野生食肉目動物、特にタヌキにおける
Bartonella
属菌の生態学的研究日本大学 大学院 獣医 学研究科 獣医学 専攻 博士課 程
水 上 昌 也
2020
目次
第
1
章 各 国の 野生 食 肉目 動物 に おけ る 病原 性Bartonella
の感 染状 況に 関す る 文献 的考 察1.1
は じめ に... 2
1.2
ネコ 型亜 目 にお け るBartonella
属 菌の 生態... 7 1.3
イヌ型 亜 目に おけ るBartonella
属菌 の生 態... 9
第
2
章 イ ヌ科 動物 の 血液 中Bartonella DNA
検出 を 目的 とし たPCR
法の 検 討2.1
はじ めに... 19 2.2
材料 およ び 方法... 22
2.2.1
検 出感 度 測定 用の プラ ス ミ ドDNA
の作 製a)
使用 菌株 とDNA
抽出b) B. rochalimae BAA-1498
T株 のrpoB
お よ びssrA
領 域 のPCR
法に よる 増 幅c) PCR
の 検 出感 度 測定 用プ ラ スミ ドDNA
溶液 の 作製d
) プラ スミ ドDNA
溶液 のコ ピ ー数 の 調整2.2.2 PCR
法の 検 出 感度 測定2.3
成績 ... 302.3.1
テ ンプ レ ー ト DNA① を 用い たPCR
法の 検出 感 度2.3.2
テ ンプ レ ー ト DNA② を 用い たPCR
法の 検出 感 度2.4
考察 ... 362.5
小括 ... 392.6
第2
章で 使用 し た試 薬類 の 組成... 41
第
3
章 タ ヌキ にお け るBartonella
感染 状 況、 疫学 要 因 な ら びに 検出 株の 遺 伝子 解 析3.1
はじ めに... 43 3.2
材料 およ び 方法... 46
3.2.1
検査 材料3.2.2 PCR
に よ るBartonella DNA
の検 出3.2.3 Bartonella DNA
シー ケン シ ング3.2.4
遺伝 子系 統解 析 およ び相 同 性解 析3.3.5
統計 解 析3.3
成績 ... 493.3.1
タ ヌキ にお け るBartonella DNA
保 有 率3.3.2
タヌ キか ら検 出 され たBartonella DNA
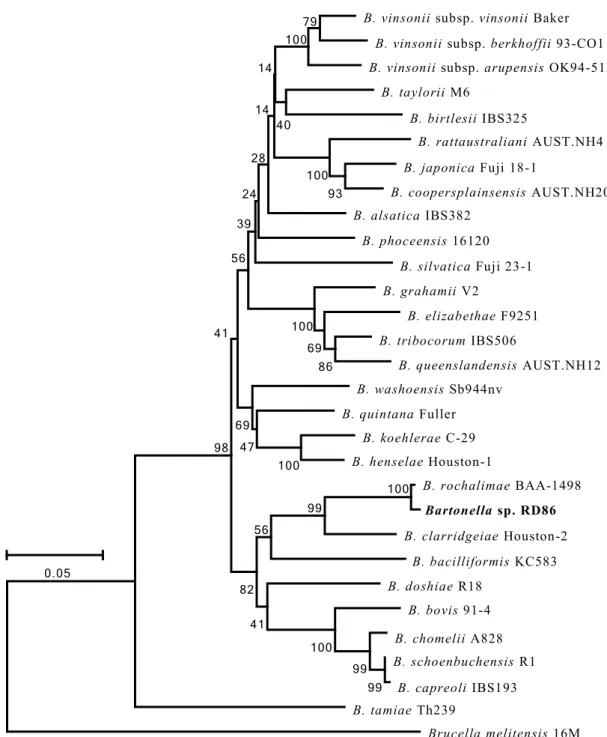
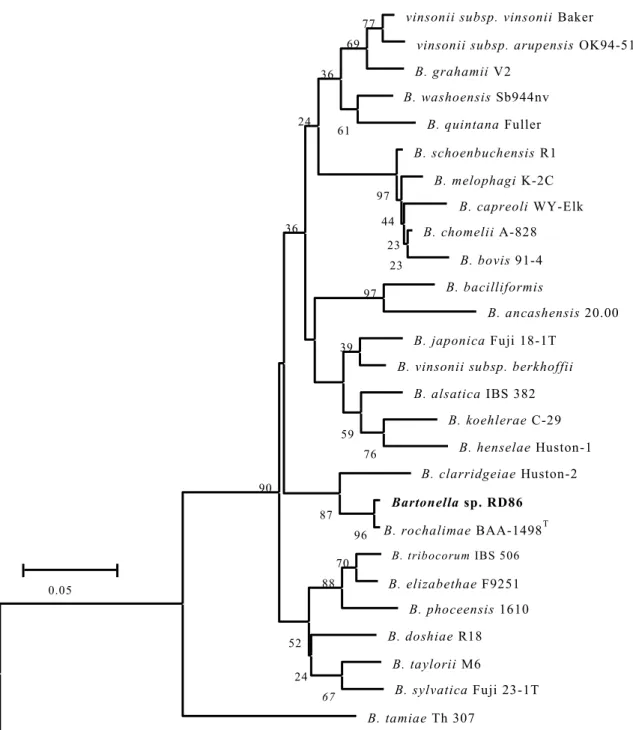
の系 統 解析3.3.3 BLAST
検 索に 基 づく タヌ キRD86
株 の 相同 性解 析3.3.4
タヌ キに おけ るB. rochalimae DNA
保 有率 に関 す る疫 学 要因3.4
考察 ... 583.5
小括 ... 62第
4
章 総 括... 64
謝辞 ... 75 引用 文献
... 76
1
第
1
章各国の野生食肉目動物における
病原性
Bartonella
の感染状況に関する文献的考察2
1.1
は じめ にわ が 国 には 、 多 種 多 様 な 野 生動 物 が 複 雑 な 生 態 環境 下 で 生 息 し 、 そ の 数 は 外来 種 を含め
31
科72
属170
種 にも達 す る[Ohdachi et al., 2009]
。この うち 、 陸 上に 生 息する野 生 食肉目 動 物は、ネ コ型亜 目動物に属 するネ コ科、マングー ス 科 およ び ジャコウ ネ コ科、イ ヌ型 亜目 に属す る イヌ科 、クマ科、イタ チ科およ び ア ライ グ マ科の計7
科32
種が 生 息して い ると 考えら れている。ネコ型亜目動物 のネコ科では、在来種のイリオモテヤマネコ が沖縄県(西表島)、ツシマヤマネ コが 長崎 県(対馬)をはじめ、外来種では、ジャコウネコ科のハクビシン が全 国 に、マングース科のフイリマングースが 沖縄県や 鹿児島 県(奄美大 島)に 生息し て いる 。イヌ 型亜目動物では、クマ科のヒグマ は北海 道に限 局して生息 して お り、ツ キノ ワ グマは北 海 道、九州 を 除く地 域 に生息し ている。また、イヌ科 のタヌ キ、
キ ツネ を はじめ、イタチ 科 のテン 、イタ チ 、イイズ ナ、オコ ジョ、ミン ク、ニホ ン アナ グ マ、外来 種 のアラ イ グマな ど は 全国に 生 息している 。
わ が 国 の野 生 動 物 の 多 く は 、人 の 生 活 圏 と 重 な るよ う に 生 息 し て お り 、 高 齢 化 に よ る 耕 作 地 の 放 置 や 若 者 の 都 市 部 へ の 流 出 に よ る 里 地 里 山 の 崩 壊 等に よ り、そ の生息 数は 近年 著し く 増加 して い る 。その結果、農・畜産業に及ぼす被 害 は年 間
10
億円以上に達し、2009
年か ら2016
年にかけては毎年ほぼ同様の被 害 額で 推 移してい る( 図1-1
