用い、細胞膜透過処置を行った。
抗体を反応させた細胞懸濁液は
Novocyte flow cytometer(ACEA Biosciences, San Diego, CA)を用いて
fluorescence-activated cell sorting(
FACS)解析された。なお、
その際に、上記蛍光標識抗体のアイソタイプコントロール抗体を適時置いて使 用した。
(
5)
Mφsの
TNF-α産生能の評価
腎細胞懸濁液を、温度応答性ポリマーを表面に固定化した
96穴ポリエチレン プレート
UpCell(セルシード、東京)に播種し、
in vitroにおいて
CpGモチーフ 含有合成オリゴヌクレオチド(CpG oligodeoxynucleotides:CpG-ODN)20 ng/mL
(
HC4033: TCCATGACGTTCCTGATGCT; Hycult Biotechnology, Uden, Nederland) を
TLR9アゴニストとして添加し、37℃で
4時間刺激培養した。なお、その際、
産生された
TNF-αを
Mφs内に蓄積させる目的で、細胞内のゴルジ体の働きを停 止させる作用のある
monensin(BD Biosciences)も共に添加した。その後、プレート温度を
37℃以下に低下させて細胞を剥離させた後に回収し、上記と同様に、
Fc
ブロッキングに続いて
APC標識抗
CD45抗体(30-F11) 、PE 標識抗
F4/80抗
体(
BM8)および
PE-Cyanine5標識抗
CD11b抗体(
M1/70)を加えインキュベー
トし、細胞膜透過処置を行った後に
FITC標識抗
TNF-α抗体(Clone: MP6-XT22;
8
eBioscience
)でインキュベートした。なお、
MP6-XT22のアイソタイプコントロ
ール抗体として、FITC 標識抗
Rat IgG1 kappa抗体(Clone: eBRG1; eBioscience)
を使用した。
FACSにて、対象の細胞集団において
TNF-αのアイソタイプコント ロールと比較し、TNF-α 陽性と判断された細胞分画の平均蛍光強度(mean
fluorescent intensity:
MFI)を、細胞の
TNF-α産生能として評価した。
(
6)
Mφsの
ROS産生の評価
腎細胞懸濁液に
Fc OxyBURST™ Green Assay Reagent(Thermo Fisher Scientific,Waltham, MA
)を添加し、
37℃で
30分間インキュベートした。本試薬は、
Fc受
容体を介し細胞内に取り込まれ
ROSを検知した際に、蛍光を発することから、
目的の細胞集団における
ROS産生は、
OxyBURST由来の蛍光が陽性である細胞 分画の
MFIまたは、蛍光陽性分画が当該集団に占める割合として
FACSにて解 析された。なお、本試薬を加えない検体の細胞をネガティブコントロールとして 使用し、OxyBURST 添加検体との差分を陽性と評価することで、各細胞が持つ 固有の自家蛍光を排除した。
(
7)
Mφsの貪食能の評価
腎細胞懸濁液を
UpCellに播種し、in vitro で
Fluoresbrite YG Microspheres 0.75 μm(
Polysciences, Warrington, PA)を添加し
37℃で
30分間培養した。対象の細胞 集団の貪食能は、本
FITC標識ビーズを貪食した結果、蛍光陽性となった細胞分 画が当該集団に占める割合として
FACSにて解析された。なお、本検討において も、
FITC標識ビーズを加えない検体をネガティブコントロールとして使用した。
(8) 腎ホモジネート
TNF-αの測定
9
腎臓組織に対し、
1%プロテアーゼ阻害剤カクテル(ナカライテスク、京都)
を添加した
radioimmunoprecipitation(RIPA)バッファー(富士フイルム和光純薬)
を細胞溶解バッファーとして加え、組織を機械的に破砕してタンパク抽出を行 った。抽出された各マウスのタンパクサンプル中の
TNF-α濃度を測定するため、
TNF alpha
マウス
ELISAキット(
BMS607-3; Thermo Fisher Scientific)を用いた。
同時に、タンパクサンプルにおける総タンパク濃度を、Pierce BCA タンパクア ッセイキット(
Thermo Fisher Scientific)を用いて測定し、これで
TNF-α濃度を 除することで、腎臓組織タンパク
1gあたりの
TNF-αレベルを定量した。
(9) 腎臓の組織学的評価
摘出した右腎臓を
4%パラホルムアルデヒド溶液にて固定後に、パラフィンに て包埋し
4 μmの組織切片を作成した。
組織切片は、
Periodic Acid-Schiff(
PAS)染色および
Masson trichrome染色を行 い、光学顕微鏡
BZ-X710(キーエンス、東京)にて観察した。糖尿病性腎症に伴 うメサンギウム基質拡大を評価するため、それぞれのマウスの
PAS染色標本に おいて
40倍拡大下で
10個の糸球体を無作為に選び、正常糸球体は
0点、メサ ンギウム領域が糸球体内に占める割合が
25%までは
1点、
25-50%は
2点、
50-75%は3
