糸状菌由来自然免疫増強化合物 gonytolide A の
TNF経路に対する作用の解析
著者
阿部 由布子
学位授与機関
Tohoku University
平成 23 年度修士論文
糸状菌由来自然免疫増強化合物 gonytolide A の
TNF 経路に対する作用の解析
東北大学大学院薬学研究科
分子薬科学専攻 医薬資源化学分野
学籍番号 B0YM1003 阿部 由布子
目次
序論 1 本論 第一章 Gonytolide A のショウジョウバエにおける自然免疫増強作用 10 の解析 第一節 DAP 型ペプチドグリカンに依存した gonytolide A の自然免疫増強 11 作用の評価 第二節 ショウジョウバエにおける自然免疫増強作用の評価 13 第三節 抗菌ペプチドの発現に対する作用 15 第四節 考察 17 第二章 Gonytolide A のヒト臍帯静脈血管内皮細胞 (HUVEC) における 18 自然免疫増強作用の解析 第一節 各種刺激下での gonytolide A のサイトカイン産生に対する作用の解析 19 第一項 TNF-α 刺激によるサイトカイン産生に対する作用 19 第二項 LPS および IL-1β 刺激によるサイトカイン産生に対する作用 21 第三項 Gonytolide A の IL-8 の発現に対する作用 24 第四項 TNF-α 刺激下での gonytolide A による免疫関連遺伝子の発現 26 第二節 TNF 経路における gonytolide A の作用 28 第一項 TNF 経路関連シグナル伝達因子のリン酸化状態への作用 30 第二項 核抽出物の NF-κB 活性に対する作用の評価 32 第三項 MAP kinase に対する作用 34 第三節 考察 37第三章 Gonytolide A による遺伝子発現変動の網羅的解析 39 第一節 Gonytolide A 単独処理で発現が変化する遺伝子 40 第一項 Gonytolide A 単独処理時の遺伝子発現変動 40 第二項 遺伝子のノックダウンによる gonytolide A の作用の評価 44 第二節 TNF-α 刺激下での gonytolide A による遺伝子発現変動 46 第三節 考察 49 結語 50 実験の部 52 参考文献 63 謝辞 70
本論文中において以下の略記を用いた.
ATF : activating transcription factor BSA : bovine serum albumin
CEBPB : CCAAT enhancer-binding protein COX : cyclooxygenase
DDIT : DNA damage-inducible transcript DAP : diaminopimelic acid
DMSO : dimethyl sulfoxide DNA : deoxyribonucleic acid Dpt : Diptericin
ELISA : enzyme-linked immunosorvent assay ERK : extracellular signal-regulated kinase FADD : Fas-associated protein with death domain FBS : fetal bovine serum
HRP : horseradish peroxidase HS : heat shock
HUVEC : normal human umbilical vein endothelial cell ICAM : Intercellular Adhesion Molecule
IFN : interferon IκB : inhibitor of κB IKK : inhibitor of κB kinase IL : interleukin
IMD : immuno deficiency
iNOS : inducible nitric oxide synthase
IRAK : interleukin-1 receptor-associated kinase JNK : c-Jun N-terminal kinase
KLF : kruppel-like factor LPS : lipopolysaccharide
mTOR : mammalian target of rapamycin MyD : myeloid differentiation factor
NF-κB : nuclear factor κB
NR4A2 : nuclear receptor subfamily 4, group A, member 2 PAMPs : pathogen-associated molecular patterns
PGN : peptideglycan
PGRP : peptideglycan recognition protein RIP : receptor-interacting protein RNA : ribonucleic acid
SDM : Schneider’s Drosophila medium
TAK1 : transforming growth factor-b activated kinase 1 TIR : Toll/interleukin-1 receptor
TLR : Toll-like receptor TNF : tumor necrosis factor TNFR : TNF receptor
TRAF : TNF receptor associated factor UAS : Upstream Activation Sequence
1
序論
