男児外性器皮膚線維芽細胞におけるアンドロゲン標
的候補遺伝子の探索
著者
中尾 佳奈子
学位授与機関
Tohoku University
学位授与番号
11301甲第19155号
URL
http://hdl.handle.net/10097/00129300
博士論文
男児外性器皮膚線維芽細胞におけるアンドロゲン標的候補遺伝子の探索
東北大学大学院医学系研究科医科学専攻 次世代小児医療講座 中尾 佳奈子
1
本博士論文は、2019 年にEndocrine Journal 誌に掲載された、
Dihydrotestosterone induces minor transcriptional alterations in genital skin fibroblasts of children with and without androgen insensitivity. (doi:
10.1507/endocrj.EJ18-0494.) を基礎論文として作成した。
基礎論文は、下記Endocrine Journal 誌のサイトに掲載されている:
2 目次 第Ⅰ章 要約 第Ⅱ章 研究背景 第Ⅲ章 研究目的 第Ⅳ章 研究方法 第Ⅴ章 研究結果 第Ⅵ章 考察 第Ⅶ章 結論 第Ⅷ章 謝辞 第Ⅸ章 文献 第Ⅹ章 図 第 XI 章 表
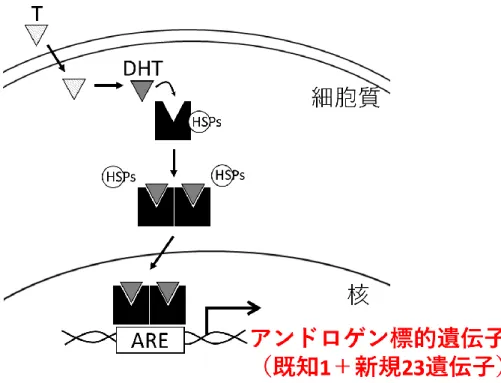
3 第Ⅰ章 要約 1 テストステロンやジヒドロテストステロン(dihydrotestosterone:DHT) 2 などの男性ホルモン(アンドロゲン)はアンドロゲン受容体(androgen 3 receptor:AR)への結合を介して外性器男性化を促す。AR は核内受容体であ 4 り、リガンドとの結合によって細胞質から核内に移行する。そして標的遺伝子 5
のアンドロゲン応答配列(androgen response element:ARE)に結合して転写
6 制御を行う。これまでにヒト外性器におけるアンドロゲン標的遺伝子として 7 APOD が同定されている。しかし、この遺伝子単独の作用だけでは複雑な外性 8 器の男性化を完全には説明できず、詳細な外性器形成におけるアンドロゲン作 9 用の分子メカニズムは未解明である。本研究は、ヒト外性器におけるアンドロ 10 ゲン標的候補遺伝子を同定することを目的とした。 11 AR 変異 p.Ala597Thr を有する部分型アンドロゲン不応症患児 1 名およ 12 び埋没陰茎男児 4 名(コントロール群)を対象とし、この 5 名の包皮由来皮膚 13 線維芽細胞を用いてトランスクリプトーム解析を行った。まず各細胞を 2 つに 14 分け、DHT またはメタノールを添加して培養した。その後、細胞から mRNA 15 を抽出し、マイクロアレイ解析を行った。コントロール群において DHT 添加 16 によって変動し、かつ、患者における変動がコントロール群よりも小さい遺伝 17 子を抽出した。発現量が著しく低い遺伝子およびシグナルデータの品質が悪い 18
4 遺伝子は解析対象から除外した。その結果、アンドロゲン標的候補遺伝子とし 19 て 24 遺伝子が同定された。7 遺伝子が近傍に ARE 候補配列を有していた。24 20 伝子の中には既知アンドロゲン標的遺伝子APOD、マウス外性器原基男性化に 21 関与すると報告されているCYP1B1、他臓器におけるアンドロゲン標的遺伝子 22 として報告のあるRGCC や HS6ST1 が含まれていた。コントロール群におけ 23 る 24 遺伝子の DHT 添加による log2発現変動量は 0.3-0.7 であった。本研究の 24 結果は、アンドロゲンによる外性器男性化の主体が、少数の特定遺伝子の大き 25 な発現変動ではなく多数の遺伝子の微細変化であることを示唆する。 26 なお、本研究で同定された 24 個のアンドロゲン標的候補遺伝子には、 27 これまでヒト疾患への関連が報告されていないものが多く含まれる。これらの 28 遺伝子の変異は、臨床的にアンドロゲン不応症と診断されるがAR 変異が認め 29 られない患者における疾患原因となっている可能性がある。また本研究によっ 30 て得られた知見は、AR 異常のためにアンドロゲン補充療法では十分な男性化 31 が認められない患者のために、AR を介さずにアンドロゲン作用を発揮する新 32 規治療薬の開発につながることが期待される。 33 34 35 36
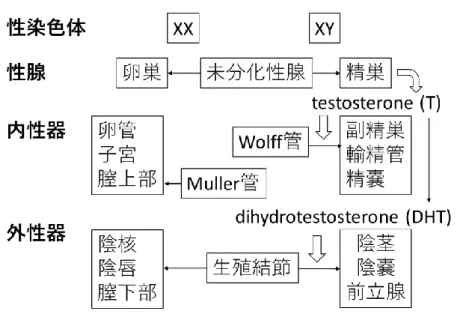
5 第Ⅱ章 研究背景 37 ヒトの性分化は遺伝学的性、すなわち Y 染色体(SRY 遺伝子)の有無によ 38 り性腺の性が決定されることから始まる(図 1)。SRY 遺伝子が存在すると 39 き、未分化性腺は精巣に分化する。そして胎盤由来 hCG の刺激により、精巣 40 のライディッヒ細胞は大量のテストステロンを産生する。そのテストステロン 41 がウォルフ管に作用すると男性型内性器が形成される。また精巣のセルトリ細 42 胞から分泌される抗ミュラー管ホルモン(AMH)の作用によりミュラー管は 43 退縮する。末梢組織でテストステロンは 5α-還元酵素により、活性の高いアン 44 ドロゲンであるジヒドロテストステロン(dihydrotestosterone:DHT)に変換 45 される。DHT はテストステロンよりも約 2 倍高い AR 結合能を持ち、解離速 46 度は逆にテストステロンの約 1/5 であることから、テストステロンよりも強い 47 アンドロゲン活性を持つ[1]。この DHT が外性器の原基である生殖結節に作用 48 すると、外性器は男性化に進み、陰茎や陰嚢、前立腺が形成される。テストス 49 テロンや DHT などのアンドロゲンは、アンドロゲン受容体(androgen 50 receptor:AR)への結合を介して男性化作用を生じる[2]。 51 AR 遺伝子は Xq11-12 上に位置し、8 エクソンから成る。AR タンパク質は 52 リガンド結合ドメイン、DNA 結合ドメイン、N 末端ドメインを有する(図 53
2)。リガンド非結合 AR は細胞質に存在し、heat shock protein 複合体
6 (HSP90、HSP70、HSP40 など)により不活化されている[3]。リガンドであ 55 るアンドロゲンがリガンド結合ドメインに結合すると、AR は活性型となって 56 ホモ二量体を形成して核へ移行[4]し、アンドロゲン標的遺伝子の近傍に存在す 57
るアンドロゲン応答配列(androgen response element:ARE)に結合して転写
58 制御を行う[5]。この転写制御には種々の転写共役因子が関わる[2]。しかし、 59 この外性器男性化に関与するアンドロゲン標的遺伝子の報告は少なく、詳細は 60 未解明である。 61 外性器における既知のアンドロゲン標的遺伝子として apolipoprotein D 62
(APOD)がある。この APOD 遺伝子は、Appari らによって、外性器における
63 アンドロゲン標的遺伝子を探索することを目的として行われた外性器皮膚の線 64 維芽細胞(fibroblast:FB)を用いたトランスクリプトーム解析で同定され 65 た。当初の解析では、正常男性の包皮由来皮膚 FB と CAIS 患者の大陰唇由来 66 皮膚 FB との間に DHT 添加による遺伝子発現量の有意な違いは認められなか 67 った[6]。しかし発生学的な由来を統一し、大陰唇に対応する陰嚢由来皮膚 FB 68 を用いて検討し直した結果、DHT 添加により 3 つの遺伝子の発現が正常男性 69 において AIS 患者よりも統計学的に有意に上昇していることが明らかになった 70 [7]。その 3 遺伝子のうちタンパク質をコードしていたのはAPOD 遺伝子だけ 71 であり、他2遺伝子はそれぞれ偽遺伝子と機能未知の遺伝子であった。APOD 72
7 は 169 アミノ酸の糖タンパク質であり、構造からリポカリンファミリーに属す 73 ると推定されている[8]。APOD はアンドロゲンやプロゲステロンなど複数の 74 疎水性リガンド結合能を有する。APOD はヒトにおいてユビキタスに発現して 75 おり、とくに精巣、脾臓、脳に高発現である。また APOD はヒト前立腺がん 76 細胞株において、アンドロゲン刺激により発現が増え、APOD 発現増加に伴っ 77 て細胞増殖が低下したと報告されている[9]。他にも乳癌細胞において、細胞増 78 殖抑制に関与することが報告されている[10]。したがって、男性の外性器皮膚 79 においても APOD はアンドロゲン依存的に発現が増え、細胞増殖の調整に関 80 与していることが推測される。しかし APOD 単独では複雑な分化・増殖の過 81 程すべてを説明することはできず、APOD よりも発現変動が軽微な未知のアン 82 ドロゲン標的分子が協働して外性器男性化に寄与している可能性が考えられ 83 る。またマウスの外性器原基やヒトの外性器以外の組織においては、アンドロ 84 ゲンの標的候補遺伝子を同定したとの報告がある。たとえば、マウスの外性器 85 原基ではCYP1B1[11]、GPER1[12]、前立腺では ESM1[13]、神経幹細胞では 86 RGCC[14]などが報告されているが、これらの遺伝子とヒト外性器の男性化と 87 の関連は明らかになっていない。 88 臨床的な観点では、AR 機能異常によって生じるアンドロゲン不応症 89