点、
75%以上は4点なる糸球体スコアを用い
0-5点で半定量的に評価し
た
4,31,32。また、糖尿病性腎症に伴う腎線維化を評価するため、
Masson trichrome染色標本において、20 倍拡大下で
4視野を無作為に撮影した後に
ImageJ-Fijiを 用いて線維化領域を抽出し、線維化率を算出した
33。
(
10) 糸球体における
Wilms tumor 1陽性細胞の評価
Wilms tumor 1(WT-1)は糸球体ポドサイトの核に発現することから、糸球体
10
における
WT-1陽性細胞数を観察することで、糸球体障害におけるポドサイト残 存の程度を評価することが出来る
7。
パラフィン包埋切片をキシレンで脱パラフィンし、エタノールで親水化の後、
pH 6
クエン酸緩衝液に浸漬し、98℃で
1時間、抗原賦活化を行った。ブロッキ ング・ワン(ナカライテスク)にて非特異的結合阻害を行った後、一次抗体とし て
100倍希釈したウサギ由来抗
WT-1ポリクローナル抗体(SC-192; Santa Cruz
Biotechnology, Dallas, TX)を
4℃にて一晩反応させた。
Dako REAL Peroxidase- Blocking Solution(Agilent, Santa Clara, CA)にて組織中の内因性ペルオキシダーゼ阻害を行った後に、二次抗体としてヒストファインシンプルステインマウス
MAX-PO(R)(ニチレイ、東京)を室温にて 40
分反応させ、DAB+ Substrate
Chromogen System
(
Agilent)を用いジアミノベンジジン染色をした。核染色はマ イヤーヘマトキシリン(富士フイルム和光純薬)を用い、その後、エタノールで 脱水、キシレンにて透徹し、封入した。
各検体において
40倍拡大下で
10個の糸球体を無作為に選び、各糸球体にお ける
WT-1陽性細胞数を数え、その平均値を算出した
7。
(
11) 間質における
alpha smooth muscle actin発現の評価
alpha smooth muscle actin(α-SMA)は線維芽細胞に発現し、腎間質線維化の際
に発現増加がみられることが知られている
34。
パラフィン包埋切片を脱パラフィン、親水化の後、pH 6 クエン酸緩衝液にて
98℃で
1時間、抗原賦活化を行った。非特異的結合阻害後に、一次抗体として
1000倍希釈したウサギ由来抗
α-SMAモノクローナル抗体(Clone: EPR5368;
abcam, Cambridge, UK
)を
4℃にて一晩反応させた。組織中の内因性ペルオキシ
ダーゼ阻害を行い、二次抗体としてヒストファインシンプルステインマウス
11
MAX-PO
(
R) (ニチレイ)を室温にて
40分反応させた後に、ジアミノベンジジ
ン染色をした。核染色はマイヤーヘマトキシリンにて行い、脱水、透徹の後に封 入をした。
各検体において
20倍拡大下で
4視野を無作為に撮影した後に
ImageJ-Fijiを用 いて
α-SMA陽性領域を抽出し、陽性面積率を算出した
33。
(
12) 腎に限局した間歇的低線量放射線照射
腎臓に限局した低線量放射線照射は、
X線照射装置
MBR-1520R-3(HITACHI、
茨城)を使用し、
db/dbマウスに対し
8週齢から開始し
1週間毎に
1回
1 Gyの 照射で
16週齢まで行われた(全
8週間) 。放射線量や照射頻度については、先行 研究を参考にし、放射線照射による全身への悪影響を極小化するべく決定され
た
15,35。放射線照射は、被照射マウスを放射線透過性のプラスチック製円筒容器
内に固定し、その外側から両側の腎臓以外の領域を厚さ
2 mmの鉛板で遮蔽する ように覆う照射ユニットを作成し、本ユニットを照射装置内に静置し、放射線照 射装置を稼働することにより行われた。なお、本
X線照射装置において、放射 線被ばくを防ぐには、厚さ
2 mmの鉛板で十分であることは、装置内において、
同鉛板の遮蔽下では放射線の検知が出来ないことをもって確認した。
(
13) 統計解析
結果は平均±標準誤差で示した。統計学的分析には、JMP Pro バージョン
14(
SAS Institute Japan、東京)を用い、統計学的有意差の検定には、
2群間かつ、
それぞれの群に対応がない場合は
Studentの
t検定を使用し、対応がある場合は
paired t検定を使用した。また、
3群以上の場合は
Tukey-Kramerの
honestly significant difference (HSD)検定を使用した。全ての分析において、有意水準
5%12
以下の場合を有意差ありとした。
13
第
3節 結果
(1) 腎における
2種類の
Mφs集団の同定
まず、一般的なマウス腎の
Mφs分布を同定するために、非糖尿病
mistyマウ スの腎を用いて解析を行った。Mφs やリンパ球を含む細胞集団は、好中球を除 くべく、前方散乱(
forward scatter:
FSC)と側方散乱(
side scatter:
SSC)によ ってゲートされた(図
1A)。ゲートされた細胞集団は、免疫細胞以外の尿細管 細胞等の腎実質細胞を依然として含むことから、それらの細胞を除外し