生物は, 長い歴史を通して常に病原微生物の感染の脅威にさらされている. 病原微生物の感染に対抗して自らの身を守り, 種を保存するために, 生物は進 化の過程で多様な生体防御機構を獲得してきた. 例えば, 皮膚は物理的な障壁 として異物の体内への侵入を防ぎ, 病原微生物の感染に対する最初の防御機構 として働く. 上皮を通過し微生物が体内に侵入した場合は,異物を排除するため に様々な免疫機構が働く. 免疫とは元々, 一度感染症にかかると二度目以降は 罹患しにくくなるという経験則に基づき研究されてきた生体防御機構である. 免疫系には大きく分けて自然免疫と獲得免疫が存在し, それぞれ異なるメカニ ズムで生体防御の役割を果たしている. 獲得免疫は, 脊椎動物のみが有する高度な生体防御機構である.1) 獲得免疫は, 個々の病原微生物に対応して働く免疫機構であり, 様々な病原体に対して一律 に働くものではない. これは T 細胞, B 細胞といったリンパ球細胞の遺伝子が 再編成されることによって多様性が生まれ, 特定の病原体を認識する受容体を 持つリンパ球のみが, その病原体の侵入時に対応するためである. 使用された リンパ球は免疫記憶として残るため, 抗原-抗体反応により異物を特異的に認識 して排除した後も, 再び異物が侵入してきた際には速やかに免疫応答すること ができる.2) しかし, 獲得免疫は, 侵入した病原体に対して長期的な防御免疫を もたらすことができるが, 異物に対する抗体が産生されるまでに一週間ほどの 時間を要するのが欠点である. 働くまでに時間がかかる獲得免疫に対して, 自然免疫は異物の侵入後直ちに 働く. 自然免疫は, ほとんどの多細胞生物が普遍的に有しており, そのメカニズ ムはショウジョウバエからヒトに至るまで相同性がある. 3) 病原微生物が上皮を 通過し, 体内へ侵入すると直ちに自然免疫が働く. 自然免疫の最初の反応は, 病 原体を認識することから始まる. 病原体の認識は, 補体を介するものと自然免 疫系の受容体によるものがある. 補体は微生物の近傍で加水分解されることに より病原体と共有結合し, 食細胞受容体のリガンドとなり, 食細胞が病原体を 取り込みやすくさせる. 一方, 自然免疫に関わる受容体は, 病原体に共通して存在する構造, 例えばグラム陰性菌の細胞壁成分であるリポポリサッカライド (lipopolysaccharide: LPS) な ど の 分 子 パ タ ー ン (pathogen-associated molecular patterns: PAMPs) を認識することによって, 限られた数の因子により無数の微生 物を認識し, 病原体の種類に応じたサイトカイン産生を誘導する. 4) サイトカイ ンを介した反応は局所性と全身性に分けられるが, 局所性の反応には感染部位 への白血球の浸潤や, 血管透過性亢進, 血小板の接着などが挙げられる. 白血球 の浸潤は, TNF-α などの刺激により血管内皮細胞上でセレクチン, ICAM-1 のよ うな接着分子の発現が誘導され, 感染部位付近の血管内皮に白血球が接着する ことによって始まる. 続いて, IL-8 に代表されるケモカインが産生されること により, 粘着した白血球は感染組織へ遊走する. 血管透過性の亢進は, COX-2, iNOS の発現が誘導されることで起こる. COX-2 や iNOS はそれぞれプロスタ グランジン類や NO の産生を促進することにより, 血管透過性を亢進して, 感 染部位への血漿の慘出を生じさせる. 血小板は血管内皮細胞上に発現した接着 分子に粘着することで, 感染部位の血流管を閉塞させ, 感染の拡大を防ぐこと に関与する. 一方, 全身性の反応には, 体温上昇や急性期タンパク質の産生など が挙げられる. 体温上昇は, IL-1, IL-6, TNF-α が視床下部に作用することによっ て誘導され, 病原微生物の増殖を抑制するとともに獲得免疫の応答を加速させ る働きがある. 5) C 反応性タンパク質 (CRP), マンナン結合レクチン (MBL) な どの急性期タンパク質の産生は, IL-6 が肝臓に作用することにより誘導される. CRP, MBL は細菌の表層に結合し, オプソニン化や補体の活性化を行う. 自然免疫を逃れたものに対して獲得免疫が働くが, 近年の研究で獲得免疫の 成立に自然免疫の活性化が必須であることが明らかにされた. 6) 獲得免疫の活性 化は, 抗原提示細胞によって細胞表面に提示された異物の断片と主要組織適合 抗原クラス II (MHC-II) の複合体を T 細胞が認識することにより成立する. こ の際, 異物 – MHC II 複合体とそれを認識する T 細胞受容体以外に, CD80 や CD86 などの補助刺激分子が必要とされる. TLR を介した異物認識によって自 然免疫が活性化されると, 補助刺激分子が細胞表面に発現し, 獲得免疫の活性 化につながる. 以上のように, 自然免疫の活性化が獲得免疫の成立を引き起こすことで病原 微生物の感染拡大を防ぐことから, 自然免疫の破綻は生体にとって重篤な傷害
