低温大気圧プラズマによる
癌細胞傷害性オートファジーの誘導
日本大学大学院医学研究科博士課程 外科系整形外科学専攻
伊藤 友久
修了年 2018 年
指導教員 齋藤 修
目次
概要 ・・・・・・・1
諸言
1. はじめに ・・・・・・・4
2. 骨肉腫 ・・・・・・・5
3. 悪性黒色腫 ・・・・・・・5
4. 低温大気圧プラズマ ・・・・・・・6
5. ミトコンドリアネットワーク ・・・・・・・7 6. 腫瘍壊死因子関連アポトーシス誘発リガンド ・・・・・・・8
7. オートファジー ・・・・・・・9
対象と方法 ・・・・・・・10
結果
1. PSMは特異的にバックグラウンドオートファジーを抑制する ・・・・・・・15
2. 高濃度PSMは特異的にミトファジーを誘発する ・・・・・・・16 3. ミトファジーの誘発は細胞死と相関する ・・・・・・・18
4. PSMは細胞条件によってオートファジー細胞死または非アポトーシス / 非オートファ
ジー細胞死を誘発する ・・・・・・・19
5. TRAILは基本的に細胞保護的オートファジーを誘発する ・・・・・・・20
考察 ・・・・・・・22
まとめ ・・・・・・・27
謝辞 ・・・・・・・28
図表 ・・・・・・・29
引用文献 ・・・・・・・46
研究業績 ・・・・・・・53
概要
背景:骨肉腫や悪性黒色腫は複数の抗癌剤に対して高い耐性を示す、代表的な治療抵抗性
腫瘍である。低温大気圧プラズマは腫瘍細胞を死滅させる一方、正常細胞に対しては影響
が非常に少ないという高い腫瘍選択性を持つことから、有望な癌治療ツールとして注目さ
れ て い る 。 私 の 所 属 研 究 室 で は 細 胞 培 養 液 に 低 温 大 気 圧 プ ラ ズ マ を 照 射 し た cold plasma-stimulated medium ( PSM ) の抗腫瘍効果を利用した新規な治療法の開発を進め ている。以前の報告では、PSMによるヒト悪性黒色腫 ( MM ) ならびに骨肉腫 ( OS ) の 細胞死が、それまでに他の細胞種で報告されていたようなカスパーゼ依存性の典型的なア
ポトーシスとは異なることを明らかにしている ( 1 ) 。今回はその他の細胞死形態として、
オートファジーに注目した。オートファジーはストレス負荷や低栄養などの細胞増殖に好
ましくない状態に対して、細胞内の恒常性を維持するために働く生理的過程であるが、条
件によっては、これを介する細胞死が誘発されること、逆にオートファジーはアポトーシ
スを負に制御しており、種々の癌の薬剤抵抗性に関与することなどが明らかになってきた。
そこで我々は、PSMによる細胞死とオートファジーの関連性を解明するために研究を行っ た。また、同じく腫瘍選択性が高く、アポトーシス細胞死の誘導物質として注目されてい
るTumor necrosis factor ( TNF ) -related apoptosis inducing ligand ( TRAIL;腫瘍壊死因
子関連アポトーシス誘発リガンド ) の抗腫瘍効果におけるオートファジーの関連性につ いても、併せて検討を行った。
目的: PSMの抗腫瘍効果にオートファジーが関与するかどうかを明らかにする。
対象と方法:PSM刺激したヒト骨肉腫、ヒト悪性黒色腫細胞株を用いて、蛍光顕微鏡によ るオートファゴソーム観察、WST-8 法による細胞生存率測定、ウエスタンブロッティング によるオートファジー関連分子の発現解析を行った。また、Tumor necrosis factor ( TNF ) -related apoptosis inducing ligand ( TRAIL;腫瘍壊死因子関連アポトーシス誘発リガン ド ) の作用を併せて検討した。
結果:PSMは細胞傷害性オートファジーを特異的に誘導することで、TRAIL抵抗性のMM やOS細胞に対して強い抗腫瘍作用を示すことを明らかにした。さらに、この癌細胞傷害性 オートファジーではミトファジーの過剰活性化が重要な役割を果たすことが示唆された。
また、PSMは細胞種または増殖条件によってオートファジー細胞死と非アポトーシス非オ ートファジー細胞死の両方を惹起すると考えられた。一方、TRAILは癌細胞において細胞
保護的オートファジーを選択的に誘発するため、それが抵抗性の一因となっていることが
示唆された。
結語:本研究により PSMが特異的に細胞傷害性オートファジーを MM並びにOS細胞に 誘発することで、抗腫瘍効果を示すことが示唆された。
緒言
1. はじめに
骨肉腫や悪性黒色腫は複数の抗癌剤に対して高い耐性があり、治療抵抗性を示す腫瘍と
して代表的である。低温大気圧プラズマは腫瘍細胞を選択的に死滅させ、非腫瘍細胞に対
しては影響を及ぼさない事が報告されており、有望な治療法として注目されている。以前
に私の所属研究室では、低温大気圧プラズマが腫瘍細胞におけるミトコンドリアネットワ
ークを崩壊させることによって、カスパーゼ非依存性細胞死を増加させる一方で、正常細
胞ではミトコンドリア形態異常を生じず、細胞死が誘導されないことを報告している
( 図1 )( 1 )。現時点では低温大気圧プラズマによる抗腫瘍効果のメカニズムはまだ断片的
にしか解明されておらず、今後の臨床応用を目標とするためには、さらなる検討が必要で
ある。今回、私は細胞死のメカニズムとしてオートファジーに着目した。本来細胞内の恒
常性を維持するために必要であるが、多くの癌細胞が生存のためにオートファジーに依存
