発光キノコ、ヤコウタケ生物発光機構研究
森 憲一
電気通信大学
2011 年 3 月
発光キノコ、ヤコウタケ生物発光機構研究
森 憲一
電気通信大学大学院 電気通信学研究科
博士(理学)の学位申請論文
2011 年 3 月
発光キノコ、ヤコウタケ生物発光機構研究
博士論文審査委員会
主査 丹羽 治樹 教授
平野 誉 准教授
中村 整 教授
白川 英樹 准教授
狩野 豊 准教授
著作権所有者
森 憲一
2011 年
Study on Bioluminescence mechanism of luminous fungus Mycena chlorophos
Kenichi Mori
ABSTRACT
Bioluminescent fungi are widely distributed on land and most belong to the class of Basidomycetes. Light of about 530 nm wavelength maximum is emitted continuously.
Despite many efforts to determine the mechanism of light emission in fungi, the detailed molecular basis for the light-emitting process remains unclear.
Mycena chlorophos (yako-take in Japanese) is distributed in subtropical areas in Japan including on Hachijo Island and the Bonin (Ogasawara in Japanese) Islands. In nature, the fruiting bodies of M. chlorophos develop mainly on decaying petioles of a palm Phenix roeberenii, and only on the day after rain falls during the rainy season (June/July and September/October). The limited distribution and the limited season for the formation of the fruiting bodies have made the chemical study of the bioluminescence mechanism difficult.
Cultivation of the fruiting bodies using peat moss as a solid medium was studied and we found that the peat moss medium was satisfactory for the production of fruiting bodies of M. chlorophos.
The characteristics of the bioluminescence by using cultivated fruiting bodies of M.
chlorophos was investigated under various conditions to obtain valuable information for the isolation of the luciferin and luciferase.
Long-lasting light emission was observed only in fresh fruiting bodies, not in the frozen and freeze-dried samples.
Although clear correlation between the light emission intensity of the fruiting bodies
and the buffer pH was not observed, the pH change of acetate and citrate buffers gave
the reverse effect against the light emission intensity. A marked change in light intensity
was observed in acetate buffer: the light intensity in acetate buffer at pH 4 decreased
rapidly within 10 min, while the initial light intensity of the pileus was maintained for a
sodium acetate did not. The light emission of the fresh fruiting bodies could be stopped
with pH 4 acetate buffer, and could be recovered at pH 6. Thus, the luciferin might be
isolated from the fruiting bodies whose the light-emission was inhibited.
発光キノコ、ヤコウタケ生物発光機構研究
森 憲一
和文概要
ヤコウタケは日本の八丈島など亜熱帯気候の地域に分布している発光キノコ である。子実体からの発光量が多く、
1~3
日間持続して緑色に発光する。ヤコ ウタケなどの発光菌類は発光機構について未解明な部分が多い。今回、発光機 構解明研究の一環として、ヤコウタケ子実体の大量確保のための実験室栽培を 試みた。その結果ヤコウタケ子実体の簡易栽培法を確立し、研究材料の大量確 保に目処を付けることができた。栽培した子実体の発光特性を検討した所、子 実体の発光を可逆的に制御することができることを見出し、ヤコウタケ生物発 光に関係する物質(ルシフェリン)の単離に道を拓くことが出来た。目 目 目
目 次 次 次 次
第 第
第 第1 1 1 1章 章 章 章 序論 序論 序論 序論 …1-18
1 1 1
1----1. 1. 1. 1. 発光生物 発光生物 発光生物 発光生物 … 1-3
1 1 1
1----1 1----1. 1 1 1. 1. 1.
発光生物発光生物発光生物発光生物とと人間とと人間人間人間とのとのとのとの関係関係関係 関係1
1 1
1----1 1 1----2. 1 2. 2. 2.
発光生物の発光生物発光生物発光生物ののの種類種類種類種類1 1 1
1----1 1----3. 1 1 3. 3. 3.
発光生物発光生物発光生物発光生物がが発光がが発光発光発光するするするする目的目的目的 目的1 1
1 1----2. 2. 2. 2. 生物発光 生物発光 生物発光 生物発光 …4-12
1 1----2 1 1 2 2----1. 2 1. 1. 1.
生物発光研究生物発光研究生物発光研究生物発光研究のののの歴史概略歴史概略歴史概略歴史概略1 1----2 1 1 2 2----2. 2 2. 2. 2.
ルシフェリン-ルシフェリンルシフェリンルシフェリン---ルシフェラーゼルシフェラーゼルシフェラーゼルシフェラーゼ反応反応反応反応1 1----2 1 1 2 2----3. 2 3. 3. 3.
ホタルホタルホタルホタルのののの生物発光生物発光生物発光生物発光1 1----2 1 1 2 2----4. 2 4. 4. 4.
ウミホタルウミホタルウミホタルウミホタルのののの生物発光生物発光生物発光生物発光1
1 1
1----2 2----5. 2 2 5. 5. 5.
オワンクラゲオワンクラゲのオワンクラゲオワンクラゲののの生物発光生物発光生物発光生物発光::: : 発光発光発光発光タンパクタンパク質タンパクタンパク質質質イクオリンイクオリンイクオリンイクオリンとととと緑色蛍光緑色蛍光タンパク緑色蛍光緑色蛍光タンパクタンパク質タンパク質質質
GFP GFP GFP GFP
1 1 1 1----2 2 2 2----6. 6. 6. 6.
生物発光の生物発光生物発光生物発光ののの特徴特徴特徴特徴ととととアプリケーションアプリケーションアプリケーションアプリケーション1 1 1
1----3. 3. 3. 3. 発光菌類 発光菌類 発光菌類 発光菌類 (luminous ( ( ( luminous luminous fungi) luminous fungi) fungi) fungi) とその研究 とその とその とその 研究 研究の 研究 の の歴史 の 歴史 歴史 歴史 … 13-15 1
1 1
1----4. 4. 4. 4. ヤコウタケ ヤコウタケ ヤコウタケ ヤコウタケ (((( Mycena Chlorophos Mycena Mycena Mycena Chlorophos Chlorophos Chlorophos )))) … 16-17
1 1 1
1----5. 5. 5. 5. 本研究 本研究の 本研究 本研究 の目的 の の 目的 目的 目的 … 18
第 第
第 第 2 2 2 2 章 章 章 章 本論 本論 本論 本論 …19-66
2 2----1. 2 2 1. 1. 1. ピートモス ピートモス ピートモス ピートモスを を を培地 を 培地として 培地 培地 として として として用 用 用 用いた いたヤコウタケ いた いた ヤコウタケ ヤコウタケの ヤコウタケ の の の 改良栽培法
改良栽培法 改良栽培法
改良栽培法の の の の確立 確立 確立 確立 2
2 2
2----2. 2. 2. 2. ヤコウタケ ヤコウタケ子実体 ヤコウタケ ヤコウタケ 子実体 子実体 子実体の の発光 の の 発光 発光の 発光 の の特徴 の 特徴 特徴 特徴 … 21-24
…19-20
2 2
2----2 2 2 2----1. 2 1. 1. 1.
ヤコウタケヤコウタケのヤコウタケヤコウタケの暗闇中のの暗闇中暗闇中暗闇中のののの発光発光発光発光2
2 2
2----2 2 2----2. 2 2. 2. 2.
ヤコウタケ子実体ヤコウタケヤコウタケヤコウタケ子実体子実体子実体のののの発光経時変化発光経時変化発光経時変化発光経時変化2 2
2----2 2 2 2----3. 2 3. 3. 3.