3 獲得免疫の活性化が起こらず致死率が高くなる. 7) 自然免疫の異常は, 遺伝的に 欠失がないときでも起こりえる. 例えば, 免疫が低下することで通常では感染 しないような病原体 (弱毒微生物, 非病原微生物) に感染しやすくなる日和見 感染が例としてあげられる. 加齢や臓器移植後の免疫製剤の使用による免疫の 低下が日和見感染の原因となる. 日和見感染症の治療は, 感染後に抗生物質や 抗炎症剤の投与を行う対症療法が主流である. また, 自然免疫の異常な活性化も, 生体にとって重大な傷害をもたらす. マク ロファージが産生するサイトカインによる反応は, 感染が局所にとどまってい るうちは感染の拡大を抑える効果的な異物排除機構として働く. しかし, 感染 が広がり全身でのサイトカインの産生を起こす場合には, 生体は敗血症と呼ば れる重篤な症状に陥る. 敗血症では, 肝臓や脾臓に存在するマクロファージが サイトカインを産生するよう促され, 全身で血管の拡張や透過性を亢進するこ とによって, 組織への血漿の漏出が広がり, 急激な血圧低下によるショック状 態となる.8) 敗血症性ショックによって, 腎臓, 心臓, 肝臓や肺など, 生命の維持 に必須の臓器は通常の血液供給が途絶え, 最終的に多臓器不全に陥り, 患者の およそ 30% は死に至る. 自然免疫の破綻は重篤な疾患につながることから, 生体防御への自然免疫の 重要性に注目が集まり, 分子機構について盛んに研究されている. 自然免疫研 究はショウジョウバエの Toll 受容体が病原体の認識に関わることが明らかに なったことを契機に発展した. 9) Toll によって活性化される Toll 経路は, ショ ウジョウバエの自然免疫シグナル経路の一つであり, グラム陽性菌や真菌の感 染に対して応答する. Toll 経路の活性化は, 細胞外に存在する PGRP-SA やグ ラム陰性菌結合タンパク質 (GNBP) などが病原体を認識することから始まる. PGRP-SA や GNBP-1 は, 下流のセリンプロテアーゼを活性化し, Spätzle を切 断する.10,11)
切断された Spätzle は Toll 受容体のリガンドとなり, Toll を介し て, dMyD88, さらに Tube へとシグナルを伝える.12)
Tube から serine/threonine kinase の Pelle へと伝達されたシグナルによって Pelle は自己リン酸化を行い, Toll-adaptor 分子複合体から解離する. その後, 細胞内シグナル伝達によって, 核内移行阻害因子 Cactus がリン酸化を受けて分解され, Cactus に阻害を受け ていた Rel タンパク質の DIF や Dorsal が核内へ移行し, Drosomycin などの 発現を誘導する. 13)
哺乳類において, Toll 経路と相同性が高い経路として発見されたのが TLR 経路である (Figure 1). 14)
Toll 経路と TLR 経路は非常に類似しており, dMyD88 は MyD88 の, Pelle は IRAK の, Cactus は IkB-α の, Dorsal, DIF は NF-κB の ホモログである. 15-18) ショウジョウバエの Toll 経路とは異なり, TLR 経路では
TLR が直接異物を認識してシグナル伝達が始まる. TLR はファミリーを形成し, 現在までに 11 種報告されている.19)
各 TLR はそれぞれ異なる異物を認識す る. TLR が異物を認識すると TLR の Toll/IL-1R (TIR) ドメインに結合したア ダプター分子 MyD88 へ, さらに MyD88 から serine/threonine kinase である IRAK へとシグナルが伝達される.20)
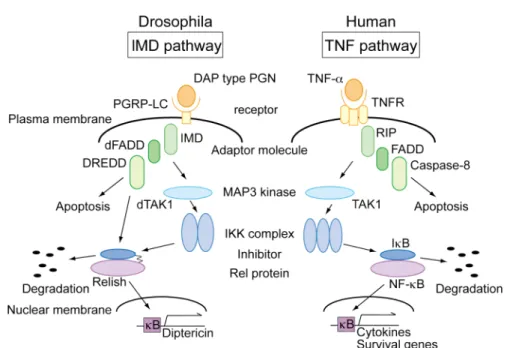
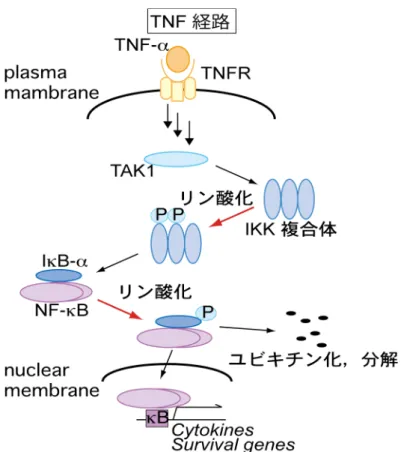
活性化された IRAK は自己リン酸化を行 い, TRAF6 と複合体を形成した後に, MAP3 kinase である TAK1 を活性化する. TAK1 は IKK 複合体のリン酸化を誘導し, 活性化した IKK 複合体が IκB-α を リン酸化することで IκB-α のユビキチン化, プロテアソームによる分解が誘導 される.21) その結果, IkB-α により阻害されていた NF-κB が活性化されて核内 に移行し, TNF-α などの種々の炎症性サイトカインや生体防御タンパク質が誘 導される. 22) また, ヒトとショウジョウバエで共通の自然免疫シグナリングとして, ヒト の TNF 経路とショウジョウバエの IMD 経路がある (Figure 2). 23) TNF 経路の 受容体である TNFR1 は TNF-α の結合によって RIP, TRAF2, そして MAP3 kinase で あ る TAK1 へ シ グ ナ ル伝 達す る. 24,25)
活性 化し た TAK1 は IκB kinase (IKK)-α/β, IKK-γ 複合体をリン酸化により活性化する. 26) リン酸化された
IKK-β は TLR 経路と同様, IκB-α をリン酸化し,27) 最終的に NF-κB が核内へ
移行し, 炎症性サイトカイン IL-8, COX-2 などの発現誘導が起こる. 28) また,
TNFR1 から伝達されるシグナルは TNF 経路以外にも数種の経路を活性化す ることが知られている. 腫瘍壊死因子である TNF-α はがん細胞では TRADD から FADD, caspase-8 へシグナルを伝達し, アポトーシスを誘導する. 29) また
TNF-α は MAP kinase 経路 を活性化することも知られている. MAP kinase 経 路は細胞の生存, 分化, 増殖, アポトーシスに深く関わっており, 30)
ERK1/2 経 路, 31)
JNK 経路, 32)
p38 経路33)の三種に大別される. いずれも TNF-α 刺激によ
って活性化し, MAP3 kinase が MAP2 kinase を, MAP2 kinase が MAP kinase を それぞれリン酸化することでシグナルを増強しながら伝達し, 最終的に活性化 した MAP kinase が下流の転写因子などを活性化する.