)。また、食肉目動物による被害は農作物にとどまら ず、特に外来種のアライグマや フイ リ マ ングース 等は、ニホンザリ ガニ 、エゾサ ン シ ョ ウ ウ オ や ア マ ミ ノ ク ロ ウ サ ギ と い っ た 日 本 固 有 種 の 捕 食 や 在 来 種 の タヌ キと 生 息域を競合 する など 、わが国の生態系に も 大 きな影響を 及ぼし ている。さ ら に、文化遺産などへの被害、住居侵 入 や直接 的 な人へ の危害など も日 本 各地 で 報 告さ れ るように な った (図1-2
)。3
以 上の よう な背 景か ら、野 生 動物に 起 因する 各種被害を 解消す る目的 で、
日 本 各 地 で 自 治 体 を 中 心 と し た 有 害 鳥 獣 駆 除 事 業 が 積 極 的 に 行 わ れ る よ う にな っ た。その結果、ハクビシン、アライグマ、ニホンアナグマ、タヌキ、クマ等の 野 生食 肉 目動物の 捕 獲頭数 は 、
2009
年 に は約22,000
頭 であったの が2016
年 には 約
77,000
頭へと 年 々 増加 して いる( 図1-3
)。また、タ ヌキ は 人家周辺に生息し て い る こ と に も 関 連 し 、 傷 病 動 物 と し て の 保 護 頭 数 が 近 年 著 し く 増 加 し てい る( 図
1-4
)。近 年、 野生 食肉 目動 物は 各種新 興 ・再興感 染症の 媒介動物 とし て重要 な 役 割を 担 っている こ とが 認知 され る よ うにな った 。米国では アライグマ 、ス カ ン ク、 ヨ ーロッパ 圏 におい て はキツ ネ などが 狂 犬病の 媒介動物と して重 要であ る 。そ の他の疾病 では 、 パス ツレ ラ 症 、エキ ノコックス 症、ア ライグマ回 虫症 な ど種 々 の細菌性・寄 生虫 性感 染 症 を 含 む 人獣共 通感染 症 の
43%
が 、食肉目動 物 に由 来 すること が 報告さ れ ている[Cleaveland et al., 2009]。わが国に おいて
も 、北 海 道のキタ キ ツネは 、 エキノ コ ックス 症 の重 要な媒介 動物と して注目す べ き食 肉目動物となっ て いる[Yimam et al., 2002]
。バ ルト ネラ 症は 、
Bartonella
属の 細 菌を原因 とする 新興・再興感 染症であ る。猫ひっかき病(Cat-scratch disease:CSD
)は、その名が示すように、猫が重要 な感染源・病原巣 とす る代 表的 な バ ル トネラ 症 であるが、近年 、コヨーテやアラ イグマ などの 野生食 肉目動 物 を自 然宿主 とするBartonella
属菌が 、人に 心内膜 炎、脾腫、関節炎などを 引 き起こ す 新 たなバ ル トネラ 症の原因と なる ことが明ら か にな っ てきた[Breitschwerdt et al., 1995]
。Bartonella
属は、グラム陰性の多形性短桿菌(0.5~ 0.6×1.0µm)で、ノミ、
シラミ、サシチョウバエなどの吸血性節足動物によって媒介される。本属菌は、
4
霊長目、齧歯目、食肉目、ウサギ目、げっ歯類などの哺乳類を自然宿主として、
宿 主 の 血 管 内 皮 細 胞 な ら び に 赤 血 球 内 に 持 続 感 染 す る こ と で 長 期 間 の 菌 血 症を 引 き起 こ す
[Breitschwerdt et al., 2010]
。本菌 の 発育には赤 血球成 分の一つで ある ヘ ミン を 必要とし、そ の発 育速 度 は極 め て遅い ため、その分離には血液 寒 天培 地 を 用いて5
%のCO
2濃度 、20
~37
℃環境下で7
~21
日間 と長期 間培養する 必要 がある。また、Bartonella
は種 々 の抗 菌 性物質 に感受性を 示すた め、本 菌を分 離 培 養す る ための優 れた 選 択培 地は 未 だ に 開発され ていな い。Bartonella henselae
は 、猫ひっ か き病の起 因菌で あり[Regnery et al., 1992]
、 猫 科動 物 を自然宿 主 とする 代 表的なBartonella
であ る。後 天性免疫不 全症候 群患 者 や臓 器 移植者な ど の免疫 不 全状態 の 人にお い ては、細菌性血 管腫症、心内膜炎、Bacillary peliosis
、Bacillary splenitis
が 重 篤 な 合 併 症 と し て 報 告 さ れ て い る[Regnery et al., 1992]
。1995
年に米国のClarridge
ら[1995]により猫から分離された B. clarridgeiae
も、CSD
の原因となる可能性が ある こ とがKordick
ら[1997]
によ り報告 された 。1999
年 にDroz
ら[1999]
は、米国 の サ ンフラ ン シスコ 湾 周 辺地域の飼 い猫 から分 離 され たB. henselae 25
株の うち 、 既 存のB. henselae
と表現型 お よび遺伝子 型が 異 なる 株 が2
種あるこ と を発見 し た。これら の 株は、gltA
領域に おける 遺伝子 解 析 では 区 別するこ と はでき な かった が 、DNA-DNA hybridization
の成 績から 、B.