している事も明らかにされている( 2 )。低温大気圧プラズマがオートファジー機能にど のような影響を与え、細胞死に関連性があるのかを解明するために本研究を行った。
2. 骨肉腫( Osteosarcoma:OS )
骨肉腫は10歳代の若年者に好発する原発性悪性骨腫瘍として最も発生頻度が高いもので あるが、人口数十万人に年間一人の発生をみる稀な癌である。定義としては、類骨・骨を
産生する未分化多形あるいは紡錘形細胞肉腫としての悪性腫瘍である。骨内と骨表面に発
生するもので分けられ、組織学的には骨芽細胞型・軟骨芽細胞型・線維芽細胞型の3つに
分類される。
1960年以前の治療は診断次第直ちに切断術が施行されていたが、結果的に肺転移により 早期死亡の結果となっていた。現在は化学療法とともに画像診断および手術療法が発達し、
その予後は飛躍的に改善され、5年生存率70%以上とされている。しかし、この20年間は 新たな治療法確立には至らず、依然として治療に難渋する腫瘍として位置付けられている。
また、手術法も大きく変換し、腫瘍型人工関節等を含めた患肢温存手術が主流となってき
ている( 3 )。
3. 悪性黒色腫( Malignant Melanoma:MM )
本邦でのメラノーマ罹患率は10万人あたり12人、死亡者は年間600人程度であり、メ ラノーマは本邦では稀な癌である。一方、米国では年間約6万人の患者が発生し、年間9,000
人が死亡しており、白人での発生が多く見られる。腫瘍の厚さが2mm以下でリンパ節転移 のない病期Ⅰであれば、90%以上の高い5年生存率が期待できるが、遠隔転移のある病期
Ⅳの進行期メラノーマの場合には生存率が9−13%と非常に予後不良である。現在は免疫チ ェックポイント阻害薬や分子標的薬を組み合わせた新しい治療法が開発されている( 4 )。
4. 低温大気圧プラズマ
物質には個体・液体・気体の状態が存在し、これは物質の三態と呼ばれ、順に分子間力
による結合が弱くなる。気体にさらにエネルギーを加えた場合、分子の電離によって電子
と陽イオンが生成される。このような、荷電粒子群を含む粒子集団がプラズマと定義され
る。プラズマの人工的な生成のためには、放電が用いられる。放電に際して気体中の電子
が印加電位からエネルギーを受け取り、衝突によって重いイオンや中性粒子へエネルギー
が伝達される。低圧下では分子間の衝突頻度が低くなり、加速を最も受けやすい電子のみ
が高速かつ高温となり、イオン温度やガス温度は高くない非平衡状態プラズマ、すなわち
低温大気圧プラズマとなる。低温大気圧プラズマは被照射物に熱ストレスを与えないため、
生体利用に用いることが可能なる( 5.6 )。
プラズマによる抗腫瘍効果は、活性酸素種( Reactive oxygen species : ROS )や活性窒
素種( Reactive nitrogen species : RNS )による影響が大きいと考えられている( 7.8.9 )。
Medium へプラズマを照射した場合、プラズマジェットの周囲ではプラズマと空気中の酸
素や窒素が反応し、ROS やRNSが発生する( 10 )。それらが、照射されたMedium内 でおそらくは水和した状態であり、その効果は数時間以上の活性を維持する事が確認され
ている( 11 )。また、凍結した場合にはその効果を数日間維持することも可能である。ROS
やRNSによってストレスを受けた癌細胞では、ミトコンドリアなどの細胞小器官の変性や DNA障害等によって細胞死が引き起こされる事が報告されているが、その抗癌活性因子の 分子的実態は完全には明らかとなっていない( 1.12.13 )。膵臓癌・大腸癌・卵巣癌など様々 な細胞株に対して、低温大気圧プラズマによる抗腫瘍細胞効果が報告されており、新たな
抗癌治療ツールとして注目を集めている( 14.15.16 )。
5. ミトコンドリアネットワーク
ミトコンドリアは網状のネットワークを有する細胞小器官であり、分裂と融合を繰り返
しながらバランスが保たれている。ミトコンドリアネットワークは細胞機能とアポトーシ
スに関して非常に重要である。分裂と融合のどちらかが障害された場合も、ミトコンドリ
アと細胞は深刻な機能不全に陥ることとなる。ミトコンドリアの分離は傷ついたミトコン
ドリアをオートファジーで除去することに役立っている。ミトコンドリア分離能が障害さ
れた場合には、広範囲に相互接続されてしまいネットワークが崩壊してしまうため、ミト
コンドリア全体の質のコントロールができなくなってしまう。一方、ミトコンドリアの融
合はミトコンドリアDNAの融合を容易にし、またミトコンドリア機能に必要な代謝産物の 交換を可能としている。これにより、ミトコンドリアは正常な呼吸機能を維持することで
きる( 1 )。
6. 腫瘍壊死因子関連アポトーシス誘発リガンド
【 Tumor necrosis factor -related apoptosis inducing ligand ( TRAIL )】
TRAILは、細胞膜に存在する特異的受容体へ結合することによってアポトーシス誘導シ
グナルを細胞内へと伝達する抗腫瘍性サイトカインである。DR4, DR5といった受容体へ結 合し三量体を形成し、さらに、FADDを介してCaspase-8と結合し複合体となる。Caspase-8 は自己分解によって活性化されアポトーシスを刺激する。アポトーシスは2つの経路によ