5
Figure 1. ショウジョウバエ Toll 経路とヒト TLR 経路
一方, ショウジョウバエの IMD 経路は PGRP-LC がグラム陰性菌の細胞壁 成分を認識することで始まる. PGRP-LC はアダプター分子である IMD を介し て dTAK1 にシグナルを伝達する. 34)
dTAK1 は DmIKK (dIKK-α, dIKK-β) 複合 体をリン酸化する.35) また, 活性化シグナルは dFADD を介して caspase であ
る DREDD を活性化する.36)
IMD 経 路 の転 写因 子であ る Rel タンパク 質 Relish は DmIKK 複合体によりリン酸化され, DREDD によって アンキリンリ ピートが切断される. Relish の転写調節ドメインが核内に移行することにより Diptericin などの抗菌ペプチドの発現が誘導される.37)
この IMD 経路と TNF 経路では, IMD は RIP と, dFADD は FADD と, DREDD は caspase-8 と, dmIKK は IKK と, Relish は NF-κB とそれぞれ非常に類似している. 38-42)
これまでの研究に関わらず, 自然免疫機構の詳細は未だ解明されていない部 分も多い. したがって, これらのシグナル伝達経路を制御する化合物を探索し, その化合物の機構を解析することで, 自然免疫の制御機構の解明および, 自然 免疫を制御する新規医薬品の創製につながると考えられる. 自然免疫関連疾患 に対する治療薬はこれまでに精力的に研究が進められており, 現在開発中のエ リトランは TLR4 のアゴニストとして働き, 敗血症の治療薬として期待される (Figure 3). 43)
また, DHMEQ は, Rel A, Rel B, および c-Rel の核への局在の阻害 をし, NF-κB の活性化を妨げるため抗がん剤や抗炎症薬として期待される. 44) しかし, まだ自然免疫を制御する薬は上市されていないため, 更なる研究が求 められている. Figure 3. 現在開発中の自然免疫制御化合物の化学構造 自然免疫制御化合物の探索に哺乳類 (細胞) を用いた場合獲得免疫の存在や 複雑なシグナル伝達系などにより化合物の自然免疫系に対する作用のみを検出
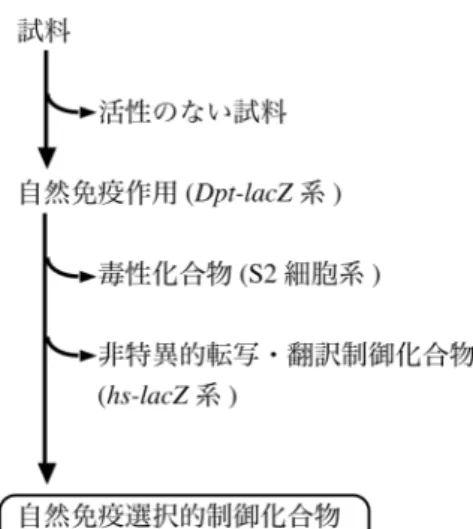
7 セイ系を構築することにより, 天然物から自然免疫制御化合物の探索を行って きた. 45,46) ショウジョウバエは獲得免疫を持たず自然免疫のみで生体防御を行っ ている. そのため, ショウジョウバエを用いることで, 獲得免疫の影響を受けず に自然免疫特異的な作用を検出することが可能であると考えられる. また, シ ョウジョウバエの自然免疫機構は前述のようにヒトの自然免疫機構と高度に保 存されており, ショウジョウバエにおいて自然免疫制御作用を示す化合物はヒ トに対しても同様の効果を示すことが期待される. さらに, ショウジョウバエ は遺伝学的解析が進んでおり, 遺伝子導入個体や欠損個体の入手が容易である ことから, 自然免疫活性の検出およびその作用機作の解析に最適なモデル生物 であると考えられる. 自然免疫制御物質の探索には, IMD 経路で産生されるショウジョウバエの抗 菌ペプチドの一つである diptericin の転写制御領域に, レポーター遺伝子であ る lacZ をつないだ外来遺伝子を導入した遺伝子組換えショウジョウバエ (Dpt-lacZ) を用いている.47) Diptericin はショウジョウバエの自然免疫活性化機 構の一つである IMD 経路の活性化によって産生が誘導される. 48) 自然免疫が 活性化され diptericin の転写が促進されると, lacZ の転写も促進され, レポー タータンパクである β-galactosidase の産生が起こる (Figure 4). したがって, 自 然免疫の活性化は β-galactosidase の産生量を測定することにより検出できる. 試料の自然免疫制御活性はこの β-galactosidase 産生量が増減するかを調べるこ とにより検出できる. すなわち, 自然免疫シグナル経路の活性化と同時に, 試料 を加え β-galactosidase の産生量が増加した場合は自然免疫活性化作用を持つ化 合物として, β-galactosidase の産生量が減少した場合は自然免疫抑制作用を持つ 化合物として考えられる. 自然免疫特異的な作用とともに, 化合物の細胞毒性 と転写・翻訳活性への作用についてもあわせて評価することで,47) 自然免疫特異 的な作用を有する化合物を選択することができる (Figure 5). 現在までに当研究室では, 多くの放線菌および糸状菌抽出物から自然免疫制 御物質の探索を行ってきた. その結果, Streptomyces 属放線菌より celastramycin A が, 49) また Aspergillus 属糸状菌より TP-1 がそれぞれ自然免疫抑制物質と して単離された (Figure 6). 50) これまで単離された化合物は自然免疫の抑制に働 き, 自然免疫を増強する化合物は全く単離されていなかった. 微生物は宿主の
Figure 4. Dpt-lacZ 系における ex vivo での β-galactosidase 活性
9 自然免疫により排除される立場にあるために, 微生物が生き残るには宿主に感 染する際に, 自然免疫を抑制する化合物を産生し自らを守る必要があると考え られる. Figure 6. 当研究室で単離された自然免疫抑制化合物の化学構造 一方, 宿主内の環境を自らの増殖に有利にするために, 微生物は宿主の免疫 を増強する化合物を産生し, 他の菌を排除する可能性がある.したがって, 新規 医薬品及び自然免疫機構のバイオプローブの開発を目指し, 自然免疫増強化合 物 の 探 索 研 究 を 行 っ た . そ の 結 果 , 本 研 究 室 の 菊 地 ら に よ っ て 糸 状 菌 Gonytrichum sp. 抽出物から,自然免疫増強作用を示す新規化合物 gonytolide A (Figure 7) が単離された. 51) この gonytolide A は, 二つの不斉中心を持つクロマ ノン誘導体が二量化した化合物 (ビスクロマノン骨格) である. Gonytolide A は Dpt-lacZ 系において自然免疫を活性化するが, その活性化作用の発現メカニズ ムは不明であった. そこで今回, 第一章で gonytolide A の作用がショウジョウバ エにおいて IMD 経路選択的であることを確認した. さらに第二章では, ヒト 細胞に対する gonytolide A の自然免疫活性を評価し, ヒト自然免疫シグナル経 路である TNF 経路関連因子のリン酸化特異的抗体を用いて自然免疫シグナル 経路における作用点の解析を試みた. さらに第三章では, gonytolide A がどのよ うな遺伝子の発現に影響を及ぼすか網羅的な解析を試みた. Figure 7. Gonytolide A の化学構造
