九州大学学術情報リポジトリ
Kyushu University Institutional Repository
ラットの外因性高コレステロール血症発症の機作と その改善に関する研究
永尾, 晃治
九州大学農学研究科食糧化学工学専攻
https://doi.org/10.11501/3135075
出版情報:Kyushu University, 1997, 博士(農学), 課程博士
第IV章 ベルオキシソームプロリファレーターによる高コレステロール血症の改差
第l節 コレステロール添加食摂取時の影響
IV
-1-1 紺青ジェンフイブロジルは、 血媛脂質調節作用をもっ薬物として高脂血症患者に対し て欧米で広く月jいられている。 この薬剤には上昇した血祭トリグリセリド濃度の低 下作用[72, 73, 74J、 rfIl疑中HDLコレステロールの上昇作HJ、 LDLコレステロー ルの低下作用などがある[75, 76J。 その作用機作は明らかでないが、 ベルオキシ ソームの活性化作JHを介して作JHすると信じられている。 第II-'Y-:第2節で示したよう に、 ExHCラットはペルオキシソームのβー酸化活性がSDラットと比較して低い。 そ こで、 本実験ではベルオキシソームをジェンフィブロジルで、活性化することが ExHCラットの高コレステロール血症の改善に資するか否かについて検討した。
IV-1-2 実験材料および方法
試薬
ジェンフィブロジルはWarner-Lambert Co. (MI, USA)より入手した。 カルボキ シメチルセルロース[sodium salt, low viscosity Jおよびコレステロールオレエートは Sigma Chemical Co.より購入した。 Triton WR1339はナカライテスクから購入したO fl-14C]オレオイノレーCoA
(57
mCi / mmol)はアマシャムジャパンから購入した。[4-14C]コレステローノレ(60mCi / mmol)はNEN Research Prod uctsから購入した。 グル タチオンはSigma Chemicals Co.から入手した。 23-ノルデオキシコール酸はSteraloids から購入した。 酸性ステロール分析用のカラムは、 AN-600(3%、 Gas chrom Q、
100/120 mesh)をガスクロ工業から購入した。
実験動物および飼料
投与薬物は、 200mgのジェンフィブロジ、ルを10m Lの1.5wt %カルボキシメチルセ
/レロース[sodium salt, low viscosity]ー脱イオン水で乳鉢で練って乳化後、 栄養カテー テルを川いて投与した。
第II章第1釘iとII;J1:ぷに予備飼育後、 ExHCラット及びSD系ラットに、 それぞれジエ ンフイブロジルを0もしくは100 mg
/
kg体重量投与する群を設けた(一群5匹、 計4 群)。 その後、 AIN-76に準じて調製した1%コレステロール食を1411間段取させた。体重の増加に応じて、 3日間隔でジェンフィブロジル投与量の設定をし、 毎日2時に 14日間投与した。 14日月、 投与12時間後の深夜1時半から 2時半の問に断頭により 屠殺し、 肝臓を摘111した。 r(rL消は、 HDLコレステロール濃度を測定後、 凍結した。
肝臓からは、 [内:ちに第II章第1節と同様の方法でミクロソーム!両分を調整した。
分析方法
血清のトリグリセリド、 コレステロール、 リン脂質、 HDLコレステロール濃度は 第II章第四îîとIrTJÆ:ぷに市販の両手京法キットで測定した。 第IIな第1節に準じて肝臓脂質 を抽出後、 化'手法にてトリグリセリド、 コレステロールおよびリン脂質濃度を測定
した。
肝臓のアポA-IV、 アポB、 アポE、 LDLレセプタ一、 コレステロール7αーヒドロキ シラーゼ、 HMG-CoA還元酵素のmRNA量を第11章第1節及び第III章第1節に準じて
測定した。 コレステロール7αーヒドロキシラーゼ活性の測定は、 第II章第1節に準じ て行った。
ミクロソームþllf分の調製
肝臓を5倍容の0.25M煎糖、 20mMβーメルカプトエタノール、 2.5mM EDTA、
75mMニコチンアミドを合んだO.05Mリン酸緩衝液(pH7.4)でホモジナイズした。 核、
ミトコンドリア!I可分を除去後、 ミクロソーム画分を超遠心分離(55P-72、 lゴjJ_、
105,000
Xg、 60分)により調製した。
HMG-CoA還元酵素活性の測定
HMG-CoA還元両手素活性はIdcらの方法[77]に従って行った。 インキュベーシヨ
ン系は10μmolリン般カリウム(pH7.2)、 2μmol2-メルカプトエタノール、 �mol
EDTA、 2μmolNADPHおよび0.05μmolDL-[3-14C] HMG-CoA (0.11μCi)とミクロソー ムを合み、 総容電を100μ!とした。 反応はHMG-CoAとNADPH の添加により開始し、
3TCで30分間インキュベートした。 反応は6N塩酸50μlの添加により停止し、 生成し たメバロン般をラクトン化させるためにさらに370Cで30分間インキュベートした。
変性したタンパク質を2,000X gで5分間遠心分離することにより沈降させた。 上清 100μiについてSilica gel 60 (Merck KGaA , Darmstadt , Germany)を用い、 展開溶媒 をベンゼン:アセトン(1:1,\八)として薄回クロマトグ ラフィーを行った。
回収率
の計算のために、 RS-[2-I�C] !'v1c\'alonic acid lactonc (1000 dpm
/
100 μL)も同様に展開 した。 メバロノラクトン!lÍJ]分は直接計測JHバイアルにかきとり、 シンチレータ- 10mlを加え、 液体シンチレーションカウンターで測定し、 外部様準線源法によりクエンチング、の補正を行った。 HM G-CoA還元酵素活性を生成したメバロン酸 pmoll
min!mg protei
nで表した。 ミクロソームのタンパク質量はLowry法[42Jで測定した。アシルCoAコレステロールアシノレトランスフエラーゼ(ACAT)活性の測定
ACAT活性はJefferyの方法[78Jに従い、[1_14C]オレオイノレ-CoAのコレステロー ルエステル画分への取り込みを指標として測定した。 反応液をあらかじめ370Cで30 分間プレインキュベート後、|司温度で[1_14C]オレオイノレーCoA(O.OlMリン酸カリウ ム緩衝液、 pH6に溶解)を添加することで反応を開始した。 最終的な反応液の組成
(0.2mlのO.lMリン酸緩衝液、 pH7.4)は100μgミクロソームタンパク質、 1mg脂肪酸
フリー牛血清アルブミン、 lmMグルタチオン、 100μM [1-14ClオレオイノレーCoA
(10,000
dpnl /nmol)であった。 15分後に4mIのクロロホルム:メタノール(2:1,
V八)を加えて反応を停止した。 外国性のコレステロールを基質として添加する実験(52 nmol, 0.3% Triton WR1339)も行った。 10μgコレステロールオレエートおよび
[4_14C]コレステロール(20,OOOdpm)をマーカーおよび内部標準としてそれぞれ添加 した。 0.8mlの水を加えて2層に 分離した後、 クロロホルム層を分取し、 コレステロー ルエステル問分をTLCにて分離した。 コレステロールとコレステローノレエステルに
相当するバンドをヨー ド蒸気でnJ祝化した後、 京jぎ取ってシンチレーションバイア ノレに分取し、 シンチレータ一(アクアゾ戸ル2) 10mlを加えて、 液体シンチレーショ
ンカウンターにて放射能活性を測定した。 ミクロソームのタンパク質量はLowry法 [ 42Jで測定したD
糞中の中性ステロールおよび胆汁酸の測定
摂食期間最後の3日間の糞を採取した。 凍結乾燥後、 糞をエタノールを用いて75 OCで加温抽山した。 抽出したステロイドを1.25N NaOHとともに1200Cで6時間オー トクレーブした後、 エーテルをJf1いて中性ステロールを抽出した。 その後、 6N塩酸 を用いて溶液を般性にし、 エーテルを用いて酸性ステロールを拍1[1した。 中性ステ ロールの測定には5αーコレスタン、 胆汁酸の測定には23-ノルデオキシコーノレ酸をそ れぞれ内部原准として川い、 ガスクロマトグラフィにてそれらを定量した[79J。
中性ステロールについてはOV-17を用い、 第II章第3節と同様の方法で行った。 胆汁 酸については、 AN-600をガラスカラム:内径3.0mmx長さ2.0mに充填し、 キャリア ガス:窒素、 カラム温度: 2500C 、 インジェクタ ・ディテクタ温度: 2800C、 ガスク
ロマトグラフ:品作GC-4CMの条件で測定を行った。
統計解析
統計解析は第II章第l節と同様に行った。
IV-1-3
結果体重、 摂食量、 食効率および肝臓重量への影響
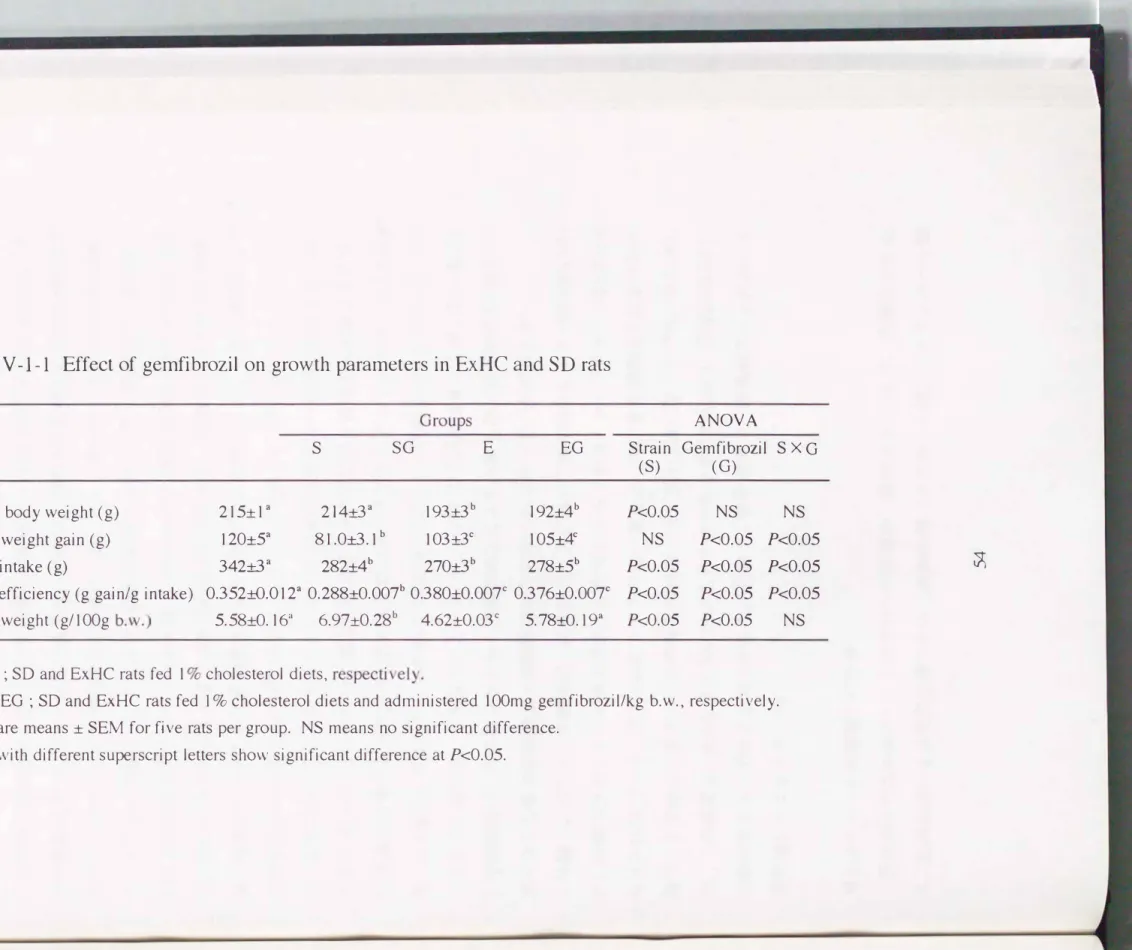
表IV-1-1に示したとおり、 ExHCラットにおいては薬剤投与による成長への影響は 見られなかった。 しかしSDラットにおいては、 薬剤投与により体重増加量、 摂食阜、
食効率ともに低下した。 以前の報告にもある通り、 両系統ラットともに薬剤投与に より肝臓重量が増加した[80J。
S わじ
町一
G
QU
EG
ANOVA
Straín Gemfíbrozíl S X G (S) (G)
Pく0.05 NS NS NS Pく0.05 Pく0.05
Pく0.05 Pく0.05 Pく0.05 訴
Pく0.05 Pく0.05 Pく0.05 Pく0.05 Pく0.05 NS
Table IV-l-l Effect of gemfibrozil on growth parameters in ExHC and SD rats
1 nítíal body weíght (g) 215:t 1 a 214:8 a 193土3b 192:t4b Body weight gain (g) 120:t5a 81.0土3.1b 103:8c 105:t4c
Food íntake (g) 342:8a 282:t4b 270土3b 278土5b
Food effíciency (g gain/g intake) 0.352:tO.012a 0.288:tO.007b 0.380:t0.007c 0.376:t0.007c Liver weight (g/100g b.w.) 5. 58:tO. 163 6.97:t0.28b 4.62:t0.03c 5. 78:tO. 193
S and E ; SD and ExHC rats fed 1 % cholesterol diets, respectively.