って進行し、1つはcaspase-3の活性化を含むextrinsic pathway、もう1つはミトコンド リア膜電位の消失、cytochrome cの放出、Caspase-9の活性化、Caspase-3の活性化を含 むinstrinsic pathway である。
TRAILの効果は腫瘍細胞特異的であり、正常細胞へ影響を及ぼさないことが報告されて いる。しかしながら、耐性を骨肉腫やメラノーマ等の腫瘍では耐性が指摘されており、他
剤の併用による耐性の克服が必要と考えられている( 17 )。
7. オートファジー
オートファジーとは、細胞内のタンパク質やオルガネラを細胞内で消化する機構であり、
栄養飢餓など様々な細胞ストレスによって誘導される。オートファジーが開始すると、脂
質二重膜からなる小胞が形成され、対象を包み込みオートファゴソームを形成する。オー
トファゴソームは、その外膜がリソソームと融合することでオートリソソームを形成し、
その内部に取り込まれた対象は加水分解酵素によって分解される。
近年では、選択性に基づいて様々な基質がオートファゴソームに取り込まれることが明
らかになっている。対象は変性タンパク質や損傷を受けたミトコンドリア、細胞内に侵入
した細菌などを含む( 18 )。オートファジーは本来細胞の恒常性を維持して細胞生存を助 ける機能であるが、条件によってはこれを介する細胞死が誘発されることが報告されてい
る。また、逆にオートファジーはアポトーシスを負に制御しており、種々の癌の薬剤抵抗
性に関与することなどが明らかになってきた( 19. 20. 21 )。
対象と方法
1. 細胞生存率測定
細胞を1×105個/wellで96well培養プレートへ播種し、72時間培養した後にWST−8法 を用いて測定した。Cell counting Kit-8( Dojindo, 熊本 )を加え、1時間インキュベート
した後に 450nm の吸光度測定を行った。吸光度測定にはマイクロプレートリーダー
( ARVO MX、パーキンエルマージャパン ) を用いた。本研究中で行っているWST-8法に
よる実験では全て、同プレート内で 3ウェル毎に同条件の試薬条件を設定し、それら3ウ ェルの細胞生存率の平均としてデータを算出している。
2. CYTO-IDおよびミトコンドリア形態イメージング
蛍光顕微鏡はオールインワン蛍光顕微鏡BZ-X700 ( KEYENCE ) を用いて観察した。
ミトコンドリアは、ミトコンドリアに選択的に局在する色素である Mito Tracker Red CMXRos ( 20 nM ラ イ フテ ク ノ ロ ジ ー ジ ャ パン ) で 染 色 し 、 核 は Hoeckst33258 ( Dojindo ) で 染 色 し て 蛍 光 顕 微 鏡 で 観 察 し た 。 オ ー ト フ ァ ジ ー の 評 価 に は CYTO-ID®Autophagy detection kit ( コスモバイオ株式会社 )を使用した。
細胞を8穴カバースリップ ( アサヒガラス, 東京 ) に1×104個/穴で播種し、24時間被
験試薬とインキュベート後に染色試薬を添加、37℃、5%CO2インキュベーター内、暗所で 1時間反応させ染色し、蛍光顕微鏡で観察とした。
3. 使用試薬
10%ウシ胎児血清( fetal bovine serum ; FBS、シグマアルドリッチ社 ) を含む高濃度グ
ル コ ー ス含 有 ダル ベ ッコ 改 変イ ー グル 培 地 [ Dulbecco’s modified Eagle’s minimal essential medium ; DMEM ( シグマアルドリッチ社 ) ] を材料としてPSMを作成した。
遺伝子組み換えヒト可溶性TRAILはエンゾライフサイエンス社 ( サンディエゴ、 USA ) から購入した。オートファジー阻害剤である Bafilomycin A1 ( Baf ), 3-Methyladenine ( 3-MA ), Chloroquine Diphosphate ( CQ )とオートファジー活性剤である Rapamycin は
それぞれ Tocris 社( ブリストル、イギリス )より購入した。全カスパーゼ阻害薬
z-VAD-FMK はメルクジャパン( 東京 )から購入した。抗酸化剤 MnTBAP はコスモバ
イオ株式会社 ( 東京 ) から購入した。非水溶性試薬は全てジメチルスルホキシドで溶解し、
ハンクス緩衝塩溶液 ( Hank’s balanced salt solution; HBSS, PH7.4 ) で希釈し、ジメチ ルスルホキシドの終濃度0.1%未満で使用した。
4. プラズマ照射
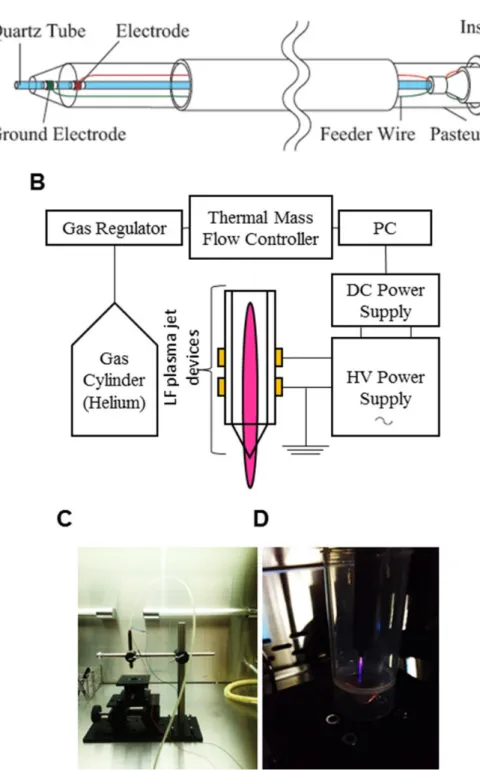
大気圧低温プラズマジェット装置を作成し、発生したAGPをDMEM培地へ照射するこ とで、cold plasma-stimulated medium ( PSM ) として使用した。私の所属研究室から既 報の論文より、使用した装置の模式図および実際の写真を引用にて提示する( 図2 )( 1 )。
ガラス管にヘリウムガスを流し、ガラス管表面の電極に電圧を印加することでプラズマジ
ェットを得ることができ、先端から照射される装置となっている。ガラス管先端はDMEM 培地へ接触しないように1cmほどの距離をおいて設置し、照射を行っている。設定は印加
電圧8kV、ヘリウムガス流量 300ml/min、照射時間 300秒で統一した。PSMを濃度調整
して用いる場合には、プラズマジェット未照射のDMEMとPSMの割合を合わせた上で調 整し、細胞培養を行った。
5. 細胞培養