henselae
と は異なる新 種 のBartonella koehlerae
として報告された。発見当初、B.koehlerae
の 人に対 す る病原 性 は 不 明であ っ た が 、2004
年、人に心内膜炎を起こ す こと がAvidor
ら[2004]
により 初 め て報告 さ れた。1995
年 にBreitschwerdt
ら[1995]は 、 米 国 の 心 内 膜 炎 の イ ヌ か ら 新 種 の
Bartonella
を分離し 、DNA-DNA hybridization
や16sRNA
の 塩基配 列を用いた 系5
統 解析の 成績から、分離 株は
B. vinsonii subsp. berkhoffii
とし て報告され た。2000
年 には 、 心内膜炎 に罹 患 した 患者 か ら もB. vinsonii subsp. berkhoffii
のDNA
と 抗 体が 検 出され、本 菌は 人に 対し ても病 原 性を示 すBartonella
である ことが 明ら か にな っ た[Chang et al., 2000]
。2007
年 には 、発 熱、 脾腫 、筋 肉痛、 関 節炎な どの多様 な症状を呈した女 性 の血 液 から、Bartonella
様 の細菌 が 分離さ れた[Eremeeva et al., 2007]
。 この 患 者 は 発 症 前 に ペ ル ー に 滞 在 し て お り 、 滞 在 中 に 多 数 の 吸 血 性 節 足 動 物 の 寄 生を 受 けて い たことか ら 、当 初、アンデス地域の ベ クター媒介 性の感染症 である カリ オ ン病が疑われた。患者から分離された株は、単極に複数の鞭毛を 保有す るな ど のB. clarridgeiae
様 の形態 を 示し[Sanchez et al., 2016]、 ま た、ペルー のヒト ノミ(
Pulex irritans
)か ら 検出 さ れた 新 種と思 わ れるBartonella
の16S-23S rRNA
遺 伝子間(ITS
)領域のDNA
配列 と 非 常に高 い 相同性(99.8%
)を示したことから、新種 の
Bartonella rochalimae
と し て命名 さ れた[Parola et al., 2002]。そ の後 、B.
rochalimae
は、米国のアライグマ、ハイイロギツネ、およびフランスのアカギツネ か ら も 分 離 さ れ 、 野 生 食 肉 目 動 物 が 自 然 病 原 巣 で あ る こ と が 明 ら か に な っ て きた
[Henn et al., 2009]。
以 上に 示し たように、食肉目動物 は、種 々の
Bartonella
の 自 然病原 巣を 形 成 し て いる こ とが 近年 明ら かに な って き たことか ら、Bartonella
菌の生態を明ら か に す るこ とはバルト ネ ラ症の 疫 学を解 明 する上 で、き わめて 重要である 。また、Bartonella
属菌の生 態解明を 行う 上で、 分離培養 法は 直近の 感染状況 を把 握し、分 離 菌 の 種 々 の 遺 伝 学 的 性 状 等 を 解 析 す る 上 で 極 め て 有 用 な 方 法 で あ る が 、 対 象 動 物 から 無菌的に採 取 した新 鮮 な血液 が 必要と なる。Polymerase chain reaction
(PCR)
法は、試料の状態に ほと ん ど影 響 される ことなく、目的と す る病原 体の 遺6
伝子を高 感度 に検 出 すること が可能で あ る。
PCR
法は、 様 々な動物 の血液や臓 器 か ら のBartonella DNA
の検出に応用される一方で、生菌・死菌の双方を検出 す る こ とや非 特異 反応 に よる偽 陽 性が問 題 となるこ とがあ る。また、抗体検出は、過 去 の 感染 を知る上で 有 用な方 法 である が 、Bartonella 属 の菌種 間の交 差反 応が 生 じ こ とが あ るた め 、 抗 体陽性 率 は 分離培 養 法や
PCR
法 の陽性率 に比べて高く な る 傾 向に ある。以上から、野生食肉目動物の
Bartonella
感染の疫 学を研 究する際 には、各 手 法 の 特性 を充分 理 解 し た上で 実 施する 必 要がある 。7
1.2
ネ コ型 亜目に お けるBartonella
属菌 の生態世 界各 国で 、 飼育 下の 猫 (
Felis domesticus
)におけ るB. henselae
の分離 培 養 法 に よ る 保 菌 率 が 検 討 さ れ て い る 。 そ の 値 は 、 ノ ル ウ ェ ー で0%
(0/100
)[Bergh et al., 2002]、日本で 6.4%(44/690
)[Maruyama et al., 2000]、オーストラリ ア(シドニー)で35.0%
(27/77
)[Branley et al., 1996]
、米国(カリフォルニア州)で
39.5%
(81/205
)[Chomel et al., 1995]
、インドネシアで43%
(5/14
)[Marston et al., 1999]
、フィリピンで55%
(17/31) [Chomel et al., 1999]、ジンバブエで 8%
(2/25)
[Kelly et al., 1998]
と いった よ うに 、国によって様々である。野生のネ コ型亜 目動 物に お いて は、アフリカの野生ライオンの5.2%
(3/58
)やチーター の33%
(1/3
)[Kelly et al., 1998]
か らB. henselae
が、チーター の5.9%
(1/17)からはB. koehlerae
が 分離 さ れている[Molia et al., 2016]
。また、米国のマウンテンライオンやボブキ ャ ット は 、B. henselae
をは じめ 、B. koehlerae subsp. boulouisii
やB. koehlerae subsp.
bothieri
といった新種 のBartonella
属 菌を保 有 してい ることも明 らかとなってい る[Chomel et al., 2016]
。 このよ う に、ネ コ 型亜目動 物の 多くが、本 菌の自 然宿 主 であ ることが 諸 外国 の 研究 者に よ り 示 されてい る (表1-1
)。わ が 国 の野 生 ネ コ 型 亜 目 で は、 特 別 天 然 記 念 物 に指 定 さ れ て い る イ リ オ モ テヤ マ ネコの
6%
(2/33
)やツシマヤマネコの7.7
%(1/13
)からB. henselae
やB. clarridgeiae DNA
が 検出さ れ ている[Tateno et al., 2013]
。さらに、 外 来種 のフ イ リマ ング ースの15%
(10/63
) やハク ビ シンの2.0%
(1/50
)から もB. henselae
が 分離 さ れている[Sato et al., 2013]
(表2-2
)。また、 高知県 ではペット とし て 飼 育 し て い た ハ ク ビ シ ン に 飼 い 主 が 引 っ 掻 か れ た 後 に 猫 ひ っ か き 病 を 発 症 し た事 例 が報 告 されてい る ことか ら[
宮 崎ら, 2001]
、野生ネコ型亜目動物も猫ひっかき8
病 を は じ め と す る バ ル ト ネ ラ 症 の 感 染 源 あ る い は 病 原 巣 と し て 重 要 な 役 割 を果 た して い るものと考え ら れる 。
9
1.3
イ ヌ型 亜目に お けるBartonella
属菌 の生態犬 (
Canis familiaris
) のBartonella
の 病 原 巣とし て の役割は 明確ではない が 、こ れ までヒト の 心 内膜炎 の 症例か ら 、B. vinsonii subsp. berkhoffii
が検出され て おり、 犬の本菌によ る 心内 膜炎 の 病 態に類 似してい ること から[Chomel et al., 2009 ]
、犬はB. vinsonii subsp. berkhoffii
の 病 原巣であ ると考 えられてい る。近 年、 各国 の 野生 イヌ 型 亜 目 におい て も
Bartonella
の保有状 況が 研究さ れ てい る(表1-1
)。 米国 の ハイイ ロ ギツネ の42%
(22/53
)、アライグマの26%
(
11/42
)、コヨー テ の9.5%
(2/21
)[Bai et al., 2016]
、フラ ンスの アカギツネ (1
頭)から人に脾腫、貧血、発熱を起こすB. rochalimae
が分離さ れてい る[Henn et al., 2009]
。また、米国のシマハイイロギツネの11.8%
(6/51) [Schaefer et al., 2011]、
ハ イイ ロ ギツネの
9.4%
(5/53
)、コヨーテの28
%(31/109
)から は人の心内膜炎 の 原因 菌となるB. vinsonii subsp. berkhoffii
が分離 さ れている[Bai et al., 2016]
。PCR
法を 用い た研 究で は 、メ キ シコのキット ギツネ の13.3%( 2/15)とコ
ヨ ーテ の5.6
%(1/18
)[López-Pérez et al., 2017]
、オーストリアの ア カギツネ の0.2
%(
1/506
)[Kaewmongkol et al., 2011]
、スペインのアカギツネの1.6
%(1/62
)とオ オ カミ の33.3
%(1/3)[Gerrikagoitia et al., 2012]、イスラ エルの キンイロジ ャッ
カルの55
%(5/9
)とアカギツネの9.1
% (1/11
)[Marciano et al., 2016]
からB.