(androgen insensitivity syndrome:AIS)の遺伝学的診断および治療に課題が
8 残る。内・外性器や性腺の発育が非典型的である疾患を性分化疾患(disorder 91 of sex development : DSD)と称し、DSD の発生頻度はおおよそ出生 2000~ 92 4500 人に 1 人と推定されている[15]。DSD の多くは染色体核型が 46,XX であ 93
り、染色体核型 46,XY である DSD は比較的まれである。AIS は 46,XY DSD
94 の原因疾患の中で最も頻度の高い疾患であり、およそ出生 10 万人に 4 人程度 95 の発生頻度である[16]。AIS が疑われるのは、①染色体核型が 46,XY、②精巣 96 が存在、③ミュラー管由来構造物は欠如、④テストステロン値は正常または高 97 値、⑤テストステロンから DHT への変換能は保たれている、という状況下で 98
不完全な男性化を呈する場合である。AIS は完全型(complete AIS:CAIS)、
99
不完全型(partial AIS:PAIS)、軽症型(mild AIS:MAIS)に分類される
100
[17]。MAIS は外性器異常を欠き男性不妊で発見される。DSD を呈するのは
101
CAIS と PAIS のみである[17]。CAIS 患児の外性器は完全女性型であるのに対
102 し、PAIS 患児では外性器の表現型に幅があり、女性型に近い陰核肥大や陰唇 103 癒合から男性型に近い二分陰嚢や尿道下裂まで様々な程度の外性器異常を呈す 104 る[17]。 105 確定診断であるAR 変異陽性率は、CAIS 患者では 95%以上であるのに対 106
し、PAIS 患者では約半数にとどまる[18]。AR 変異陰性例で AIS を発症する機
107
序として、転写共役因子や AR の標的遺伝子および AR シグナル経路に関わる
9 分子の質的・量的異常が関与している可能性が想定されている[19]が、特定の 109 疾患原因分子は同定されていない。 110 治療面では、CAIS 患児は社会的女児として養育することが一般的であ 111 り、適切な時期(一般的にはテストステロンが末梢でエストロゲンに変換され 112 て生じる思春期以降)での精巣摘出術と、必要に応じて女性ホルモン補充療法 113 や膣形成術を行う。PAIS 患児では男性化不全の程度などを考慮して社会的性 114 を決定する。社会的女児として養育される場合は CAIS の治療に準じ、必要に 115 応じて外性器形成術を行う。社会的男児として養育される PAIS 患者では、外 116 性器形成術とアンドロゲン補充療法を行う。しかし PAIS 患者に対するアンド 117 ロゲン補充療法では、男性化作用を生じるまでにしばしばアンドロゲン大量投 118 与を要する[20]。特にリガンド結合ドメインに変異を持つ患者はアンドロゲン 119
補充に抵抗性であるとされる。AIS の約半数が PAIS であり、また AIS で同定
120 されるAR 変異の約半数以上はリガンド結合ドメインに存在[17]することから 121 概算すると、AIS 患者の約 4 人に1人はアンドロゲン補充に抵抗性を示す。こ 122 れらの患者に対しては、新規治療法の開発が望まれる。 123 そこで本研究ではヒト外性器の男性化にかかわる未知のアンドロゲン標的 124 遺伝子の探索のため、アンドロゲン感受性のある組織に対してアンドロゲンを 125 添加し変動する遺伝子を抽出する研究を計画した。用いる組織は外性器原基で 126
10 ある生殖結節が最も理想的だが入手は極めて困難であるため、本研究では比較 127 的容易に入手できる組織として男児の外性器皮膚を用いることとした。特に研 128 究のために患者に余分な侵襲が加わることがないよう、原疾患の治療目的で行 129 われる外性器形成術の際に余剰で出る組織を利用した。外性器形成術は通常、 130 前思春期に行われるため、用いる組織は前思春期男児皮膚となるが、生理的な 131 アンドロゲンが低い前思春期の矮小陰茎患児においてアンドロゲンの投与によ 132 り陰茎長の伸長が観察されることから[21]、前思春期においてもアンドロゲン 133 ‐AR シグナル経路は働いており研究対象として問題ないと考えた。そして正 134 常男児との比較には AIS 患者の外性器皮膚を用いることとした。アンドロゲン 135 作用には、上記に記した古典的な経路のほか、非古典的経路が存在する可能性 136 があり[22]、中には AR を介さない経路の存在も示唆されている[23]。AIS 患 137 者と比較して正常男児において相対的にアンドロゲンによって大きく変動した 138 遺伝子群を抽出することで、この AR を介さない非古典的経路の影響を最小限 139 に抑えることが可能になると考えた。なお対象とする AIS 患者は PAIS 患者と 140 した。理想的には対象とする AIS 患者は AR 機能が完全に失われている CAIS 141 患者が正常コントロールとの差が大きくなるため望ましい。以前は CAIS であ 142 っても診断目的で外性器皮膚を採取しアンドロゲン結合能を測定することが広 143 く行われていたため、CAIS 患者由来皮膚を入手することが比較的容易であっ 144
11 た。しかし昨今では極力不必要な侵襲は避け、臨床的特徴と遺伝学的検査によ 145 り診断されるようになり、手術を受ける AIS 患者は PAIS 患者に限定されてい 146 るためである。 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162
12 第Ⅲ章 研究目的 163 本研究は、ヒト外性器におけるアンドロゲン標的候補遺伝子を同定するこ 164 とを目的とした。特に、遺伝子発現変動が比較的軽微な遺伝子にも注目して探 165 索を行った。 166 167 168 169 170 171 172 173 174 175 176 177 178 179 180
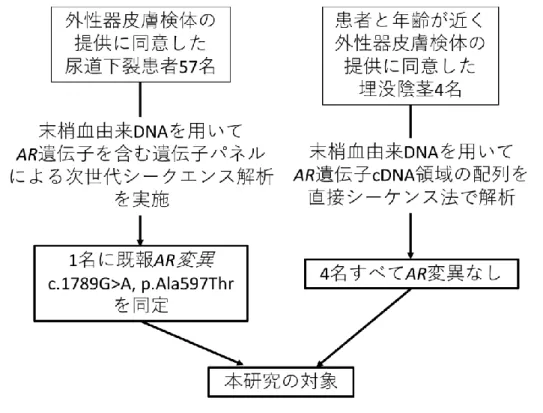
13 第Ⅳ章 研究方法 181 1. 倫理上の配慮 182 本研究は、国立成育医療研究センターの倫理審査委員会で承認を受けた 183 『性分化疾患・性成熟疾患・生殖機能障害における遺伝的原因の探索(倫理委 184 員会承認番号 512)』の一環として行われた。実施にあたっては共同研究者が説 185 明文書を用いて、可能な限り研究対象者本人、研究対象者が未成年の場合は保 186 護者にも説明を行い、文書同意を得た。研究への参加に同意した場合にも随時 187 同意を撤回できる旨も説明した。血液は通常の術前検査時にあわせて、年齢や 188 体格に応じて 3-10 ml 余分に採取した。皮膚組織は治療目的の手術において余 189 剰に出た組織を用いることで、患者への負担が最小限になるよう配慮した。検 190 体と臨床情報は連結可能匿名化され、解析担当者には匿名化した試料と臨床情 191 報のみが供与された。 192 193 2. 研究対象の選択 194 研究対象の選択フローを図 3 に示す。外性器皮膚検体の提供に同意した尿 195 道下裂患者 57 名のうち、次の項に示す末梢血由来 DNA を用いた次世代シーク 196 エンス解析によりAR 遺伝子に変異を同定した患者を“AIS 患者”として本研究 197 の対象とした。また、選ばれた AIS 患者と年齢が近く、皮膚が採取された外性 198
14 器の部位(包皮、陰嚢など)が同一で、外性器皮膚検体の提供に同意した埋没 199 陰茎(正常な大きさの陰茎が、包皮長が短いために矮小陰茎のように見えてい 200 る状態[24])4 名のうち、直接シークエンス法でAR 遺伝子に変異を認めなか 201 った者を“正常コントロール”として本研究の対象とした。 202 203 3. 次世代シークエンス解析 204
カスタム Haloplex Target Enrichment System (アジレント・テクノロジー
205 社、パロアルト、カルフォルニア州、米国)を用いて、末梢血由来 DNA からラ 206 イブラリ作成を行った。ライブラリ作成手順は、製造者の提供するプロトコー 207 ルに従って標準的な流れで行った。カスタムデザイン遺伝子パネルには、AR 208 遺伝子の他、尿道下裂に関与する既知/候補遺伝子 24 個(ATF3、BMP4、 209 BMP7、BNC2、CTGF、CYP1A1、CYR61、DGKK、EGF、ESR1、ESR2、 210 EGF8、FGFR2、GSTM1、GSTT1、HOXA4、HOXB6、HSD3B2、 211 HSD17B3、MAMLD1、MID1、NR5A1、SRD5A2、WT1)を掲載した 212 (Design ID 02185-1348467147)。作成したライブラリは、次世代シークエン 213 サーMiSeq(イルミナ社、サンディエゴ、カリフォルニア州、米国)により 214 150 塩基のペアエンド解析を行った。得られた配列情報は Surecall ソフトウェ 215 ア(アジレント・テクノロジー社)によりアライメントを行い、SAMtools 216