本論
第一章 Gonytolide A のショウジョウバエにおける自然免疫増強作
用の解析
ショウジョウバエは病原微生物の感染に対して, 脂肪体での抗菌ペプチドの産 生によって対抗する. 産生された抗菌ペプチドは, 病原微生物の細胞膜に結合し 膜に穴をあけることで抗菌作用を示す. このショウジョウバエにおける抗菌ペプ チドの産生誘導は, パターン認識タンパク質による病原体の認識とその下流の自 然免疫経路の活性化によって起きる. ショウジョウバエのパターン認識タンパク 質は, 病原体の細胞壁成分のある共通の分子パターンを認識し, その結果, 分子 パターンに応じた自然免疫経路を活性化する. グラム陽性菌の感染時には, PGRP-SA が働く. PGRP-SA は体液中でグラム陽 性菌 の Lys 型ペプチドグリカンを認識し, Spätzle が活性化される. 活性化 Spätzle は Toll 受容体を介して Toll 経路を活性化し, 抗菌ペプチド Drosomycin の産生を誘導する. 一方, グラム陰性菌が有する DAP 型ペプチドグリカンは, PGRP-LC 及び PGRP-LE に認識される. PGRP-LC, PGRP-LE はいずれも IMD 経路を活性化し, 抗菌ペプチド Diptericin の産生を引き起こす. 本章では, gonytolide A の自然免疫増強作用を詳細に検討するために, IMD 経 路のリガンドである DAP 型ペプチドグリカン刺激下での gonytolide A の自然 免疫増強作用の濃度依存性及び, 時間依存性を調査した. 続いて, gonytolide A の 作 用 が IMD 経 路 に 依 存 的 か 調 べ る た め に , リ ガ ン ド を 加 え な い 場 合 の β-galactosidase 活性を測定した. さらに, 熱刺激依存的に β-galactosidase を産生 するショウジョウバエを用いて, gonytolide A の作用が自然免疫特異的であるか 調査した.11
第一節 DAP 型ペプチドグリカンに依存した gonytolide A の自然
免疫増強
作用の評価
ショウジョウバエの自然免疫経路の一つである IMD 経路は, DAP 型ペプチ ドグリカンにより活性化され Diptericin を産生する. そこで, diptericin プロモ ーターの活性をレポーター遺伝子 lacZ を用いて測定し, gonytolide A のショウ ジョウバエの自然免疫に対する作用を評価した. DAP 型ペプチドグリカン刺激 時における diptericin プロモーター活性を測定したところ, gonytolide A の処理 により 10 µg/ml で約 3 倍, 100 µg/ml で約 5 倍の活性となった (Figure 8a). 一方, 1 µg/ml 以下では上昇はほとんど認められなかった. また, gonytolide A に よるプロモーター活性の経時変化を調べたところ, DAP 型ペプチドグリカン刺 激 12 時間後から活性の上昇が認められ, 18 時間後で増加率が最大となった (Figure 8b) .続いて, gonytolide A による diptericin プロモーターの活性の増強が DAP 型 ペプチドグリカンに依存しているのか, gonytolide A 単独でも生じるのか調べる ために, gonytolide A 単独処理でのプロモーター活性を測定した (Figure 8c). そ の結果, 無刺激下では gonytolide A を作用させてもプロモーター活性の上昇は ほとんど認められず, 100 µg/ml でわずかに認められるに留まった.
Figure 8. Gonytolide A の diptericin プロモーターに対する作用
a) DAP 型ペプチドグリカン刺激下での diptericin プロモーター活性に対する作用. Gonytolide A, DAP 型ペプチドグリカンを含む培地中で Dpt-lacZ 幼虫を ex vivo 培養し, 12 時間後の β-galactosidase 活性を測定した. 縦軸は gonytolide A 0 µg/ml の値の相対値. 横軸は gonytolide A の濃度. b) Gonytolide A の作用の時間依存性. DMSO あるいは gonytolide A 及び DAP 型ペ プチドグリカンを含む培地中で Dpt-lacZ 幼虫を ex vivo 培養し, 0, 4, 8, 12, 18, 24 時間後の β-galactosidase 活性を測定した. 縦軸は β-galactosidase/protein . 横軸は幼虫の培養時間. c) Gonytolide A 単独処理時の diptericin プロモーター活性に対する作用. Gonytolide A を含む培地 中で Dpt-lacZ 幼虫を ex vivo 培養し, 12 時間後の β-galactosidase 活性を測定した. 縦軸は
13
第二節 ショウジョウバエにおける自然免疫増強作用の評価
DAP 型ペプチドグリカン刺激下で gonytolide A は濃度依存的に diptericin のプロモーター活性を上げることが明らかとなったが, gonytolide A が特定のシ グナルに依存せずに転写・翻訳量を増加させる可能性が考えられた. そこで, 熱 刺激に依存し β-galactosidase を産生する hs-GAL4/UAS-lacZ 系統のショウジョ ウバエを用いて gonytolide A の転写・翻訳に対する作用を調べた (Figure 9). そ の結果, hs-lacZ 系では gonytolide A による β-galactosidase の産生の増加は
diptericin プロモーターで増強作用が強く認められた 100 µg/ml においても認 められなかった. 続いて, gonytolide A のショウジョウバエ S2 細胞に対する細胞毒性を評価し たところ, 作用のある 10 µg/ml まで gonytolide A は細胞毒性を示さなかった (Figure 10).
Figure 9. Gonytolide A の熱刺激プロモーターに対する作用 Gonytolide A を含む培地に hs-GAL4/UAS-lacZ 系統の幼虫を加え, 37˚C, 30 分間熱処理した後に, 25˚C で 18 時間培養し, 18 時間後の β-galactosidase 活性を測定した. 縦軸は gonytolide A 0 µg/ml の値の相対値. 横軸は gonytolide A の濃度. Figure 10. Gonytolide A のショウジョウバエ S2 細胞の生存率に対する作用 S2 細胞を gonytolide A を含む培地中で培養し, 24 時間後に細胞数測定試薬を加え, 25̊C で 4 時間培養した. 測定試薬を加えた直後と 4 時間後の 450 nm の吸光度を測定した. 縦軸は gonytolide A 0 µg/ml の値の相対値. 横軸は gonytolide A の濃度.