SG and EG ; SD and ExHC rats fed 1 % cholesterol diets and administered 100mg gemfibrozil/kg b.w., respectively.
Values are means :t SEM for five rats per group. NS means no significant difference.
Values with different superscript letters show significant difference at Pく0.05.
血清および肝臓脂質濃度への影響
薬剤投与の血清および肝臓脂質濃度への影響は、 表IV-1-2に示した。 両系統ラッ
トの週齢は同 'であったが体重は異なったので、 標準化のために結果は100g体吾、11 たりで示した。 これまでの報告と同様に[11, 12J、 食事コレステロールはExHC ラットにおいて血清コレステロール濃度を顕著に上昇させた。 薬剤投与により非 HDL両分のコレステロール(non HDL-cholesterol)濃度がExHCラットで低下した が、 SDラットでにはそのような影響は見られなかった。 HDLコレステロール濃度 は、 両系統ラットともに薬剤投与により上昇した。 血清トリグリセリド濃度に対す る薬剤投与の彰響は系統間で異なり、 SDラットで低下したがExHCラットで上昇し た。 血清中のトリグリセリドは主に非HDL画分のリボタンパク質に存在するため、
非HDL!両分のコレステロール濃度に対するトリグリセリド濃度の比を取ることで HDL画分以外のリボタンパク質の大まかなトリグリセリド/コレステロール比を求め た。 その結果、 ExHCラットの非HDL画分のリボタンパク質Iflのトリグリセリド/コ レステロール比は、 薬剤投与によりSDラットのものと同程度まで上昇した。 血清リ
ン脂質濃度は、 両系統ラットともに薬剤投与により上昇した。 投与期間を1週間とし た場合でも血清脂質濃度への影響は同じ結果であった(データは示さず)。
肝臓コレステロール濃度は、 両系統ラットにおいて薬剤投与により同程度低下し た。 肝臓トリグリセリド濃度は第II章第2節の結果と同様、 コレステロール摂取によ る上昇はSDラットに比べExHCラットで低い値を示した。 また薬剤投与により両系 統ラットにおいて低下し、 その濃度はSDラットに比べExHCラットで低い値を示し た。 この現象は、 ExHCラットにおけるトリグリセリドの合成低下[第II章第2節]
に起因するものであると考えられた。 肝臓リン脂質濃度は、 両系統ラットにおいて 薬剤投与により上昇した。
糞中ステロール排濯量への影響
薬剤投与の糞rflステロイド、排出t量への影響を、 表IV-1-3に示した。 薬剤投与によ り、 腸内細菌による代謝産物である二次胆汁酸(リソコール酸、 デオキシコール駿
Table IV-1-2 Effect of gemfibrozil on serum and hepatic lipids in ExHC and SD rats
Groups ANOVA
S SG E EG Strain Gemfibrozil S X G
Serum lipids (mgl100g b.w.) Total cholesterol
non HDL-cholesterol HDL-cholesterol Triglyceride
Triglyceride /non HDL cholesterol Phospholi pid
Hepatic lipids (mgl1 OOg b.w.) Total cholesterol
Triglyceride Phospholi pid
(S)
5.49:tO.183 5.36:tO.28a 13.6:t0.9b 7.64:t0.32c Pく0.05 4.20:tO.153 3.45:t0.233 12.9士0.9b 6.07:tO.29c Pく0.05 1. 29:tO. 093 1.91 :t0.09b 0.72:t0.03 c 1.57:tO.06d Pく0.05 15.2:t1.93 9.93士0.79b 7.17土0.57b 17.1 :t1.1 3 NS 16.6:t2.43 12.9土0.33 2.55:tO.25b 12.7:tO.73 Pく0.05 9.90:tO.303 10.4:t:0.5ab 9.60:t0.313 11.3:tO.4b NS
233:t22a 411:t46a 129:t 3a
102土10b 227:t12a 142:t12b NS 160土37b 85.9:t6.8bc 42.5:t4.3 c Pく0.05 192土 8b 102:t 3c 154:t 3d Pく0.05 S and E � SD and ExHC rats fed 1 % cholesterol diets, respectively.
(G)
Pく0.05 Pく0.05 Pく0.05 Pく0.05 Pく0.05 NS
NS Pく0.05 Pく0.05 Pく0.05 Pく0.05 NS
Pく0.05 NS Pく0.05 Pく0.05 Pく0.05 NS
SG and EG � SD and ExHC rats fed 1 % cholesterol diets and administered 100mg gemfibrozil/kg b.w., respectively.
\ VC )
Table IV -1-3 Effect of �emfibrozil on fecal steroids excretion in ExHCand SD rats
。
Neutral sterols (mg/day/lOOg b.w.) Total neutral sterols
Cholesterol Coprostanol
Bile acids (mg/day/100g b.w.) Total bile acids
Chenodeoxycholic acid Cholic acid
ß-muricholic acid
S
31.0土2.0ab 27. 1 ::t2. oab 3.89:tO.66
2.06士0.22a
Groups SG
34.0:t2.2a 31.0:t2.9a 2.98:tl.28
2.45土0.35ab
E
28.0:t1.5b 23.9:t1.3b 4.08:t0.17
3.09:t0.18b
EG
32.7:tl.3ab 29.8:t1.3ab 2.82:tO.38
4.08:t0.37C 0.0348:tO.0051 a 0.159::tO.051 b 0.0111 :tO.0044a 0.302士0.042C 0.0349:tO.0047 0.0266:tO.0126 0.0332:tO.0189 0.0429:tO.0142
0.707:t0.103a 1.71 :t0.37b 0.559:tO.083a 2.90:tO.37C
A NOVA
Strain Gemfibrozil S X G
(S) (G)
NS NS NS
Pく0.05 NS NS NS
Pく0.05 NS Pく0.05 NS
NS NS
Pく0.05 NS Pく0.05 Pく0.05
NS NS
Pく0.05 Pく0.05 Lithocholic acid 0.183::tO.039a 0.0638士0.0138b 0.378:tO.034c 0.0889:t0.0352ab Pく0.05 Pく0.05 Pく0.05 Deoxycholic acid 0.281 ::tO.042a 0.0740:tO.0122b 0.371 :tO.024c 0.0792:tO.0108b NS Pく0.05 NS α村βmuricholic acid 0.379:tO.036ab 0.317:tO.063a 0.514:t0.075ab O. 584::tO. 079b Pく0.05 NS NS hyo+urso deoxycholic acid 0.443::tO.161 a 0.0965:tO.0303a 1.22:t0.21 b 0.0911 :t0.0204a pく0.05 Pく0.05 Pく0.05 Chenodeoxy+Lithocholic acid 0.2 1 8:tO.040a O. 222:tO. 047a 0.389:t0.034 b 0.391 :t0.030b Pく0.05 NS
Cholic+Deoxycholic acid 0.316:tO.040a 0.101 :t0.007b 0.404:t0.037c 0.122:tO.009b NS Pく0.05 Cheno+Litho/Cholic+Deoxycholic 0.206:tO.026a 0.768:!::O.169b 0.338:t0.045a 1.08:tO.07c Pく0.05 Pく0.05
S and E � SD and ExHC rats fed 1 % cholesterol diets, respective ly.
SG and EG : SD and ExHC rats fed 1 % cholesterol diets and administered 100mg gemfibrozil/kg b.w., respectively.