15 0.1.17 ソフトウェア(http://samtools.sourceforge.net)を用いて、バリエーシ 217 ョンコールを行った。 218 219 4. 直接シークエンス法 220 正常コントロールのAR 変異解析、および AIS 患者で次世代シークエンス 221 解析により同定された塩基置換の確認を,直接シークエンス法で行った。末梢 222 血由来ゲノム DNA からAR 遺伝子の翻訳領域を PCR 増幅するのに用いたプラ 223
イマーの一覧を表 1 に示す。PCR 増幅には、AmpliTaq Gold 360 Master Mix
224 (サーモフィッシャー・サイエンティフィック社、ウォルサム、マサチューセ 225 ッツ州、米国)を用いて、製造者のプロトコールに従った標準的な流れで行っ 226 た。得られた PCR 産物は、ExoSAP-IT(アフィメトリクス社、サンタクラ 227 ラ、カリフォルニア州、アメリカ合衆国)により酵素的に未反応 dNTP とプラ 228 イマー処理を行った。精製 PCR 産物は、BigDye Terminator v3.1(サーモフィ 229 ッシャーサイエンティフィック社、ウォルサム、マサチューセッツ州、米国) 230 を用いてシークエンシング反応を行ったのち、エタノール沈殿により精製を行 231 った。得られたシークエンス産物は、キャピラリーシークエンサーApplied 232
Biosystems 3130xl Genetic Analyzer(アプライド・バイオシステムズ社、フォ
233
スターシティ、カリフォルニア州、米国)によって塩基配列を決定した。
16 235 5. 細胞培養 236 本研究で用いた皮膚 FB は、尿道下裂または埋没陰茎に対して全身麻酔下 237 で共同研究者により行われた修復術の際に余剰で出た包皮由来皮膚組織から樹 238 立した。採取された組織片は、1 名につきおよそ 5 ㎜大が 1~2 切片である。組 239 織は 1 g/L グルコースと 10% ウシ胎児血清 (シグマ・アルドリッチ社、セン 240 トルイス、ミズーリ州、米国)と 10%ペニシリン-ストレプトマイシン溶液が含 241 まれる DMEM 培地(ナカライテスク社、京都、日本)に入れ、当施設に届け 242 られた。初代培養を行う際には、6 穴プレート上にメスで組織に幾筋も切り込 243 みを入れて固定し、37℃、CO2 5%に保たれたインキュベータ内で培養した。 244 培養には 1 g/L グルコースと 10% ウシ胎児血清 (シグマ・アルドリッチ社)と 245 1%ペニシリン-ストレプトマイシン溶液が含まれる DMEM 培地(ナカライテ 246 スク社)を用いた。生えてきた FB を 10 cm プレートに移して 80%コンフルエ 247
ントまで拡げたのち、トリプシン処理により細胞を剥がし、Cell Reservoir One
248 (ナカライテスク社)1 mL に入れて液体窒素のなかで凍結保存した。6 穴プレ 249 ートからの培養を繰り返すことで、1 名につき初代培養細胞(1 回のみ継代) 250 を 5 本ずつ凍結保存した。実験を始める際には凍結細胞を 37℃水浴で温めて迅 251 速(1 分以内)に融解し、10 ㎝シャーレ 2 枚に播種した。その後 10 ㎝シャー 252
17 レ 1 枚は 80%コンフルエントに達した段階で再び凍結保存し、もう 1 枚はおよ 253 そ 1 週間に 1 回程度の継代を行いながら実験に用いた。本実験終了までの継代 254 数は 10 回未満になるようにし、10 回を超える場合はこれを破棄して新たに凍 255 結細胞を融解して用いた。また継代数が 10 回未満であっても、細胞増殖速度 256 の低下や形態変化が生じてきた場合には新たに凍結細胞を融解して用い、対象 257 細胞間の増殖速度や状態等にばらつきが生じないよう配慮した。 258 259
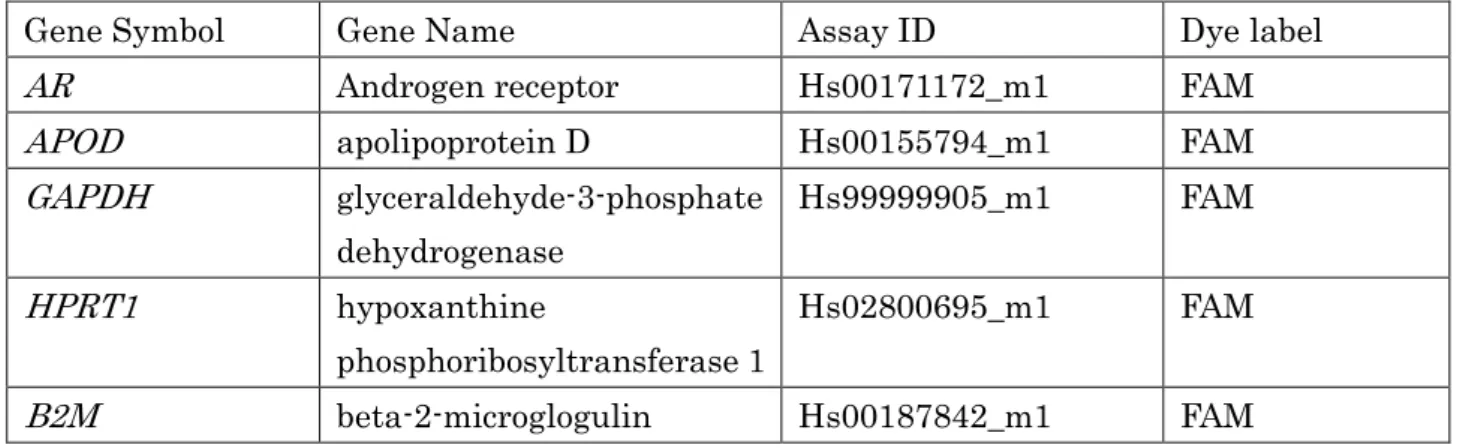
6. 定量 Reverse transcription‐polymerase chain reaction (qRT-PCR)法を用
260 いた条件検討 261 本実験に先立ち、DHT 添加濃度と添加後の培養時間を決定するため、 262 qRT-PCR 法により、AR 遺伝子発現上昇を指標とした条件検討を行った。条件 263 検討には、コントロール 1 名の細胞を用いた。 264 まずはΔΔCt 法に用いる内部標準遺伝子の決定のため、AR 遺伝子と、3 265 つの内部標準遺伝子GAPDH、HPRT1、B2M の増幅効率を検量線の傾きおよ 266 び直線性の比較によって検討した。6 穴プレートに FB を播種し(2× 105 cells/ 267 ウェル)、100%コンフルエントに達した段階で、トリプシン処理により細胞を 268
回収した。その後 RNeasy Plus Mini キット(キアゲン社, ヒルデン, ドイツ)を
269
用いて、製造者の標準プロトコールに従い total RNA 抽出を行った。電気泳動
18
で 2 本のリボソーム RNA バンドが見えることを確認したのち、High-Capacity
271
cDNA Reverse Transcription キット(サーモフィッシャーサイエンティフィッ
272
ク社)を用いて、製造者の標準プロトコールに従い cDNA 合成を行った。そし
273
て得られた cDNA を 7 段階希釈したサンプルを鋳型とし、各 TaqMan プロー
274
ブ(表 2)と TaqMan Gene Expression Assays(サーモフィッシャーサイエン
275 ティフィック社)を用いて Applied Biosystems 7500 リアルタイム PCR システ 276 ム(サーモフィッシャーサイエンティフィック社)で増幅反応させた。増幅反 277 応は triplicate で測定した。 278 次に、AR 遺伝子と増幅効率が同程度と判定された遺伝子を内部標準とし 279 て、ΔΔCt 法により DHT 添加時間の検討を行った。DHT(シグマ・アルド 280 リッチ社)は製造者の用法記載に基づきメタノールに溶解して用いた。まず 12 281 穴プレートに FB を播種し(1× 105 cells/ウェル)、80%コンフルエント近くに 282 達した段階で DHT を終濃度がそれぞれ 0 nM (メタノールのみ。終濃度 283 0.3%)、1 nM, 10 nM, 100 nM となるように調整して添加し、24 時間培養し 284 た。その後トリプシン処理により細胞を回収した。前述の通りの方法で qRT-285 PCR を行った。用いた内部標準遺伝子の TaqMan プローブを表 2 に示す。増 286 幅反応は triplicate で測定した。得られたデータをもとに、ΔΔCt 法によって 287 DHT 添加なし(メタノールのみ)の場合と比較して最もAR 遺伝子の発現を 288
19 増加させる DHT 濃度を決定した。増幅反応は triplicate で測定した。 289 最後に、決定した濃度の DHT を添加した後の培養時間を決定するための 290 実験を行った。まず 10 cm ディッシュ 6 枚に FB を播種し(5 × 105 cells/ディ 291 ッシュ)、おおよそすべての細胞が 80%コンフルエント近くに達した段階で、3 292 枚には DHT を添加し、残り 3 枚にはメタノールのみを添加した。そして、 293 DHT 添加細胞 1 枚と DHT 非添加細胞 1 枚を 1 セットとし、それぞれ 24 時 294 間、48 時間、72 時間培養した後、トリプシン処理により細胞を回収した。そ 295
して上記と同様の手順で、total RNA 抽出、電気泳動、cDNA 合成を行った。
296 そして前述の通りの方法で qRT-PCR を行った。増幅反応は triplicate で測定し 297 た。得られたデータをもとに、ΔΔCt 法によって、DHT 添加なし(メタノー 298 ルのみ)の場合と比較して最もAR 遺伝子の発現を増加させる DHT 添加後の 299 培養時間を決定した。 300 301 7. AIS 患者由来皮膚 FB の評価 302 AIS 患者由来皮膚細胞における相対的アンドロゲン作用不全の確認のた 303 め、条件検討によって決定した濃度の DHT を添加して培養したのち、外性器 304 皮膚の既知アンドロゲン標的遺伝子であるAPOD 遺伝子の発現量をコントロ 305 ール 4 名と比較した。まず 10 cm ディッシュに各細胞あたり 2 枚ずつ FB を播 306
20 種し(5 × 105 cells/ディッシュ)、おおよそすべての細胞が 80%コンフルエン 307 ト近くに達した段階で、1 枚には DHT を条件検討で決定した濃度で添加し、 308 残り 1 枚にはメタノールのみを添加した。そして条件検討で決定した時間の培 309 養を行ったのち、トリプシン処理により細胞を回収した。そして上記と同様の 310
手順で total RNA 抽出、電気泳動、cDNA 合成を行った。そして前述の通りの
311 方法で qRT-PCR を行った。増幅反応は triplicate で測定した。得られたデータ 312 をもとに、ΔΔCt 法によって DHT 添加なし(エタノールのみ)の場合と比較 313 したAR 遺伝子の発現増加率を求めた。 314 315 8. 本実験 316 AIS 患児 1 名とコントロール 4 名由来の皮膚 FB をそれぞれ 10 cm ディッ 317 シュ 2 枚ずつに撒き(5 × 105 cells/ディッシュ)、24 時間培養しておよそ 80% 318 コンフルエントになるようにした。その後、1 枚には DHT を条件検討で決定 319 した濃度で添加し、残り 1 枚にはメタノールのみを添加した。そして条件検討 320 で決定した時間の培養を行ったのち、トリプシン処理により 10 検体からそれ 321 ぞれ細胞を回収した。そして上記と同様の手順で total RNA 抽出を行った。 322 323 9. マイクロアレイ解析 324