15
第三節 抗菌ペプチドの発現に対する作用
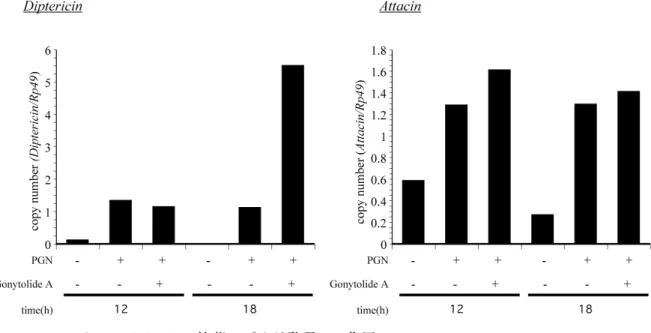
前節で, gonytolide A は DAP 型ペプチドグリカン刺激下で diptericin プロモ ーターの活性を増加させた. そこで, 実際に Diptericin の産生量が gonytolide A で増加するか否かを調べるために, IMD 経路の活性化によって発現する二つの 抗菌ペプチド diptericin, attacin の mRNA 量を測定した (Fig. 11). その結果, DAP 型ペプチドグリカン刺激 12 時間後では gonytolide A の有無で diptericin の発現に変化が認められなかったが, 18 時間後において gonytolide A 処理で
diptericin の発現が約 5 倍増加した. 一方, attacin の発現量は, DAP 型ペプチ
ドグリカン刺激 12, 18 時間後のどちらにおいても gonytolide A の処理の有無で 変化しなかった.
Figure 11. Gonytolide A の抗菌ペプチド発現への作用
DMSO あるいは gontolideA 10 µg/ml , 及び DAP 型ペプチドグリカン 100 ng/ml を含む培地中 で, Dpt-lacZ 幼虫を 12, 18 時間 ex vivo 培養したときの diptericin, 及び attacin の RNA 量を 測定した. 縦軸は copy number (1/Rp49) .
17
第四節 考察
当研究室で行われたショウジョウバエを用いた自然免疫制御化合物のスクリ ーニングにより, 新規化合物 gonytolide A が自然免疫増強化合物として見いだ された. Gonytolide A はスクリーニングの評価系の一つである Dpt-lacZ 系にお いて β-galactosidase 活性を顕著に増加させる. 本章では, gonytolide A の示す β-galactosidase 活性が自然免疫選択的であることを確かめるために, gonytolide A の diptericin プロモーターの活性に対する作用を検討した. DAP 型ペプチドグリカン刺激下では, gonytolide A は 3 µg/ml 以上の時濃度 依存的に diptericin プロモーター活性を上昇した. また, DAP 型ペプチドグリ カン刺激後の diptericin プロモーター活性を経時的に調査したところ, 刺激後 12 時間から gonytolide A によるプロモーター活性の上昇が認められ, 18 時間 後に最大となった. 続いて, gonytolide A による diptericin プロモーターの活性 の増強が DAP 型ペプチドグリカンに依存して生じるのかを調べるために, gonytolide A 単独処理時の diptericin プロモーター活性を測定した. その結果, gonytolide A は単独では diptericin プロモーターの活性化を引き起こさないこ とが明らかとなった. したがって, gonytolide A による diptericin プロモーター の活性化は DAP 型ペプチドグリカンに依存して生じることが示された. 続いて, gonytolide A が実際に Diptericin の産生量を増加させるのかを確かめ るために抗菌ペプチド diptericin 及び attacin の RNA 量を測定した. その結 果 , gonytolide A は DAP 型 ペ プ チ ド グ リ カ ン 刺 激 18 時 間 後 に お い て ,diptericin の RNA 量を増加させた. したがって, gonytolide A による diptericin
プロモーターの活性化は転写レベルで生じることが示された. Gonytolide A に よる diptericin の発現量の増加が DAP 型ペプチドグリカン刺激の 12 時間後 では認められないにもかかわらず, 18 時間後で見られたことから, gonytolide A は diptericin の発現量自体を増加させるのではなく, mRNA の分解を妨げるこ とで対照に比べ発現量の低下が抑制されたと考えた. 一方, gonytolide A は Toll 経路及び IMD 経路の活性化で産生される attacin の発現には影響を与えなか った. attacin と diptericin の発現パターンを比較すると attacin の発現時間が 早いことから, attacin と diptericin の発現を制御する因子やメカニズムが異な る可能性が考えられ, そのメカニズムの違いが gonytolide A による作用の違い を生じたと考えられた.