Mφsと リンパ球のみを解析の対象とするため、さらに汎白血球マーカーである
CD45陽性の細胞集団をゲートした(図
1B) 。同集団を、汎
Mφsマーカーである
F4/80と接着因子である
CD11bによって展開したところ、F4/80
-CD11b-で表現 されるリンパ球に加えて、
F4/80low CD11bhighを示す細胞集団(以下、
CD11bhigh Mφs)およびF4/80high CD11blowを示す細胞集団(以下、CD11b
low Mφs)が同定された(図
1C) 。次に、それぞれにおける、骨髄由来マーカー
Ly6Cの発現を検 討したところ(図
1Dおよび
E)、CD11b
high Mφsについては、陽性率
61.4
±
0.7%であったが、
CD11blow Mφsについては、陽性率
3.0±
0.3%にとどま った(図
1F)。なお、2 型糖尿病モデルである
db/dbマウスにおいても、これら
Mφsにおける
Ly6C発現は同様であった(図
1G) 。以上から、
CD11bhigh Mφsは、骨髄由来(Bone marrow-derived:BM-)Mφs の性質をもち、CD11b
low Mφsは組織固有(
Tissue resident:
Res-)
Mφsの性質をもつと考えた。
(
2) 糖尿病の有無と腎
TNF-αおよび各
Mφs機能の関連
16
週齢の糖尿病モデル
db/dbマウス群と、非糖尿病
mistyマウス群における
比較を行った。腎に浸潤した
CD11bhigh Mφs中の
TLR9発現は、
db/db群におい
て、misty 群より高く(図
2A)、CD11b
low Mφsでも同様であった(図
2B)。腎ホ
14
モジネートの
TNF-αは、
db/db群において、
misty群より著しく高かった (図
2C) 。
CD11blow Mφs中の
ROS産生細胞率も、
db/db群において
misty群より高く(図
2D) 、
CD11blow Mφs中のビーズ貪食細胞率もまた同様に、
db/db群が
misty群よ り高かった(図
2E)。
(3) 糖尿病性腎症の進行に伴う尿アルブミン排泄と
CD11bhigh Mφsの増加
db/dbマウス
5-20週齢と
mistyマウス
8-20週齢を用い、
5・
8・
12・
16・
20週 齢の時点において尿アルブミン排泄量を測定した後に、腎白血球に占める
CD11bhigh Mφsと
CD11blow Mφsの割合を解析した。以後の実験では、これらのマ ウス群を
16週齢以上の加齢群と
12週齢以下の若齢群に分けて、 解析を行った。
尿アルブミン排泄量については、
db/dbマウス加齢群は
db/dbマウス若齢群に比 べ有意に多かったが(図
3A)、misty マウスにおいては、加齢群および若齢群共 に尿アルブミン排泄をほぼ認めなかった。腎
CD11bhigh Mφs(
%)についても、
db/db
マウスにおいては加齢群が若齢群に比べ有意に高かったが、misty マウス
の加齢群と若齢群との間に差を認めなかった(図
3B、
Cおよび
D) 。
db/db
加齢群においては、尿アルブミン排泄量が増加していたことから、糖尿
病性腎症の進行期にあると考えられた。同時に、腎への
CD11bhigh Mφs浸潤の増 加が観察されたことから、以後は、腎症進行と腎
Mφsの関連を検討する目的で、
db/db
加齢群に着目して解析を行い、それと週齢を一致させた
mistyマウスを正
常対照群として使用することにした。
(4) 腎症進行モデルにおける
CD11bhigh Mφsと
CD11blow Mφsの性質
db/db
加齢群において、
CD11bhigh Mφsおよび
CD11blow Mφsにおける
M1マー
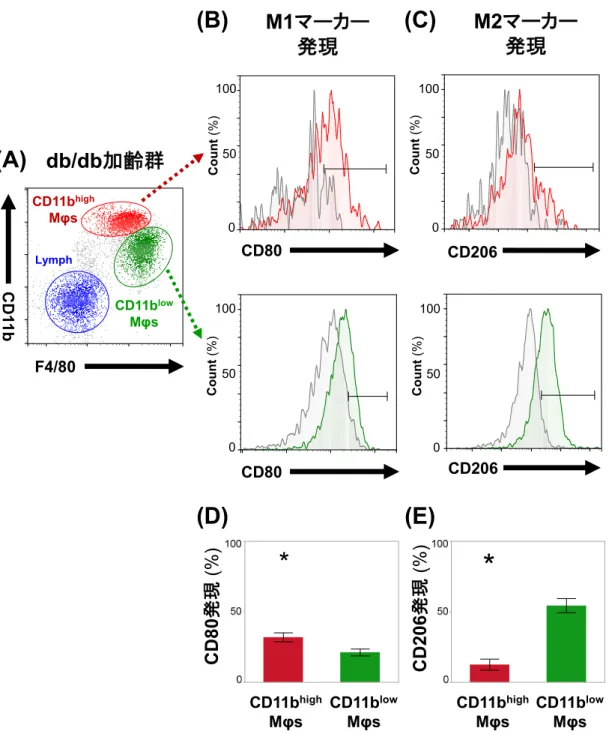
カー(CD80)および
M2マーカー(CD206)の発現を、フローサイトメトリーを
15
用いて検討した(図
4A-E) 。
CD80発現についての
CD11bhigh Mφsおよび
CD11blow Mφsにおける代表的な結果を図
4Bに、
CD206発現についての結果を図
4Cに示 す。
Mφsの
CD80発現は、
CD11bhigh Mφsにおいて、
CD11blow Mφsに比べ有意に 高かったが(38.0±2.8%および
23.3±1.8%)(図 4D)、CD206発現は低かった
(
15.9±
2.8%および
53.8±
3.0%) (図
4E) 。これらの結果は
mistyマウスでも同 様に観察された(データ示さず) 。
(5) 腎症進行モデルにおける
CD11bhigh Mφsと
CD11blow Mφsの機能
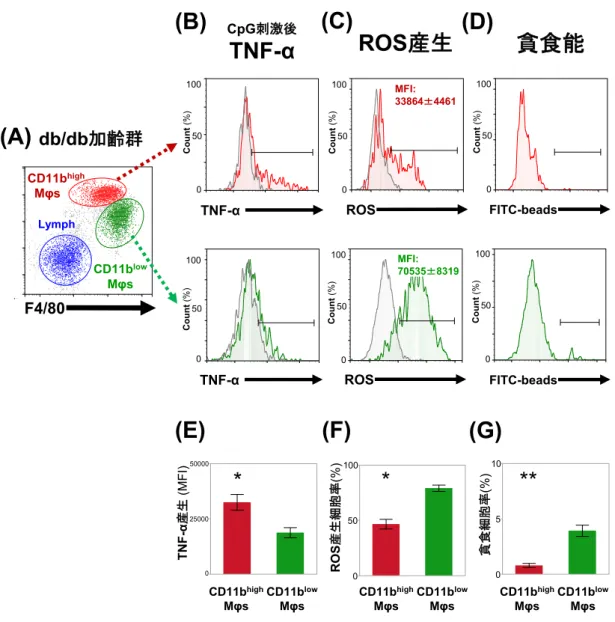
Mφs
の機能として、
TNF-α産生能、
ROS産生および貪食能を評価した。
TNF- α産生能は、 腎
Mφsにおける
TLR9の糖尿病性腎症への関与を検討する目的で、
採取した
Mφsを
TLR9アゴニストである
CpG-ODNとゴルジ体機能阻害薬
(monensin)で培養し、各
Mφs内に
TLR9を介した刺激により産生された
TNF- αを蓄積させ、これを