21
抽出した total RNA は、遺伝子発現解析用マイクロアレイ(SurePrint G3,
325
Human Gene Expression 8 x 60 k v3.0,アジレント・テクノロジー社)に供した。
326
マイクロアレイ解析は、タカラバイオ (滋賀、日本)に委託して行った。得ら
327
れたマイクロアレイデータは NCBI’s Gene Expression Omnibus
328
(https://www.ncbi.nlm.nih.gov/geo/)に登録した(GEO accession number
329 GSE121712)。 330 はじめに、得られたマイクロアレイデータは GeneSpring software (アジレ 331 ント・テクノロジー社)を使い、各サンプルの 75 パーセンタイルをそろえる正 332
規化を行った。次に、Agilent Feature Extraction software を用いて低発現の遺
333 伝子やシグナル検出が不良な検体を含む遺伝子を除外した。具体的には、まず 334 コントロール 4 名 8 検体中 3 検体以上かつ患児 1 名 2 検体中 1 検体以上におい 335 て raw データのシグナル値が 100 よりも大きいものを抽出した。続いて 336 “detected(シグナル検出)”フラグがコントロール 4 名 8 検体全てについてお 337 り、かつ患児 1 名 2 サンプルの少なくとも 1 サンプルについている遺伝子を抽 338 出した。抽出された遺伝子毎に患児 1 名とコントロール 4 名それぞれの DHT 339 添加時のシグナル値を DHT 非添加時(メタノールのみ)のシグナル値で割っ 340 た値(fold change:FC)を計算し、底2で対数変換を行った。 341 次の 5 条件全てを満たす遺伝子をアンドロゲン標的候補遺伝子と定義した。 342
22
i) コントロール 4 名の平均|log2 FC| ≥ 0.3
343
ii) コントロール 3 名以上で各|log2 FC| ≥ 0.3
344
iii) 患児の|log2 FC| < コントロール 4 名の平均|log2 FC|
345 iv) コントロール 4 名全てで log2 FC の正負が一致 346 v) タンパク質をコードしている遺伝子 347 この log2 FC 0.3(FC 1.2)というカットオフは、先行研究で神経幹細胞における 348 アンドロゲン標的遺伝子の抽出の際に設定されたカットオフを参考に決定した 349 [14]。AIS 患者とコントロール 4 名由来の皮膚 FB における DHT 添加による 350 APOD の発現を qRT-PCR 法で評価した実験結果を用いて、この絞り込み基準 351 のシミュレーションをすると、APOD が“アンドロゲン標的候補遺伝子”として 352 抽出されることを確認した。 353 コントロール 4 名における各遺伝子の DHT 添加時のシグナル値と非添加 354 時 のシグナル値の統計学的有意差の有無は、 エクセル および GeneSpring 355 software (アジレント・テクノロジー社)を用いてスチューデントの t 検定を行い 356 評価した。GeneSpring では多重比較検定補正(Benjamini-Hochberg 法)も行っ 357 た。 358 359 10. アンドロゲン標的候補遺伝子の評価 360
23 抽 出 さ れ た ア ン ド ロ ゲ ン 標 的 候 補 遺 伝 子 に つ い て 、 第 一 に Pubmed 361 (https://www.ncbi.nlm.nih.gov/pubmed/)で既報にあたり、アンドロゲン作用や 362 他の性ホルモンとの関連が報告されているか否かを調べた。検索は、論文発行年 363 の 制 限 は 設 け ず 、 各 遺 伝 子 名 364
に ”function” 、 ”androgen” 、 ”testosterone” 、 ”male” 、 ”external genitalia
365 development”、”estrogen”、”steroid”などの語句を組み合わせて行った。検索で 366 みつかった文献に掲載されている参考文献も適宜参照した。 367 第二に、当該遺伝子のイントロンおよびエクソン領域に ARE 候補配列が存 368
在するか否かを調べた。ARE 配列は、既報で chromatin immunoprecipitation data
369 に基づき ARE と報告されている 15-mer(AGAACANNNTGTTCT)を用いた 370 [5]。ARE は回文配列状に並んだ 6 塩基 2 つ(AGAACA と TGTTCT)と、そ 371 れらをつなぐ任意の 3 塩基(NNN)から成る。この ARE と 12-mer 以上一致 372 (任意の配列 3 塩基を除くと 9-mer 以上一致)しているものを ARE 候補配列と 373 定義した。 374
第三に、NCBI Gene Expression Omnibus データベースに登録されている正
375 常男性の外性器(主に陰嚢)由来皮膚 FB と AIS 患者の大陰唇/陰嚢由来皮膚 FB 376 での遺伝子発現に関するマイクロアレイデータ(GSE3872、GSE6796、GSE6797) 377 を NextBio (https://www.nextbio.com/) (イルミナ社)を用いて参照した。最後 378
24
に、DAVID 6.8 (https://david.ncifcrf.gov/)を用いて gene ontology(GO)解析
379 を行った[25, 26]。 380 381 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396
25 第Ⅴ章 研究結果 397 1. 研究対象 398 対象者の選択フローを図 3 に示す。外性器皮膚検体の提供に同意した尿道 399 下裂患者 57 名に対して次世代シークエンス解析を行い、1 名にAR 遺伝子の 400
c.1789G>A, p.Ala597Thr 変異(RNA transcript ID:ENST00000374690.9)を
401 ヘミ接合性に同定した。この変異は直接シークエンス法で確認した(図 4)。患 402 児は出生時より尿道下裂を指摘され、月齢 18 のときに修復術を受けている。 403 この患児 1 名を AIS 患者として本研究の対象とした。 404 また、AIS 患児と年齢が近く、外性器皮膚検体の提供に同意した埋没陰茎 405 4 名では、直接シークエンス法で解析したAR 遺伝子に変異は認めなかった。 406 この 4 名は外性器修復術を月齢 6 から月齢 21 のときに受けていた(表 3)。こ 407 れら 4 名をコントロール群として本研究の対象とした。 408
AIS 患者で同定したAR 遺伝子 p.Ala597Thr 変異は DNA 結合ドメインに
409 位置する。この変異は独立した家系の複数の PAIS 患者で報告のある既報変異 410 である[27, 28]。初報告は 1992 年に Klocker らによって報告された独立した 2 411 家系 5 名の PAIS 患者である[27]。そのうち 1 家系 3 兄弟の陰嚢皮膚由来 FB 412 に対するトリチウム標識 DHT の結合能評価の結果、DHT に対する変異 AR タ 413 ンパク質の解離定数および最大結合部位数は健常男児と同等であったと報告さ 414
26 れている。一方、変異 AR タンパク質の転写活性能は、野生型と p.Ala597Thr 415 変異型 AR 発現ベクターをレポーターベクターすなわちホルモン応答配列を持 416 つプロモーター制御下クロラムフェニコール(CAT)発現ベクターと共に 417 COS-7 細胞に一過性発現させた系で評価され、変異型 AR の転写活性能は野生 418 型の約 1/3 であったと報告されている[27]。AR タンパク質の DNA ドメイン 419 は 2 つのジンクフィンガーから成り、1 つ目のジンクフィンガーは proximal 420
box (P-box)と呼ばれ、結合する ARE 配列の識別に働く[29]。そして 2 つ目の
421 ジンクフィンガーは distal box(D-box)と呼ばれ、AR 二量体化に関与する。 422 D-box の中でも特に 5 つのアミノ酸が重要であり、Ala597はこの 5 つのアミノ 423 酸のうちの一つである(図 5)。従って、p.Ala597Thr 変異 AR ではホモ二量体 424 化不全によって転写活性能の低下が生じていると考えられる。 425 426 2. qRT-PCR 法を用いた条件検討結果 427 AR 遺伝子と内部標準遺伝子の増幅効率を比較した結果を図 6 に示す。AR 428 遺伝子とGAPDH、HPRT1、B2M すべてで検量線の傾きが同程度であったこ 429 とから、いずれも AR 遺伝子の発現量をΔΔCt 法で調べる際の内部標準として 430 用いることができると判断した。そこで、以下の条件検討では GAPDH を内部 431 標準として用いた。 432