第二章 Gonytolide A のヒト臍帯静脈血管内皮細胞 (HUVEC) に
おける自然免疫増強作用の解析
前章で gonytolide A はショウジョウバエの自然免疫応答シグナル伝達経路で ある IMD 経路において, diptericin プロモーターの活性を増強することが明ら かとなった. Gonytolide A のヒトへの適用を目的として, ショウジョウバエの IMD 経路と相同性のあるヒトの TNF 経路への gonytolide A の作用について 検討した. 病原微生物が上皮組織を通過すると, 食細胞によって TNF-α などのサイト カインの産生が起こる. 産生された TNF-α は細胞膜表面に存在する TNFR に 結合し, 細胞内シグナル伝達によって NF-κB の活性化を誘導する. このシグナ ル伝達に関与するタンパク質が TNF 経路と IMD 経路では保存されているこ とから, IMD 経路に作用する化合物 gonytolide A は TNF 経路にも作用するこ とが期待される. そこで本章では, TNF-α の標的細胞の一つである血管内皮細 胞用いてヒトの自然免疫に対する作用を解析した. 第一節で gonytolide A のサ イトカインの産生に対する作用を評価し, 第二節では gonytolide A の TNF 経 路に対する作用を検討した.19
第一節 各種刺激下での gonytolide A のサイトカイン産生に
対する作用の解析
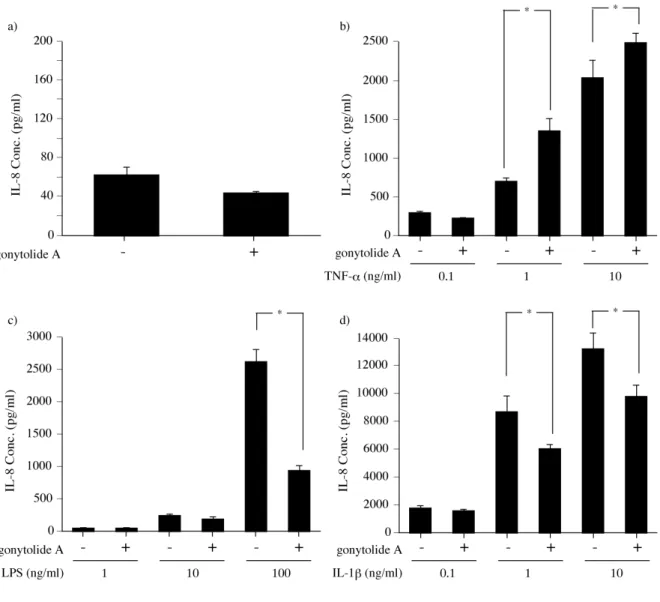
第一項 TNF-α 刺激によるサイトカイン産生に対する作用 HUVEC では TNF-α の刺激により TNF 経路が活性化され, IL-8 などの炎 症性サイトカインを産生する. IL-8 は好中球の活性化と遊走を惹起するサイト カインであり, 異物の侵入に伴う初期の炎症反応に深く関係する. そこで, ヒト における gonytolide A の自然免疫活性を評価するために, TNF-α 刺激下での IL-8 産生量を測定した. Gonytolide A で 1.5 時間前処理した HUVEC を TNF-α (1 ng/ml) で刺激し, 16 時間後の培養上清中の IL-8 を ELISA で測定した. TNF-α 刺激による HUVEC での IL-8 の産生量は 680 pg/ml であった (Figure 12a). Gonytolide A を 1 µg/ml 以下で前処理したときの IL-8 はおよそ 650 ng/ml であり化合物無 処理時と同程度であったが, 3 µg/ml, 10 µg/ml で処理した場合, それぞれ 970 pg/ml, 1200 pg/ml に増加した. また, gonytolide A の HUVEC への細胞毒性を検 討した結果, 10 µg/ml 以下では毒性を示さなかった (Figure 12a). このことから, gonytolide A は TNF-α 刺激による炎症性サイトカインの産生を増加させたと 考えられる. 従って, gonytolide A はショウジョウバエだけでなくヒト細胞に対 しても自然免疫経路に作用することが示唆された. 続いて, gonytolide A の IL-8 産生に対する作用を経時的に調べたところ, TNF-α 刺激 16 時間後より gonytolide A 処理群での IL-8 の産生増加が認めら れた (Figure 12b).Figure 12. Gonytolide A の HUVEC における自然免疫活性
a) Gonytolide A の HUVEC に対する作用. Gonytolide A の自然免疫活性 (■), 細胞生存率 (●) に対する作用を測定した. 自然免疫活性: HUVEC を 37˚C で 12 時間培養した後, gonytolide A で 1.5 時間処理し, TNF-α で刺激した. 刺激後 16 時間の培養上清を回収し, ELISA にて IL-8 産生量を測定した. 縦軸は IL-8 産生量. 横軸は gonytolide A の濃度. 細胞毒性: HUVEC を gonytolide A を加えた培地中で 24 時間培養し, 細胞数測定試薬を加え, 4 時間後の 450 nm の吸光度を測定した. 測定試薬を加えた直後と 4 時間後の 450 nm の吸光度を測定した. 縦軸 は gonytolide A 0 µg/ml の時の値に対する相対値. 横軸は gonytolide A の濃度. * p < 0.05 b) Gonytolide A の作用の時間依存性. HUVEC を gonytolide A で 1.5 時間処理し, 0, 12, 16, 20, 24 時間後の培養上清を回収し, IL-8 産生量を測定した. 縦軸は IL-8 産生量. 横軸は TNF-α 刺
21
第二項 LPS および IL-1β 刺激によるサイトカイン産生に対する作用
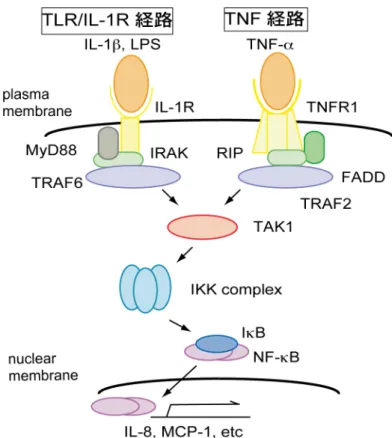
Gonytolide A のシグナル経路への特異性を検討するために, リガンドとして IL-1β, LPS を用いて, IL-8 産生量の調査をした. IL-1β, LPS はそれぞれ IL-1R, TLR4 のリガンドであり, MyD88 依存的なシグナル伝達によって NF-κB を活 性化する (Figure 13). ショウジョウバエでは MyD88 依存的シグナル伝達経路 として Toll 経路があり, IL-1β, LPS によって活性化される IL-1R/TLR 経路と Toll 経路は高く保存されている. TNF 経路と IL-1R/TLR 経路はともに NF-κB を活性化し, サイトカインなどの産生を誘導するが, 上流でのシグナル伝達に 関わる因子が異なっている. そこで, gonytolide A が TNF-α だけでなく IL-1β, LPS での刺激に対しても作用するか調べるために, HUVEC を gonytolide A で 1.5 時間前処理し, IL-1β および LPS での刺激後 16 時間での IL-8 の産生量 を ELISA で検出した (Figure 14). IL-8 の産生量は TNF-α 刺激時では TNF-α の濃度 1 ng/ml, 10 ng/ml で, gonytolide A 処理によって増加した. 無刺激時では増加は認められなかった. 一 方, LPS 刺激では LPS が 1 ng/ml, 10 ng/ml では IL-8 量に変化が見られなかっ たが, 100 ng/ml で 36 % に減少した. また, IL-1β 刺激では, 0.1 ng/ml では変化 がなかったが, 1, 10 ng/ml ではそれぞれ 70, 74% に減少した.