FACSで測定することで評価した(図
5) 。
db/db
加齢群において、これら
Mφsの機能を
CD11bhigh Mφsと
CD11blow Mφsに分けて解析した結果を示す(図
6A-G) 。
CD11bhigh Mφsと
CD11blow Mφsの
TNF- α産生能、ROS 産生およびビーズ貪食能の代表例を、それぞれ、図
6B、Cおよ び
Dに示している。肝臓の
Mφsと同様に
14,16、TNF-α産生能を示す
MFIは
、CD11bhigh Mφs
において、CD11b
low Mφsに比べ高かった(29406±2388 および
14668±
1036) (図
6E) 。
ROS
産生細胞の
MFIは、CD11b
high Mφsにおいて、CD11b
low Mφsに比べ低く
(
33864±
4461および
70535±
8319) (図
6C) 、さらに細胞集団に占める
ROS産 生細胞の割合も同様に低かった(46.6±4.4%および
79.0±2.8%)(図
6F)。
貪食細胞の割合も同様に、
CD11bhigh Mφsにおいて、
CD11blow Mφsに比べ低か
った(0.78±0.18%および
3.92±0.50%)(図
6G)。
16
(
6) 糖尿病性腎症の進行と腎
Mφsの機能変化
CD11bhigh Mφs
中の
TLR9発現は、db/db 加齢群において若齢群より有意に高 かったが(図
7A) 、
CD11blow Mφs中の
TLR9発現は、
db/db加齢群と若齢群との 間に差は無かった(図
7B)。
CD11bhigh Mφs
の
TNF-α産生能は、
db/db加齢群において若齢群より高かった
(図
7C)。
ROS
産生
CD11blow Mφsの割合は、
db/db加齢群において若齢群より高く(図
7D)、貪食
CD11blow Mφsの割合も同様に、db/db 加齢群が若齢群より高かった
(図
7E) 。
(
7) 糸球体における
Mφsの分布と
TNF-α産生能
db/db
マウス
16週齢の同一個体から得た糸球体ならびに腎全体における
Mφs分布について解析した。糸球体においては、
Mφsのほとんどを
CD11bhigh Mφsが 占めていた(図
8A)。さらに、糸球体由来の
CD11bhigh Mφsの
TNF-α産生能は、
腎全体由来のそれと比べ有意に高かった(図
8Bおよび
C) 。
(
8) 腎に限局した間歇的低線量放射線照射が腎症へ及ぼす効果
腎限局放射線照射は、
db/dbマウスに対し、
8週齢から
16週齢までの間、
1週 間に
1 Gyずつ、計
8週間行われた。その際に作成し使用した遮蔽装置を示す(図
9Aおよび
B)。図
9Aは斜め上方から撮影した装置、図
9Bは横から撮影した装 置の画像である。
尿アルブミン排泄量は、照射群(Irradiation:
Ir群)において、非照射群(Non-
irradiation:
Non-ir群)よりも有意に少なかった(図
9C)。血清クレアチニン(
Cr)
は両群間で差は無く(図
9D)、末梢血白血球数、体重、飲水量、摂餌量や尿量も
17
差を認めなかった(データ示さず) 。しかしながら、腎重量および腎に浸潤した 白血球数は、
Ir群において、
Non-ir群よりも有意に少なかった(図
9Eおよび
F)。 これは、放射線照射が、糖尿病性腎症による炎症を伴う腎腫大を改善した結果と 考えられた。
糸球体病変について、
PAS染色切片を用いて評価したところ、
Ir群において メサンギウム増加・硬化所見の改善(図
10Aおよび
B)ならびに有意な糸球体スコアの低下がみられた(図
10C) 。さらに、糸球体
WT-1陽性細胞数は、
Ir群 において有意に多かった(図
10D-F)。
間質病変について、
Masson trichrome染色切片を用いて評価したところ、
Ir群 において有意に腎線維化が少なかった(図
11A-C)。また、α-SMA の陽性面積率 については、両群間に差は見られなかった(図
11D-F) 。
(
9) 腎に限局した間歇的低線量放射線照射が腎
Mφsへ及ぼす効果
CD11bhigh Mφs
と
CD11blow Mφsの分布が
Non-ir群と
Ir群の間で異なっており
(図
12A) 、その比は
Ir群に有意に低かった(図
12B) 。各
Mφsの片腎あたりの 浸潤数については、Ir 群において、Non-ir 群よりも
CD11bhigh Mφs数が有意に少 なく(図
12C) 、
CD11blow Mφsは少ない傾向にあった(図
12D) 。これは、腎にお いても
CD11bhigh Mφsが放射線感受性であり、
CD11blow Mφsが放射線抵抗性であ ることを示すと考えられた。この現象は、糖尿病である
db/dbマウスほど明らか ではなかったが、非糖尿病マウスである
mistyマウスにおいても観察された(デ ータ示さず) 。
CD11bhigh Mφs
および
CD11blow Mφs中の
TLR9発現は、Ir 群において、Non-ir
群よりも有意に低かった(図
13Aおよび
B) 。さらに、
CD11bhigh Mφsの
TNF-α産生能、腎ホモジネートの
TNF-αや
ROS産生
CD11blow Mφsの割合は、Ir 群に
18