ヤコウタケヤコウタケ子実体ヤコウタケヤコウタケ子実体の子実体子実体ののの発光発光発光発光スペクトルスペクトルスペクトル スペクトル2
2 2
2----2 2 2----4. 2 4. 4. 4.
マイクロスコープマイクロスコープをマイクロスコープマイクロスコープををを用用いた用用いたいたヤコウタケいたヤコウタケヤコウタケ細胞ヤコウタケ細胞の細胞細胞ののの観察観察観察観察2 2 2
2----3. 3. 3. 3. ヤコウタケ ヤコウタケの ヤコウタケ ヤコウタケ の の の状態 状態による 状態 状態 による による による発光量変化 発光量変化 発光量変化 発光量変化 … 25-29
2 2
2 2----3 3 3----1. 3 1. 1. 1.
凍結乾燥凍結乾燥ヤコウタケ凍結乾燥凍結乾燥ヤコウタケのヤコウタケヤコウタケののの調製調製調製と調製とと発光反応と発光反応発光反応 発光反応2
2 2
2----3 3 3----2. 3 2. 2. 2.
ヤコウタケヤコウタケ組織ヤコウタケヤコウタケ組織組織組織をを破砕をを破砕破砕破砕したしたした状態した状態での状態状態でのでの発光での発光発光発光2 2 2
2----4. 4. 4. 4. ヤコウタケ ヤコウタケの ヤコウタケ ヤコウタケ の の の発光 発光 発光 発光と と酸素 と と 酸素 酸素の 酸素 の の関係 の 関係 関係 関係 … 30-33 2
2 2
2----5. 5. 5. 5. ヤコウタケ ヤコウタケ発光 ヤコウタケ ヤコウタケ 発光 発光 発光を を持続 を を 持続 持続 持続させる させる させる最適温度 させる 最適温度 最適温度 最適温度 … 33-34 2
2 2
2----6. 6. 6. 6. 凍結乾燥 凍結乾燥ヤコウタケ 凍結乾燥 凍結乾燥 ヤコウタケ ヤコウタケ ヤコウタケにおける における における における ルシフェリン
ルシフェリン ルシフェリン
ルシフェリン‐ ‐ ‐ルシフェラーゼ ‐ ルシフェラーゼ ルシフェラーゼ反応確認 ルシフェラーゼ 反応確認 反応確認 反応確認 … 35-37 2
2 2
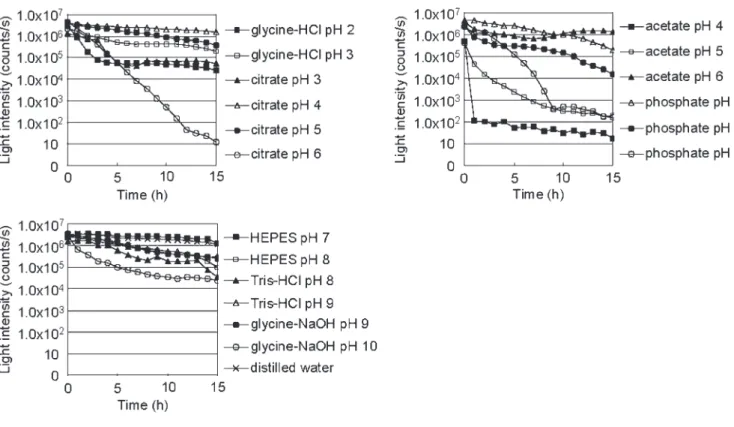
2----7. 7. 7. 7. 生 生 生 生の の の のヤコウタケ ヤコウタケ切片 ヤコウタケ ヤコウタケ 切片 切片を 切片 を を を用 用 用 用いた いた いた いた種 種々 種 種 々 々 々の の の の緩衝液 緩衝液 緩衝液 緩衝液、 、 、 、 pH pH pH pH 中 中での 中 中 での での での ヤコウタケ
ヤコウタケ ヤコウタケ
ヤコウタケ発光経時変化 発光経時変化 発光経時変化 発光経時変化の の の の測定 測定 測定 測定 2
2 2
2----8. 8. 8. 8. 酢酸緩衝液中 酢酸緩衝液中での 酢酸緩衝液中 酢酸緩衝液中 でのヤコウタケ での での ヤコウタケ ヤコウタケ ヤコウタケ発光 発光 発光 発光 … 41-45 2----8 2 2 2 8 8----1. 8 1. 1. 1.
酢酸酢酸、酢酸酢酸、、酢酸、酢酸酢酸酢酸Na Na Na Na
中中での中中でのヤコウタケでのでのヤコウタケヤコウタケヤコウタケ発光量変化発光量変化発光量変化発光量変化2 2 2----8 2 8 8----2. 8 2. 2. 2.
異異なる異異なるなるなるpH pH pH pH
のののの酢酸緩衝液酢酸緩衝液酢酸緩衝液酢酸緩衝液をを用をを用用いた用いたいたいた ヤコウタケヤコウタケヤコウタケ
ヤコウタケ発光発光発光発光ののののスイッチングスイッチングスイッチングスイッチング
2 2 2 2----8 8 8----3. 8 3. 3. 3.
酢酸処理前後酢酸処理前後の酢酸処理前後酢酸処理前後のヤコウタケののヤコウタケヤコウタケヤコウタケ発光発光発光スペクトル発光スペクトルスペクトルのスペクトルののの測定測定測定測定2 2 2
2----9. 9. 9. 9. ヤコウタケ ヤコウタケ発光 ヤコウタケ ヤコウタケ 発光 発光 発光への への各種添加物 への への 各種添加物 各種添加物の 各種添加物 の の の効果検証 効果検証 効果検証 効果検証 … 46-54 2 2 2 2----9 9 9----1. 9 1. 1. 1.
ヤコウタケヤコウタケ発光ヤコウタケヤコウタケ発光への発光発光へのへのへのカルボンカルボンカルボンカルボン酸類酸類酸類の酸類ののの添加添加添加添加…38-40
2----9 2 2 2 9 9----3. 9 3. 3. 3.
ヤコヤコウタケヤコヤコウタケウタケ発光ウタケ発光への発光発光へのへのへのアミンアミンアミンアミン類類類の類のの添加の添加添加 添加2 2 2 2----9 9 9----4. 9 4. 4. 4.
ヤコウタケ発光ヤコウタケヤコウタケヤコウタケ発光発光発光へのへのへのへの添加剤効果添加剤効果添加剤効果添加剤効果のまとめのまとめのまとめ のまとめ2 2 2
2----10. 10. 10. 10. ヤコウタケ ヤコウタケ ヤコウタケ ヤコウタケ発光 発光に 発光 発光 に に に対 対 対 対する するキレート する する キレート キレート剤 キレート 剤 剤の 剤 の効果 の の 効果 効果 効果 … 55-59 2
2 2
2----11. 11. 11. 11. ヤコウタケ ヤコウタケ ヤコウタケ ヤコウタケ発光物質可溶化 発光物質可溶化 発光物質可溶化の 発光物質可溶化 の試 の の 試 試 試み み み み … 60-66 2 2 2----11 2 11 11----1. 11 1. 1. 1.
界面活性剤界面活性剤を界面活性剤界面活性剤を用をを用用用いたいたいたいたヤコウタケヤコウタケヤコウタケヤコウタケ発光可溶化試験発光可溶化試験発光可溶化試験 発光可溶化試験2 2 2----11 2 11 11----2. 11 2. 2. 2.