細胞株は骨肉腫細胞株 ( HOS、143B、 SAOS-2、 MG63 ) と悪性黒色腫細胞株
( A549、A375、A2058 )を使用しており、それぞれヘルスリサーチソリューションバン ク ( 大阪 ) より購入した。細胞はFBSを含むDMEMを用いて、5%CO2インキュベータ
ーにて37℃環境下で培養した。
6. ウエスタンブロッティング
細胞をCa2+-, Mg2+-フリーのPBSで洗浄し、プロテアーゼおよびホスファターゼ阻害
剤カクテル( Sigma-Aldrich )を含む CelLytic lysis buffer ( Sigma-Aldrich ) で, 30分 間振盪して可溶化した。細胞残骸を20,000 x g , 15分間の遠心で除去し、上清を集めて, そ のタンパク質量をBCA protein アッセイ ( Thermo Fisher Scientific ) で測定した. 等量 のタンパク質を 4 – 12% NuPage Bis-Tris acrylamide gels ( Invitrogen ) を用いたポリ
ア ク リ ル ア ミ ド ゲ ル 電 気 泳 動 で 分 離 し 、polyvinylidene difluoride membranes ( Immobilon-P; Millipore, Bedford, MA )に転写した. ブロットを0.05% Tween 20を含む トリス緩衝液 ( TBST; Sigma-Aldrich ) で洗浄し、2% nonfat dry milkを含むTBSTで30
分間ブロッキングした。このブロットを p62 ( MBL ), LC3, GAPDH ( Cell Signaling Technology )に対する一次抗体と4°C でovernight 2% nonfat dry milkを含む TBST 中
でインキュベートした。ブロットを2回洗浄後、HRP標識ヤギ抗ウサギまたは抗マウスIgG 抗体 ( Cell Signaling Technology, Inc. ) と室温で1時間インキュベートした。ブロットを TBST で 3回 洗浄 した 後、enhanced chemiluminescence reagent ( GE Healthcare, Fairfield, CT ) でシグナルを増強し、LAS-4000 camera system ( Fujifilm, Tokyo, Japan ) を用いて可視化した。
7. 統計処理
実験群間の統計的有意性は分散分析および Tukey test で解析し、値は平均±標準偏差で表 記し、p<0.05を有意とした。
結果
1. PSMは特異的にバックグラウンドオートファジーを抑制する
PSMによる細胞死におけるオートファジーの役割について洞察を得るために、PSMの オートファジー調節能を種々の癌細胞で調べた。CYTO-ID Greenは、細胞にトランスフェ クションすることなく、オートファジー小胞を染色できるカチオン型両媒性色素である。
生細胞におけるイメージングの結果、A375ならびにHOS細胞では、通常の細胞培養 ( 10%
FCSを含むDMEMを用いた培養 ) 条件でもかなりの数のCYTO-IDドットが観察された。
このCYTO-IDドットはRapamycin ( 0.5 μM ) で著しく増加する一方、3-methyladenine
( 3-MA, 5 mM ) 処理により完全に消失することから、オートファゴソームを示すことが確
認された( 図3A, B )。PSMによる時間処理は、HOS、 A549細胞を顕著に傷害した。
その結果、接着性の紡錘形細胞は丸くなり、一部は接着性と膜の統合性を失った。これに
付随して、バックグラウンドのCYTO-IDドットは顕著に減少した。一方、正常のHDFで はバックグラウンドのCYTO-IDドットをほとんど認められず、PSM処理による変化もな かった( 図4 )。この結果は、PSMが特異的にバックグラウンドオートファジーを抑制す ることを示す。これに対して、TRAIL処理はCYTO-IDドットを著しく増加させた( 図8 )。 所属研究室では安定した培養状態の得られているHDFを正常細胞として用いており、プラ
ズマがHDFに対しては細胞死を誘導しない点についても既報であり ( 図1 )( 1 )、今回 は正常細胞に関する追加実験は行わなかった。
2. 高濃度PSMは特異的にミトファジーを誘発する
以前の報告にて、PSM刺激が顕著なミトコンドリアの傷害が付随する、ミトコンドリア の過剰な断片化とクラスター形成を誘発することで、非アポトーシス/非オートファジー細 胞死が誘導されることが明らかになっている ( 1 ) 。本研究では非アポトーシス/非オート ファジー細胞死とは別にオートファジー、特にミトファジーによる細胞死の存在を検討し
た。損傷されたミトコンドリアはオートファジーの過程で除去され、このミトファジーと
呼ばれる生理的、細胞保護的な現象はこのオルガネラの品質管理ならびに細胞生存にとっ
て極めて重要である。その一方で、過剰なミトファジーは、神経変性疾患、糖尿病、がん
等の様々な病態の原因となることが示されている。よって、PSMがミトファジーを引き起 こすかどうかを検討した。この目的のために、CYTO-IDで生細胞のオートファゴソームを、
ミトコンドリア局在性色素MitoTracker Red CMXRosでミトコンドリアをそれぞれ染色し て蛍光顕微鏡で観察した。未処理のA375細胞核の周囲にフィラメント状のミトコンドリア が、細胞質に散在したオートファゴソームがそれぞれ観察され、両者の局在は異なってい