おいて、
Non-ir群よりも明らかに低かった。 (図
13C、
Dおよび
E) 。しかしなが ら、特記すべきことに、貪食
CD11blow Mφsの割合は、Ir 群において、Non-ir 群 よりも明らかに高かった(図
13F) 。
以上の結果をまとめると、表のようになる。
19
第
3章 研究
2糖尿病性腎症マウスモデルにおける
C-Cケモカイン受容体タイプ
2アンタゴニストの 治療効果
第
1節 背景および目的
C-C
ケモカイン受容体タイプ
2(C-C chemokine receptor type 2:CCR2)は単球の遊走を促進させ、慢性炎症性疾患における治療標的であることが示されてい る。腎臓病学の領域においても、この
CCR2のリガンドである単球走化性タン パク質
1(
Monocyte chemotactic protein-1:
MCP-1、別名
CCL2)の中和作用をも つ薬剤
NOX-E36(emapticap pegol)による
MCP-1の阻害が、糖尿病性
apoEノッ クアウトマウスにおいて、炎症性
Mφsの腎への浸潤と分化を抑制し、糖尿病性 腎症の改善効果を示したことが報告されている
36。さらに、
CCR2を直接阻害す る検討として、
CCR2アンタゴニストである
RS102895を
db/dbマウスに投与し たところ、耐糖能の改善とともに、腎臓への
Mφs浸潤の抑制、酸化ストレス減 少による糸球体におけるネフリンの保護、尿蛋白の減少等の腎症改善効果がみ られたことが報告されている
31。これまで、
CCR2アンタゴニストとしては、様々 な物質が報告されているが
6,31,37,38、
CCR2アンタゴニストには、げっ歯類に対す る交叉反応性の欠如という固有の特性が存在することが報告されていることか ら、
CCR2への選択的結合性を高めた薬剤を選択することが、その治療効果を検 討する上で重要である
39。
CCR2
アンタゴニストの一つである、
INCB3344(別名、
15a)は半数阻害濃度
10 nM
でマウス単球への
MCP-1の結合を阻害し、ERK リン酸化や化学走化性と
いった
MCP-1を介した作用に対する用量依存的な阻害効果を示す
39。また、
INCB3344
は
CCR2に対する高い選択性と、げっ歯類での良好なバイオアベイラ
20
ビリティをもち、マウスモデルにおける臓器への
Mφs浸潤の用量依存的な阻害 をもたらすことが報告されている
39。さらに、
INCB3344は、約
5nMの解離定数 である高親和性でヒトの
CCR2にも結合可能であり、その結合は迅速かつ可逆 的であることから、ヒトへの臨床応用も期待可能な薬剤である
40,41。近年では、
INCB3344
が、その長い薬物動態的半減期により、高い
CCR2占有率を維持する
ことで、これを投与された
apoEノックアウトマウスにおいて、循環血中の
CCR2陽性単球数と頚動脈および大動脈起始部のアテロームサイズの減少がみられた ことが報告されている
41。
そこで、
INCB3344を糖尿病マウスモデルに投与し、腎における
Mφsの分布
や機能の変化、それに関連した糖尿病性腎症に対する治療効果が得られるかを
検討した。
21
第
2節 材料と方法
(1) 使用実験動物
(
2) 尿アルブミン排泄量、血清クレアチニン値および随時血糖値の測定
(3) 腎臓の細胞懸濁液の作成
(
4) フローサイトメトリー
(5)
Mφsの
TNF-α産生能の評価
(
6)
Mφsの
ROS産生の評価
(7)
Mφsの貪食能の評価
(
8) 腎臓の組織学的評価
(9) 糸球体における
Wilms tumor 1陽性細胞の評価 以上については、すべて研究
1と同様の手法で行った。
(
10)
INCB3344の投与
Bot
らが検討し得られた結果より、治療効果が見込める投与量として
5 mg/kg体重
/日を設定した
41。マウス投与用の薬液は、
INCB3344を
10%ジメチルスルホ キシド(DMSO)含有生理食塩水に溶解し、1 回の
INCB3344投与量が上記にな り、かつ
1回の投与液量が
10mL/kg体重になるように作成した。治療群
(INCB3344 群)においては、これを週
3回、
8週齢から
16週齢の
8週間、腹腔 内注射し、対照群(
Vehicle群)においては、同量の
10%DMSOを同様に腹腔内 注射した。
(11) 統計解析
研究
1と同様の方法で行った。なお、
mistyマウスにおける
Vehicle群および
INCB3344群、
db/dbマウスにおける
Vehicle群および
INCB3344群の計
4群を対
22
象に解析する際、
misty-INCB3344群と
db/db-Vehicle群との比較については、有
意差を表記しなかった。
23
第
3節 結果
(1)
INCB3344が腎症および全身に及ぼす効果
尿アルブミン排泄量は、
db/dbマウスの
10%DMSO腹腔内投与群(
db/db- Vehicle群)において、misty-Vehicle 群よりも有意に多く、INCB3344 腹腔内投 与群(
db/db-INCB3344群)において、
db/db-Vehicle群よりも有意に少なかった
(図
14A)。血清