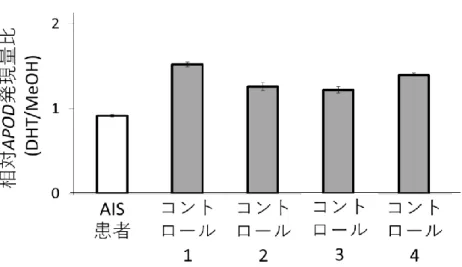
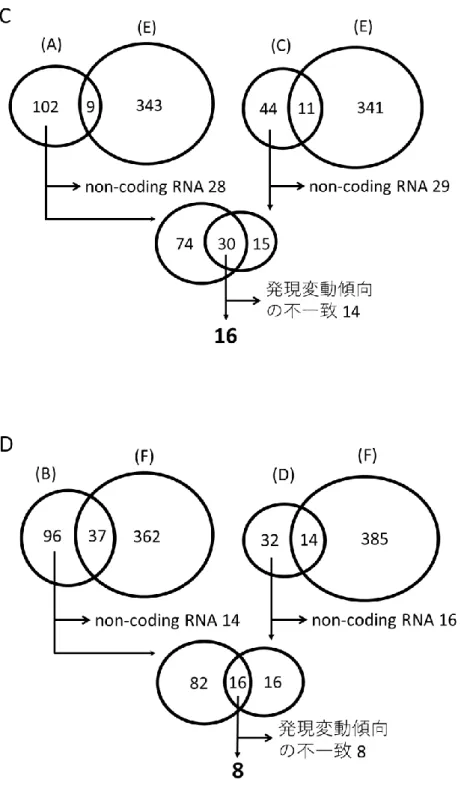
27 DHT をそれぞれ終濃度 0 nM (メタノール)、1 nM, 10 nM, 100 nM で添加 433 した後のAR 遺伝子発現を調べた結果を図 7A に示す。DHT 1 nM 添加によ 434 り、非添加(0 nM)の場合と比べて AR 遺伝子発現は約 2 倍に増加しており、 435 DHT 添加量増加に伴うAR 遺伝子発現のさらなる増加は認めなかった。そこ 436 で DHT 添加濃度は 1 nM に決定した。 437 次に DHT 1 nM 添加後の培養時間の検討を行った。培養時間 24、48、72 438 時間のうち AR 遺伝子発現が DHT 添加によって最も上昇していたのは、72 時 439 間培養であった(図 7B)。そこで培養時間は 72 時間に決定した。 440 441 3. AIS 患者由来皮膚 FB の評価 442 AIS 患者とコントロール 4 名の DHT 1 nM 添加によるAPOD 遺伝子の相 443 対発現量比の結果を図 8 に示す。コントロール 4 名由来皮膚 FB では 1.2~1.5 444 倍のAPOD 遺伝子の発現上昇を認めたのに対し、AIS 患者では DHT 添加によ 445 るAPOD 遺伝子の発現変動がなかった。 446 447 4. マイクロアレイ解析 448 マイクロアレイ解析の結果として、58,341 遺伝子について 5 名 10 検体の 449 シグナル情報を得た。データの正規化の後、まずコントロール 4 名 8 検体で低 450
28 発現遺伝子とシグナル検出が不良な検体を含む遺伝子を除外し、58,341 遺伝子 451 から 16,705 遺伝子を抽出した。次に、その 16,705 遺伝子について 4 名それぞ 452 れの log2FC を求めた。そして 4 名の平均|log2FC |が 0.3 以上であるものを選 453 び、244 遺伝子を得た。このうち DHT 添加により発現が上昇していたのは 454 111 遺伝子、低下していたのは 133 遺伝子であった。また、コントロール 4 名 455 中 3 名以上で|log2FC |が 0.3 以上であるものは 101 遺伝子であった。このうち 456 DHT 添加により発現が上昇していたのは 55 遺伝子、低下していたのは 46 遺 457 伝子であった(図 9A)。同様に、患者 1 名 2 検体についてもデータの正規化後 458 に低発現遺伝子とシグナル検出が不良な検体を含む遺伝子を除外し、58,341 遺 459 伝子から 16,511 遺伝子を抽出した。その後|log2FC |が 0.3 以上であるものを抽 460 出し、751 遺伝子を得た。このうち DHT 添加により発現が上昇していたのは 461 352 遺伝子、低下していたのは 399 遺伝子であった(図 9B)。 462 次に、コントロール 4 名において 2 通りの抽出方法で選ばれた DHT 添加 463 により発現が上昇している遺伝子(図 9A(A)と(C))から AIS 患者でも 464
DHT 添加により発現が上昇していた遺伝子(図 9B(E))と non-coding RNA
465 を除外し、それぞれ 104 遺伝子と 45 遺伝子を得た(図 9C)。その後、それら 466 に共通している 30 遺伝子を選び、その中からコントロール 4 名全てで DHT 467 添加により発現が上昇している 16 遺伝子をアンドロゲン標的候補遺伝子とし 468
29 た。 469 同様に、コントロール 4 名において 2 通りの抽出方法で選ばれた DHT 添 470 加により発現が低下していた遺伝子(図 9A(B)と(D))から、AIS 患者でも 471 DHT 添加により発現が低下していた遺伝子(図 9B(F))と non-coding RNA 472 を除外し、それぞれ 98 遺伝子と 32 遺伝子を得た(図 9D)。その後それらに共 473 通している 16 遺伝子を選び、その中からコントロール 4 名全てで DHT 添加 474 により発現が低下している 8 遺伝子をアンドロゲン標的候補遺伝子とした。以 475 上より、合わせて 24 遺伝子をアンドロゲン標的候補遺伝子として選出した。 476 24 遺伝子の一覧を表 3 に示す。DHT 添加により発現が上昇していた 16 遺伝 477 子のコントロール 4 名の平均 log2 FC は 0.30 から 0.74 であり、DHT 添加によ 478 り発現が低下していた 8 遺伝子のコントロール 4 名の平均 log2 FC は-0.30 から 479 -0.99 であった。24 遺伝子のうち、コントロール 4 名における DHT 添加時の 480 シグナル値と非添加時のシグナル値に統計学的に有意な差を認めた遺伝子はな 481 かった。 482 483 5. アンドロゲン標的候補遺伝子の評価 484 Pubmed を用いた文献検索により、24 遺伝子のうち 11 遺伝子にアンドロ 485 ゲンや他の性ホルモンとの関連を示唆する報告を認めた(表 3)。特にAPOD 486
30 はヒト陰嚢皮膚 FB においてアンドロゲン標的遺伝子として報告されており、 487 Cyp1b1 はマウスの外性器原基からの雄性化に関与すると報告されている。な 488 お除外した 87 個の non-coding RNA には、前立腺がん[30]や乳がん[31]など 489 の悪性腫瘍の増殖に関与すると報告のあるH19 が含まれていた。H19 遺伝子 490 の log2 FC は、患者では 0.02、コントロールではそれぞれ 0.38、0.25、0.27、 491 0.11 であった。 492
ARE 候補配列は 24 遺伝子のうち 7 遺伝子に認め、KIAA1598 と CNIH3
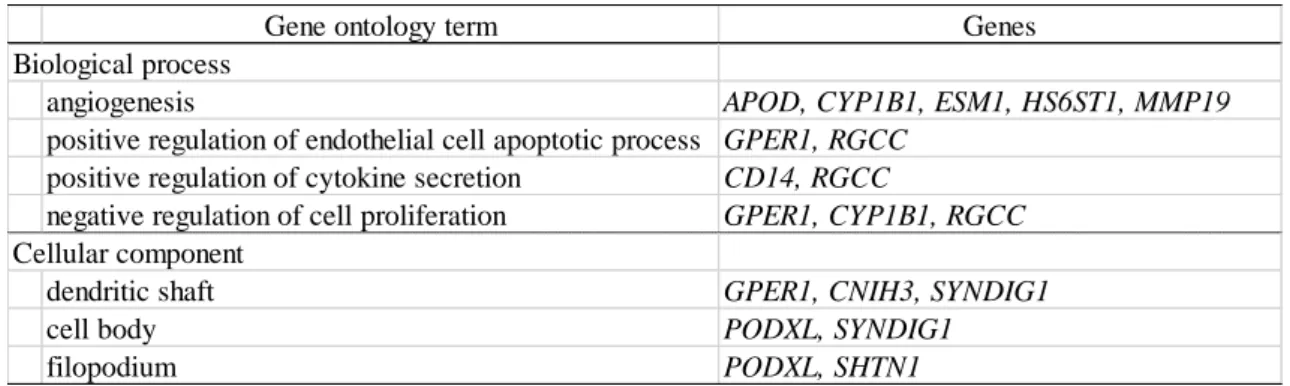
493 は複数の候補配列を周囲に有していた(表 3)。データベースに DHT により発 494 現がコントロール群で上昇していた 16 遺伝子のうち 8 遺伝子は、データベー 495 スに登録されている DHT 非添加のヒト外性器皮膚を用いたマイクロアレイ解 496 析で AIS 患者よりも健常男性において発現が高かった(表 3)。CYP1B1 は今 497 回の研究では DHT 添加によりコントロール群で発現が上昇していたが、デー 498 タベースのマイクロアレイ解析の結果では、正常男性の方が低発現であった。 499 また DHT により発現がコントロール群で低下していた 8 遺伝子のうち 2 遺伝 500 子においてデータベースに登録があったが、いずれも正常男性の方が高発現で 501 あった。GO 解析では、APOD、CYP1B1、HS6ST1 を含む 5 遺伝子が血管新 502 生に関与することが示唆された。その他、細胞のアポトーシスや細胞分化に関 503 与する遺伝子が複数含まれていた(表 4)。 504
31 第Ⅵ章 考察 505 1. 本研究によって得られた知見 506 今回の研究は、DHT 添加による発現変動が小さい遺伝子群にも注目する 507 ことで、既知のヒト外性器皮膚におけるアンドロゲン標的遺伝子の他に新たに 508 23 個のアンドロゲン標的候補遺伝子を同定した(図 10)。これはヒト性分化に 509 おける外性器の発生・分化の分子メカニズム解明の一助となる成果である。今 510 回の研究では、正常コントロール 4 名の外性器皮膚 FB において DHT 添加に 511 より統計学的に有意に変動した遺伝子はなかった。この結果はアンドロゲン誘 512 導性の外性器男性化が少数の遺伝子の大きな発現変動による現象ではないこと 513 を示唆する。今回の研究で同定された 24 個のアンドロゲン標的候補遺伝子の 514 DHT 添加による発現変動幅は比較的小さく、外性器男性化は発現変動幅の小 515 さな遺伝子が多く協調して生じていると推測される。 516 517 2. 同定されたアンドロゲン標的候補遺伝子の機能 518 24 遺伝子にはヒト外性器皮膚においてアンドロゲン標的遺伝子として報告 519 されているAPOD[7]が含まれ、正常コントロール 4 名において DHT 添加に 520 より発現が上昇していた。Hornig らは、ヒト外性器皮膚 FB のAPOD 遺伝子 521
mRNA 測定により AR 機能が推定でき、AR 陰性 AIS 患者の診断に有用である