た。これに対してPSM処理を施した場合は、ミトコンドリアを著しく断片化・凝集させ、
これらの傷害された丸い細胞では、CYTO-IDドットはクラスターを形成した。さらに、二
つの蛍光をオーバーレイすると、CYTO-IDドットとMitoTrackerシグナルがかなりの部分 で重なり黄色の蛍光が見られ、ミトファジーが誘導された ( 図5A,B )。これ対して、TRAIL
処理では、ミトコンドリアは断片化・凝集せず、ミトファジーは観察されなかった( 図5C )。
さらに、HOS及びA2058細胞株を用いて、ミトコンドリア形態の変化におけるPSM濃 度依存性を観察し調べた。濃度設定として、培養細胞全体に核変性が見られ細胞死が誘導
される100% PSMを高濃度と設定し、それと比較するため、生存細胞が散見し核変性や細
胞死が極端に見られない濃度として25% PSMを低濃度として設定した。高濃度PSM添加 によってミトコンドリアの著しい断片化とクラスター形成に加えて、ミトコンドリアとオ
ートファゴソームの共在が見られた。これに対して、低濃度PSM添加ではバックグラウン
ドのCYTO-IDドットは著名に減少したが、ミトコンドリアの形態には大きな変化は見られ
なかった( 図6A )。注目すべきことに、強力なオートファジー促進剤であるrapamycin はオートファゴソームを増加させ、その一部はミトコンドリアと共存していた( 図6A )。 高解像度の解析から、低濃度PSM処理ではミトコンドリアの短縮化が起こり、高濃度PSM 処理では点状となったミトコンドリアの凝集が観察された ( 図6A,B )。
3. ミトファジーの誘発は細胞死と相関する
WST-8法を用いてA2058並びにHOS細胞株の生存率を測定した。A2058ならびにHOS
細胞株の生存率を測定すると、いずれの細胞でも高濃度PSMでは細胞死が誘導されている が、低濃度PSMでは細胞死は誘導されず ( 図7A,B )、顕微鏡下の観察結果と一致した。
また、HOS細胞株ではPSM25%からPSM濃度を濃くするごとに段階的に細胞死が誘導さ れたが、A2058細胞株ではPSM25%と50%での細胞生存率に差は見られず、100%で強く 細胞死が誘導された。次に、ウエスタンブロッティングによりオートファジー関連分子の
動態を解析した。低濃度PSMは、オートファジー関連アダプターp62分子のタンパク質レ ベルを添加後24から48時間で増加させたが、オートファゴソーム形成に必須なLC3-II レベルは増加させなかった。これに対して高濃度PSMは添加後36から48時間で両者のレ ベルを増加させた( 図7C )。これらの結果は、ミトファジーの誘発は細胞死と相関する ことを示す。
4. PSMは細胞条件によってオートファジー細胞死または非アポトーシス/非オートファ ジー細胞死を誘発する
PSMの抗腫瘍効果におけるオートファジーの役割をさらに調べるために、作用メカニズ
ムの異なる薬理学的オートファジー阻害剤のPSM細胞毒性に対する効果を検討した。
HOS・MG63細胞株では、3-MAならびにBaf両者でPSMによる細胞死が抑制された ( 図
8A,C )。SAOS-2細胞では3-MAでは効果が見られなかったが、Bafで細胞死が抑制された
( 図8B )。これらのオートファジー阻害剤の効果は、PSM濃度が低いほど顕著であった。
その一方、HOS, 143B, A2058細胞を用いた他の実験では、3-MA, Baf, CQのいずれのオー トファジー阻害剤によっても細胞死は抑制されなかった( 図9A,C,E )。また、これらの
細胞死はカスパーゼ阻害剤z-VAD-FMKでは抑制されなかった。一方、HOS,143B細胞株 では、これらの細胞死は抗酸化剤 MnTBAPによって抑制され、この効果もPSM濃度が低 いほど顕著であった( 図9B,D )。これらの結果は、PSMは細胞条件によってオートファ ジー細胞死または非アポトーシス/非オートファジー細胞死を誘発することを示す。しかし ながら、A2058細胞株ではMnTBAPによる細胞死抑制が見られず、細胞株間での差異が 見られた。
5. TRAILは基本的に細胞保護的オートファジーを誘発する
TRAIL処理は癌細胞でCYTO-IDドットを増加させ、オートファジーによる細胞死抑制
が薬剤抵抗性の原因となる可能性が示唆された( 図5C )。よって、顕微鏡およびWST-8 法での観察により、TRAILとオートファジーの関連について検討を行った。結果として
TRAILで72時間処理しても、A2058, SAOS-2, HOS細胞いずれも生存率はほとんど低下
しなかった。それに対して、3-MAおよびCQは濃度依存的に生存率を減少させたが、Baf は効果がなかった ( 図10A-D )。細胞毒性の低い濃度の3-MAを併用するとTRAILによる 細胞死が増強され、HOS細胞ではこの増強効果がz-VAD-FMKによって抑制されたが、
SAOS-2細胞はほとんど抑制されなかった( 図10C,D )。次にTRAILと3-MA併用のオ ートファジーならびにミトコンドリア形態に対する作用を解析した。TRAIL単独はHOS 細胞で、CYTO-IDドットを増加させ、3-MAはそれらを消失させた( 図11 )。また、ミ トコンドリアはTRAILによって過剰融合し、一方3-MAによって分裂し、凝集した。しか し、いずれの場合でもCYTO-IDドットはミトコンドリアと別な位置に観察された。TRAIL と3-MAを併用すると、CYTO-IDドットは大きなクラスターを形成し、ミトコンドリアと 共存した ( 図11 )。これらの結果より、TRAILは細胞保護的なオートファジーを誘発し、