酵素酵素を酵素酵素を用をを用用いた用いたいたいたヤコウタケヤコウタケヤコウタケヤコウタケ発光可溶化試験発光可溶化試験発光可溶化試験発光可溶化試験第 第
第 第3 3 3 3章 章 章 章 結語 結語 結語 結語 … 67-69
第 第
第 第4 4 4 4章 章 章 章 実験 実験 実験 実験の の部 の の 部 部 部 … 70-92
4 4----1. 4 4 1. 1. 1. 一般操作 一般操作 一般操作 一般操作 … 70-73
4 4----2. 4 4 2. 2. 2. 第 第 第2 第 2章 2 2 章 章の 章 の の実験方法 の 実験方法 実験方法 実験方法 … 74-92
参考文献 参考文献 参考文献
参考文献 … 93-95
謝辞 謝辞 謝辞
謝辞 …96
第 第
第 第1 1 1 1章 章 章 章 序論 序論 序論 序論
1 1
1 1----1. 1. 1. 1. 発光生物 発光生物 発光生物 発光生物
(1)(2)1 1
1 1----1 1 1 1----1. 1. 1. 1.
発光生物発光生物発光生物発光生物とと人間とと人間人間人間とのとのとの関係との関係関係 関係発光生物が生み出す可視光は熱を放出しない冷光であるという特徴を持っている。暗 闇のなかに浮かび上がる発光生物の光は古代から人類にとって興味深いものであった ため、多くの研究者が発光生物の調査、研究を行ったという記録がある。(3)ある種の発 光キノコや朽木に発生した発光バクテリアが夜に淡く発光するのを古代ギリシアの哲
学者
Aristotle(384-322 B.C.)が記述している。ローマ時代の政治家であり博物学者であ
った
Gaius Plinius Secundus
はカモメ貝やある種のクラゲが発光するのを記録している。また、Robert Boyle(1627-1691)は真空ポンプを用いて朽木の発光(菌類の発光)
には空気が必要であると示している。生物発光の近代的な研究は
19
世紀からになりそ れについては後述するが、生物発光の光は古代から我々人類を魅了し続けている。1 1
1 1----1 1 1 1----2. 2. 2. 2.
発光生物の発光生物発光生物発光生物ののの種類種類種類種類地球上には数多くの発光生物が存在することが知られている。例えばバクテリア、渦 鞭毛藻類(海洋プランクトン)、菌類(発光キノコ)、腔腸動物(クラゲなど)、環形動 物(ミミズ、ゴカイなど)、甲殻類(ウミホタル、エビ)、軟体動物(ラチア)、昆虫類
(ホタル、鉄道虫、ツチボタル:アラキノカンパ)などである。中でも有名な生物とし てホタル(Fig.1-1)、ウミホタル(Fig.1-2)などが挙げられる。発光する生物の大きさも小
さいものではバクテリア、細菌類、大きいものでは甲殻類、昆虫など様々である。発光 生物の生息地は海か陸かに大別できる。種としては海に生息しているものが多く、特に 深海では未知の発光生物が多数生息していると考えられている。
生存環境が違うように発光方法にも違いがある。ホタルやオワンクラゲ
(Fig.1-3)
の様 に発光器官を持ち特定部分で発光するもの、ウミホタルや発光巻貝ラチア(Fig.1-4)など の発光液を体外に分泌するものなどがある。また、生物発光は生物種自らが発光する一 次発光と寄生もしくは共生生物が発光する二次発光に分けることができる。先に挙げた 発光生物は一次発光である。二次発光の生物の例としてはマツカサウオなどがあり、二 次発光生物の発光器の中で発光バクテリアが共生することで発光している。発光の様式も二種類あり、両者はフラッシュ発光とグロー発光と呼ばれている。フラ ッシュ発光は比較的強い光を短時間で放出する発光であり、ホタルや渦鞭毛藻などの発 光生物で見られる。それに対してグロー発光はある程度の発光量を保ちながら長時間持 続的に発光する様式で、バクテリアや発光キノコなどに見られる。
1 1
1 1----1 1 1 1----3. 3. 3. 3.
発光生物発光生物発光生物発光生物がが発光がが発光発光発光するするする目的する目的目的 目的生物発光の進化的な起源は細胞毒性のある活性酸素の除去であったと考えられてい る。その後の進化過程で発光システムが様々な用途に利用されるようになったと考察さ れている(Table.1-5)。例を挙げると、ホタル、ホタルイカ、発光ゴカイなど雌雄間の合 図として発光すると考えられている。発光キノコは光で虫を誘引して胞子を遠方に拡散 させる。ウミホタルなどは外敵への威嚇のために発光する。ハダカイワシなどは自分の 影を消して敵に見つからないように発光する(
Counter shading
)。多くの深海性発光 魚は眼の付近に発光器を有していて視界を確保する。チョウチンアンコウやアラキノカ ンパなどは発光を疑似餌として用いて捕食動物を誘引している。このように発光生物は様々な目的で発光を利用している。
発光の目的 代表的な生物
求愛行動 ホタル、ホタルイカ、発光ゴカイ、ウミホタル
胞子の拡散(媒介者へのアピール) 発光キノコ
威嚇、おとり、忌避行動 ウミホタル、発光ムカデ、ウミサボテン、エダウミウシ
カウンターシェーディング(影を消す) サクラエビ、ハダカイワシ、ミッドシップマン
照明 深海性発光魚
疑似餌 チョウチンアンコウ、アラキノカンパ
Fig.1-1
ホタルFig.1-3
オワンクラゲFig.1-2
ウミホタルFig.1-4
ラチア 写真提供:産業技術総合研究所近江谷克裕博士 写真提供:産業技術総合研究所 近江谷克裕博士
1 1
1 1----2. 2. 2. 2. 生物発光 生物発光 生物発光 生物発光
(1)(2)1 1
1 1----2 2 2 2----1. 1. 1. 1.
生物発光研究の生物発光研究生物発光研究生物発光研究のの歴史概略の歴史概略歴史概略 歴史概略神秘的な輝きを放つ発光生物は多くの研究者の興味を引き長年にわたり研究されてき た。生物発光の物質的、化学的な研究は
19
世紀後半に始まった。1885
年にDubois
は、ヒカリコメツキ虫(
Phyrophorus )の発光器を熱水抽出した溶液の上清と、発光器を冷水
抽出してから発光が認められなくなるまで放置した溶液を調製し、両者を試験管内で混 ぜることによって再び発光することを確認した。熱水抽出液には熱に安定な基質(ルシ フェリン)
、冷水抽出液には酵素(
ルシフェラーゼ)
を含んでいると考えた。そこで、これ らを混ぜた時に起こる発光反応をルシフェリン-ルシフェラーゼ反応(L-L反応)と名付けた(4)(5)。その後、生物発光研究の進歩に寄与したのは
E. Newton Harvey (1887-1959)
である。
Harvey
は生物発光の魅力にとりつかれ世界を渡り歩き、発光生物にはL-L
反応を示すものと示さないものがあることを見出した。
L-L
反応を示す生物例としてはホ タル類やヒカリコメツキ等の発光甲虫、海洋甲殻類のウミホタル、多毛類の発光ゴカイ などであり、示さないものは発光クラゲやホタルイカ、発光キノコ類などであった。そ れらの研究結果を元に生物発光に関する300
を超える出版物を執筆した。その中でも1952
年に出版された著書『Bioluminescence
』(6)は長く生物発光のバイブルとみなされ てきた。他にも様々な研究者が世界に生息する発光生物の研究を行ってきた。ここでは 代表的な発光生物であるホタルなどについて、生物発光機構などを説明する。1 1 1
1----2 2 2----2. 2 2. 2. 2.