Cr値も、db/db-INCB3344 群において、db/db-Vehicle 群よりも 有意に低かった(図
14B) 。随時血糖値、末梢血白血球数および体重は、
mistyおよび
db/dbマウス内の両群間に差は無かった(図
14C、14Dおよび
14E)。
糸球体病変について、
PAS染色切片を用いて評価したところ、
db/db-INCB3344
群において、メサンギウム増加・硬化所見の改善がみられ(図
15Aおよび
B) 、有意に糸球体スコアが低下していた(図
15C) 。さらに、糸球体
WT-1陽性細胞数は、db/db-INCB3344 群において有意に多かった(図
15D-F)。
(2)
INCB3344が腎臓の炎症や
Mφs分布に及ぼす効果
腎重量および腎に浸潤した白血球数は、
mistyおよび
db/dbマウス内の両群間 で有意な差を認めなかったが(図
16Aおよび
B)、CD11b
highMφs数は、db/db-
Vehicle群において
misty-Vehicle群よりも有意に多く、
db/db-INCB3344群にお いて
db/db-Vehicle群よりも有意に少なかった(図
16C)。なお、CD11b
lowMφs数については
mistyおよび
db/dbマウス内の両群間に差を認めなかった(図
16D)。
(3)
INCB3344が腎
Mφsの
M1/M2フェノタイプや機能に及ぼす効果
CD11bhigh Mφs
および
CD11blow Mφsにおける
M1マーカー(
CD80)発現は、
db/db-Vehicle
群において
misty-Vehicle群よりも有意に高く、db/db-INCB3344 群
24
において
db/db-Vehicle群よりも有意に低かった(図
17Aおよび
B) 。さらに、
CD11bhigh Mφs
における
M2マーカー(CD206)発現は
mistyおよび
db/dbマウス 内の両群間に差はみられなかったが(図
17C) 、
CD11blow Mφsにおける
CD206発 現は、db/db-INCB3344 群で有意に高かった(図
17D)。
CD11bhigh Mφs
および
CD11blow Mφsにおける
TLR9発現は、
db/db-Vehicle群に 比べ、db/db-INCB3344 群において有意に低かった(図
18Aおよび
B)。
CD11bhigh Mφs
および
CD11blow Mφsにおける
TNF-α産生能は、
db/db-Vehicle群において
misty-Vehicle群よりも有意に高く、
db/db-INCB3344群において
db/db-Vehicle
群よりも有意に低かった(図
19Aおよび
B) 。さらに、腎ホモジネートの
TNF-α
も同様の結果であった(図
19C)。また、ROS 産生 CD11b
low Mφsの割合 は、
misty-INCB3344群および
db/db-INCB3344群において、各マウス種の
Vehicle群に比べ有意に少なかった(図
19D)。なお、貪食
CD11blow Mφsの割合は
mistyおよび
db/dbマウス内の両群間に差はみられなかった(図
19E) 。
25
第
4章 研究
3糖尿病性腎症マウスモデルにおける
L-カルニチンの治療効果ならびに腎 Mφsに与 える影響
第
1節 背景および目的
糖尿病性腎症マウスモデルにおいては、その病態にミトコンドリア機能異常 が関与し
42、ミトコンドリアに局在し、ミトコンドリア由来
ROSの消去系酵素 として働く
Superoxide dismutase 2(
SOD2)
43のダウンレギュレーションが起こ っていることが報告されている
44-46。同様に、高脂肪食を摂取した肥満マウスモ デルでは、ミトコンドリア機能異常によるミトコンドリア由来の
ROS産生亢進 が起こり、ミトコンドリア内の脂質
β酸化が阻害され、細胞内に脂質が集積し た結果、腎糸球体における血管内皮細胞および足細胞の障害により蛋白尿が誘 発され、尿細管細胞の障害により間質の炎症が生じることが示されている
47。ま た、敗血症マウスモデルにおいては、循環血中のミトコンドリア由来
DNA(mtDNA)が
TLR9活性化を引き起こし、敗血症性腎障害を増悪させるとの報 告もある
25。近年、
L-カルニチン(
L-carnitine:
LC)は透析患者におけるカルニ チン欠乏症や、それに由来する赤血球造血刺激因子製剤抵抗性貧血に対し使用 されるようになってきており、脂肪酸がミトコンドリアに利用される際の担体 として働くとともに、ミトコンドリア保護効果を持つことが知られ
48-50、SOD2 の回復に寄与する可能性も報告されている
51-54。そこで、
L-カルニチンを、肥満 を有する
2型糖尿病マウスモデルに投与し、腎におけるミトコンドリア保護効 果を通じて、糖尿病性腎症に好ましい効果が得られるかを検討した。その際に、
腎
Mφsの分布や機能について影響を及ぼすか併せて検討した。
26
第
2節 材料と方法
(1) 使用実験動物
(
2) 尿アルブミン排泄量、血清クレアチニン値および随時血糖値の測定
(3) 腎臓の細胞懸濁液の作成
(
4) フローサイトメトリー
(5)
Mφsの
TNF-α産生能の評価
(
6)