その抑制は細胞種によって異なるモードの細胞死を促進してTRAILの抗腫瘍効果を増強す
ることが示唆される。ただし、3-MA併用による細胞死の増加が実際に相乗的に誘導される ものであるかは、本実験のみでは確実なものではない。
考察
この研究で、我々は、PSMのオートファジーに対する作用をヒトMMおよびOS細胞で 解析した。ウエスタンブロッティング分析の結果、致死的な濃度のPSMがオートファゴソ ーム形成に必須なLC3-IIを増加させたのに対して、非致死的濃度のPSMではそのような 効果は見られなかった ( 図7C )。さらに、このPSMの細胞毒性は薬理学的なオートファ ジー阻害剤である3-MAや Bafにより強く抑制された( 図8 )ことから、このオートフ ァジーが細胞死の要因であると考えられる。ただし、細胞種によりこれらのオートファジ
ー阻害剤の効果が異なったことより、オートファジー過程でのIII型PI3Kやオートファゴ ソームとリソソーム小胞の融合メカニズムに異なる細胞株では相違があるのかもしれない。
特にA2058細胞株においては、他の細胞株と比して異なる反応を呈していることから、株
間での差異を強く伺わせた。また、培養細胞のコンディションによってもPSMへの反応に 誤差が見られるため、本研究における限界の一つと考えられる。
しかしながら、オートファジー細胞死の誘発におけるミトファジーの増加の付随は注目 すべき知見である。当研究室の以前の報告に一致して、このPSMは濃度依存的にミトコン ドリアネットワークの異常を誘発した。細胞毒性を示す濃度のPSMはミトコンドリアの過 剰な断片化とクラスター形成を惹起し、一方、細胞毒性を示さない濃度では中程度の断片
化のみが誘発された( 図5,6 )。CYTO-IDを用いた生細胞イメージングで、MMおよび OS細胞では、栄養十分でストレスのない培養条件下でも一定のCYTO-IDドットが観察さ れたことからバックグラウンドのオートファゴソーム生成( オートファジー活性化 )が 起きていると考えられる ( 図3,4 ) 。このオートファジーは、細胞毒性を示す濃度のPSM によって強く抑制され ( 図5,6 )、オートファジー阻害剤3-MAや CQ単独の長時間処理で 特異的に細胞生存率が著明に低下した( 図9 )ことから、このバックグラウンドのオート ファジーは基本的に、特に低栄養条件下で、細胞保護的に働くと考えられる。注目すべき
ことにCYTO-IDを用いた生細胞イメージングの結果、このPSMに反応して、オートファ
ゴソームの状態ならびに存在位置に顕著な変化が認められた。未刺激の細胞では、オート
ファゴソームは微小な散在した粒子として存在し、その大部分は細胞質のミトコンドリア
とは異なる位置に観察された。一方、PSM処理後では、オートファゴソームはフォーカス を形成して、クラスター化したミトコンドリアと共存した。ミトファジーにおいては、損
傷されたミトコンドリアがオートファゴソームに包含されるため、両者は共存することに
なる。また、所属研究室からの報告では過剰なミトコンドリアの断片化とクラスター形成
が起きる場合には、ミトコンドリアは激しく損傷し、その膜電位ならびに完全性を喪失す
ることが示されている ( 7 )。さらに、ミトコンドリア膜電位の喪失は、ミトファジーの主
要な引き金である ( 22 ) 。これらを考慮すると、本研究で見出された現象はミトファジー である可能性が高い。ミトファジーは、本来損傷されたミトコンドリアを排除して、ミト
コンドリアの品質管理ならびに細胞生存に寄与する、生理的で細胞保護的な仕組みである。
しかし、何らかの異常で、過度のミトファジーが生じると、様々な病理学的状態を引き起
こし、神経変性疾患、糖尿病、癌等の様々な病態の原因となることが明らかにされてきた
( 22. 23 )。したがって、PSM処理によって本来生体防御的に働くミトファジーが過度に活
性化され、細胞傷害的に働いていることが推察される。
また、TRAILはオートファゴソーム形成を増加させたが、その細胞毒性は3-MAで抑制 されずむしろ増強される傾向があった( 図5,11 )ことから、これらの癌細胞において
TRAILは基本的に細胞保護的なオートファジーを促進することが示唆された。この結果は、
これまでの肺、膀胱、大腸直腸、および肝臓の様々な癌細胞種における報告と一致する
( 24.25 )。一方、TRAILは細胞障害性のオートファジーの要因と考えられるミトファジー
をほとんど誘発しなかった( 図11 )。この性質は、細胞保護的オートファジーを主として
活性化することが示されているtemozolomide, epirubicin, sorafenib等の様々な抗癌剤と 同様である( 26.27.28.29 )。興味あることに、バックグラウンドのオートファジーを3-MA で抑制すると、TRAILはミトファジーを増加できるようになり( 図11 )、MMやOS細
胞に対する抗腫瘍効果が増強された事が示唆された( 図10 )が、この増強はアポトーシ スによるものであった。この知見は、ミトファジーがオートファジー細胞死だけではなく、