ルシフェリンルシフェリン-ルシフェリンルシフェリン--ルシフェラーゼ-ルシフェラーゼルシフェラーゼ反応ルシフェラーゼ反応反応反応Dubois
が発見したL-L
反応(4)(5)は有機化学的に研究され、その化学反応の概要が明らかとなってきている。
L-L
反応ではルシフェリンがルシフェラーゼの触媒作用により 酸化され不安定中間体を形成し、この中間体が分解して発光体(
オキシルシフェリン)
の 励起分子を生成し、励起分子が基底状態に戻る際のエネルギーが、光として外部に放出 される(Fig.1-6)
。研究の初期には
Harvey
は二枚貝のルシフェリンとウミホタルのルシフェリンが異 なることから、ルシフェリンおよびルシフェラーゼは種によって異なると考えていた(6)。 その後オワンクラゲの発光基質であるセレンテラジン(coelenterazine)
は、ホタルイカ プレルシフェリン(Watasenia preluciferin)、レニラルシフェリン(Renilla luciferin)と して、クラゲ、エビ、イカ、魚類など異なる海洋生物に共通の発光基質として用いられ ていることが1970-1980
に相次いで発見された(7)。これまでに、様々な発光基質ルシフFig.1-6 L-L
反応の機構概要 ルシフェリンルシフェリン過酸化物
励起一重項状態
(S
1)
オキシルシフェリン*基底状態
(S
0)
ルシフェラーゼオキシルシフェリン
O
2発光
ルシフェリン + O2 オキシルシフェリン* オキシルシフェリン + 光
エネルギーレベル
ルシフェリン
ルシフェリン過酸化物
励起一重項状態
(S
1)
オキシルシフェリン*オキシルシフェリン*
基底状態
(S
0)
ルシフェラーゼオキシルシフェリン
O
2発光
ルシフェリン + O2 オキシルシフェリン* オキシルシフェリン + 光
エネルギーレベル
ミホタル、発光バクテリア、腔腸動物(クラゲ)、ラチア、ミミズ、オキアミ、渦鞭毛 藻などがルシフェリンの構造や発光反応機構が提唱されている発光生物である。代表的 なルシフェリンを
Fig.1-7
に示す。Cypridina luciferin
ウミホタルルシフェリン
R
1= R
2= OH
: coelenterazine(セレンテラジン)= Renilla luciferin(ウミシイタケルシフェリン)
= Watasenia preluciferin(ホタルイカプレルシフェリン)
R
1= R
2= OSO
3H
:Watasenia luciferin
(ホタルイカルシフェリン)
NH
N N
R2
R1
O
NH
N N
O
HN
NH NH2
NH
H Me
H
O CHO Me
Me
Me
Latia luciferin
ラチア(発光巻貝)ルシフェリン
Firefly luciferin
ホタルルシフェリンS
N N
HO S
H COOH
NH HN
NH HN HO
2C
CO
2H O O
R R
R = OH
: Euphausiid luciferin オキアミルシフェリンR = H
: Dinoflagellate luciferinN H O
CHO
n-C
13H
27-CHO
発光バクテリアルシフェリン陸棲ミミズ
Diplocardia
ルシフェリン1 1 1
1----2 2 2 2----3. 3. 3. 3.
ホタルホタルホタルホタルのの生物発光のの生物発光生物発光生物発光 ( (8) ( (8)8)8)発光する甲虫であるホタル(Fig.1-1)の発光は最も有名な生物発光である。1917 年
Harvey
は北米産ホタルPhotinus pyralis
の生物発光系がDubois
の提案したL-L
反応 を示すことを見出した。その後McElroy
がホタルの発光にはATP
とマグネシウムイオ ンが必要であることを確認し、その後ホタルルシフェラーゼの結晶化、ホタルルシフェ リンの単離に成功した(9)(10)。その後の研究でホタル生物発光はFig.1-8
に示すL-L
反応 であることが確認された。ホタルの生物発光の発光効率は88%
(近江谷らの最近の研 究では41%
(11))と報告されている。発光効率は①「酸化反応(1→5)の化学収率」②「励起一重項状態(
S
1)の5a*
が生成する効率」③「5a*
から蛍光を発する効率」の 掛け算から求められる値で、88%(41%)であるホタル発光反応は極めて効率よく化学
反応エネルギーを光エネルギーに変換していることがわかる。S N
S
N CO2H
HO S
N S
N CO2AMP HO
ATP, Mg2+
ルシフェラーゼ
O2 ルシフェラーゼ
AMP
光
CO2 111
1
(ホタルルシフェリン) 2222
33
33 4444
5 55
5a* a* a* (励起状態)a* 555b* 5b* b* (励起状態)b* 5555
(ホタルオキシルシフェリン)
S N
S N HO
OO O
OAMP
S N
S N O
O O O
S N
S N O
O S
N S N O
O
S N
S N HO
O
Fig.1-8
ホタル生物発光反応1 1 1
1----2 2 2 2----4. 4. 4. 4.
ウミホタルウミホタルウミホタルウミホタルのの生物発光のの生物発光生物発光生物発光 ( (12) ( (12)12)12)ウミホタル
(Fig.1-2)
は沿岸の砂地に生息する甲殻類である。敵に襲われた時などに青 く発光する発光液を放出する。1916年にHarvey
がウミホタルの発光系がL-L
反応型 であることが確認された後、様々な研究者によってウミホタル発光に関する研究が行わ れ、ウミホタルルシフェリンの構造、生物発光反応機構はFig.1-9
に示すものであると 確認された。なおウミホタル発光系の生物発光効率は約28%
である。O2 ルシフェラーゼ
CO2 CO2
6 6 6 6
(ウミホタルルシフェリン) 7777 8888
9 99
9****(励起状態) 9999
(ウミホタルオキシルシフェリン)
NH N N O
NH
HN NH2 NH
N N NH
NH
HN NH
NH2 O
N N N O OO
N N NH
O O O
N N NH
NH
HN NH
NH2 O
青色発光 青色発光
Fig.1-9
ウミホタル生物発光反応
1----2 1 1 1 2 2----5. 2 5. 5. 5.