NS NS NS
ト ザ1
およびウノレソデオキシコール酸)の排j世が減少した。 逆に薬剤投与により、 コレス テローノレや一次胆汁酸(ケノデオキシコール酸およびβ-ムリコール酸)の排池が増 加した。 全体的には、 総中性ステロールおよび総胆汁酸の排濯が薬剤投与により増 加した。 SDラットと比較すると、 ExHCラットの胆汁酸の排池量の増加が顕著であっ た。 また薬剤投与により、 ジヒドロキシ胆汁酸であるケノデオキシコール酸とその
次胆汁酸であるリソコール酸の和は両系統ラットとも影響を受けなかったが、 ト リヒドロキシ日li汁酸であるコール酸とその二次胆汁酸であるデオキシコール酸の和 は低下し、 そのため、 ケノデオキシコール酸+リソコール酸/コール酸+デ、オキシコー ノレ酸の比は上昇した。 ジヒドロキシ胆汁酸の量と比は、 SDラットに比べExHCラッ
トで高い値を示した。
コレステローノレ代謝関連酵素活性への影響
薬剤投与のコレステロール代謝関連酵素活性への影響は、 表IV-1-4に示した。
ExHCラットにおいては、 薬剤投与によりコレステロール7α-ヒドロキシラーゼ活 性は上昇したがHMG-CoA還元酵素活性は影響を受けなかった。 対照的にSDラット
においては、 薬剤投与によりHMG
-CoA還元酵素活性は上昇したがコレステロール
7αーヒドロキシラーゼ活性は影響を受けなかった。 ACAT活性は、 測定系へのコレ ステロールの添加・無添加にかかわらず薬剤投与により両系統ラットにおいて低下し、 SDラットに比べてExHCラットで高い値を示した。
コレステロール代謝関連タンパク質およびアボタンパク質のmRNA発現への影響
薬剤投与のコレステロール代謝関連タンパク質およびアボタンパク質のmRNA発 現への影響は、 図IV-l-l、 図IV-1-2および表IV-1-4に示した。 コレステロール7αー ヒドロキシラーゼおよび、HMG-CoA還元酵素のmRNAレベルは、 薬剤投与に対して 酵素活性と同様の傾向で応答した。 LDLレセプタ-mRNAレベルは、 薬剤投与によ り両系統ラットとも影響を受けなかった。 アポBおよびアポA-IVmRNAレベルは、
薬剤投与により阿系統ラットにおいて低下した。 アポBmRNAレベルは、 SDラット に比ベExHCラットで高い値を示した。 アポEmRNAレベルは、 両系統ラットにお
Table IV -1-4 Effect of gemfi brozil on hepatic mRNA abundance, acti vities of cholesterol 7α-hydroxylase,
HMG-CoA reductase and ACAT in ExHC and SD rats
Groups ANOVA
S SG E EG Strain Gemfibrozil S X G
(S) (G)
Proteins
7α-hydroxylase (nmol/hr/mg protein) 1.91:1:0.213 1.73:tO.233 1.65:1:0.243 3 .38:i:Ü.31 b Pく0.05 Pく0.05 Pく0.05 HMG-CoA reductase
(nmol/min/mg protein) 0.0316:t0.0 1313 0.113:tO.0 19bO.0364:i:Ü.013630.0503:i:Ü.0161 3 NS Pく0.05 Pく0.05 ACAT (nmol/min/mg protein)
cholesterol no addi tion 0.505:t0.028!1 0.355:tO.031 b 0.668:i:Ü.029 0.561 :t0.0733(; Pく0.05 Pく0.05 NS cholesterol addi tion 1.02:1:0.043 0.370:t0.033 b 1. 19:tO.07c 0.546:i:Ü.029d P<0.05 P<0.05 NS
mRNAs (arbitrary unit)
7α-hydroxylase 100:t13日 1 06:t 163 133:t 53 213:t45b Pく0.05 NS NS
HMG-CoA reductase 100:t 73 135:t 6b 112:1: 33 107:1: 53 NS Pく0.05 Pく0.05
LD L rece ptor 100:t10 ] 04:1: 6 ]23:t 5 113:t 8 NS NS NS
Apo A-IV ]00土 73 35.8:1:4.0b 79.5:1:1.7c 36.9土1.6b Pく0.05 Pく0.05 Pく0.05
Apo B 100:t 63 89.3:1:4.33 142:t 6b 124:1: 3c Pく0.05 Pく0.05 NS
Apo E 1 00:t 43 87.9:t4.33 130:1: 7b 122:1: 4b Pく0.05 NS NS
S and E � SD and ExHC rats fed 1 % cholesterol diets, respectively.
SG and EG � SD and ExHC rats fed 1 % cholesterol diets and administered 100mg gemfibrozil/kg b.w., respectively.
。 げ\1
ApoA・IV ApoB ApoE
S
10μg5μg
SG
10μg5μg
E
10μg5μg
EG
10flg5μg
Fig.IV-l・1 Slot blot hybridization of hepatic apolipoproteins mRNA
S and E � SD and ExHC rats fed 1 % cholesterol diets, respectively.
SG and EG ; SD and ExHC rats fed 1 % cholesterol diets and administered 100mg gemfibrozillkg body weight (b.\V.), respectively.
Two different amounts of total RNA (10 and 5μg) were blotted for analysis.
4.4Kb -
2.3Kb. 2.0Kb -
Cholesterol 7α-hydroxylase
S SG E EG
司唾・
6.3Kb・
4.4Kb -
2.3Kb - 2.0Kb -
H九1G-CoA reductase S SG E EG
..
6.3Kb -
4.4Kb -
2.3Kb - 2.0Kb -
LDL receptor S SG E EG
Fig.IV-l・2 Northern blot hybridization of hepatic mRNAs S and E ; SD and ExHC rats fed 1 % cholesterol dicts, respectively.
SG and EG ; SD and ExHC rats fcd 1 % cholesterol diets and administered 100mg gemfibrozil/kg body weight, rcspectively.
Cholestcrol 7α-hydroxylasc mRNA of 3.7 Kb band and HMG-CoA reductase mRNA of 5.0 Kb band are indicated by the a汀ow. LDL receptor mRNA of 5.3 Kb band is shown as the single band. The position of marker DNAs with 2.0, 2.3, 4.4 and 6.3 Kb is shown.
いて薬剤投与による影響を受けなかったが、 SDラットに比べExHCラットで高い値 を示した。
V-1-
4Jラ然
以前の報告と一致して、 SDラットにおいて薬剤投与により血清のトリグリセリド 濃度の低下とHDLコレステロール濃度の上昇が見られた[80J。 一方ExHCラット では、 薬剤投与によりHDL阿分以外のコレステロール濃度が低ドし、 HDLコレステ ロール濃度がj二持した。 しかしExHCラットでは、 薬剤投与による血清のトリグリ セリド濃度の低下は見られず、 逆に上昇したo Krauseらは本薬剤を投与したSDラッ トでは、 肝臓からのVLDLの分泌が減少することやリボタンパク質リバーゼ、活性の
上好によるVLDL見化のjt進により、 血清トリグリセリド濃度低下が起こると報告 している[81, 82J。
食事コレステロールは、 ラットJJF臓における脂肪酸やトリグリセリドの合成を増
加させ、 それにイ'i':って肝臓や血清11]にトリグリセリドが蓄積することが報告されて いる[83J。 本実験では、 両系統ラットに対する薬剤投与により食事コレステロー ル誘発J性の肝臓のトリグリセリドの蓄積を抑制していることから、 薬剤投与は食事 コレステロール誘発性の肝臓のトリグリセリド合成を抑制しているものと考えられ た。 よって、 本薬剤はExHCラットのトリグリセリドを合むリボタンパク質の分泌 を抑制しているものと思われるロ 本薬剤の効果についてKrauseらは、 一晩絶食させ た場合、 血祭トリグリセリドは低下しないことも報告している[81J0 ExHCラッ トの血清トリグリセリド濃度に対する本薬剤の役割を理解するためには、 このラッ トのトリグリセリドに富むリボタンパク質の代謝について更なる研究が必要である。