アポトーシスの調節にも関与することを伺わせた。しかし、TRAILと3-MAの併用のミト コンドリア形態に対する作用は、PSMのそれと全く同一ではないため、増強される細胞死 モードの違いは、このミトコンドリアネットワークに対する作用の相違に起因することも
考えられる。これらの問題について現在さらに研究中である。また、本研究中では検討さ
れていないが、PSMとTRAILを併用した場合、PSMによるオートファジー動態への影響
が、TRAILにおける薬剤抵抗性克服の一助となる可能性もあり、今後の課題と考えられた。
本研究で検討された細胞死経路についてまとめたシェーマを提示する( 図12 )。PSM がミトコンドリアの分裂・凝集を誘導する結果、ミトコンドリアネットワークの崩壊によ
って非アポトーシス/非オートファジー細胞死を起こすことは、所属する研究室論文にて既 報である( 1 )。さらに今回の研究の結果、3-MAやCQなどのオートファジー抑制剤を用 いることで抑制される細胞死、すなわち細胞障害性オートファジーの存在が示唆された。
また、TRAILと3-MAを併用した場合には、TRAIL抵抗性腫瘍における抵抗要素として考
えられる細胞保護的オートファジーが3-MAによって抑制される結果、TRAILによるアポ トーシス細胞死に対する抑制が解除され細胞死が誘導される事が推察された。
将来的な展望として、PSMを抗癌治療として使用する場合には、やはりその投与方法に ついて検討する必要がある。現在まで、プラズマジェットをマウスの皮下腫瘍モデルに対
して直接照射することで抗腫瘍効果を確認するといった報告は散見される( 30.31 )が、
これではより深部に存在する腫瘍に対して適用することはできない。今後新たな方法とし
て、プラズマジェットの照射対象として生理食塩水など有害性のない物質を選択し、生体
投与を可能とすることが必要と考えている。生体への投与経路としては、経静脈的投与・
経口投与・局所投与などが考慮され、水分形態であればこれらの投与が考慮することがで
きる。生体内投与における確実な安全性についても検討されている報告は見られるが、人
体において確実に言及されたものはない。細胞株の状態には左右されるものの、プラズマ
は低濃度でも細胞死を誘導する事は可能なため、確実なTreatment Windowについても検 討が必要と考える ( 32 ) 。
本研究は、PSMがTRAILや他の抗がん剤が誘発できない細胞障害性オートファジーを 強く誘発して、高い薬剤耐性を持つMMおよびOS細胞を効率的に死滅させることを初め て明らかにした。この発見は、現行の治療法では十分な治療効果が得られない腫瘍に対し
て、PSMは代替療法として十分に期待されるものであることを示している。
まとめ
本研究によりPSMが特異的に細胞傷害性オートファジーをMM並びにOS細胞に誘発 することで、抗腫瘍効果を示すことが示唆された。
謝辞
本研究に関し、利益相反はありません。
本研究に際して、研究内容、学会発表、論文のご指導と、様々なご指導を頂きました鈴木
良弘博士( 前生理学分野助教、現一般社団法人プラズマ化学生物学研究所代表理事、英国 王立医学協会フェロー )に深謝いたします。また、研究のご指導、サポートをして頂いた 鈴木美喜研究員に深謝いたします。
本研究に際して、ご指導を賜りました日本大学医学部 整形外科学系 整形外科学分野
斎藤修准教授、徳橋泰明教授 ならびに 日本大学医学部 皮膚科学系 皮膚科分野
落合豊子先生、小野江明日香先生に深謝いたします。
A
B
0 20 40 60 80 100 120
Cell viability (%)
***
***
***
PSM
HDF A375
NT He-gas
PI
FITC-Annexin Ⅴ
図1
Oncotarget 19912
www.impactjournals.com/oncotarget
medium considerably increased annexin V-positive cells in A375 cells, but not in HDFs (Figure 3A). However, the cell death induced by AGP-activated medium was not inhibited by treatment with the caspase-3/7-specific inhibitor z-DEVD-fluo r ome t hyl ket one (FMK) (Figure 3C) or general caspase inhibitor z-VAD-FMK up to 100 μM for 24 h (data not shown). These results indicate that
AGP-activated medium increases caspase-independent cell death in a tumor-specific manner. As reported previously [29], treatment with 100 μM H2O2 resulted in considerable cell death, which was quite resistant to the caspase inhibitors (Figure 3C). In addition, necrostatin up to 100 μM had minimal effects on the cell death triggered by AGP-activated medium or H2O2.