rochalimae
のDNA
が 検 出され て いる。 さ らに、米 国のハ イイロギツ ネから 採取 し たヒ ト ノミの一 種(Pulex spp.) [Schaefer et al., 2012]や チリのイヌ から採 取し
た ノミ[Pérez-Martínez et al., 2009]
か ら もB. rochalimae
のDNA
が 検出さ れた こ と から 、B. rochalimae
の自然宿主 はア ライグマ やキ ツネな どの野生 イヌ 型亜目 動 物で あ ることが 明 らかと な り、Pulex 属の ノ ミが本 菌の 主要な ベクターである と考え られ てい る 。 東部 ス ロバ キア のア カギツ ネに おけ るノ ミの寄 生率 は72%
10
(
56/78
)と 非常 に 高 い 値であ り[Kočišová et al., 2006]
、また同国 の アカギ ツネ か ら 回収 し たノミの4.7%
(19/407
)からBartonella DNA
が検出さ れて いる こと か ら、ノミは アカ ギツ ネに おい てBartonella
の 重 要なベ クターの一 つであ る 可能性 が 示唆 さ れている[Víchová et al., 2018]。さら に 、イラ クのジャッ カルの 12.3%
(
7/57
)からは 、 新種 と思 われ るBartonella merieuxii
のDNA
が検出されている[Chomel et al., 2012]
。 以 上 に 示 し た よ う に 、 各 国 の イ ヌ 科 動 物 も 、 種 々 のBartonella
の自 然病 原巣 を形 成し て い ると考 え られる(表1-1
)。また、米国では コ ヨー テ に咬まれた 人 が 、バルトネラ症を発症した事例が報告[Chang et al., 2000]
さ れ て い る こ と か ら 、 野 生 イ ヌ 型 亜 目 も 人 の バ ル ト ネ ラ 症 の 感 染 源 と し て 注目 す る必 要 がある。
わ が国 の野 生イ ヌ型 亜目 で は 、テン の
12.5
%(1/8
)から病原性は不明で あ るが 、 北米のジ リ ス由来のB. washoensis
と 高い相同性 を示す 株が 分離されて い る。さらに、ニホ ンア ナグ マの6.7%( 1/15)から新種と思われる Bartonella
も 分 離さ れ ている[Sato et al., 2012]
(表1-2
)。以上から、各国 の野 生食 肉 目動 物 は、数 種の病 原性
Bartonella
を保 有し て お り、特 にネコ型亜目 で はB. henselae、 B. clarridgeiae
、お よびB. koehlerae
が、イ ヌ型 亜 目では、
B. rochalimae
お よ びB. vinsonii subsp. berkhoffii
が それ ぞれ の 亜 目の 動 物におけ る主 要な 菌種 で あ る こ とが判明 した。さ らに 、亜目 ご と に固有 の 菌種 を 保有して い ること 、ネ コ 型亜 目 お よびイヌ 型 亜目の動物は、それぞれ新 種 のBartonella
を 保 有して い ること も確 認 され た。一方 、わが国固有 のイヌ 型亜 目 動物 で あるタヌ キ は、人の生活圏に密接した環境に生息している ため、有害獣 と し て 駆 除 さ れ た り 、 傷 病 獣 と し て 保 護 さ れ る 件 数 は 年 々 増 加 し て い る に もか かわらず、その病原性Bartonella
の保 有 状況に ついては未 だ 検 討 されていな い。11
したがって、 タヌ キ におけ る
B. rochalimae
を 含む病原性Bartonella
の感染状況 を 明 ら か に す る こ と は 、 わ が 国 に お け る バ ル ト ネ ラ 症 の 疫 学 を 解 明 す る 上 で重 要 な課 題 であると 思 われた。さら に、食 肉 目動物 由 来のBartonella
に感染した人 の 事 例 が 日 本 や 米 国 で 報 告 さ れ て い る こ と か ら 、 野 生 食 肉 目 動 物 を 取 り 扱 う際 は 、 本 症 の 病 原 巣 で あ る こ と を 十 分 認 識 し た 上 で 対 応 す る 必 要 が あ る と 思 われ た 。12
図
1-1
わが 国の 野生 食肉 目動 物 によ る 農業被害 額の推 移 (2009~ 2016
年)0 2 4 6 8 10 12 14 16
タヌキ アライグマ ハクビシン クマ
2009 2010 2011 2012 2013 2014 2015 2016
年
被
害
額
出典:農林水産省(
https://www.maff.go.jp/
)農作物被害状況 よ り( 億 円)
13
図
1-2
食肉目動物による各種被害a.
タヌキによるトウモロコシの食害(出典
URL:https://plaza.rakuten.co.jp/hananoyakata/diary/201708290000/) b.
ハクビシンによる天井裏の糞尿(出典
URL:https://npo-hakubishin.com) c.
床下のアナグマ(出典
URL
:https://mosika1.hamazo.tv/e7173670.html
)d.