いずれにせよこの薬物は、 表IV-2に示すようにExHCラットの非HDL画分のリボタ ンパク質中のトリグリセリド/コレステロール比をSDラットのものと同程度まで 界させることによって、 この薬剤を投与しないコレステロール負荷ExHCラットで 見られるコレステロールに富むβ-VLDL様の粒子の血中での蓄積[12Jを抑制して いるIIJ能性がある。
本実験において、 本薬剤は糞中への中性ステロール、 特にコレステロールの排池
を増加させた。 ヒトにおいては本薬剤が食事コレステロールの吸収を抑制すること [84J、 ヒト[84JとWistarラット[85Jにおいて本薬剤が糞中へのコレステロー
/レの排池を増加させることが報告されている。 従って本薬剤は食事コレステロール の吸収抑制もしくはコレステロールの排池増加を引き起こしている可能性がある。
今泉らはコレステロールの吸収はSDラットとExHCラットで違いが無かったことを ぶしている[12J0 rfl性ステロールに加え、 薬剤投与により糞中への胆汁酸の排世 が、 特にExHCラットで増加した。 この結果は、 ヒトにおいて薬剤投与により胆汁 酸の分泌が抑制されること[74Jとは異なるが、 Wistarラットに対する本薬剤投与 の結果[85Jとは 』致する。 このような薬剤投与による糞中への中性ステローノレお よび胆汁酸の排前増加が版肉となって、 肝臓でのコレステロール蓄積の顕著な抑制 が起こったものと考えられる。
ACATの活性は肝臓コレステロール濃度によって制御されているので
[86, 87J、 本実験における薬剤投与による本酵素活性の低下の原肉の一部は、 上 述の肝臓へのコレステロールの蓄積の低下によるも のと考えられる。 なお、 薬物の 本酵素の発現や、 活性に対する直接的作用の可能性もある。 薬剤投与による本酵素 活性の低下はSDラットでより顕著であったことから、 コレステロールエステルに富 むリボタンパク質の合成と分泌の抑制に異なった影響をもたらしていると考えられ る。 ひいては、 ExHCラットにおける非HDL画分のトリグリセリド/コレステロール 比にも影響し、 この両分のコレステロール低下の原因の一つになったと考えられる。
薬剤投与による肝臓コレステローノレ濃度の低下は、 しかし、 LDLレセプターの
mR NAの発現には影響を与えなかった。 Matsuokaらも、 本薬剤がウサギの肝臓の
LDLレセプタ-mRNA最には影響しなかったことを示している[88J。 よって、
薬剤投与したラットにおける非HDL画分のコレステロール濃度には、 LDLレセプター を介した取り込みの影響は顕著ではないと思われる。
薬剤処理により7αーヒドロキシラーゼ、の活'性やmRNAレベルの増加がExHCラッ
トにおいて示された。 7α-ヒドロキシラーゼは胆汁酸合成の律速酵素であり、 本酵 素活性は、 高コレステロール食摂食や胆汁酸結合性イオン交換樹脂による胆汁酸プー ルの減少により誘導されることが報告されている[89J。 薬剤投与により肝臓コレ
ステロール濃度はrt1tl系統ラットにおいて低下したことから、 ExHCラットにおける
7αーヒドロキシラーゼの活性やそのmRNAの上昇は、 このラットに対する薬剤の特 別な作用に基づくものであると考えられた。 実際、 薬剤投与による胆汁酸の排前 はSDラットに比べExHCラットで高aい値を示した。 それに加えて、 薬剤投与により
トリヒドロキシ目立汁隊(コール酸とその二次胆汁酸であるデオキシコール酸)に対 するジヒドロキシ胆汁酸(ケノデオキシコール酸とその二次胆汁酸であるリソコー /レ駿)の比がよ白川lし、 その他はSDラットに比べExHCラットで高い値を示した。 ケ ノデオキシコール般を経山して合成されるβームリコール駿[89Jの排前量も、 薬剤 投与によりよ自力11しSDラットに比べExHCラットで高い値を示した。 本薬剤がジヒド ロキシ胆汁般とトリヒドロキ、ンJjhU卜酸の合成の比〉容に影響することはWistarラット でも示されている[85J。 ジヒドロキシ�Il汁酸とトリヒドロキシ1311汁般の分岐点は
12αヒドロキシラーゼが作川する過程に存伝すると考えられることから[89J、 こ の薬剤投与によって7αーヒドロキシラーゼ、以後の段階の胆汁酸の合成系がExHCラッ トでSDラットよりもより活性化を受け、 そのために7αーヒドロキシラーゼの活性 や発現がup regulateされたと考えられた。
7αーヒドロキシラーゼの場合とは対照的に、 薬剤投与によるHMG-CoA還元酵素 の活性とmRNAレベルの増加がSDラットでのみ示された。 これはWistarラットにお ける結果[85Jや、 SDラットにおいてe4C]酢酸からのコレステロールの合成が増加 した我々の結果(未発表)と一致する。 また薬物投与によるSDラットでの本酵素の mRNAレベルの増加はわずかであるので、 HMG-CoA還元酵素やLDLレセプターの mRNA発現を調節しているsterol regulatory element binding protein [90J等の転写
因子の活性化に対する本薬剤の寄与は小さく、 転写以降の過程に本薬物は作用して いるものと考えられた。 いずれにしても本実験では食事にコレステロールが負荷さ れていることから、 本薬剤のHMG-CoA還元酵素への影響が系統によって異なった 理由について明らかにする事は、 ExHCラットのコレステロール代謝を理解する で重要である。
先の報告と同様、 薬剤投与によりアポA-IV mRNA [91JとアポB mRNA [92J のレベルが両系統ラットにおいて低下した。 アポEmRNAが薬剤投与により影響を
受けないことはStaelsらの報告と一致する[92J。 アポBは肝臓におけるトリグリセ リドに富むリボタンパク質の形成の抑制に関与し、 アポA-IVは庶糖食や魚、油を摂食 したラットにおいて肝臓のトリグリセリドの会合や輸送に関係するという報告があ る[93Jが、 薬物投与ExHCラットの血清非HDLコレステロールの低下に対するこ れらアボタンパク質遺伝子の発現の影響については、 更に検討が必要である口
要するに本実験の結果から、 ベルオキシソームプロリファレーターによりExHC ラットにおけるl匂コレステロール血症が抑制されることが示された。 この抑制には、
ベルオキシソームの活性化に基づく、 ACAT活性の抑制やコレステロール7αーヒド ロキシラーゼ、の活性化を介した胆汁酸合成の充進が関与することが示唆された。
V-1-5 小t丙
コレステロールr111:jExHCラットとSDラットのコレステロール代謝に対する、 ベ ルオキシソームプロリファレーターであるジェンフィブロジルの投与の影響を調べ た。 HDLコレステロール濃度は、 両系統ラットともにこの薬剤投与により上昇し、
非HDL岡分のコレステロール濃度はExHCラットでのみ低下した。 血清トリグリセ リド濃度はSDラットで低下したがExHCラットでは上昇した。 そのため、 ExHCラッ トの非HDL画分のトリグリセリド/コレステロール比は、 SDラットのものと同程度
まで上昇した。 総中性ステロールおよび総胆汁酸の排池が薬剤投与により増加し、
SDラットと比較してExHCラットの胆汁酸の排池量は高い値を示した。 薬剤投与に より、 ExHCラットにおいてコレステロール7α-ヒドロキシラーゼ活性が上昇し、
対照的にSDラットにおいてHMG-CoA還元酵素活性が上昇した。 ACAT活性は、 薬 剤投与により両系統ラットにおいて低下し、 SDラットに比ベてExHCラットで高い 値を示した。 コレステロール7αーヒドロキシラーゼおよび、HMG-CoA還元酵素の mRNAレベルは、 酵素活性と同様の傾向で応答した。 LDLレセプタ-mRNAレベル は、 薬剤投与の影響を受けなかった。 アポBおよびアポA-IV mRNAレベルは、 薬剤 投与により両系統ラットにおいて低下した。 アポBとアポEのmRNAレベルはExHC ラットで高く、 アポA-IV mRNAレベルはExHCラットで低かった。 これらの結果は、
ペルオキシソームプロリファレーターに対するコレステロール代謝系の応答の程度
は、 ラットの系統によって異なることを示す。
第2節 核内転写|母子の発現に対する影響
IV
-2-1
出荷1:)第IV章第1 .節にぶしたように、 ジェンフィブロジルはExHCラットの肝臓のコレ ステロール7α-ヒドロキシラーゼの活性を充進することにより、 糞中への胆汁酸の 排池を促進させ、 ひいてはExHCラットの高コレステロール血症の症状を改善した 可能性がある。 ノド両手来のmRNAレベルもExHCラットでは顕著な上封があったこと から、 本両手ぷの発射はこのラットで特異的に誘導された可能性がある。 ジェンフイ ブロジルはベルオキシソームプロリファレーターのー稀であり、 肝臓の多くの遺伝
子の発現に影響を及ぼすという報作がある[94, 95, 96J 0 -般的に、 真核生物の
遺伝子の転写は、 プロモーターのような発現制御に関係する遺伝子配列(制御エレ メント)とその配列に結合する特異的な核内タンパク(転写閃チ)との相互作用に よって制御されている事が知られている[97, 98, 99J。 ベルオキシソームの増殖 によって幾つかの転写ft1チが誘導される事は既に報告されており[100, 101J、 コ レステロール7α-ヒドロキシラーゼのプロモーター領域にその制御エレメントが幾 っか存在する[102, 103, 104J。 本節では、 それら転写因子のうちのNuclear factor
KB (NF-KB)について、 ジェンフィブロジルによるコレステロール7α-ヒドロキシ ラーゼ遺伝子発現の誘導との関係をSDラット及び、ExHCラットを用いて検討した。
IV-2-2
実験材料および方法試薬