32 と提唱した[32]。APOD はヒトにおいてユビキタスに発現しており、とくに精 523 巣、脾臓、中枢神経に高発現である[8]。APOD は 169 アミノ酸から成る、19 524 kDa の糖タンパク質であり、構造から疎水的な有機物の運搬体であるリポカリ 525 ンファミリーに属すると考えられている[8]。APOD は細胞外に分泌されて働 526 くと考えられており、たとえばグリア細胞で作られた APOD は細胞外小胞に 527 より分泌され、パラクライン作用によって神経細胞に取り込まれて神経細胞保 528 護に寄与すると報告されている[33]。APOD は複数のリガンド結合能を有し、 529 アンドロゲンやエストロゲン、プロゲステロン、HDL や LDL、アラキドン酸 530 などと結合する[33]。またAPOD はげっ歯類の海綿体を人工的に虚血状態に 531 したときに最も強く発現が誘導される遺伝子であると報告されている[34]。血 532 管内皮細胞はパラクライン作用により近接する血管壁細胞のAPOD 発現量を 533 低下させることで壁細胞の接着を促し、血管新生の開始にはこの内皮細胞と壁 534 細胞の接着を抑制することが必要であることから、APOD 発現量増加は血管新 535 生に寄与すると考えられている[35]。ヒトにおいて外因性にアンドロゲンを投 536 与すると外性器の血管新生が促されることが知られており[36]、また一般に組 537 織の分化・成長は血管新生を伴いながら進行することから、生体内における外 538 性器の分化・成長の際には、アンドロゲン作用により外性器皮膚で発現が増え 539 た APOD がパラクライン作用により近隣の血管内皮細胞や壁細胞に働いて血 540
33 管新生を促している可能性がある。あるいは血管内皮細胞にもアンドロゲン受 541 容体は発現している[37]ことから、アンドロゲンと直接結合した APOD が血 542 管内皮にアンドロゲンを運搬する役割を担っているのかもしれない。 543 また、興味深い遺伝子として DHT 添加によりコントロール群で 6 番目に 544 発現が上昇していたCYP1B1 が挙げられる。Cyp1b1 は実験動物に抗エストロ 545 ゲン様の生殖毒性を生じる催奇性物質 2,3,7,8-tetrachlorodibenzo-p-dioxin 546 (TCDD)のターゲットとして同定され[38]、マウスで TCDD の抗エストロゲ 547 ン作用発揮に関与することが示唆されている[39]。ヒトにおいても CYP1B1 は 548 エストロゲン、テストステロン、プロゲステロンなどのステロイド代謝に関与 549 し、ホルモン不活化作用を有している[40]。しかし DHT は主に 3α-ヒドロキ 550 システロイドデヒドロゲナーゼにより活性の低い 3α-アンドロスタンジオール 551 に変換され[41]、CYP1B1 は DHT 代謝には関与しない。従って、本研究で 552 DHT 添加によりCYP1B1 の発現が上昇した原因は DHT 代謝のためではない 553 と考えられる。この遺伝子はマウスにおいて外性器原基である生殖結節に発現 554 しており、雌雄分化が始まった直後である 15.5 dpc(交配後 15.5 日目)の外性 555 器原基において、雌マウスと比して雄マウスで発現が高かったと報告されてい 556 る[11]。CYP1B1 は前立腺がんや乳がん、卵巣がんなどホルモン関連がん組織 557 において正常組織よりも発現が高いことが知られており、CYP1B1 には Wnt/ 558
34 βカテニンシグナル伝達経路の活性化による細胞増殖を促す機能があると報告 559 されている[42]。本研究は、マウスと同様にヒトにおいても CYP1B1 が細胞増 560 殖作用などを介して外性器の分化・成長に関与している可能性を示唆する。 561 今回最も DHT 添加により発現が上昇していた遺伝子はRGCC であった。 562 この遺伝子は神経幹細胞に DHT を添加し RNA シークエンス法で変動遺伝子 563 の解析をした研究において、DHT 添加により最も発現が上昇していた遺伝子 564 であり、また AR ノックダウンでこの DHT 添加による発現変動は消失したと 565 報告されている[14]。RGCC は 137 アミノ酸から成る 14.7 kDa のタンパク質 566 である。RGCC はユビキタスに発現しており、種々の細胞において増殖に関与 567 すると報告されている[43]。RGCC は血管平滑筋細胞においては、サイクリン 568 依存性キナーゼ p34CDK2の酵素活性を高めることで、細胞周期 S 期への移行を 569 促す[44]。その一方で RGCC は神経膠腫細胞においてはポロ様キナーゼ 1 と 570 複合体を作り、G2/M 移行期で細胞周期を停止させる。したがって RGCC は 571 細胞によって異なる細胞増殖への作用を持っている可能性があり、外性器皮膚 572 細胞の増殖に対してどのように作用するかについては、さらなる検討が必要で 573 ある。 574 そのほかに文献検索の結果、アンドロゲン作用との関連が示唆された遺伝 575 子は以下の 5 遺伝子である。 576
35 SPATA21 は別名 Spergen-2 と呼ばれ、今回の研究で DHT 添加により正 577 常コントロール群の皮膚 FB で 2 番目に発現が上昇していた遺伝子である。 578 Spergen-2 はアミノ酸 500 個から成る 56 kDa のタンパク質をコードする。 579 Spergen-2 は、ラットの精巣に特異的に発現している遺伝子の一つとしてディ 580 ファレンシャル・ディスプレイ法を用いて同定された[45]。Spergen-2 は核移 581 行シグナルと推定される配列を N 端に有し、細胞内では精原細胞の核に局在し 582 ていた[45]。ヒトにおける Spergen-2 の機能はほとんど明らかになっておら 583 ず、今回の研究で DHT 添加により外性器皮膚 FB で遺伝子発現が上昇した意 584 義も不明だが、精子形成はアンドロゲン作用により生じ、Spergen-2 はヒト精 585 巣でも高発現であることから、ヒト精巣における役割について今後さらなる研 586 究が望まれる。 587 RIMBP3(KIAA1666)は前シナプス・アクティブゾーンタンパク質 RIM 588 および電位依存性カルシウムチャネルの結合分子として同定された RIMBP の 589 一つである。RIMBP1 および RIMBP2 は主に脳に特異的に発現するのに対 590 し、最後に見つかった RIMBP3 はマウスにおいて精巣特異的に発現し精子の頭 591 部の形成に関与していると報告されている[46]。ヒトにおけるアンドロゲン感 592 受性組織における報告は本研究が初めてであり、その意義については今後の検 593 証が必要である。 594
36 HS6ST1 はヘパラン硫酸の 6 位の水酸基に硫酸基を転移する反応を担うヘ 595 パラン硫酸 6 スルホトランスフェラーゼの一種である。ヘパラン硫酸は付加さ 596 れている硫酸基の状態に応じて成長因子シグナル伝達経路の調節に関与する。 597 胎生 16 日目の雄マウス尿生殖洞では、雌マウス尿生殖洞よりもヘパラン硫酸 598 が硫酸化されており、またHs6st1 は発生段階の雄マウス前立腺に強く発現し 599 ていることから、Hs6st1 はアンドロゲン作用に伴う前立腺発生に関与している 600 と推測されている[47]。本研究においても DHT 添加により正常コントロール 601 群の外性器皮膚 FB において発現が上昇していたことから、外性器皮膚組織の 602 アンドロゲン依存的な増殖に関与している可能性がある。 603 ESM1 は臍帯静脈の上皮細胞由来 cDNA ライブラリから見つかった遺伝子 604 であり、上皮細胞特異的分子 1、別名エンドカンというタンパク質をコードし 605 ている。エンドカンは 50 kDa のプロテオグリカンであり、活発に分裂増殖し 606 ている組織に発現が高いと報告されている[13]。前立腺がん組織の検討では、 607 正常の前立腺組織に比べてESM1 の発現が増加しており、ESM1 発現は AR 発 608 現と相関していたと報告されている[13]。本研究では DHT 添加により正常コ 609 ントロール群の外性器皮膚 FB で発現が低下していたことから、組織によって 610 異なる作用を持つ可能性が考えられる。 611 GPER1 は 7 回膜貫通型 G タンパク質共役受容体であり、エストロゲン作 612
37 用のうちエストロゲン受容体を介さない非古典的経路(non-genomic 613 pathway)に関与すると報告されている。スナネズミの精巣上体の免疫染色に 614 よる検討において、繁殖期には精巣上体の主細胞と基底細胞の細胞質に 615 GPER1 は強く染色された。一方、非繁殖期では染色が減弱していた。また去 616 勢により GPER1 は核に局在し、テストステロン補充により細胞質にも局在す 617 るようになったことから、テストステロンもGPER1 の発現制御に一部関与し 618 ている可能性があると報告されている[12]。ただし本研究においては正常コン 619 トロール群の皮膚 FB において DHT 添加によりGPER1 発現は低下してお 620 り、組織による発現の違いや種差が影響している可能性がある。 621 またアンドロゲン以外の性ホルモンとの関連が文献的に報告されている遺 622 伝子は以下の 3 遺伝子である。これらについてはアンドロゲン感受性組織や細 623 胞での研究は本研究以外にないことから、アンドロゲン作用との関連について 624 はさらなる研究が必要である。 625
ADH1C はアルコール脱水素酵素(alcohol dehydrogenase:ADH)の 1 種