オワンクラゲのオワンクラゲオワンクラゲオワンクラゲののの生物発光生物発光生物発光:生物発光:発光::発光発光タンパク発光タンパクタンパクタンパク質質質質イクオリンイクオリンとイクオリンイクオリンととと緑色蛍光緑色蛍光緑色蛍光タン緑色蛍光タンタンタン パクパクパクパク質質質質
GFP GFP GFP GFP
(13)(13)(13)(13)オワンクラゲ(Fig.1-3)は無色透明のクラゲで、刺激によって傘の縁が緑色に発光する 発光生物である。生物発光は
L-L
反応で発光すると考えるのが一般的であったが、オ ワンクラゲの生物発光はL-L
反応を示さなかったため、研究は難航した。1962 年にShimomura
、Johnson
などによってオワンクラゲから発光タンパク質イクオリンと緑色蛍光タンパク質(GFP)が単離され、この発光性を基に新たな生物発光の機構が提唱 された。この生物発光では、アポタンパクに発光基質が結合した発光タンパクが、金属 イオンなどの引き金物質の作用により発光する。このイクオリンのアポタンパクに結合 している発光基質が、セレンテラジンである。セレンテラジンを酸素化物の形で取り込 んでいる発光タンパク質複合体イクオリンは、カルシウムイオンが付加することで高次 構造が変化して、酸素化物から不安定中間体を形成し、それが分解する際に励起分子生 じる。クラゲの体内ではその励起状態のエネルギーが緑色蛍光タンパク質(GFP:
Green
Fluorescent Protein)
に移動して、GFP
の発色団が励起状態になり、基底状態に戻る時 に緑色の生物発光波長(510 nm)を発する(Fig.1-10)。GFP
が系内に存在しなければイク オリンからの青色発光が観測される(460 nm)
。現在、数種類の発光タンパクが腔腸動物 や原虫、ウロコムシなどから発見されている(14)。また、イクオリンと共に発見された緑 色蛍光タンパク質(GFP)は広く生物発光の分子生物学的応用に用いられ、生体内イメー ジング技術のツールとして発展に貢献し広く普及したことで、Shimomura
らが2008年に ノーベル化学賞を受賞したことは記憶に新しい。O2
アポイクオリン
(アポタンパク質)
Ca2+
Ca2+
Ca2+
Ca2+
セレンテラジン( 10101010 ) イクオリン(11111111)
12 12 12 12
励起状態の青色蛍光タンパク質(BFP*) 励起状態のセレンテラミド
(13131313****)
Ca2+
CO2 NH
N N
O OH
HO N
N N OHOO
OH
HO
N N NH
O O O
HO
OH
N N NH
O
O
OH
N N
O O
NH HO
HO NH O H
H Ca2+
Ca2+
励起状態のBFP**** 13
1313 13****
Ca2+
Ca2+
基底状態基底状態 基底状態基底状態ののののBFPBFPBFPBFP
1313 1313
青色発光
励起状態励起状態
励起状態励起状態ののののGFPGFPGFPGFP****
基底基底基底
基底状態状態状態の状態ののGFPのGFPGFPGFP
エネルギー移動
緑色蛍光1414 1414 14 14 14 14**** N
N NH O
O
OH N N
O O
NH O
HO NH O H
H
N N NH
O
HO
OH
**
**
1 1----2 1 1 2 2----6. 2 6. 6. 6.
生物発光の生物発光生物発光生物発光ののの特徴特徴特徴特徴ととととアプリケーションアプリケーションアプリケーションアプリケーション生物発光の最大の特徴は発光量子収率の高さである。電子材料として最近注目されてい る発光ダイオードで約
4%、有機 EL
材料で約7%、また一般的な化学発光では 0.1%以
下の量子収率であるが、生物発光の量子収率はウミホタルでは28
%,
ホタルでは88
%(41%(11))と桁違いに良いことが分かる。この理由としては、発光効率が①「ルシフ ェリン酸化反応の化学収率」②「励起一重項状態(
S
1)が生成する効率」③「励起一重 項状態から蛍光を発する効率」の掛け算から求められる値であり、生物発光反応はいず れの効率も高いことがわかる。特に①の効率が化学発光などと比較して高いため、極め て効率よく化学反応エネルギーが光エネルギーに変換されている。その結果、熱によっ て放出される白熱光とは異なり、生物発光は熱の発生をほとんど伴わない発光=冷光で 発光する。そのため生物発光系は、地上における最も効率のよいエネルギー変換系の一 つであると言われている。このような利点から、現在生物発光のアプリケーション研究 が盛んに行われている(Fig.1-11)。生物発光の化学的な研究は
20
世紀に大きく進んだ。今日では生物発光反応は様々な 分野での分析ツールとして用いられている。例えばホタルの生物発光系はATP
の検出 や細胞内の遺伝子発現のプロモーターアッセイに用いられている。オワンクラゲのエク オリンはカルシウムイオンとの結合をトリガーとして発光する発光タンパクである。そ のため、生体内で信号物質として使われている細胞内カルシウムイオン濃度のモニタリ ングに用いられている。ウミホタルルシフェリンのアナログは活性酸素の検出試薬とし て用いられている。加えてエクオリンと同時に発見されたGFP
は蛍光性のあるタンパ ク質であり、バイオイメージングツールとして生化学の分野で広く用いられている。繋がった。このような生物発光を用いた応用(アプリケーション)は生物発光を化学的 に理解しようとする研究から生み出されたものである。
今後もその応用の発展が期待される。そのためにも「発光機構に関する基礎研究の積 み重ね」と「新規な発光系・発光物質の探索」は必須の研究テーマであると私は考えて いる。
衛生検査キット
(キッコーマン株式会社)
薬物効果による腫瘍細胞退縮の様子
(Xenogen社)
レポーターアッセイキット
(東洋ビーネット株式会社)
(環境汚染モニタリング、生命検出系)
微生物が持つ
ATP
をモニタリングすることで、食品などに 混入する微生物の検出・定量を行う。また、同様の理由で宇 宙などでの地球型生命の検出にも用いられた。(プロモーターアッセイ:遺伝子発現検出系)
遺伝子の転写活性を評価、転写産物をモニタリングする。
(生体内イメージング)
ルシフェラーゼ、または
GFP
を生体内で発現させ、マークしたタンパク質などの物質の動態を観察する。
Fig.1-11 生物発光の応用例
1----3 1 1 1 3 3 3. . . . 発光菌類 ( 発光菌類 発光菌類 発光菌類 ( ( (luminous luminous luminous luminous fungi) fungi) fungi) fungi) とその とその とその とその研究 研究 研究の 研究 の歴史 の の 歴史 歴史 歴史
(15)(15)(15)(15)生物の発光は古代から人類に認識されて利用されていた。原住民が夜の森を歩くため の明かりとして発光キノコを利用していたという記録が存在する。発光という現象と菌 類が直接繋がったのは
19
世紀半ばで、J. F. Heller (1813-1920)
が朽木の発光と動物の 発光はそれぞれ菌とバクテリアが原因だと示した。発光菌類は持続的な発光を行い、極 大発光波長が520~530 nm
であるなどの特徴を持ち、様々な発光菌類の種が世界中に 分布している。時代が進み、W. Pfeffer (1845-1920) が
Dubois
のL-L
反応(4)(5)を発光菌類に対して 応用したことが菌類の発光についての近代的アプローチの始まりであった。しかし、発 光菌類、発光キノコ類においては、Duboisが考案した伝統的な熱水抽出物と冷水抽出物に よるL-L反応を示さない例が多かった。これは、ルシフェリンが低濃度でしか存在しない、または不安定であることがL-L反応を示さない原因と推測されていた。
1962年にAirth、Foersterらが始めて発光菌類でL-L反応を確認したとの報告がなされ
たが、色々問題点も多かった(16)(17)。具体的には、まずナラタケ(
Armillaria mellea
)から熱 水抽出物(ルシフェリン)を、エノキダケ(Collybia velutipes