T4 Polynucleotide kinaseは宝酒造(東京)、 Sephadex-G50カラムはファルマシア
ジャパン、[y_32PJATP("-'3000Cνmmol)は日本アマシャムから購入した。 一本鎖ム 成オリゴヌクレオチドの合成は、
北海道システム ・サイエンス(株) (北海道)に
依頼した。実験動物および飼料
ラットの削料、 飼育、 ジェンフィブロジルの投与および屠殺の方法は第IV章第1 節と同様に行った。
分析)i法
ベルオキシソームのß-酸化活性の測定は、 第II章第2節に準じて行った。
核内タンパク質のおblff
ラットJl ffll援の妓内タンパク質の抽出は、 基本的にFrainら、 および、Dignamらによ る方法[105‘ 106Jに従って行った。 即ち、 19肝臓、可たり2mlの比率でバッファ-A
(10mM Hepes-KOH緩衝液(pH7.9)、 10mM KCl、 0.15mM spermin、 0.5mM spermidine、 O.lmM EDTA、 0.35M sucrose、 0.5mM DTI、 0.5mM
phenylmethylsulfonyl fluoride、 14 μg / ml aprotinin)を加えて、 テフロンホモジナイ
ザ、ーを用いて90%以上の細胞が破砕されるまでホモジナイズした。 ホモジネートを ナイロンメッシュで糖、過後、 500Xgで10分間遠心分離した。 沈殿を6mlのバッファー Aに懸濁し、 等量のバッファ-B(バッファ-Aのo.35M sucroseを0.5Msucroseに
置換した)に した。 1,500X gで15分間遠心分離後、 沈殿を2mlのバッファーC
(10mM Hepes-KOH緩衝液(pH7.9)、 10mM KCl、 0.15mM spermin、 0.5mM spermidine、 O.lmM EDTA、 0.5mM D1T、 10% glycerol、 0.5mM
phenylmethylsulfonyl fluoride、 14μg/ml aprotinin)で懸濁した。 1,500X gで、10分間遠
心分離後、 沈殿を2倍容のバッファーD(20mM Hepes-KOH緩衝液(pH7.9)、
0.33M NaCl、 1.5mM MgC12、 0.5mM DTI、 0.5mM phenylmethylsulfonyl fluoride、
14μg/ml aprotinin、 100/0 gl ycerol )で懸濁したロ 懸濁液に5M NaCl溶液をゆっくり加 え、 終濃度を0.33Mにした。 ホモジナイズ後氷中に45分間放置し、 8,000Xgで15分間 遠心分離した口 上清中に核内タンパク質を得、 -800Cに保存した。
合成オリゴヌクレオチドの療識
GTG1TGTGATGGGAAGCITC-3')、 およびそのアンチセンス配列(5'-
GAAGC1TCCCATCACAACAC-3')の20mer 高本鎖オリゴヌクレオチドを合成した。
2本鎖オリゴヌクレオチドは、 相補的な2本の1本鎖オリゴヌクレオチドを等量
5mg/mlの濃度で、 10mMトリス塩酸緩衝液(pH7.5)、 50mM NaCl、 10mM MgCl2
を合む溶液中で900C、 1分間加熱後、 1時間室温にて放置アニーリングすること によって調製した。 5pmolの 2本鎖オリゴヌクレオチドを、[y-32P1ATPでT4 Polynucleotide kinaseを用いて5'-末端標識した。 反応溶液はTE緩衝液(10mMトリス塩酸緩衝液 (pH8.0)、 1mM EDTA)で平衡化したセファデックス(Sephadex)G50スピンカ ラム(ファルマシアジャパン)にかけ、 遠心分離 によって溶出した標識化オリゴヌ クレオチドを阿収した。
ゲルシフトアッセイによる核内転写因子の発現測定
5μgの核内タンパク質を、 30mMトリス塩駿緩衝液(pH7.4)、 0.6mM EDTA、
0.6mMD1T、 30mM KCl、 12% glycerol、 3μg 2本鎖ポリ(dI-dC)を合む26μlの反 応液中で250C、 30分間インキュベートした[107, 108J。 またコンペティションアツ セイは、 標識化オリゴヌクレオチドを加えるのと同時に、 標識化していなし、2本鎖 オリゴヌクレオチドを加えてインキュベートを行った。 遊離DNAとDNA-タンパク
質複合体は、 TAE緩衝液(6.7mMトリス、 3.3mM酢酸ナトリウム、 1mM EDTA、
pH7.9)中で、 6%のポリアクリルアミドゲル(13x 13cm)を用いて分離した。 電気 泳動後、 ゲルは滅紙上に転写し、 乾燥後オートラジオグラフィーを行った。
統計解析
統計解析は第11章第1節と同線に行った。
IV-2-3
結果ラットの成長への影響
表V-2-1に示したように、 ジェンフィブロジル投与のラットの成長への影響は、
IV
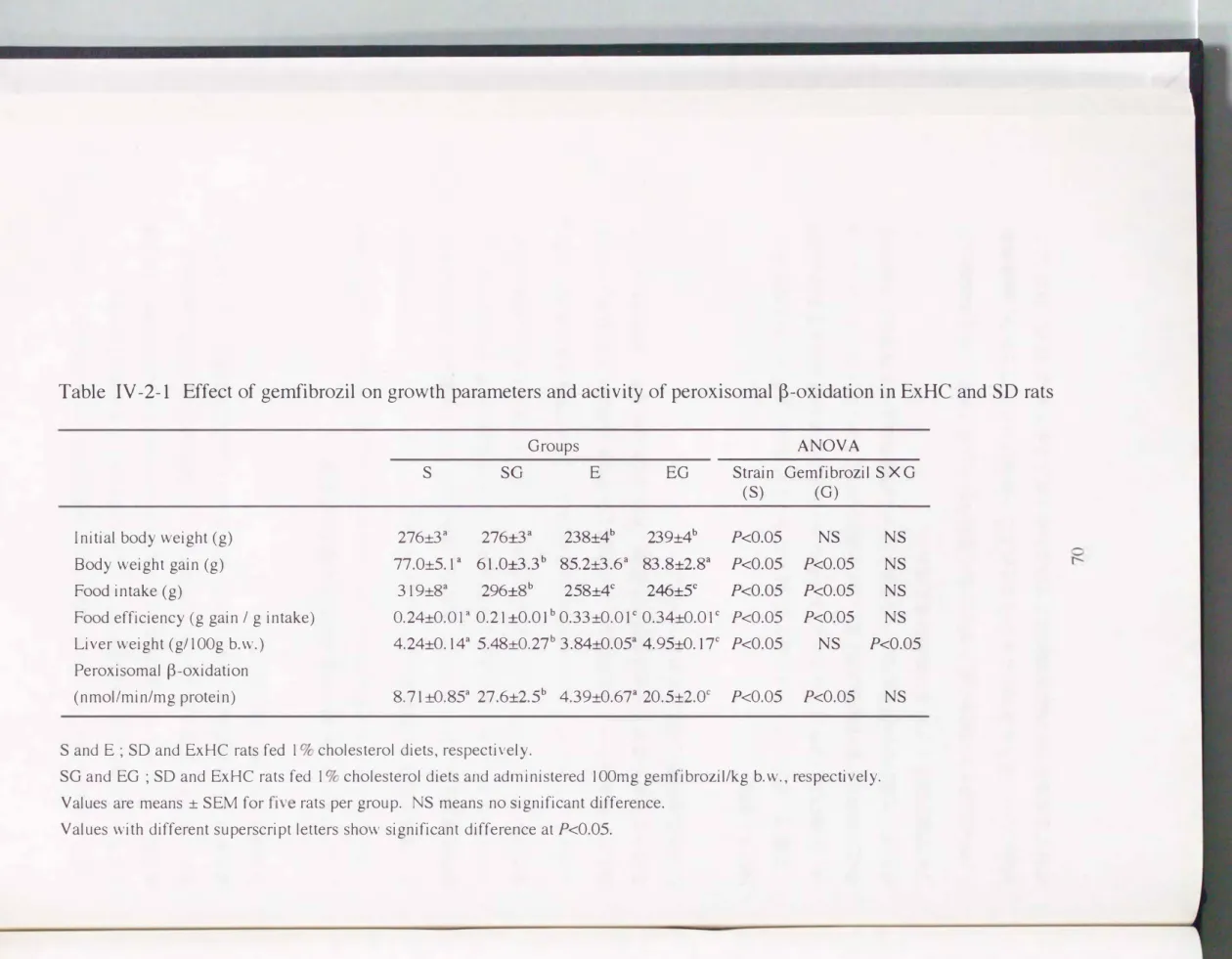
立1節と同様の傾向を示し、 E xHCラットにおいては薬剤投与による成長への影響Table IV -2-1 Effect of gemfibrozil on growth parameters and activity of peroxisomal ß-oxidation in ExHC and SD rats
Groups ANOVA
S SG E EG Strain Gemfibrozil S X G
lnitial body weight (g) Body weight gain (g) Food intake (g)
Food efficiency (g gain / g intake)
(S)
276:t3詞 276:t3a 238土4b 239:t4b Pく0.05 77.0:t5.1 a 61.0:t3.3b 85.2:t3.6a 83.8:t2.8a Pく0.05 319:t8a 296:t8b 258:t4C 246:t5C Pく0.05 O. 24:tO. 01 a 0.21士0.01b 0.33:t0.Ol c 0.34:tO.01 c Pく0.05
(G)
NS NS Pく0.05 NS Pく0.05 NS Pく0.05 NS Liver weight (g1100g b.w.) 4.24:tO.14a 5.48:t0.27b 3.84:t0.05a 4.95:tO.17c Pく0.05 NS Pく0.05 Peroxisomal ß-oxidation
(nmol/min/mg protein) 8.71 ::t:O.85a 27.6:t2.5b 4.39:t0.67a 20.5:t2.0c Pく0.05 Pく0.05
S and E ; SD and ExHC rats fed 1 % cholesterol diets, respectively.
SG and EG ; SD and ExHC rats fed 1 % cholesterol diets and administered 100mg gemfibrozil/kg b.w., respectively.
NS
r ト-,
は見られなかったが、 SDラットにおいては、 薬剤投与により体重増加量、 侵食'T"-e.、
食効率ともに低下した。 両系統ラットともに薬剤投与により肝臓重量が増加した。
両系統ラットにおいて、 薬剤投与により顕著にペルオキシソームの脂肪酸β-酸化活 性が上昇した。 肝臓重量およびベルオキシソームの脂肪酸β-酸化活性の結果より、
薬剤投与により両系統ラットにおいてベルオキシソームの誘導が起こっていること が示唆された[80, 81, 82, 92J。
ジェンフィブロジル投与の核内転写因子発現への影響
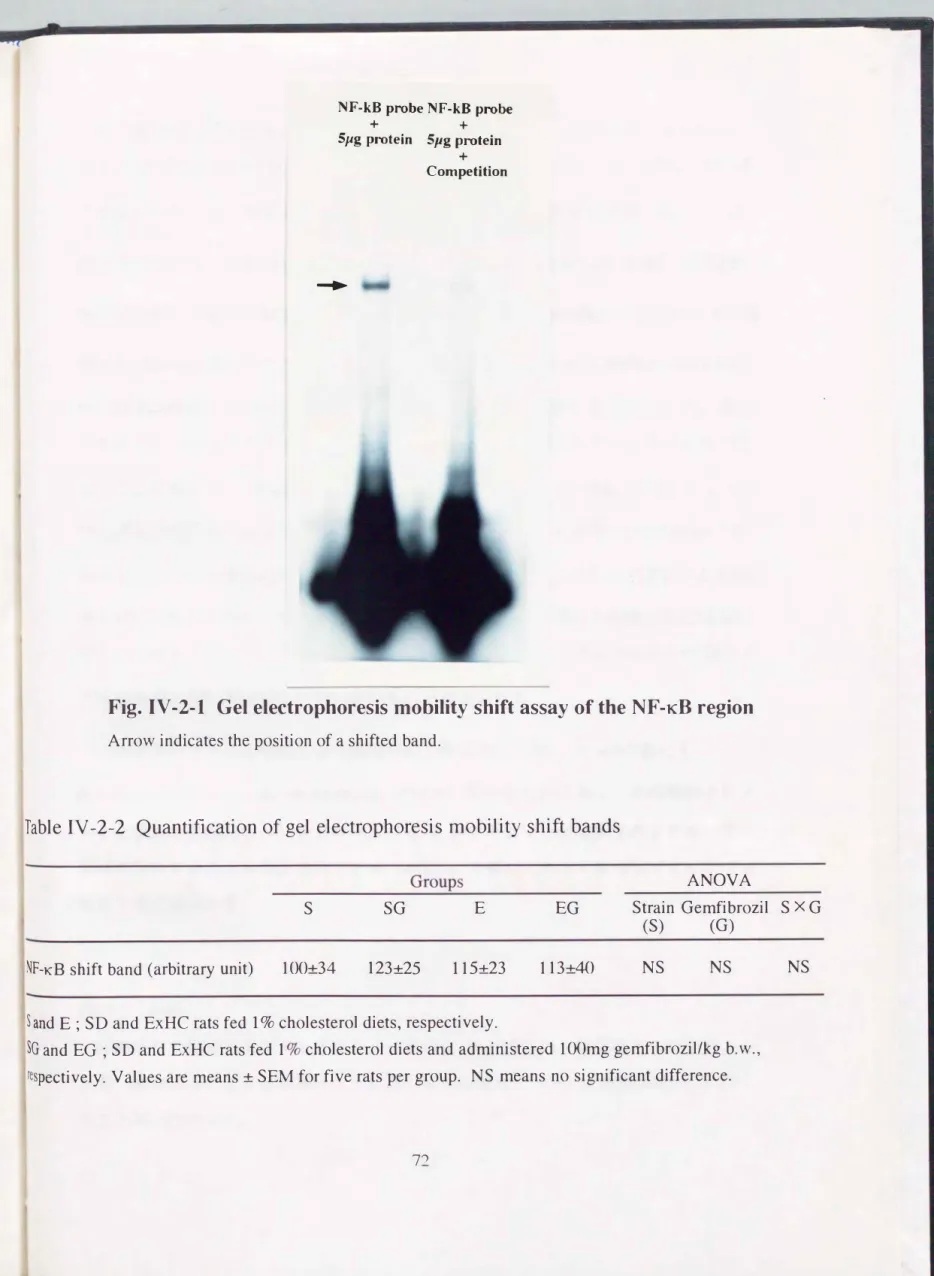
ゲルシフトアッセイ法を用いて、 コレステロール7α-ヒドロキシラーゼ、プロモー
ター領域に結合する核内転写因子NF-KBの発現に対するジェンフィブロジル投与の 影響を検討した。 凶IV-2-1に示した通り、 核内タンパク質と標識オリゴヌクレオチ ドのインキュベーションにより1本のシフトバンドが検出された(左のレーン)。
またコンペティションアッセイによりそのシフトバンドが消失したことから(右の
レーン)、 このシフトバンドがNF-KB由来のDNA-タンパク質複合体であることを確 認した。 両系統ラットの薬剤投与・非投与群それぞれから抽出した核内タンパク質 を用いてゲルシフトアッセイを行った結果、 表IV-2-2に示す通り、 群問でシフトバ ンドの放射活性に差は見られなかった。
IV-2-4 考察
前節で、 ジェンフイブロジルによるExHCラットの高コレステロール血症改善に は、 肝臓のコレステロール7αーヒドロキシラーゼ遺伝子の発現元進よる本酵素活性 の増加が関与する可能性を示した。 この薬剤投与によるコレステロール7αーヒドロ キシラーゼ遺伝子の誘導には、 ベルオキシソームの増殖によって誘導される転写因
子の増加が関与している可能性が考えられた。
NF-KBはBリンパ球中の転写因子として発見されて以来[109J、 多くの異なった 細胞において様々な薬物により活性化されることが報告されたことから、 細胞刺激 に対する重要な包指的転写調節因子ではないかと考えられている[110, 111J。 こ
NF-kB probe NF-kB probe
+ +
5μg protein 5pg protein + Competition
-1・b
Fig.IV-2-1 Gel electrophoresis mobility shift assay of the NF-KB region Arrow indicates the position of a shifted band.