626 であり、主に肝臓においてエタノールをアセトアルデヒドに変換する役割を持 627 つ[48]。胎内で母体を介して活性の高いエストロゲンであるエストラジオール-628 17βに曝露された雄ブタの前思春期の前立腺において ADH1C の発現が低下し 629 ていたと報告されている[49]。エストロゲン作用とアンドロゲン作用はしばし 630
38 ば拮抗的に働くことから、生理的な意義は明らかではないが、本研究において 631 アンドロゲン作用により発現が上昇していたことはこの先行研究の結果と矛盾 632 しないと考える。 633 IMPA2 はイノシトール合成酵素であるイノシトールモノフォスファター 634 ゼ(IMPase)をコードしている。IMPA2 は双極性障害の連鎖解析で疾患感受性 635 遺伝子座とされた 18p11.2 に存在しており、IMPA2 のプロモーター領域の遺 636 伝子多型と双極性障害発症との関連が報告されている[50]。IMPase は双極性 637 障害の治療薬として用いられるリチウムにより抑制され、イノシトールおよび 638 その代謝産物の産生が減ることで治療効果を発揮すると考えられている[51]。 639 またIMPA2 は受精卵が着床しうる増殖した子宮内膜において、前増殖期の子 640 宮内膜と比較して遺伝子発現が増加していたと報告されている[52]。 641 MMP19 は亜鉛やカルシウムなどの金属を必要とするタンパク質分解酵 642 素、マトリックスメタロプロテアーゼをコードしている。hCG 刺激によって、 643 ヒト卵胞の顆粒膜細胞および莢膜細胞でMMP19 発現が上昇することから、 644 MMP19 は排卵に関与していると推測されている[53]。 645 以下に記す 13 遺伝子については、検索し得た範囲ではアンドロゲンや他 646 の性ホルモンとの関連を示唆する文献はなかった。そのうちSQRDL、NFE2、 647 CD14 の 3 遺伝子は、データベースの健常男性皮膚において AIS 患者と比べた 648
39 発現レベルが本研究の結果と同様に高かったことから、アンドロゲン作用と関 649 係している可能性があるが、今後の検証が必要である。以下これら 10 遺伝子 650 について、現時点まで報告されている事柄を簡単に記す。 651 SQRDL(別名 SQOR)はミトコンドリアで硫化水素の代謝に関与する酵 652 素である。硫化水素は生体内で作られるガス状伝達物質で、心血管系などにお 653 いけるシグナル伝達に関与する[54]。 654 KIAA1598(別名 SHTN1、Shootin1)には、2 つのアイソザイムがある。 655 1 つは主に脳に発現しているShootin1a であり、神経の極性化と軸索伸長に重 656 要な役割を果たす[55]。もう 1 つのアイソザイムは上皮細胞を含む全身に発現 657 しているShootin1b だが、初報告が 2016 年であり各組織でどんな役割を果た 658 しているかはほとんど明らかになっていない[55]。 659 NFE2 は巨核球の成熟および血小板産生に関与する転写因子であり、NFE2 660 ノックアウトマウスは重度の血小板減少による出血により死亡する[56]。ヒト 661 では造血幹細胞の機能維持や T 細胞系の分化にも関与する[57]。 662 CD14 はグリコシルホスファチジルイノシトールアンカー型タンパク質で 663 あり、単球、マクロファージ、好中球、B 細胞、樹状細胞、上皮細胞などに発 664 現している。CD14 はグラム陰性細菌の表面に存在するリポ多糖を認識・運搬 665 することでマクロファージなどの免疫細胞を活性化するシグナル伝達の中心的 666
40 役割を担っている[58]。 667 SAP25 はヒストン脱アセチル化酵素 Sin3 と結合してタンパク質複合体を 668 作り転写抑制に関与する分子 SAP25 をコードする遺伝子として同定された。 669 SAP25 は核と細胞質を流通するシャトルタンパク質と考えられている[59]。 670 HSPB3 は分子シャペロンとして働く熱ショックプロテインの一つをコー 671 ドしており、末梢神経に発現している[60]。HSPB3 のミスセンス変異による 672 遺伝性ニューロパチーも報告されている[61]。 673
SRMS は別名 PTK 70(Protein tyrosine kinase 70)とも呼ばれる非受容体
674
型チロシンキナーゼをコードする。ホスフォプロテオミクス解析により、細胞
675
増殖を抑制しアポトーシスを促す機能を持つことから腫瘍抑制因子とされる
676
DOK1(downstream of tyrosine kinases 1/docking protein 1)が基質であるこ
677 とが報告されている[62]。 678 PODXL は CD34 ファミリーに属する膜貫通型糖タンパク質をコードす 679 る。PODXL は糸球体上皮細胞および足細胞、造血幹細胞、血管上皮細胞など 680 に発現し、細胞接着阻害に関与する[63]。PODXL の過剰発現は乳がんや前立 681 腺がん[64]、肝臓がんなどの腫瘍において悪性度の高さや不良な予後と関連す 682 ると報告されている[65]。 683 SYNDIG1 は早いシナプス応答を担う AMPA 型グルタミン酸受容体に直接 684
41 結合して興奮性シナプスの成熟を助けるタンパク質をコードする[66]。 685 KRT86 は TypeII 毛髪ケラチン hHb6 をコードし、常染色体優性遺伝形式 686 の毛髪疾患である連珠毛の家系の連鎖解析により同定された[67]。 687
CNIH3 は CNIH2 と共に中枢神経に広く分布する AMPA 型グルタミン酸
688 受容体に結合し複合体を構成し、AMPA の不活化と脱感作の速度を調節してい 689 る CNIH3 をコードする[68]。 690 KIF24 はセリンスレオニンキナーゼ Nek2 にリン酸化されると構造変化を 691 起こして活性型となり、一次繊毛形成を阻害して有糸分裂を生じさせる KIF24 692 タンパク質をコードする[69]。また原因不明の続発性無月経患者のマイクロア 693 レイ解析による検討で KIF24 遺伝子を含む領域(9 番染色体短腕 13.3)185 kb 694 に重複を認め、KIF24 過剰発現と卵巣機能不全の関連が示唆されている[70]。 695
DOCK5 は DOCK ファミリーの一種で、低分子量 GTPase である Rac1 の
696
グアニンヌクレオチド交換因子として働く dedicator of cytokinesis protein 5 を
697 コードする。DOCK5 は破骨細胞、好中球、肥満細胞などに発現する[71]。 698 その他、除外した 87 個の non-coding RNA にはH19 遺伝子が含まれてい 699 た。H19 はコントロール群において DHT 添加により発現が上昇していた。 700 H19 は発生過程の胚において発現が高く、母由来アリルからのみ発現するイン 701 プリンティング遺伝子である[72]。成人においては前立腺がん[30]や乳がん 702
42 [31]など様々ながんの増殖にも関与するとして注目されている。H19 の RNA 703 は 5 エクソンから成る。最初のエクソンに腫瘍抑制遺伝子の retinoblastoma 704 (RB) 遺伝子発現を抑制することで細胞増殖を促進する microRNA675 705 (miR675) が存在する[73]。また 2-5 エクソン断片は版抑制遺伝子として働く 706 let-7 microRNA の生物活性を抑えることで、細胞増殖を促すと考えられている 707 [74]。 708 データベースに登録のあるマイクロアレイ解析では、10 遺伝子が AIS 患 709 者よりも正常男性の外性器皮膚における発現が高く、1 遺伝子は逆に正常男性 710 の外性器皮膚における発現の方が AIS 患者よりも低かった(表 3)。このうち、 711 RGCC、ADH1C、IMPA2、SQRDL、APOD、NFE2、CD14、HSPB3 は本研 712 究の発現変動パターンと一致していた。しかしCYP1B1 と ESM1、DOCK5 は 713 本研究と異なる変動パターンであった。乖離の理由としては、データベースに 714 登録されているマイクロアレイ解析では DHT 添加はしておらず、定常状態に 715 おける男性外性器皮膚を用いているという点が考えられる。すなわち生理的な 716 in vivo でのアンドロゲン作用と薬理的な in vitro でのアンドロゲン作用との違 717 いによって遺伝子発現に差異が出た可能性がある。また生体内では、アンドロ 718 ゲンだけでなく、テストステロンから変換されて生じるエストロゲンなど他の 719 ホルモンの影響も受けている影響も排除できない。 720