)から破砕沈殿物(不溶性ルシ フェラーゼ)を得た。その熱水抽出物とNADHをインキュベートした後、ルシフェラーゼ としてエノキダケの破砕沈殿物を添加することで発光が確認された。ここから、Fig.1-12 に示すような 2 段階の反応による発光機構が提唱された。しかし、ここで行われた発光実 験ではルシフェリンとルシフェラーゼを異なる種から抽出している点、またエノキダケは 発光しない種であるという点が問題であった。また L-L 反応において、ルシフェリンは発光した後エミッターである発光体(オキシル シフェリン)に変化すると考えられるので、抽出物の蛍光スペクトルから発光菌類のオキ シルシフェリンを推定、単離、構造決定するという試みも様々な研究者によってなされて きた。一例を挙げると、日本などに分布しているツキヨタケ(
Pleurotus japonicus
syn.Lampteromyces japonicus
)からは発光エミッターとしてLampteroflavinがGoto(Isobe)ら によって単離されている(18)(19)。しかし、これらの物質は生物発光の発光エミッター候補と はなるものの、in vivo
の生物発光に関連しているかは確認されておらず発光キノコ類の生 物発光をin vitro
で再現できていない。また、Shimomuraらは発光菌類の研究を通じていくつかの特徴を見出した。一つは、数 種の発光菌類からルシフェリンとして抽出した物質が H2O2と Fe2+存在下において化学発 光を起こすこと。もう一つは発光菌類のどの種においても、生物発光を触媒する酵素(ル シフェラーゼ)の役割を担う酵素が発見できなかったことである。これらの実験結果から
Shimomura は発光菌類の発光には(O2-)スーパーオキシドアニオンが関連し、また発光
X
(luciferin precursor)
NAD(P)H/H+ XH2 + NAD(P)+
+ soluble enzyme
(reductase)
XH2 + O2 X’ + H2O + Light
particulate enzyme
(luciferase) (oxyluciferin)
Fig.1-12 Airth、Foersterが提唱した発光機構 (luciferin)
(luciferin)
反応がルシフェラーゼによって触媒されない反応と推測した。そして
P. stipticus
子実体よ りパナール(panal)と命名した物質を単離し、P. stipticus
はパナール誘導体を発光基質とし て、酵素ではなくカチオン性界面活性剤が反応を触媒するという発光反応(Fig.1-13)を提案 した(20-23)。このように発光菌類の研究は行われているものの、今日まで
in vitro
で発光菌類の生物発 光を再現したという報告はなく、発光物質は全くの未解明な状態にある。Fig.1-13 Shimomuraが提唱したpanalの発光機構(20-23)
OH
O HOOC
O H
OH
O HOOC
NR H
OH
HOOC N R
OH
OH
HOOC N R
OH
panal
RNH2
pH 3.5, 24 h, surfactant
Probable cyclic product
H2O2/Fe2+
pH 7-8, cationic surfactant
Light + products
1 1
1 1----4. 4. 4. 4. ヤコウタケ ヤコウタケ ヤコウタケ ヤコウタケ (((( Mycena Mycena Chlorophos Mycena Mycena Chlorophos Chlorophos Chlorophos ))))
ヤコウタケ(
Mycena chlorophos
)はクヌギタケ属の腐生菌の発光キノコである(
Fig.1-10
)。主に亜熱帯性気候の八丈島、小笠原諸島、東南アジアなどといった極限られた場所にのみ分布している。子実体の形状は傘径
7~27 mm、柄の高さ 10~23 mm
である。天然の状態では6~7
月、9
月といった雨季の晴れ間に、フェニックスやシンノ ウヤシ、ビロウヤシといった椰子の枯葉や幹に発生し、約3
日間連続的に発光する。ヤ コウタケの発光色は、淡い緑色(λmax= 525 nm
)である。ヤコウタケの発光強度は、発光キノコの中でも強く、毒キノコとしても有名なツキヨタケよりも強い。天然のヤコ ウタケ子実体はまばらにしか生育しておらず、実験試料として大量に確保することは非 現実的であるが、1995 年に三洋電機株式会社の新津らが、実験室内でのヤコウタケ子 実体の栽培に成功した(24)(25)。新津の協力を得て、当研究室においても実験室レベルで の栽培により実験試料としてのヤコウタケを自給することに成功している。
Fig.1-10
天然のヤコウタケ 左上:明所下 右上:暗闇中下:天然の生育状態(フェニックスに発生)
(写真提供:市野亨氏)
拡大
拡大
拡大
拡大
1 1
1 1----5. 5. 5. 5. 本研究 本研究の 本研究 本研究 の の の目的 目的 目的 目的
本研究の目的は、ヤコウタケ生物発光研究を通して、発光キノコ類、発光菌類の生物 発光機構の解明を行うことである。
発光菌類は歴史的に見て、生物発光について研究がほとんど進んでいない。多くの努 力にもかかわらず、ヤコウタケなどの発光菌類の発光系が
L-L
反応であるという確た る証拠すら示されておらず、その発光機構は未知である。本研究では、発光菌類のヤコウタケの生物発光を対象として研究を行った。ヤコウタ ケは培養が可能であるため新鮮なサンプルを必要に応じて大量に入手可能であるとい う点は生物発光研究において大きなアドバンテージとなる。
もしヤコウタケの発光系を解明することができれば新しい発光基質、発光系をバイオ イメージング技術などに提供することができる可能性がある。またその長時間持続発光 性を活かして新しいツールを開発できる、などの有用な応用が期待できる。このような 観点でヤコウタケ発光機構研究を始めた。
第 第 第
第 2 2 2 2 章 章 章 章 本論 本論 本論 本論
2 2
2 2----1. 1. 1. 1. ピートモス ピートモスを ピートモス ピートモス を培地 を を 培地 培地として 培地 として として用 として 用いた 用 用 いた いた いたヤコウタケ ヤコウタケ ヤコウタケ ヤコウタケの の改良栽培法 の の 改良栽培法 改良栽培法 改良栽培法の の の確 の 確 確 確 立
立 立 立
ヤコウタケの人工栽培は、すでに新津らによって確立されている(22)(23)。この方法で は培地である腐葉土は熱風乾燥させ直径
2 mm
に粉砕したものを使用する必要があり、培地の調製に時間と手間を要する。今回、腐葉土の代わりにピートモスを培地として用 いたヤコウタケの栽培法を確立した。Fig.2-1 に示すヤコウタケのライフサイクルの中で 培地をピートモスに変更することで、培地調製の労力を軽減した。
ピートモス(
peat moss)
は、水ゴケが腐植し堆積したものを乾燥・粉砕した農業・園 芸用の土である。主な産地はカナダ、北ヨーロッパ、サハリン、北海道などであり、通 気性、保水性、保肥性が高く、有機酸を含むため通常弱酸性(約pH 4
)を示す。ヤコウタケ菌糸の最適生育
pH
は4
であり酸性条件を好むので、酸性であるピートモ ス培地はヤコウタケ栽培に適しているといえる。特に前処理を必要とすることなく、栄 養源である米糠(もしくは小麦粉)と含水率調整のみを行えばよいので、比較的手軽に ヤコウタケ栽培を行うことができた。胞子
菌糸体
菌糸が蔓延した ピートモス培地 原基
子実体
約4週間 約
1
週間約
1
週間Fig.2-1
ヤコウタケのライフサイクル2 2
2----2. 2 2. 2. 2. ヤコウタケ ヤコウタケ子実体 ヤコウタケ ヤコウタケ 子実体 子実体 子実体の の発光 の の 発光 発光の 発光 の の特徴 の 特徴 特徴 特徴
栽培で得たヤコウタケ子実体を用いて、発光の経時変化と発光スペクトルの測定、お よび光学顕微鏡による観察を行った。
2 2 2 2----2 2 2 2----1. 1. 1. 1.