Table IV -2-2 Quantification of gel electrophoresis mobility shift bands
S E
W一
G
CJ
EG
ANOVA
Strain Gemfibrozil S X G
(S) (G)
NF-KB shift band (arbitrary unit) l00:t34 123:t25 115:t23 113:t40 NS NS NS
hnd E ; SD and ExHC rats fed 1 % cholesterol diets, respectively.
�G and EG ; SD and ExHC rats fed 1 % cholesterol diets and administered l00mg gemfibrozil/kg b.w.,
町spectively. Values are means :t SEM for five rats per group. NS means no significant difference.
の転J1/l刈子が、 ペルオキシソームの増殖により誘導されることおよびコレステロー ル7α-ヒドロキシラーゼ、遺伝子のプロモーター領域に制御エレメントを持っている
ことなどから、 NF-KB発現へのジェンフイブロジル投与の影響を検討した。 NF-KB は2つのサブユニット(p50とp65)から成り、 通常はInhibitor Kl3 (IKB)と不活性
別の複合体を形成して細胞質内に存在し、 刺激に応じてIKBが離れることにより活性
別として核内に移行することが知られている。 本実験の結果から核内にNF-KBが存 布する事は雌認されたが、 その誘導は本薬剤投与による影響を受けなかった。 同じ ペルオキシソームプロリファレーターであるciprofibra teで、ベルオキシソームを活性 化することにより、 NF-KBの誘導が起こることが報行されているが[100J、 ジェン フィブロジル投与によりなぜ誘持されなかったのか、 また本薬剤とciprofibrateの作
川の違いについては明らかでない。 いずれにせよ、 NF-KBが何らかの肉子により誘 導を受けて核内に移行していることは示されたが、 その誘導は本薬剤投与に特異的 なものではなく、 よって本薬剤によるコレステロール7αーヒドロキシラーゼ遺伝子 発現の誘導にNF-Kl3は関与していないものと考えられた。
ベルオキシソームの増殖により誘導される転写因子には、 NF-Kl3の他にも
peroxísome proliferator actÍvated rcceptor (PPAR)等が知られており、 その制御エレメ ントと類似した配列がコレステロール7αーヒドロキシラーゼ遺伝子のプロモーター 領域に存在することも報告されている[112J。 今後はこれらの転写因子についても 検討する必要がある。
IV-2-5
小括肝臓重量の増加および、ベルオキシソームのβー酸化活性の上昇が起こっていたこと から、 薬剤投与により両系統ラットにおいてベルオキシソームの増殖が起こってい ることが/J�II愛された。
ゲルシフトアッセイ法を用いて、 コレステロール7α-ヒドロキシラーゼ、プロモー
タ
ー領域に結合する核 内
転写因子NF-KBの発現に対するジェンフィブロジル投与の 影響を検討したところ、 群聞でシフトバンドの放射活性に差が見られず、 ジェンフイ ブロジル投与によるコレステロール7αーヒドロキシラーゼ遺伝子発現の誘導ヘNF-KBの関与は明らかにされなかった。
第V章 高コレステロール血症発症に関与する遺伝子の検索
V-
1系者言
ExHCラットの高コレステロール血症発症には多数の遺伝子が関与していること が推定されている[14] 0 ExHCラットに特異的な発現パターンを示す遺伝子群の 検索を行うことで、 高コレステロール血症発症に関与する疾患遺伝子群を検出する ことが期待できる。 本実験では、 検体問で、のmRNAの発現量の違いを1枚のゲル上 にフィンガープリントすることで区別でき、 稀少な転写産物の検出に適したReverse
transcriptase (RT) -polymerase chain reaction (PCR) differential display (00)法 及びRNA arbitrary primed (RAP) -PCR 00法、 発現強度に顕著に違いがある転写 産物の検出に適しており比較検体問でcONAのハイブリッドを形成させ、 ハイブリツ ドを形成しない検体特異的遺伝子を増幅するPCR-select cONA subtraction法を用い、
対照群としてSDラットの遺伝f発現パターンを比較することで、 高コレステローノレ 血症発症に関与する遺伝子群の検索を行った。
V-2
実験材料および方法 試薬o ligotex -dT30、 T4 Polynucleotide kinaseは宝酒造、 Operon 10-mer kitsはOperon Technologies (CA, USA)、 Random primersはGIBCO BRL (NY, USA)、
SuperTaq DNA polymeraseはサワディ ・テクノロジー(東京)、 Ready to Go First
strand cONA synthesis kit、 dNTPはフアルマシアジャパンから購入した。 ターアンカー プライマー(3'-GTlSG-5')の合成は、 サワディ
・
テクノロジーに依頼した。実験動物および飼料
第II章第1節と同様の食事でSOおよび、ExHCラット を1%コレステロール添加及び、
無添加の純化食で1週間飼育した。 屠殺後直ちに肝臓を摘出し、 液体窒素で凍結後- 800Cに保存した。
RT-PCR differentiaJ dispJay法による疾患遺伝子群の検索
RT-PCR differential displayはItoらの方法を[113J Fukumaki (九州大学医学部)
らが改変したものに従った。 jJ法の概略は凶V-lに示した。 肝臓の総RNAは第II章 第1節と|司伎な方法により調整した。 総RNA500μg から0 ligotex-dT30を用いて、 Poly (A)+mRNAを調製した。 Poly (A)+ mRNA 2μgと3'-アンカープライマ-5μgを用い、
First strand cDNA synthesis kitによりcDNAを合成した。 アンカープライマーのラベ
リングは、 50pmolのGTlSGを[y_32PJATPでT4 Polynucleotide kinaseを用いて5'-末端標 識した(総屯50μ1)0 First strand cDNA 2μl 、 標識アンカープライマー1μl、 18μM イ壬意プライマーlμi、 2.5mM dNTP 1.6μl、 10x PCRバッファ-2μl 、 TaqDNA
polymerase 1 U (総量20μ1)を、 サーマルサイクラー(ASTEC PC-800、 アステック、
福岡)により940C-3分を l サイクル、 950C-15秒、 400C-2分、 720C-1分を 2 5 サイク ル、 720C-5分を1サイクルのプログラムで明幅させた。 増幅産物を6%アクリルアミ ドゲルにて2000V, 5時間泳動した後、 ゲノレを750Cで乾燥しX線フィルムに感光させ た。 x線フィルムにオートラジオグラフィーされたフインガープリントから目的の バンドを選別し、 ゲルからそのバンドを切り出して増幅産物を抽出した。
RAP-PCR differential display法による疾患遺伝子群の検索
方法の原理は図V-1に示した[114}。 肝臓の総RNAは第II章第1節と同様な方法 により調整した。 総RNA
500μgから01igotex-dT30を用いて、 Poly (A)+ mRNAを調
製した。 Poly (A) + mRNA2μgと6merのランダムプライマ-5μgを用い、 First strand cDNA synthesis kitによりcDNAを合成した。 First strand cDNA 1.5μl、 2種類の任意ー プライマーをそれぞれ50 pmol、 lOmM dNTP4μi、 10XPCRバッファ-5μi 、 Taq DNA polymerase 4U (総量50μ1)を、 サーマルサイクラー(ASTEC
PC-800)によ
り940C-25秒、 320C-1分、 720C-30秒を45サイクル、 720C-5分を1サイクルのプログラ ムで増幅させた。 増幅産物を2%アガロースゲルにて泳動した。 ゲルを1μg/mlのエチ ジウムブロマイドで染色し、 UV照射して目的のバンドを選別し、 ゲルからそのバ ンドを切り出して増幅産物を抽出した。RT-PCR DD
RNA CAAA...AAA
4吋婿…
anchor prime町r
First strand synthesis
RNA CAAA...AAA
cDNA GTTT....TTTG
Second strand synthesis random primer
ACGTACG…川合併
cDNA GTTT.…TTTG
�
PCR amplification ACGTACGGTTT....TTTG
RAP圃PCR DD
RNA AAAAA
倒…V///.NNNNNN
random hexamer primer
+
First strand synthesisRNA AAAAA
cDNA - NNNNNN
+
Second strand s川esis random primerACGTACG……働。
cDNA NNNNNN
nH O &E・‘
a c,・・E 0・
m a Rc p・
EE台曹v
ACGTACG
NNNNNN
Fig.V-l Schematic diagram ofthe RAP-PCR and RT・PCR differential display technique
PCR-select cDNA subtraction法による疾患遺伝子群の検索
PCR-select cDNA subtraction kit (Clonetech Lab. Inc., CA, USA)を用いて行い、
方法の原理は|ヌlV-2に示した[115J。 肝臓の総RNAは第II章第1節と同様な方法に より調整した。 総RNA500μgからOligotex-dT30を川いて、 Poly (A)+ mRNAを調製
した。 Poly(A)+ mRNA 2μgにlOpmol cDNA synthesis primer (ターTTTTGTA CA GCTT30- 3 ,)を加え、 逆転写反応によりFirst-strandcDNAを合成した。 その後DNApolymerasc およびDNA ligascをJHいて、 てイヌ鎖cDNAを合成した。 合成した二本鎖cDNAのうち、_-}iの検体(テスター)については半分にわけ、 異なったアダプタ一(アダプタ-
1、 2)をそれぞれ付加した(テスター1、 2)口 対照検体(ドライバ一)にはアダプ ターは付加しない。 DNAの彼雑j交を低くするためにドライバーを制限酵素(RsaI) により断片化し、 ドライバーとテスター1、 ドライパーとテスター2を680C8時間イン キュベートすることにより、 ハイブリッド形成させた。 両サンプノレを一つにまとめ、
新たにドライバーを加えてアニーリング(980C2分)し、 680Cで一晩インキュベート した。 最初のインキュベーションにおいて、 ドライバーと共通配列を持たないテス ターに特異的強発現を示すcDNA配列はハイブリッドを形成せず、 2回目のインキュ ベーションにおいて、 テスター特異的配列を持ち異なったアダプターを持った cDNA 同士がハイブリッドを形成した。 テスター特異的cDNA配列は、 アダプタ-1 および2と相同配列を持つプライマー(プライマ-1 : 5'-CTAATACGACTCACTATA GGGC-3'-.