Mφsの
ROS産生の評価
(7)
Mφsの貪食能の評価
(
8) 腎ホモジネート
TNF-αの測定
(9) 腎臓の組織学的評価
(
10) 糸球体における
Wilms tumor 1陽性細胞の評価 以上については、すべて研究
1と同様の手法で行った。
(11) 腎実質細胞および
Mφsにおけるミトコンドリア由来
ROS産生の評価 腎細胞懸濁液に
MitoSox™ red mitochondrial superoxide indicator(
Thermo FisherScientific)を添加し、37℃で10
分間インキュベートした。本試薬は、細胞内の
ミトコンドリア内に取り込まれ
ROSを検知した際に、蛍光を発する性質を持つ ことから、解析対象の細胞集団のミトコンドリア由来
ROS産生は、当該集団に
おける
MitoSoxの蛍光陽性細胞が集団の全細胞に占める割合(
MitoSox陽性細胞
数/対象集団の全細胞数)として
FACSにて評価された(同時に、対象の細胞群 の
MFIも評価した)。なお、本試薬を加えなかった検体を対照として使用し、
MitoSox
添加検体と比較することで、細胞固有の自家蛍光を排除し、MitoSox に
よる蛍光陽性細胞を同定した。
27
(
12) 近位尿細管細胞におけるミトコンドリア由来
ROS産生の評価
腎細胞懸濁液に
MitoSox™をインキュベートした後に、Fcブロッカー(Clone:
2.4 G2, BD Biosciences
)による非特異的ブロッキングを行い、
APC標識抗
CD45抗体(Clone: 30-F11,eBioscience)および近位尿細管細胞のマーカーである
Lotus tetragonolobus lectin(
LTL; Vector Laboratories, Burlingame, CA)を加え
55、
4℃下 で
15分間インキュベートした。FACS 解析においては、まず非白血球の細胞群 として
CD45陰性の細胞群を同定し、その中で
LTL陽性の細胞集団を近位尿細 管細胞として同定した。その際、
LTLを加えない検体を同時に解析し、細胞固有 の自家蛍光を排除した。近位尿細管細胞中の
MitoSox陽性細胞率を前項の手法 により評価した。
(13) 腎組織における
Superoxide dismutase 2発現の評価
抗
SOD2抗体を用いた免疫組織化学により、腎(特に尿細管)における
SOD2発現を評価した。
まず、パラフィン包埋切片を脱パラフィン、親水化の後、
pH 6クエン酸緩衝 液にて
98℃で 1時間、抗原賦活化を行った。非特異的結合阻害後に、一次抗体 として
1000倍希釈したウサギ由来抗
SOD2ポリクローナル抗体 (
ab13534, abcam)
を
4℃にて一晩反応させた。組織中の内因性ペルオキシダーゼ阻害を行い、二次抗体としてヒストファインシンプルステインマウス
MAX-PO(
R)(ニチレイ)
を室温にて
40分反応させた後に、ジアミノベンジジン染色をした。核染色はマ イヤーヘマトキシリンにて行い、脱水、透徹の後に封入をした。
各切片において
20倍拡大下で
4視野を無作為に撮影し、ImageJ-Fiji を用いて
SOD2陽性領域を抽出し、陽性面積率を算出した
33。
28
(
14)
L-カルニチンの投与
事前検討として
db/dbマウスの飲水量を測定したところ、8 週齢から
16週齢 においては、平均
10 mL/日程度であった。既報から、
L-カルニチン
L-酒石酸塩
(LC;富士フイルム和光純薬)の一日投与量を
25 mg/個体と決定したため56、
同
2.5 g/Lの溶液を調製し、これを飲水として
8週齢から
16週齢までの
8週間
経口投与した(LC 群) 。なお、対照群として通常飲水群(Vehicle 群)を設定し た。
(
15) 統計解析
研究
1と同様の方法で行った。なお、misty マウスにおける
Vehicle群および
LC群、
db/dbマウスにおける
Vehicle群および
LC群の計
4群を対象に解析する
際、misty-LC 群と
db/db-Vehicle群との比較については、有意差を表記しなかっ
た。
29
第
3節 結果
(1) 糖尿病モデルマウスの腎実質細胞および腎
Mφsにおけるミトコンドリア 由来
ROSの解析
腎細胞懸濁液サンプルから、フローサイトメトリーによって、
CD45陰性の細 胞群を同定し、腎実質細胞として解析した。腎実質細胞におけるミトコンドリア 由来
ROS陽性細胞率は、16 週齢の
db/dbマウスで、同週齢の
mistyマウスに比 べ、有意に高かった(図
20A) 。腎
CD11bhigh Mφsにおけるミトコンドリア由来
ROS陽性細胞率は、
db/dbマウスと
mistyマウスの間で差はなかったが (図
20B)、
CD11blow Mφs
については、腎実質細胞と同様に、
db/dbマウス群で有意に高かっ
た(図
20C)。
(2)
L-カルニチンが腎実質細胞およびMφsのミトコンドリア由来
ROSに及ぼ
す効果
LC
を
8週間飲水に混ぜ経口投与した
db/dbマウス群(db/db-LC 群)は、通常
飲水
db/dbマウス群(
db/db-Vehicle群)に比べ、腎実質細胞におけるミトコンド