アライグマによる咬傷(出典 URL:https://www.whistlerquestion.com/news/local-news/local-woman-attacked-by-raccoon-
after-rescuing-dog-1.23106017)
14
0 10,000 20,000 30,000 40,000 50,000 60,000 70,000 80,000 90,000
2009 2010 2011 2012 2013 2014 2015 2016
タヌキ アライグマ ニホンアナグマ ハクビシン クマ
年
捕
獲 頭
数
図
1-3
有害 鳥獣 捕獲 によ る 野 生食肉 目 動物の 捕獲頭数の 推移 (2009~2016
年)出 典: 環境 省 野生鳥 獣の保護管 理
(
http://www.env.go.jp/nature/choju/index.html
)15
図
1-4
日本の傷病鳥獣としての食肉目動物の保護頭数の推移0 100 200 300 400 500 600 700 800 900 1000
2009 2010 2011 2012 2013 2014 2015 2016
タヌキ ニホンアナグマ ハクビシン
保 護 頭 数
年
出 典 : 環 境 省 野 生 鳥 獣 の 保 護 管 理
(
http://www.env.go.jp/nature/choju/index.html
)16
表
1-1
世界の野生食肉目動物から検出されたBartonella
属菌16
亜目 自然宿主 菌種 人に対する症状 引用文献
ネコ型亜目
ボブキャット
B. henselae 患部発赤、リンパ節腫脹、発熱
(猫ひっかき病)
Chomel et al., 2016
ピューマチーター
Molia et al., 2016
ライオンリビアヤマネコ
Gerrikagoitia et al., 2012
チーター B. koehlerae 心内膜炎
Molia et al., 2016
イヌ型亜目
ハイイロギツネ
B. vinsonii subsp. berkhoffii 心内膜炎
López-Pérez et al., 2017
シマハイイロギツネ
Schaefer et al., 2011
コヨーテ López-Pérez et al., 2017
アライグマ
B. rochalimae 脾腫、貧血、発熱
Henn et al., 2009
キンイロジャッカル
Marciano et al., 2016
コヨーテ López-Pérez et al., 2017
ハイイロオオカミ
Gerrikagoitia et al., 2012
17
表
1-2
日本の野生食肉目動物から分離されたBartonella
属菌.
17
亜目 自然宿主 菌種 引用文献
ネコ型亜目
フイリマングース
B. henselae
Sato et al., 2013 ハクビシン
イリオモテヤマネコ
Tateno et al., 2013 ツシマヤマネコ
イヌ型亜目 ニホンアナグマ Bartonella sp.
Sato et al., 2012
テン B. washoensis-like bacterium
18
第
2
章イヌ科動物の血液中
Bartonella DNA
検出を目的としたPCR
法の検討19
2.1
はじめ に各 種動 物の 血液 を用 いて
Bartonella
感 染の有無を 検討す る場合、 間接蛍 光 抗体法 による 抗 体検 出 法[常 岡 ら, 1998]と PCR
法や分 離培養 法による抗 原検 出 法 が あ る 。 し か し な が ら 間 接 蛍 光 抗 体 法 で 本 菌 に 対 す る 特 異 抗 体 を 検 出 する た めの野 生動物の 二次 抗体 の 入手は 困 難で 、また 、Bartonella は 、Q 熱やクラミ ジ ア肺 炎 の原因菌であ るCoxiella burnetii
やChlamydia pneumoniae
と交差 反応 を 示 すた め[
常岡ら ,2001]
、野生動物におけるBartonella
の感染を 血 清学的 に証 明 するこ とは 極 めて難 しい 。 一方、 血液か らBartonella
を分離す る 分離 培 養法とBartonella DNA
を 検出 するPCR
法で は 、 分離株や 検出さ れたDNA
の塩 基配 列 か ら 、 宿 主 に 感 染 し て い る 菌 種 を 同 定 す る と と も に ゲ ノ ム 性 状 を 詳 細 に 解 析す るこ と が可 能であ る[La Scola et al., 2003]。 特 に、培養細 胞を用 いた感染実 験で
分 離株 の 病原性も 解 析でき る ことか ら[Eicher et al., 2012]
、バルトネ ラ症の 疫学 を 研究 す る上で分 離 培養法 は 理想的 な 方法で あ るとい える[Gutiérrez et al., 2017]。
猫 にお ける
Bartonella
感染を調べる場合、分離培養法が 広 く用い られ て お り、米 国[Regnery et al., 1992]
、ドイ ツ[Heller et al., 1997]
、 フランス[Heller et al., 1997]
、 日 本[Maruyama et al., 2004]
な ど を は じ め 、 世 界 各 地 の 猫 に お け るBartonella
菌の保有状況が検討されている 。Bartonella
はその発 育に血 液成分 を必要とするため、分離には、ウサギやヒツジの血液を加えた栄養 価 の高い ハー ト インフュ ージョ ン寒 天培地が 用いら れ て いる
[Regnery et al., 1992]。 血液 寒天培
地 上に おけ るBartonella
の発育速度は、大腸菌やサルモネラ などと比 べ ると極め て 遅く、 可視化コロニ ー の形成 ま で最低2
週 間 を要す る。 また、Bartonella
は、種 々 の 抗 生 物 質 に 対 し 感 受 性 を 示 す た め 、 優 れ た 選 択 分 離 培 地 は 未 だ に 開 発さ れ てお らず、本菌 を 分離 す る場 合 、検 体 から無 菌 的に採 血した 新 鮮 血液が 必要 と
20
な る。しかしながら、タヌキを含 む野 生動物 から 血液を 採取する 場合 、狩 猟で 捕 獲した個体、交通 事故死 あ るい は 野外 斃 死個体 等 か ら採血する ことが 多いた め、
無 菌 的 な 新 鮮 血 液 を 入 手 す る こ と は 、 極 め て 困 難 で あ る 。 一 方 、
PCR
法 は 、Bartonella
の特 定の 遺 伝子 領 域 を高感度、かつ特異的に増幅する適切なプライマー が あ れ ば 、 血 液 検 体 の 雑 菌 汚 染 の 有 無 に か か わ ら ず 、 対 象 と な る 動 物 の