43 AR 変異陰性例で AIS を発症する機序として、転写共役因子や AR 標的遺 721 伝子および AR シグナル経路に関わる分子の質的・量的異常が関与している可 722 能性が想定されている[19, 75]が、特定の疾患原因分子は同定されていない。 723 本研究で同定された 24 のアンドロゲン標的遺伝子の中に、AR 変異陰性の臨床 724 的 AIS 患者の新たな疾患原因遺伝子が含まれる可能性がある。 725 一般的に受容体異常症の重症例では治療に難渋することが多い。例えばイ 726 ンスリン受容体異常症ではインスリン作用不全による高血糖を呈し、重症例で 727 は幼児期に死亡する[76]。高用量インスリンを用いても十分な血糖降下が得ら 728 れない患者では IGF-1 治療が試みられることがある。これは IGF-1 がインスリ 729 ン受容体を介さずにインスリン様の血糖降下作用を生じるためである[77]。こ 730 の例のように、本研究の結果が AR 受容体の下流で働く分子メカニズム解明に 731 寄与すれば、AR 異常のためにアンドロゲン補充療法では十分な男性化が認め 732 られない AIS 患者のために AR を介さずにアンドロゲン作用を発揮する新規治 733 療薬の開発につながる可能性がある。 734 735 3. 本研究の限界 736 本研究には次の 4 つの限界がある。一つ目は、研究対象となる AIS 患者が 737 1 名しか見つからなかったことである。これにより変異による AR 機能不全に 738
44 よる影響と個人のばらつきを区別することはできず、同定した 24 個のアンド 739 ロゲン標的候補遺伝子の中に偽陽性、あるいは除外した遺伝子の中に偽陰性が 740 含まれている可能性は否定できない。AIS 患者としても、理想的には完全に 741 AR 機能を喪失している CAIS 患者の検体を用いることが望ましいが、診断的 742 に外性器皮膚を採取することが一般的ではなくなった現在においては、今後も 743 使用可能な CAIS 由来検体を用いることは期待できない。今後は正常男児由来 744 皮膚 FB のAR をノックアウトあるいはノックダウンした皮膚 FB を用いた検 745 証を検討したい。二つ目に、本研究では培養皮膚 FB を用いている。分化後の 746 皮膚 FB には生殖結節のような多分化能はなく、従って本研究の結果は多様な 747 細胞間での相互作用が生じうる生理的な外性器の状態を完全には反映していな 748 いと考えられる。三つ目に、本研究では DHT 添加による比較的短期的な影響 749 に着目して調べた。しかし外性器の男性化には、長期的なアンドロゲン曝露に 750 よる連続的な事象が寄与する可能性がある。四つ目に、アンドロゲンによる男 751 性化作用には多くの転写因子や転写調節因子が関わる[2]。従って本研究のみで 752 は同定された遺伝子の全てが AR を介して作用していると断定することはでき 753 ず、24 遺伝子の作用機序について今後の検証が必要である。今後は検体数を増 754 やした追試と、同定した候補遺伝子のノックダウンや過剰発現などの in vitro 755 実験により各遺伝子が外性器皮膚で果たす役割についての検討を深めたい。 756
45 第Ⅶ章 結論 757 ヒト外性器におけるアンドロゲン標的候補遺伝子を 24 個同定した。本研 758 究の結果は、アンドロゲンによる外性器男性化の主体が、少数の特定遺伝子の 759 大きな発現変動ではなく、多数の遺伝子の微細変化であることを示唆する。 760 761 762 763 764 765 766 767 768 769 770 771 772 773 774
46 第Ⅷ章 謝辞 775 本研究を遂行するにあたり,御指導を頂きました以下の方々に深く御礼申し上 776 げます。 777 国立成育医療研究センター分子内分泌研究部:深見真紀先生、宮戸真美先生、 778 五十嵐麻紀先生、鳴海覚志先生、三代川温子様.国立成育医療研究センター免 779 疫アレルギー・感染研究部:松本健治先生、原真理子先生.国立成育医療研究 780 センター研究所所長:松原洋一先生.東北大学大学院医学系研究科小児病態学 781 分野:呉繁夫先生.名古屋市立大学大学院医学研究科 小児泌尿器科学分野: 782 林祐太郎先生、水野健太郎先生.福島県立医科大学医学部 泌尿器科学講座: 783 小島祥敬先生. 784 785 また,本研究の実施にあたり御協力下さいました患者様とその御家族に深く感 786 謝を申し上げます. 787 788 789 790 791 792
47
第Ⅸ章 参考文献
793
1.Grino PB, Griffin JE, Wilson JD (1990) Testosterone at high concentrations
794
interacts with the human androgen receptor similarly to dihydrotestosterone.
795
Endocrinology 126: 1165-1172.
796
2.Heemers HV, Tindall DJ (2007) Androgen receptor (AR) coregulators: a
797
diversity of functions converging on and regulating the AR transcriptional
798
complex. Endocr Rev 28: 778-808.
799
3.Pratt WB, Toft DO (1997) Steroid receptor interactions with heat shock
800
protein and immunophilin chaperones. Endocr Rev 18: 306-360.
801
4.Jenster G, Trapman J, Brinkmann AO (1993) Nuclear import of the human
802
androgen receptor. Biochem J 293 ( Pt 3): 761-768.
803
5.Wilson S, Qi J, Filipp FV (2016) Refinement of the androgen response
804
element based on ChIP-Seq in androgen-insensitive and androgen-responsive
805
prostate cancer cell lines. Sci Rep 6: 32611.
806
6.Holterhus PM, Hiort O, Demeter J, Brown PO, Brooks JD (2003) Differential
807
gene-expression patterns in genital fibroblasts of normal males and 46,XY
808
females with androgen insensitivity syndrome: evidence for early programming
809
involving the androgen receptor. Genome Biol 4: R37.
48
7.Appari M, Werner R, Wunsch L, Cario G, Demeter J, et al. (2009)
811
Apolipoprotein D (APOD) is a putative biomarker of androgen receptor
812
function in androgen insensitivity syndrome. J Mol Med (Berl) 87: 623-632.
813
8.Rassart E, Bedirian A, Do Carmo S, Guinard O, Sirois J, et al. (2000)
814
Apolipoprotein D. Biochim Biophys Acta 1482: 185-198.
815
9.Simard J, Veilleux R, de Launoit Y, Haagensen DE, Labrie F (1991)
816
Stimulation of apolipoprotein D secretion by steroids coincides with inhibition
817
of cell proliferation in human LNCaP prostate cancer cells. Cancer Res 51:
4336-818
4341.
819
10.Koistinen H, Hautala LC, Seppala M, Stenman UH, Laakkonen P, et al.
820
(2009) The role of glycodelin in cell differentiation and tumor growth. Scand J
821
Clin Lab Invest 69: 452-459.
822
11.Nishida H, Miyagawa S, Matsumaru D, Wada Y, Satoh Y, et al. (2008) Gene
823
expression analyses on embryonic external genitalia: identification of regulatory
824
genes possibly involved in masculinization processes. Congenit Anom (Kyoto)
825
48: 63-67.
826
12.Menad R, Fernini M, Smai S, Bonnet X, Gernigon-Spychalowicz T, et al.
827
(2017) GPER1 in sand rat epididymis: Effects of seasonal variations, castration