ヤコウタケのヤコウタケヤコウタケヤコウタケのの暗闇中の暗闇中暗闇中の暗闇中ののの発光発光発光発光栽培したヤコウタケ子実体は持続発光し、暗闇中において淡い緑色に発光した。また、
主に子実体の傘の部分が強く発光しており、柄の部分の発光は微弱であった。ヤコウタ ケの傘の部分が特に強く発光していることから、ヤコウタケの発光物質は傘の部分に多 く存在していると推測された。ヤコウタケの柄部分は例えるならタンポポの茎のように なっており、周りが繊維質の柄で中空になっている。ヤコウタケの柄の中空部分には水 分が充填されていた。ヤコウタケ菌柄から傘部まで水分を送るルートであるとも考えら れた。
Fig.2-2
ヤコウタケ写真 左:明所中 右:暗室中
2 2 2 2----2 2 2 2----2. 2. 2. 2.
ヤコウタケ子実体ヤコウタケヤコウタケヤコウタケ子実体子実体の子実体のの発光経時変化の発光経時変化発光経時変化発光経時変化新津のデータによれば、ヤコウタケ子実体の発光量は子実体形成後成長に伴い発光量 は増大し、約
24
時間後に極大に達した。その後徐々に発光は減衰し、約72
時間後(約3
日後)には発光を終了する(Fig.2-3)
(22)。000 0 0.1 0.10.1 0.1 0.2 0.20.2 0.2 0.30.30.3 0.3 0.4 0.40.4 0.4 0.5 0.50.5 0.5 0.60.60.6 0.6 0.70.70.7 0.7 0.8 0.80.8 0.8 0.9 0.90.9 0.9 111 1
0 00
0 20202020 4 04 04 04 0 60606060 子実体形成
子実体形成子実体形成
子実体形成からのからのからの経過時間からの経過時間経過時間経過時間 ((((時間時間時間時間))))
N o rm al iz e d I n te n si ty N o rm al iz e d I n te n si ty N o rm al iz e d I n te n si ty N o rm al iz e d I n te n si ty
Fig.2-3
ヤコウタケ子実体の発光経時変化((22)より抜粋・改変)ヤコウタケの外見状態も時間経過によって変化した。Fig.2-2 のように子実体は形成 した直後は綺麗な乳白色をしているが、時間が経過すると色が茶色がかってゆき子実体 がしおれていくのが確認できた。
また、約
72
時間もの長時間持続発光を行う発光生物は希少であり、その点からヤコウ タケは非常に興味深い研究対象である。発光強度とヤコウタケの鮮度に関連性がある(新鮮であるほど強く発光する)ことから、胞子形成のような生命活動が盛んに行われ ているほど発光量が増加する傾向があると考えられる。
2 2 2 2----2 2 2 2----3. 3. 3. 3.
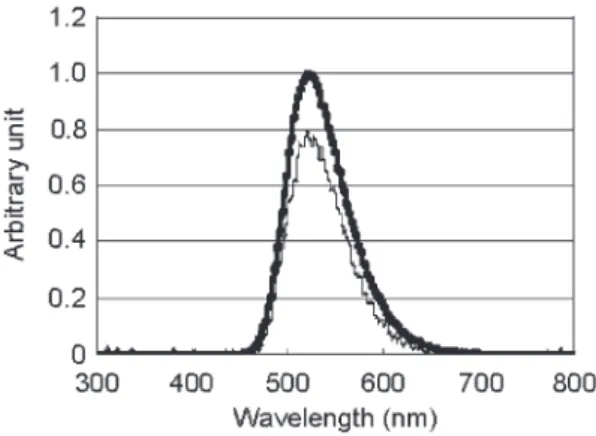
ヤコウタケ子実体ヤコウタケヤコウタケヤコウタケ子実体子実体の子実体のの発光の発光発光発光スペクトルスペクトルスペクトルスペクトルヤコウタケの発光色は淡い緑色であり、発光スペクトルは極大発光波長が
525 nm
で あった(Fig.2-4)。000 0 0.2 0.20.2 0.2 0.4 0.40.4 0.4 0.6 0.60.6 0.6 0.8 0.80.8 0.8 111 1
400 400 400
400 450450450450 500500500500 550550550550 600600600600 650650650650 700700700700 wavelength (nm)
wavelength (nm) wavelength (nm) wavelength (nm)
N o rm al iz e d I n te n si ty N o rm al iz e d I n te n si ty N o rm al iz e d I n te n si ty N o rm al iz e d I n te n si ty
2----2 2 2 2 2 2 2----4. 4. 4. 4.
マイクロスコープマイクロスコープマイクロスコープマイクロスコープををを用を用いた用用いたいたいたヤコウタケヤコウタケヤコウタケヤコウタケ細胞細胞細胞の細胞のの観察の観察観察 観察株式会社キーエンスに協力していただきヤコウタケ細胞をマイクロスコープ(株式会 社キーエンス製マイクロスコープ
VHX-1000
)で撮影した(Fig.2-5)
。ヤコウタケ傘の部 分のひだ状の組織を切り取り撮影したところ、白い粒のようなものが多く観察できた。この粒は光学顕微鏡でヤコウタケ破砕物を観察した際にも多く観察できたものであっ
Fig.2-4
ヤコウタケ子実体の発光スペクトル日本分光株式会社製 FD-6500 にて測定
マイクロスコープなどで観察するには弱い発光であるので発光部の観察はできなかっ た。
Fig.2-5
ヤコウタケ子実体傘部切片のマイクロスコープ写真2 2
2 2----3. 3. 3. 3. ヤコウタケ ヤコウタケの ヤコウタケ ヤコウタケ の状態 の の 状態 状態 状態による による による による発光量変化 発光量変化 発光量変化 発光量変化
ヤコウタケが長時間発光を行うといっても長くて三日程度であり、継続的にヤコウタ ケを使用した実験を行うためには、定期的なヤコウタケ栽培が必要であった。しかしヤ コウタケ栽培には植菌から子実体形成まで最短でも二ヶ月程度はかかり、培地調整など にかかる労力は少なくなかった。そこでヤコウタケを保存性が高く、実験にも用いやす いものにするために、冷凍、凍結乾燥などによるサンプル保存を試みた。
また、ヤコウタケ子実体の発光は組織を破砕することで発光強度が減少する傾向があ った。破砕によってヤコウタケの発光が失活してしまうと、発光を手がかりとした発光 活性の測定や発光物質の抽出、保存などができなくなる。そこで、ヤコウタケ発光は破 砕によってどのように発光強度が変化するか測定した。
2 2
2 2----3 3 3 3----1. 1. 1. 1.