2:
5' -TGT AGCGTGAAGACGACAGAA-3,)を付加し、 940C-30秒、 680C- 30秒、 720C-1.5分を27サイクルのプログラムで、 サーマルサイクラー(ASTEC PC- 800)によりPCRにかけることで増幅された口 その後、 増幅産物を2%アガロースゲ ノレにて泳動し、 ゲルからそのバンドを切り出して増幅産物を抽出した。
V-3
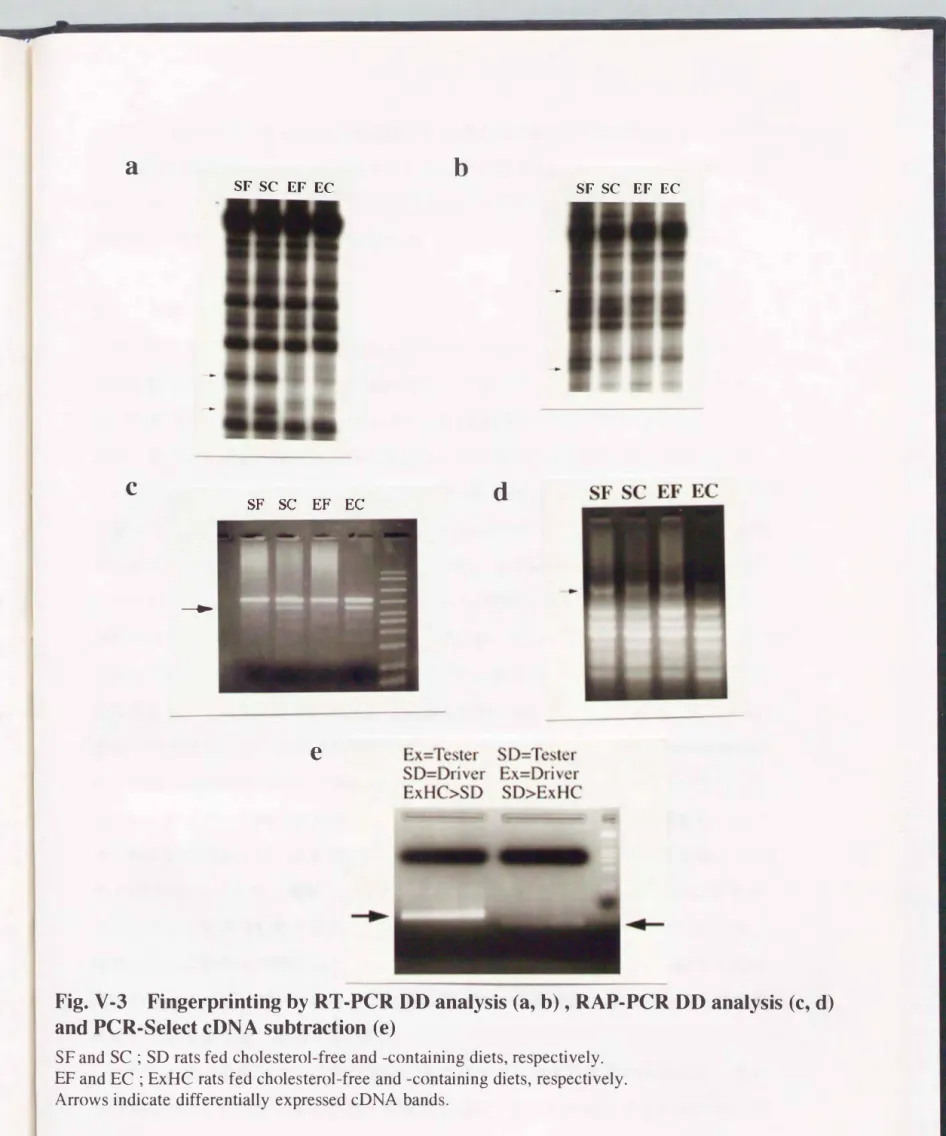
結果関V-3に示した通り、 RT-PCR 00法(a, b)およびRAP-PCRDD法(C, d)によ り、 両系統ラットにおける肝臓でのmRNA発現をそれぞれフィンガープリントする
Tester and driver single-strand cDNA are prepapared from the two mRNA samples under comparison.
RNA
RNA cDNA
AAAACAUGUUCGAAA...AAA
吋'��ÃoY:i岬・ TTTT GT AC AAGCTTT….TTT
cDNA synthesis primer AAAACAUGUUCGAAA...
↓
AAATTTT GTAC AAGCTTT….TTT
Tester and driver double-stranded cDNA are prepapared with using DNA polymerase.
Two tester populations are created with different adaptors, but driver cDNA has no adaptors.
Tester cDNA with adaptor 1
・Eコ
Driver cDNA (30x excess) Tester cDNA with adaptor 2 露翠zz}
a
b c
d
C二回
-E二コ -コ
仁二E・
-=コ
a, b, c, d + e
First hybridization
-二コ
mffZL}
医E2J mI:Z】
Second hybridization: IIÚX samples, add fresh
denatured dri ver, and anneal
E乙E2
ロ�
* Type e molecules are formed only if the tZ:Z:m.i sequence is absent from the driver cDNA.
-•
Add primers・+lm1 Amplify by PCR mm m
Fig.V-2 Schematic diagram ofthe PCR・.Select cDNA subtraction technique
a
b
SF SC EF EC SF SC EF EC
C SF SC EF EC
d
一一・ー
一 -EF 一
二 一
..
e
Fig.
V-3Fingerprinting by RT・PCR DD analysis (a, b) , RAP-PCR DD analysis (c, d) andPCR・Select cDNA subtraction (e)
SF and SC ; SD rats fed cholesterol-free and -containing diets, respectively.
EF and EC ; ExHC rats fed cholesterol-free and -containing diets, respectively.
Arrows indicate differentially expressed cDNA bands.
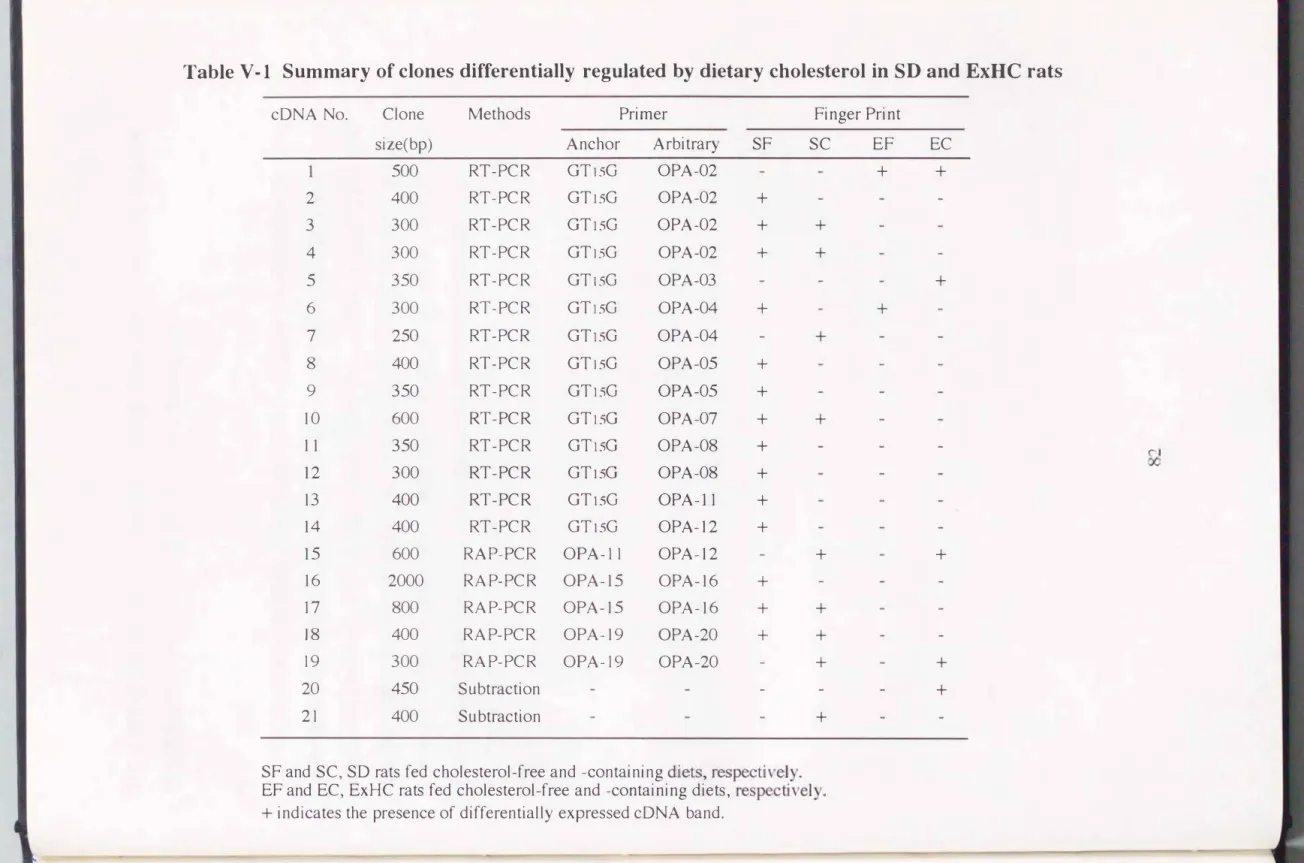
いてコレステロール妓食により強発現している遺伝チをそれぞれ検IHした。 3つの 検索方法から選別された、 ドid系統ラットおよび、食事コレステローノレ蚊食において発 現パターンの呉なった疾忠遺伝子の候補を表V-1にまとめた。 特徴的な発現パター ンをぷす21例のやが!日遺伝子を検lHできた。
V-4
考察RT-PCR DD法、 RAP-PCR DD法のどちらのプ'j1.去によっても、 r�lä系統ラットにお ける肝臓でのmRNA発現について良好なフィンガープリントを得ることができたo RT-PCR 00法はRAP-PCR00法に比べ、 RIを使川することやゲルからのバンドの 選別、 切りl11しなどの傑作がよti雑であるという短所があったが、 フィンガープリン トのバンドの数がより多く作られた。 RAP-PCR 00法はRT-PCR 00法に比べ、 フイ
ンガープリントのバンドの数が少ないという短所があったが、 J弘行: 1二の手間は簡単 で実験にかかるl昨日uが短いという利点があった。 またRT-PCR 00法では、 逆転'R
にオリゴdTプライマーを)11いるので、 cDNAの3'領域が増幅されてくることが多く、
翻訳領域についての↑!?報がイ!?られる頻度が低いが、 RAP-PCR DD法は任意プライマー で逆転写を行うため、 cDNAの増rPMされる領域に偏りはなく、 翻訳領域の情報が得 られることが多い。 それぞれに欠点はあるものの、 両DD法とも特異的に発現する 遺伝子の同定法として非常にイi効であるといえる。 PCR-select cDNA subtraction法 は 2種類の細胞のmRNAをcDNAに逆転写し(一方がテスターで他店がドライ
ノミー)、 テスターに対して大日過剰のドライバーでハイブリットを形成させ、 テス ター特異的に発現している配列を濃縮して、 その後PCRで増幅する方法である。 SD および、ExHCラットから調製したmRNAサンプルを、 それぞれテスターおよびドラ イバーとして組み合わせを変えて反応させることで、 両系統ラットそれぞれに強く 発現している配列を噌rPIAすることができた。 本方法は転写因子のような稀少な転写 産物の検出には適していないが、 比較検体問で特に発現強度に違いがあるものを検 出するには大変イf効であると思われた。
これらの方法を川いて、 21伺の遺伝子をクローニングすることに成功した。 ラッ トの系統やコレステロールの負荷に特徴的な遺伝子がクローニングされたので、 と
Table V -1 Summary of clones differentially regulated by dietary cholesterol in SD and ExHC rats
cDNA No. Clone Methods Primer Finger Print
size(bp) Anchor Arbitrary SF SC EF EC
500 RT-PCR GTI5G OPA-02 + +
2 400 RT-PCR GTI5G OPA-02 +
3 300 RT-PCR GT15G OPA-02 + +
4 300 RT-PCR GTI5G OPA-02 + +
5 350 RT-PCR GT15G OPA-03 +
6 300 RT-PCR GTI5G OPA-04 + +
7 250 RT-PCR GT15G OPA-04 +
8 400 RT-PCR GTI5G OPA-05 + 9 350 RT-PCR GTI5G OPA-05 +
10 600 RT-PCR GTI5G OPA-07 + +
1 1 350 RT-PCR GTlSG OPA-08 + ぐ∞、}
12 300 RT-PCR GTlSG OPA-08 + 13 400 RT-PCR GTlsG OPA-l1 + 14 400 RT-PCR GTlSG OPA-12 +
15 600 RAP-PCR OPA-11 OPA-12 + +
16 2000 RAP-PCR OPA-15 OPA-16 +
17 800 RAP-PCR OPA-15 OPA-16 + + ]8 400 RAP-PCR OPA-19 OPA-20 + +
19 300 RAP-PCR OPA-19 OPA-20 + +
20 450 Subtraction +
21 400 S u btracti on +
SF and SC, SD rats fed cholesterol-free and ←COntaining diets,respree csptlevceulyv .