リア由来
ROS陽性細胞率が有意に低かった(図
21A)。また、腎
CD11bhigh Mφsにおけるミトコンドリア由来
ROS陽性細胞率は、 両群の間で差は無かったが (図
21B)、CD11b
low Mφsでは、db/db-LC 群で有意に低かった(図
21C)。
(3)
L-カルニチンが近位尿細管細胞のミトコンドリアに及ぼす効果近位尿細管細胞(
Proximal tubular cells:
PT cells)中のミトコンドリア由来
ROS陽性細胞率は、
db/db-Vehicle群において
misty-Vehicle群よりも有意に高く、
db/db- LC群において
db/db-Vehicle群よりも有意に低かった(図
22Aおよび
B) 。
さらに、電子顕微鏡にて
db/db-Vehicle群および
db/db-LC群の近位尿細管細胞
30
のミトコンドリア形態を観察したところ、
db/db-Vehicle群において数多くみられ ていたミトコンドリアの膨化やクリステの消失が、db/db-LC 群においては改善 を認めていた(図
22Cおよび
D) 。
(
4)
L-カルニチンが腎
SOD2発現に及ぼす効果
腎切片中の
SOD2陽性面積率は、db/db-LC 群において、db/db-Vehicle 群より 有意に高く、
LCによる
SOD2発現の回復がうかがわれた(図
23A、
Bおよび
C) 。
(
5)
L-カルニチンが腎症へ及ぼす効果
尿アルブミン排泄量は、db/db-Vehicle 群において
misty-Vehicle群よりも有意 に高く、
db/db-LC群において
db/db-Vehicle群よりも有意に低かった(図
24A) 。 血清
Crは群間に差が無かった(図
24B)。なお、腎重量および腎浸潤白血球数 は、
db/db-LC群において有意に少なかった(図
24Cおよび
D) 。これは、
L-カル ニチン投与が糖尿病性腎症による炎症性腎腫大を改善した結果と考えられた。
体重、摂餌量および随時血糖値については、
mistyマウスおよび
db/dbマウス
内の
Vehicleおよび
LC両群間に差を認めなかった(図
25A、Bおよび
C)。
糸球体病変について、
PAS染色切片の評価を行ったところ、
db/db-LC群にお いて糖尿病性腎症進行に関連するメサンギウム増加・硬化所見の改善がみられ
(図
26Aおよび
B) 、糸球体スコアも低下していた(図
26C) 。さらに、糸球体
WT-1陽性細胞数は、db/db-LC 群において有意に多かった(図
26D-F)。
間質病変については、
Masson trichrome染色切片の評価を行ったところ、
db/db- LC群において、有意に腎線維化が少なかった(図
27A-C)。
(6)
L-カルニチンが腎Mφsの分布や機能に及ぼす効果
31
CD11bhigh Mφs
と
CD11blow Mφsの分布も
Vehicle群と
LC群の間で異なってお り、腎白血球に占めるそれらの比は、
db/db-LC群において、
db/db-Vehicle群より も有意に低く(図
28A) 、その理由として、腎
CD11bhigh Mφsの減少が考えられ た。
CD11bhigh Mφs
および
CD11blow Mφsの
TLR9発現は、
db/db-Vehicle群において
misty-Vehicle群よりも高く、db/db-LC 群において
db/db-Vehicle群よりも明らか に低かった(図
28Bおよび
C) 。
CD11bhigh Mφs
の
TNF-α産生能については、
db/db-LC群において
db/db-Vehicle群よりも低い傾向がみられ(図
29A) 、腎ホモジネートの
TNF-αは
db/db-Vehicle群において
misty-Vehicle群よりも高く、db/db-LC 群において
db/db-Vehicle群よ
りも有意に低かった(図
29B) 。
ROS産生
CD11blow Mφsの割合もまた、
db/db- Vehicle群において
misty-Vehicle群よりも高く、
db/db-LC群において
db/db-Vehicle群よりも低かった。 (図
29C) 。しかしながら、特記すべきことに、貪食
CD11blow Mφsの割合は、
db/db-LC群において、
db/db-Vehicle群よりも高かった(図
29D)。32
第
5章 考察
本研究は研究
1、研究2および研究
3から構成されているため、まず研究
1の 考察を述べ、次の研究の考察へと続き、最後に全者に共通する腎
Mφsの糖尿病 性腎症における役割を論じることとする。
研究1. 腎臓における機能に着目した
2種類のマクロファージの同定と糖尿病性腎症 病態との関わりの検討
(1) マウス腎臓における
CD11bhigh Mφsと
CD11blow Mφsの同定、糖尿病性腎症 における
CD11bhigh Mφsと
TLR9の関与
まず、肝臓における
Mφsの分類にならい
12,13、マウスの腎臓において、
F4/80low CD11bhigh Mφs(以下、
CD11bhigh Mφs)および
F4/80high CD11blow Mφs(以下、
CD11blowMφs)からなる 2