Figure 1: Experimental setup for AGP generation and preparation of AGP-activated medium. (A, B) Schematic diagrams of the AGP jet device (A) and the whole AGP generation system operated in helium gas (B). (C, D) Photos of the AGP jet device (C) and a typical image of AGP generated by DBD of helium gas (D).
図2
図3
A375
Control Rapamycin 3-MA
A
CYTO-IDHoe33342
Supplementary Figure S1
Control Rapamycin 3-MA
HOS
B
CYTO-IDHoe33342
025/12018
図4
B
ControlPSM (100 %)
CYTO-ID
BF MitoTracker Overlay
A
A375
ControlPSM (100 %) 18/1683
CYTO-ID
BF MitoTracker Overlay
Figure 1
114/12786
TRAIL
025/12045
022/11852
022/11964
022/11955
Control
MitoTracker
A375 18/1673
Overlay
25/12045
C
BF CYTO-ID図5
ControlRapamycin3-MA
Cyt-ID MitoTracker Overlay Zoom
HOS
PSM (100 %)PSM (25 %)
Figure 2
A2058
PSM (25%)PSM (100%)Control
A
B
図6
pmTOR P62
GAPDH
PSM 25% PSM 100%
LC3-I/II
C 12 24 36 48 72 (h) C 12 24 36 48 72 (h)
Figure 3
A B
0 20 40 60 80 100 120
A2058
Viability (%)
0 20 40 60 80 100 120
HOS
Viability (%)
143B
D
143B
ControlPSM (100 %)PSM (25 %)
CYTO-ID
BF MitoTracker Overlay
C
図7
✳ ✳
✳
(h)
0 20 40 60 80 100
120 MG63
Viability (%)
0 20 40 60 80 100
120 SAOS-2
Viability (%)
A
B
Figure 4
Viability (%)
C
0 10 20 30 40 50 60 70 80 90
100 HOS
図8
✳ ✳
✳ ✳
✳
0 20 40 60 80 100 120
140 HOS
Viability (%)
0 20 40 60 80 100 120 140
160 HOS
Viability (%)
A B
0 20 40 60 80 100
120 143B
0 20 40 60 80 100 120
140 143B
Viability (%) Viability (%)
C D
0 20 40 60 80 100 120
140 A2058
Viability (%) Viability (%)
0 20 40 60 80 100 120
140 A2058
E F
Figure 5 図9
✳ ✳
✳
✳
0 20 40 60 80 100 120 140 160 180
Control TRAIL 3-MA 1.3 3-MA 2.5 3-MA 5 TRAIL + 3-MA 1.3 TRAIL + 3-MA 2.5 TRAIL + 3-MA 5 CQ 100 CQ 300 TRAIL + CQ 100 TRAIL + CQ 300 Baf 100 Baf 300 TRAIL + Baf 100 TRAIL + Baf 300 A2058
Viability (%)
0 20 40 60 80 100 120
Control TRAIL 3-MA 1.3 3-MA 2.5 3-MA 5 TRAIL + 3-MA 1.3 TRAIL + 3-MA 2.5 TRAIL + 3-MA 5 CQ 100 CQ 300 TRAIL + CQ 100 TRAIL + CQ 300 Baf 100 Baf 300 TRAIL + Baf 100 TRAIL + Baf 300 SAOS-2
Viability (%)Viability (%)
0 20 40 60 80 100 120 140
HOS
Viability (%)
0 20 40 60 80 100 120
SAOS-2
Figure 6
A B
C D
図10
✳ ✳ ✳ ✳
025/12045 ControlTRAIL3-MA
Cyt-ID MitoTracker Overlay Zoom
TRAIL + 3-MA
HOS
Figure 7 図11
PSM
Apoptosis TRAIL
Mitochondrial fragmentation
Apoptotic Cell death
Mitochondrial clustering
Autophagy (toxic) Autophagy
(protective)
Non-apoptotic Non-autophagic death
Autophagic death
apoptosis- resistant cancer cells Inhibition
Flow
Promotion Cell death
Figure 8
TRAIL 3-MA 3-MA
CQ
図12
図1. PSMの腫瘍選択性( 参考文献1より引用 )
(A)各種細胞株をPSM単独培地で培養した場合の細胞生存率。HDFおよびMelanocytes
では有意な細胞死は見られない。A375( 悪性黒色腫 )、A549 ( 肺癌 )、MG63( 骨肉腫 ) 細胞株では有意な細胞死が見られる。***P < 0.001
(B)Annexin ⅤとPropidium iodide染色を用いたフローサイトメトリー。Mediumおよ びPSM単独培地でそれぞれ48時間培養を行った場合、A375細胞株に比してHDF細胞株
ではAnnexin Ⅴ陽性細胞が少ない事がわかる。
図2. 低温大気圧プラズマ照射装置( 参考文献1より引用 )
(A)(B) 低温大気圧プラズマ照射装置の模式図 (C) 実際に実験で使用している装置
(D) ガラス管先端より発生したプラズマをmediumへ向かって照射している
図3. PSMによるバックグラウンドオートファジーの抑制
(A)(B) A375, HOS細胞株ではPSM処理によってバックグラウンドのCYTO-IDドットが 著名に減少した。このCYITO-IDドットはrapamycinにて著しく増加し、3-MAにて完全 に消失した。