れらを解析することによって、 ExHCラットの食事コレステロールに対する易応父 性の機構を遺伝子のレベルで追求することが可能となった。
V-5 小す舌
RT-PCRDD法およびRAP-PCRDD法により、
両系統ラットにおける肝臓でのmRNA発現をそれぞれフインガープリントすることができた。 またPCR-select
cDNA subtraction法により、 両系統ラットにおいてコレステロール摂食により強発 現している遺伝子をそれぞれ検出した。 これらの方法によって、 両系統ラットおよ び食事コレステロール侵食において発現ノミターンの異なった、 高コレステロール血 症発症に関係する可能性がある21個の遺伝子の候補を選別する事に成功した。第VI章 総括
食事コレステロール摂取に対して高応答性を示し、 容易に高コレステロール血症 を呈する外因性高コレステロール血症(ExHC)ラットにおける高コレステロール
血症発症機作の解明とその病態発症に対する食事および薬物の改善効果について検 討した。
まず、 食事コレステロールに対する肝臓のコレステロール代謝関連タンパク質の 遺伝子発現の応答について、 ExHCラットとSDラットで比較検討した。 食事コレス テロールに応答してExHCラットのコレステロール7αーヒドロキシラーゼ、mRNAi!
とその活性は増加した。 食事コレステロールに応答してExHCラットのLDLレセプ タ-mRNA量は低下したが、 LDLレセプタータンパク質量はラットの系統や食事コ レステロールの影響を受けなかった。 アポA-IV mRNA量は食事コレステロールの 影響を受けなかったが、 ExHCラットで低い値を示し、 血清アポA-IV濃度も同様で あった。 これらの結果から、
ExHCラットにおける高コレステロール血症の発症は、
コレステロール代謝関連遺伝子の発現のみで説明することはできず、 その他の脂質 代謝経路との相互作用についても検討する必要があると考えられた。
ExHCラットの肝臓トリグリセリド濃度はSDラットのそれと比較して低いことが 知られている。 そこで肝臓のトリグリセリド代謝関連酵素の活性と遺伝子発現に及 ぼす食事コレステロールの影響についてSDラットと比較検討した。 脂肪酸合成酵素 の活性はSDラットに比べE泣-ICラットにおいて低い値を示した。 NADPHの供給系 であるグルコース6-リン酸デヒドロゲナーゼおよびリンゴ酸酵素活性は、 系統およ び食事による影響は見られなかった。 トリグリセリド合成の律速酵素として知られ るホスファチジン酸ホスホヒドラーゼ活性は、 SDラットに比べExHCラットにおい て低い値を示した。 ミトコンドリアにおける脂肪酸酸化の律速酵素であるカルニチ ンアシルトランスフエラーゼ活性は、 SDラットに比べExHCラットにおいて高い値 を示した。 ベルオキシソームのß-酸化活性は、 SDラットに比べExHCラットにおい て低い値を示した。 脂肪酸合成酵素、 グ、ルコース6-リン酸デヒドロゲナーゼおよび リンゴ、酸酵素のmRNAの発現量は、 SDラットに比べExHCラットにおいていずれも
低いイiむをぶした。 これらの結果から、 ExHCラットではコレステロールの摂取とは 関係なく脂肪椴酸化系の充進と脂肪酸およびトリグリセリド合成系の抑制が起こっ ており、 脂肪般の合成に関しては転写レベルで抑制されていることが示された。 し たがって、 ExHCラットの川二臓におけるリボタンパク質合成の際、 トリグリセリド 利JHの低ドがIJ IきI�こされるものと考えられた。
これまでの結*から、 トリグリセリドが少ないコレステロールエステルに富むß
VLDL様の粒子の肝臓からの分泌増加が、 ExHCラットの高コレステロール血症発症 に関与しているものとJfえられることから、 1%コレステローノレ食で1週間飼育した
SD
および�ExHCラットの、 ゴ、ルジ装置中の新/主リポタンパク質の脂質組成を検討し た。 dく1.006g / mLfful分に科!吋する新生VLDLljìの各脂質の別合には両系統ラットに おいて追いが見られた。 日IJち、 コレステロールエステルとタンパク質の割合は、 SD ラットに比べExHCラットで、向く、 逆にトリグリセリドの割合は、 SDラットに比べ ExHCラットで、低いイI立を示した。 コレステローノレエステル/トリグリセリド比を比較 すると、 SDラットに比ベExHCラットで顕著に高い値を示した。 これらの結果から、ExHCラットでは、 JÚllf 1での巽化が遅いβ-VLDL様の粒子の分泌増加が起こってお り、 このことがExHCラットの高コレステロール血症発症の原因であると結論づけ た。
更に、 このようなExHCラットの高コレステロール血症に対する食事成分および
薬物による改善について検討を行った。
まず、 食事脂肪酸の種類とExHCラットのコレステロール代謝との関係について 検討した。
コレステロール無添加条件で、は、 血清のコレステロール、 トリグリセリド、 リン 脂質濃度はすべて、 オリーブ油食群>ココナツ油食群>サフラワ一泊食群の順であっ た。 食事のモノ不飽和脂肪酸は多価不飽和脂肪酸、 飽和脂肪酸と比較して、 ExHC ラットのLDLレセプターの発現を抑制する事で血中コレステロール濃度を上昇させ る可能性がぷされた。
コレステローノレ添加食の条件下では、 血清の総コレステロール濃度はオリーブ油 食摂食群において顕著に上昇した。 ココナツ油食群はオリーブ油食群と比較して、
コレステロール7αーヒドロキシラーゼの活性およびmRNAの発現の克進などにより、
血清コレステロール濃度上昇を抑制するようであった。 サフラワ一泊食群による血 清コレステロール濃度上昇抑制機構は明らかでないが、 ExHCラットにおける肝臓 トリグリセリド濃度の低下が顕著で、ないことから、 トリグリセリド代謝を介して、
高コレステロール血症抑制に作用する可能性がある。 なお、 LDLレセプターmRNA レベルに対して食事脂肪の種類は影響しなかった。
次に、 ExHCラットではベルオキシソームのβ-酸化活性が低下していたことから、
ベルオキシソームプロリファレーターの一種であり、 血清トリグリセリド低下剤と して用いられているジェンフィブロジルのExHCラットの高コレステロール血症発 症への影響を検討した。 この薬剤投与により、 ExHCラットのコレステロール負荷 に基づくHDLコレステロール濃度の低下が改善され、 非HDL画分のコレステロール 濃度の上昇が抑制された。 ExHCラットの非HDL両分のリボタンパク質中のトリグ リセリド/コレステロール比は、 薬剤投与によりSDラットのものと同程度まで上昇 した。 薬剤投与したExHCラットでは、 肝臓のコレステロール7 a-ヒドロキシラー ゼ活性とそのmRNAおよび胆汁酸の排離電などの増加とACAT活性の低下が観察さ れた。 SDラットでは薬剤投与によってACAT活性の低下は認められたが、 コレステ ロール7αーヒドロキシラーゼに対する影響は見られなかった。 しかしHMG-CoA還 元酵素の活性の顕著な噌加が見られた。 これらの結果から、 ExHCラットで低下し ているベルオキシソームの活性を上昇させることによって、 ACAT活性、 胆汁酸合 成やコレステロールに富むリボタンパク質の分泌に改善効果がもたらされ、 高コレ ステロール血症が抑制されると考えられた。
またベルオキシソームプロリファレーターによるExHCラットのコレステロール
7 a-ヒドロキシラーゼ遺伝子発現の上昇の機構を調べる目的で核内転写因子NF-KB の発現量を測定した。 コレステロール7α-ヒドロキシラーゼ遺伝子のプロモーター
領域はNF-KBの結合領域を有することから、 この配列をプロープにしてゲルシフト アッセイ法により、 ジェンフイブロジル投与の影響について評価した。 この配列を 認識している証拠となるシフトバンドが核内タンパク質中に確認されたが、 ジェン