凍結乾燥凍結乾燥凍結乾燥凍結乾燥ヤコウタケヤコウタケのヤコウタケヤコウタケのの調製の調製調製調製とととと発光反応発光反応発光反応発光反応まず凍結ヤコウタケは新鮮なヤコウタケを液体窒素で瞬間的に凍結処理することで 得た。また、凍結乾燥ヤコウタケは液体窒素で冷凍したヤコウタケ子実体を凍結乾燥機 で一晩乾燥することで得ることができた。各サンプルの特徴としては、凍結サンプルは 凍結状態ならば発光していないが融解が始まると発光を開始した。また凍結乾燥サンプ
ルは
Fig.2-6
で示すように白色で、子実体傘の部分はかなりもろくすぐ崩壊するが、柄の部分は強靭な繊維質のおかげで硬い。凍結乾燥ヤコウタケは乾燥状態では発光はして いないが、これに純水、緩衝液などの水分を加えると発光を開始することを発見した。
また、低温乾燥条件で
1
~2
ヶ月発光活性を維持したまま保存できることがわかった。ルを比較した(Fig.2-7)。その結果、凍結乾燥ヤコウタケの発光スペクトルは凍結乾燥前 の生のヤコウタケ子実体とほぼ一致していた。このことから、両者は同一の発色団、発 光機構で発光しているものと予想でき、凍結乾燥サンプルの研究を行うことでもヤコウ タケの研究を進めることが可能であると考えられた。
Fig.2-6
凍結乾燥ヤコウタケ1.0
0.8
0.6
0.4
0.2
0.0
N o rm a li z e d I n te n s it y
700 600
500 400
300
Wavelength(nm)
生の子実体切片
凍結乾燥子実体
生ヤコウタケ、凍結ヤコウタケ、凍結乾燥ヤコウタケ、それぞれの発光の経時変化は
Fig.2-8
に示した。生ヤコウタケサンプルは持続的に強く発光しているが、冷凍サンプルと凍結乾燥サンプルは異なる経時変化を示した。冷凍サンプルは解凍と同時に発光開 始し、その後ゆっくりと発光量が減少し続けた。また、凍結乾燥サンプルは緩衝液を添 加すると同時に生サンプルに近い発光強度で発光するが、その後は急激に発光量が減少 した。
ヤコウタケ子実体の発光は凍結処理を行うと持続的に発光しなくなり、徐々にもしく は急激に発光量が減少した。おそらく生体内での発光関連物質の生合成経路が凍結処理 によって停止してしまい、残存している発光物質だけで発光するため発光量が除々に低 下してしまうのではないかと考察できる。つまり冷凍処理、凍結乾燥処理はヤコウタケ 発光に大きな影響を与えていることがわかる。このことから、発光菌類の最大の特徴で ある持続発光の原因について調査するためには、生のヤコウタケサンプルを用いて研究 を行う必要があると考えた。また、今回調製したヤコウタケ凍結乾燥子実体は常温で保 存可能であるなど保存性に優れ、水または緩衝液を添加した時点から発光を開始する点 から、発光のコントロールが行いやすいサンプルであると言える。
Light intensity (counts/sec) Fresh sample Frozen sample
Freeze-dried sample
0 1.0x108 1.0x107 1.0x106 1.0x105
1.0x103 1.0x104
1.0x102 10
Light intensity (counts/sec) Fresh sample Frozen sample
Freeze-dried sample
0 1.0x108 1.0x107 1.0x106 1.0x105
1.0x103 1.0x104
1.0x102 10
2 2 2
2----3 3 3----2. 3 2. 2. 2.
ヤコウタケヤコウタケ組織ヤコウタケヤコウタケ組織組織組織をを破砕をを破砕破砕破砕したしたしたした状態状態での状態状態でのでの発光での発光発光 発光ヤコウタケ子実体の発光は破砕することで急激に減少する。この現象はヤコウタケを 乳鉢などで破砕する際に目視で確認していた。この破砕による発光量減少を詳しく調べ るために、ヤコウタケ子実体切片と緩衝液などを用いて、破砕の効果を発光測定で確認 した(Fig.2-9)。
その結果、ヤコウタケ組織破砕時の発光量減衰速度は未破砕サンプルと比較してとて も早い。また、破砕しただけで初期発光量も約十分の一になってしまっているのが確認 できる。
破砕によってヤコウタケの発光量が減少してしまう理由としては、①細胞組織として 発光を行っているため破砕処理により組織が壊れて発光物質生産が止まる。また②破砕 による組織表面積増加で内部の酸化速度が速くなった結果発光物質が減少した、という 可能性が考えられる。
破砕方法(乳鉢、ヒスコトロン)による発光減少効果の違いは確認できなかった。な お、ヤコウタケは繊維が強いためか完全な懸濁液にはならなかった。
破砕による急激な発光量減少は、低温下(約
4℃)では緩和することが判明した。そ
の場合、破砕により発光量は十分の一程度になり発光量の減少速度も未破砕のサンプル よりも早かった。1010 1010 100 100 100 100 1x10 1x10 1x10 1x10 3333 1x10 1x10 1x10 1x10 4444 1x10 1x10 1x10 1x10 5555 1x101x10 1x101x10 6666 1x10 1x10 1x10 1x10 7777
0 00
0 5555 10101010 15151515
切片 切片
切片 切片と と と破砕物 と 破砕物 破砕物 破砕物の の の の比較 比較 比較 比較
切片切片切片
切片+++超純水 +超純水 超純水 超純水 AveAveAveAve....
切片 切片切片
切片+++NaPB Ave+NaPB AveNaPB AveNaPB Ave....
切片 切片切片
切片+++Tris Ave+Tris AveTris AveTris Ave....
破砕切片破砕切片破砕切片
破砕切片++++超純水 超純水 超純水 Ave超純水 AveAveAve....
破砕切片 破砕切片破砕切片
破砕切片++++NaPB AveNaPB AveNaPB Ave....NaPB Ave 破砕切片破砕切片破砕切片
破砕切片++++Tris AveTris AveTris Ave....Tris Ave
L u m in e sc e n c e i n te n si ty L u m in e sc e n c e i n te n si ty L u m in e sc e n c e i n te n si ty L u m in e sc e n c e i n te n si ty
time (h) time (h) time (h) time (h)
Fig.2-9
ヤコウタケの破砕による発光量変化(n=3)
超純水、リン酸緩衝液
(NaPB)
、Tris-HCl
緩衝液中での切片、破砕切片それぞれの 発光量経時変化L ig h t in te n s it y ( c o u n ts /s e c )
2 2
2 2----4. 4. 4. 4. ヤコウタケ ヤコウタケの ヤコウタケ ヤコウタケ の の の発光 発光と 発光 発光 と と と酸素 酸素 酸素の 酸素 の の関係 の 関係 関係 関係
生物発光に酸素が必要であることを証明する実験が行われたのは古く、1667 年であ
る。
Robert Boyle
は1667
年に真空ポンプを用いて発光菌類が付着した木片の発光が真空状態で消失することから、発光には空気が必要であることを見出している。またその 後のホタル発光などの化学的な研究から、空気に含まれる発光に必要な物質は酸素であ ることが判明している。
一般的に生物発光は発光する際に酸素が必要である。発光タンパクにおいても基質と 酸素が結びついた状態で存在する。発光菌類においては発光に酸素が必要であるとわか っているが、ヤコウタケの発光、特に子実体の発光に酸素が関連しているかは解明して いなかった。そこでヤコウタケの発光における酸素の関与を調べることにした。具体的 な実験方法としては伝統的な方法を踏まえて、真空ポンプを用いてヤコウタケを酸素が 存在しない状況(真空、もしくはアルゴン雰囲気下)に置いた場合の発光の変化を観察 した
(Fig.2-10)
。空気 空気 空気 空気
アルゴン アルゴン アルゴン アルゴン
アルゴン アルゴン アルゴン アルゴン
酸素 酸素 酸素 酸素
(
(
(
(空気環境下 空気環境下 空気環境下 空気環境下) ) ) ) ( (アルゴン ( ( アルゴン アルゴン雰囲気下 アルゴン 雰囲気下) 雰囲気下 雰囲気下 ) ) ) ( (酸素 ( ( 酸素 酸素 酸素または または または または 空気雰囲気下 空気雰囲気下 空気雰囲気下 空気雰囲気下) ) ) )
発光 発光 発光
発光 発光停止 発光停止 発光停止 発光停止 再発光 再発光 再発光 再発光
Fig.2-10
雰囲気の変化とヤコウタケの発光(
((
(ヤコウタケヤコウタケヤコウタケヤコウタケ子実体子実体子実体子実体のののの発光状態発光状態発光状態)発光状態)) )
(上の発光の状態をデジタルカメラ(8秒露光)で撮影した写真)