Bartonella DNA
の 保有状 況 を正確 に 把握で き ると考え られる 。こ れま で、
PCR
法 を 用いて 様 々な野 生 食肉目動 物からBartonella DNA
が 検出されている。Hennら[2009 ]は 、 Bartonella
のITS
領域、クエン酸 合成酵 素遺 伝子(gltA
)、RNA
ポリ メラ ーゼ サ ブ ユニッ ト β(rpoB
)、および 細胞分 裂 タンパク質 (
ftsZ)領 域 を そ れぞれ 標的とした PCR
法によって、フランスの1
頭の アカ ギツ ネ の血液から
Bartonella DNA
を 検 出し、 その塩基配列からB. rochalimae
で あっ たことを明らか に してい る 。Gerrikagoitia
ら[2012]
は 、gltA
領域を標的と し たPCR
法に より 、 スペイ ン の1
頭 のハイ イ ロオオ カミの 肝臓およ び脾臓 からB. rochalimae
を検 出し てい る。López-Pérez ら[2017]も、gltA 領域を 標 的 としたPCR
法でハイイロ ギツネ5
頭 の 血液か らB. rochalimae DNA
を 検出してい る。Bartonella
の 菌 種 を同 定す る 場 合、gltA
領域とrpoB
領域 の 塩基配 列に お け るBartonella
標 準 株との 相 同性値 を用 い 、gltA
領域で は96.0%未満 、 rpoB
領 域が
95.4%
未満であれば 、解析に用いたBartonella
株は 既存種とは 別 種であ ると 定義 され る
[La Scola et al., 2003 ]
。このことから、両領域 の塩基配 列 解析は 検出 した
Bartonella
の 菌種 を同 定する 上で 必 須であ ると考 えられ てきた 。しか しながら 、
gltA
領 域を 増幅 する プラ イマ ー は 、マウス、ラ ット、ヒ トなどの宿 主 由来 ゲ ノムとアニ ール反 応 し、PCR
法におい て、 顕著な 非特異 反 応が生じる 可能性が あ る こ と が 指 摘 さ れ て い る[Diaz et al., 2012]
。 ま た 、 複 数 の げ っ 歯 類 由 来21
Bartonella
株を 用い た ゲノム解析では、異なる株間で同領域の 遺伝子 組換えが生じ る 可 能 性 が 報 告 さ れ て い る
[Paziewska et al., 2011 ]
。 こ れ ら の 背 景 か ら 、Bartonella
の菌 種 を 同 定 する 場 合 、gltA 領域 以 外の複 数の遺伝子 領域を 解析する こ とが推 奨されるよう に なった 。Diaz
ら[2012]は 、 Bartonella
の トランスファー メ ッセ ン ジャーRNA(ssrA)領域を標的とした real-time PCR
法 によるスク リー ニ ング 検 出法を開 発 した。 ま た、ssrA
領域に 基づく 系統 解析の成績 がgltA
領 域 に 基 づ く も の と 非 常 に 近 似 し て い た こ と か ら 、Bartonella
の 疫 学 研 究 に お い てssrA
領 域 を用 いたPCR
法も 広 く 用 いられ る よ うになっ た[Marciano et al., 2016 ]。第
2
章 では 、 タヌ キを 含 む イ ヌ科動 物 の血液 中 か らBartonella DNA
を高 感 度か つ特 異的に 検 出可能 なPCR
法 の確立 を 目的とし て 研究を行っ た。 特に、rpoB
領域 とssrA
領 域 の遺 伝子 を 標的と したPCR
法の 有 用性とその 諸 条件 につ い て検 討 した。ま た、 血液 検体 中 にはPCR
反 応を阻害す る物質 が存在する こと も 知ら れ ているこ と から 、それらがPCR
反応の感度にどの程度の影響を及ぼす か につ い ても同時 に 検討す る ことと し た。22
2.2
材料お よび方 法2.2.1
タヌ キBartonlla DNA
陽 性 株を用 い た予備 実験-
70
℃で 保管し て いたタ ヌ キ血液 を 室温で 解凍したの ち、DNeasy Blood and Tissue Kit( QIAGEN
社) を 用い てDNA
を抽出 した。 タヌ キ 血液200µl
か ら 抽 出し たDNA
を 無 作為 に選 択 し 、 後述のrpoB
領域を標 的とし たPCR
法[Renesto et al., 2001]
でBartonella DNA
の検 出 を行っ た。 本PCR
法で 陽性とな っ たRD198
株、443
株 、474
株を用 い て 、さらにgltA
およ びssrA
領域のPCR
法 を 実施 し、それぞれの 増幅産 物 の 泳動像 を 比 較検討 した。なお、陽性コント ロ ー ルに は、B. arsatica IBS 382
T株 を用い た 。2.2.2
検出 感度 測定 用の プ ラスミ ドDNA
の 作 製a)
使用菌 株とDNA
抽出-
70℃ で凍 結保 存し てい た B. rochalimae BAA-1498
T株の 一白菌 耳量 を、5%
ウ サギ血 液加 ハート イ ンフュ ー ジョン 寒天 培地(HIA
)*1 に塗 抹 して 、35℃
、5%CO
2の気相で7~ 10
日間培養し た 。HIA
上に発育した菌株を適当量掻き取り、リ ン酸 緩衝 生理食 塩 水(
PBS) 500μl
に 懸濁 し て遠 心洗浄 した 後、InstaGene Matrix
(
Bio-Rad
社)を 用いて 菌体 か らDNA
を 抽出し た。b) B. rochalimae BAA-1498
T株のrpoB
およびssrA
領域のPCR
法 に よる増 幅Bartonella
属 菌 のrpoB
およびssrA
領 域 を増幅する プライ マー を それぞれ 使 用し た(表2-1
)。各領域 を 増 幅 するForward
、Reverse
プ ライマー(10μM
)を1μl
ず つ、DNA ポリ メ ラーゼ にTakaRa EX Taq HS( TaKaRa
社) を0.1μl、 dNTP
溶 液を1.6μl
、反応 用buffer
の2 μl
をそれ ぞ れ 加 え、Nuclease Free Water
(以 下 、23
NFW
)13.3µl
で全 量を20µl
とした(表2-2
)。各 領域のPCR
法は 表2-3
に 示す 条 件下 で行 った。 両PCR
の増 幅 産 物を3% アガロ ースゲルを 用いて それぞ れ電 気
泳 動し 、0.5µg/ml
のエ チジ ウム ブ ロ マイド で 染 色 済のゲルか ら、UV
照 射下でPCR
産 物を 切 り出した。Wizard SV Gel and PCR Clean-Up system(Promega
社)を用 い てPCR
産 物を 精製 し、 これ をイ ンサー ト 用DNA
とした 。c) PCR
検 出感度測 定 用 の プラス ミ ドDNA
の 作製pGEM T-easy Vector
シ ステム(Promega 社)を用い て 、添付のプロトコー ル に従 っ てインサート 用DNA
とpGEM T-easy Vector
を ラ イゲーショ ンし、コ ン ピ テン ト セル(大 腸菌DH5α
株 )を 形質転 換 した 。2% X-Gal
(Invitrogen
社) を40μl
、200mM Isopropyl β-D-1-thiogalactopyranoside
(IPTG, Invitroge
社)の40μl
を 塗 付し た アンピシ リ ン含 有Luria Bertani( LBamp)寒天培地*
2に形 質転換 した 菌 を塗 沫し 、37℃
で16
時間、好気培養 し た 。培地上 に発育 し たコロニー を3
~5
個 釣菌し、PCR
法 によ って 目 的のDNA
が挿入 さ れたプ ラスミドが 大腸菌 内に 存在 す るこ とを 確認し た 。なお 、PCR
法 には、 プ ラスミ ドDNA
のSP6
およ びT7
プ ロ モー タ ー領域 に それ ぞれ 結 合するSP6
プ ライマー (5’-TATTTAGGTGACACT ATAG-3’)と T7
プ ラ イマー (5’-TAATACGACTCACTATAGGG-3´) を使用 した 。 PCR
反応液の組成は表2-2
に、反応条件は表2-3
にそれぞれ示すとおりである。形 質転 換済 の 大腸 菌 を
LBamp
液体培 地*
3に接種 し、37℃
で 一晩激 しく 振 とう培養した。培 養 した 大 腸 菌からWizard Plus Minipreps DNA Purification System
(Promega
社)を用いて , プラスミドDNA
を 抽 出 した。すな わち、 培養液3ml
を1,930
×g
、10
分間 遠心分 離(EX-136
:TOMY
社)した後、上清を除去し た 。大 腸 菌の 沈 査にCell Resuspension Solution 250µl
を加 えて混和し 、1.5ml 尖 底プ ラ ス チッ ク チューブ に 移した 。Cell Lysis Solution
の250µl
を加えて転 倒混和 し、24
さ らに
Alkaline Protease Solution
を10µl
加 え て転倒混 和した 後、室温で5
分間 静 置し た 。反応後 、Neutralization Solution
の350µl
を加えて転倒混和し、15,700
×g で
1
分間 遠心 分離(5415R:Eppendorf
社)した。次いで、ミニカラムのメン ブ レン 上 にColumn Wash Solution
の750µl
を 加えて15,700×g
で1
分間遠 心洗 浄 し、 こ の洗 浄工程 を2
回繰り返した 。メン ブ レンを 洗浄後、 ミニカラムを滅菌1.5ml
マイ クロ チュ ーブに 装 着し た。NFW
を30µl
加えて 室温で1
分間静置した後 、
15,700×g
、1
分 間遠 心分 離し 、 精 製 済 の プラスミ ドDNA
溶 液 を回収 した 。NanoDrop 1000 Spectrophotometer (Thermo Scientific
社)を用いて rpoB
領 域あ る いはssrA
領域がそれ ぞれ 組 み込 ま れ たプラ ス ミドDNA
溶液 の 濃 度を測 定し た 後 、NFW
を用いて 両 プラス ミ ド溶液 と もに6×10
7copy/µl
になるよう 濃度を 調整 し た。d
)プ ラ スミ ドDNA
溶 液の コピ ー数の 調 整2.2.2-c
)で作製したプラスミドDNA
溶液(6×10
7copy/µl)を原液として、
6×10
7copy/µl
~ 6×10-1copy/µl
に な るようNFW
を用い て10
倍 階段希釈し た 。 続 いて 、NFW
を45µl
ずつ 入れ た9
本の1.5ml
チ ューブ それ ぞれ に、プラスミドDNA
溶液 の各希釈系 列 から分 取 した5µl
を加えてよく混合し、これらをPCR
法 の 検出 感 度の測定 に 用いる 希 釈列( テ ン プレートDNA
① ) とし た(図2-1
)。また、血液中に存在する
PCR
阻 害 物質の影 響を検 討する ために、犬血液 の抽出DNA
液を加えた希釈列を 作 製 した。 す なわち 、 あらかじめBartonella
の 感染が陰性 であるこ とを確認し て おいた ビーグル犬 から採取 した200µl
の血 液 か ら、DNeasy Blood and Tissue Kit
(QIAGEN
社)を用い てDNA
を45µl
抽 出し 、 こ れを 希 釈列作製 用の イ ヌ血 液抽 出 液 と した 。イ ヌ血 液抽 出液を
45µl
ずつ 入 れた1.5ml
のチュー ブ9
本に 、 プラス ミ ド25
DNA
溶 液 の各 希釈 列か ら分 取した5µl
を加 え て混合し、 これら を イヌ科動物の 血 液か らBartonella DNA
を検 出 するPCR
法の検出感度の測定に用いる 希釈 列( テン プ レート
DNA② )と し た (図 2-2)。
26
表
2-1. PCR
に用 い たプラ イ マーの 塩 基配列 と 増幅塩 基長表
2-2
.PCR
に用 い た試薬の 組成×10 Ex Taq Buffer( Takara
社)2.0 µl
dNTP Mixture(TaKaRa
社)1.6 µl
TaKaRa Ex Taq HS(TaKaRa
社)0.1 µl
DNA
溶液 (20ng/μl) 1.0 µl
10µM
各 プラ イマ ー 各1.0 µl
Nuclease Free Water 13.3 µl
計
20.0 µl
表
2-3. gltA、 rpoB
お よ びssrA
領域 のPCR
の反応 条 件標 的 遺 伝 子 反 応 温 度 (℃) 時 間 ( 秒 ) 回 数
gltA
熱変 性 増幅 伸長 反応
熱変 性
アニ ーリ ン グ 伸長
94 94 53 72 72
300 20 30 30 300
1 35 1
rpoB
熱変 性 増幅 伸長 反応
熱変 性
アニ ーリ ン グ 伸長
94 94 53 72 72
120 30 30 60 120
1 35 1
ssrA
熱変 性 増幅 伸長 反応
熱変 性
アニ ーリ ン グ 伸長
94 94 53 72 72
120 30 30 60 120
1 35
1
標 的 遺 伝 子 プ ラ イ マ ー 塩 基 配 列 増 幅 塩 基 長
(bp)
gltA BhCS.781p 5’-GGGGACCAGCTCATGGTGG-3
BhCS.1137n 5’-CGATGCACAATCATTTCTGG-3’ 379
rpoB 1400F 5´-CGCATTGGCTTACTTCGTATG-3´
2300R 5´-GTAGACTGATTAGAACGCTG-3´ 893 ssrA ssrA-F 5’-GCTATGGTAATAAATGGACAATGA-3´
ssrA-R 5´-GTAGACTGATTAGAACGCTG-3´
301
27
図
2-1
テンプレートDNA
① の 作製過 程血 液中 の阻害物質の影 響 を受け ず に
Bartonella DNA
を検 出するPCR
法の 感 度測 定 系プラスミド DNA を 10 倍階段希 釈
6 × 10
7copy/µl
6× 10
6copy/µl
6 × 10
5copy/µl
6 × 10
4copy/µl
6×10
3copy/µl
6×10
2copy/µl
6 × 10
1copy/µl
6×10
0copy/µl
6 × 10
-1copy/µl
Nuclease Free Water 45µl
5µl ずつ添加
6 × 10
6copy/µl
6 × 10
5copy/µl
6 × 10
4copy/µl
6×10
3copy/µl
6 × 10
2copy/µl
6 × 12
1copy/µl
6×10
0copy/µl
6 × 10
-1copy/µl
6 × 10
-2copy/µl
28
図
2-2
テンプレートDNA
②の 作 製過 程イ ヌ血 液 中から
Bartnella DNA
を検出 す るPCR
法 の感度測定 系プラスミド DNA を 10 倍階段希 釈
6 × 10
7copy/µl
6 × 10
6copy/µl
6 × 10
5copy/µl
6×10
4copy/µl
6 × 10
3copy/µl
6 × 10
2copy/µl
6×10
1copy/µl
6 × 10
0copy/µl
6×10
-1copy/µl
イヌ血液抽出液 45µl
5µl ずつ添加
6 × 10
6copy/µl
6 × 10
5copy/µl
6 × 10
4copy/µl
6×10
3copy/µl
6 × 10
2copy/µl
6 × 12
1copy/µl
6×10
0copy/µl
6 × 10
-1copy/µl
6 × 10
-2copy/µl
29
2.2.3 PCR
法の検出 感 度の測 定2.2.2-d
)で作製したテンプレー トDNA①および DNA② を用い、 Bartonella
のrpoB
領 域とssrA
領 域を増 幅 するPCR
法の検出感度を測定した。すなわち、PCR
反 応 用チ ュー ブ 内に,6×10
6~6×10-2copy/µl
に希釈した テンプ レートDNA
① ある い は
DNA②を 1μl、Forward
およびReverse
プライマー(10µM)をそれぞ れ1μl
、DNA
ポリ メラー ゼ のTakaRa EX Taq HS
(TaKaRa
社 )を0.1μl
、dNTP
溶 液を1.6μl
、反応 用buffer
を2 μl
そ れぞれ 加 え 、PCR
反応 を行 った。PCR
産物 を 電 気泳 動 し、染色 ・ 脱色し た 後、UV
照 射 下で増幅 バンド の有 無を確認し た 。各 遺伝 子領 域に 対 して 、 上 記 の
PCR
法 を7
回 ずつ行い、4
回以 上陽 性を 示 し、 かつプラスミドDNA
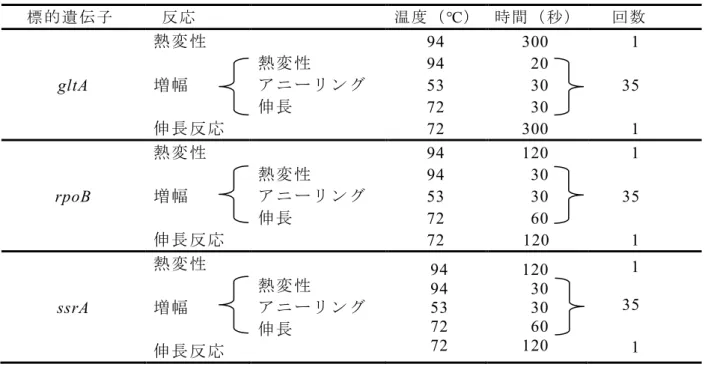
の コピ ー 数が最 も 少ない 希釈 を検出限界 とした 。30
2.3
成 績2.3.1
タ ヌキBartonlla DNA
陽 性 株を 用 いた予備 実験gltA
、rpoB
、ssrA
の各 領域 を 標 的とし たPCR
法にお ける予 備実験の結果、gltA
領 域 を 標 的 と し たPCR
産 物 の 泳 動 像 に は 、 特 異 的 と さ れ る 増 幅 バ ン ド(379bp)
周辺 に非 特異 反応 が多 く 出現 し 、判別 が 困難であった。一方、rpoB お よ びssrA
領域 の 泳動 像で は、 目的 と する増 幅 領域周辺 に 非 特異反応は 少なく 、特 異 的なバ ンドの 判 別は 容 易 で あった ( 図2-3)。
2.3.2
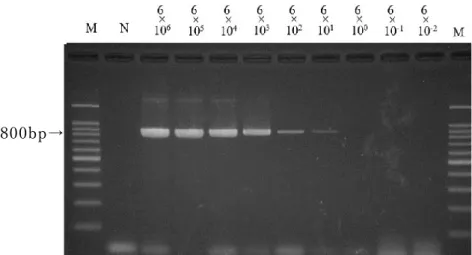
テ ンプレートDNA
①を用いたPCR
法の検 出感度rpoB
領域 を標 的と し たPCR
法で は 、7
回 実 施したうち の4
回に おい て6×10
1copy/tube
まで800bp
付 近 に明瞭 な 特 異 バンド が検出され た (図2-4)。
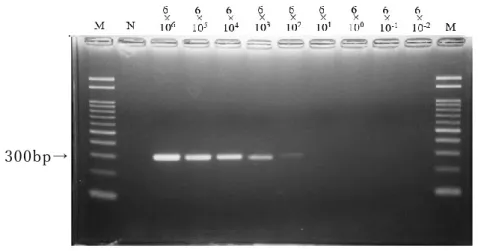
ssrA
領 域を 標的 とし たPCR
法 で は 、7
回実 施した うちの4
回に おい て6×10
2copy/tube
まで300bp
付 近 に明瞭 な 特 異 バンド が検出され た (図2-5)。
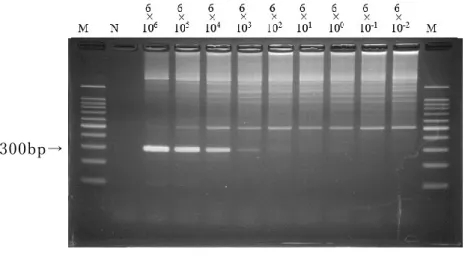
2.3.3
テン プレ ートDNA②を用 い た PCR
法 の 検出感 度rpoB
領域 を標 的と し たPCR
法で は 、7
回 実 施したうち の4
回に おい て6×10
3copy/tube
まで800bp
付 近 に明瞭 な 特 異 バンド が検出され た (図2-6)。
ssrA
領 域 を 標 的 と し たPCR
で は 、7
回 実 施 し た う ち の4
回 に お い て6×10
2copy/tube
まで300bp
付 近 に明瞭 な 特 異 バンド が検出され た (図2-7
)。31
A
B
C
図
2-3
タヌキDNA
溶液 のgltA
(A
)、rpoB
(B
)、ssrA
(C
)領域 を 増幅するPCR
泳 動像の 比較M: 100bp DNA
マ ーカ ー、N:
陰 性 コント ロ ール、P:
陽性コ ントロ ール、1; RD198
株、2: RD443 株、3: RD474
株800bp→
300bp→ 300bp→
32
M: 100bp DNA
マーカ ー 、N:
陰 性コント ロール図
2-4
テンプレートDNA①を 用 いた rpoB
領域にお けるPCR
法の 検 出感 度測定800bp→
33
M: 100bp DNA
マー カ ー、N:
陰性コン トロー ル図
2-5
テンプレートDNA①を用 い た ssrA
領域に おけるPCR
法の 検 出感 度測定300bp→
34
M: 100bp DNA
マ ー カー、N:
陰性コ ントロ ール 図2-6
テンプレートDNA② を 用いた rpoB
領域にお けるPCR
法の 検 出感 度測定800bp→
35 300bp→
M: 100bp DNA
マ ー カー、N:
陰性 コント ロール図