Figure 13. TLR/IL-1R 経路と TNF 経路
TLR/IL-1 経路, TNF 経路はともに自然免疫や炎症に重要な役割を果たしており, TAK1 で活性
23
Figure 14. 各種刺激による IL-8 産生への作用
DMSO (gonytolide A (-)), または gonytolide A 10 µg/ml (gonytolide A (+)) で HUVEC を 1.5 時間 前処理した後, a) 無刺激下, あるいは各濃度の b) TNF-α , c) LPS , d) IL-1β で刺激し, 16 時間培 養したときの IL-8 産生量を ELISA で測定した. 縦軸は IL-8 産生量. * p<0.05
第三項 Gonytolide A の IL-8 の発現に対する作用
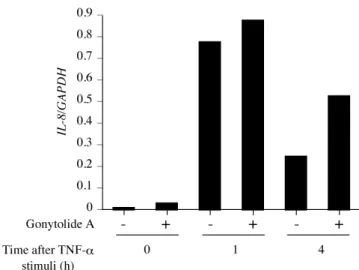
前項で ELISA により, 各種刺激下での gonytolide A による IL-8 産生量の 変化について評価した. Gonytolide A は TNF-α 刺激下では IL-8 の産生を正に 制御した. この変化が実際に転写レベルで生じるのか調べるために, TNF-α 刺 激下で IL-8 の mRNA 量を測定した (Figure 15). その結果, TNF-α 刺激後 4 時間の時, gonytolide A 処理で IL-8 の発現量が約 2 倍増加した.
25
Figure 15. TNF-α 刺激下の IL-8 発現
DMSO (gonytolide A (-)), または gonytolide A 10 µg/ml (gonytolide A (+)) で HUVEC を 1.5 時間 前処理した後, TNF-α 1 ng/ml による刺激 0, 1, 4 時間後の IL-8 の RNA 量を測定した. 縦軸は copy number (1/GAPDH).
第四項 TNF-α 刺激下での gonytolide A による免疫関連遺伝子の発現
前項で TNF-α 刺激下で, gonytolide A は IL-8 の発現を増加させることが明 らかとなった. TNF-α は TNF 受容体に結合し, TNF 経路を活性化することで, サイトカインや COX-2, ICAM1 といった炎症関連因子の発現を誘導する. そこ で, gonytolide A が TNF-α 刺激下で IL6, COX-2, ICAM1 の発現にどのように影 響するかを調査した (Figure 16). RT-PCR により mRNA 量に変化が認められ るかを調べた結果, IL-6 は TNF-α 刺激後 0, 1, 4 時間後のいずれにおいても gonytolide A の処理の有無で発現量に変化が認められなかった. 一方, COX-2 は TNF-α による刺激がない場合でも gonytolide A により発現量が約 2 倍増加し, 刺激後 1 時間の際にも gonytolide A により発現量の増加は見られたが, TNF-α 刺激 4 時間後ではほとんど発現していなかった. ICAM1 は TNF-α 無刺激時及 び刺激 1 時間後の時には gonytolide A による発現増加は起こらなかったが, 刺激 4 時間後の時には gonytolide A 処理で約 2 倍発現が増加した.
27
Figure 16. TNF-α 刺激時の IL-6, COX-2, ICAM1 の発現への作用
HUVEC を 12 時間培養し, DMSO または gonytolide A 10 µg/ml で 1.5 時間処理した後, TNF-α 1 ng/ml で刺激し 0, 1, 4 時間後の IL-6, COX-2, 及び ICAM1 の RNA 量を測定した. 縦 軸は copy number (1/GAPDH). * p<0.05
第二節 TNF 経路における gonytolide A の作用
前節で gonytolide A はショウジョウバエだけでなくヒト細胞においても自然 免疫経路に対して作用を示すことが明らかとなった. TNF 経路では TNF-α 刺 激によって IKK がリン酸化され活性化し, 活性化した IKK は IκB-α の Ser32, Ser36 をリン酸化する (Figure 17). リン酸化された IκB-α はポリユビキ チン化されプロテアソームによって分解される. IκB-α は NF-κB を細胞質にと どめる作用をしているため, IκB-α が分解されることにより NF-κB は核内へ移 行する. その結果, 転写因子である NF-κB によって各種炎症性サイトカインが 産生される. そこで, gonytolide A の TNF 経路に対する作用を検討するため, HUVEC を用いてシグナル伝達因子の活性化, 及び NF-κB に対する作用を調 査した.
29
Figure 17. TNF-α 刺激による IKK, IkB-α のリン酸化
TNF-α 刺激により TAK1 などの MAP3 キナーゼが活性化すると, IKK のリン酸化が誘導され る. リン酸化により活性化した IKK 複合体は IκB-α をリン酸化する. リン酸化された IκB-α はユビキチン化を受けプロテアソームにより分解する.その結果, NF-κB (p50, p65) 複合体は核 内に移行し各種サイトカインの産生を誘導する
第一項 TNF 経路関連シグナル伝達因子のリン酸化状態への作用
HUVEC では, TNF-α による TNF 経路の活性化を介して, 各種サイトカイ ンが産生される. そこで, gonytolide A によって TNF 経路の IKK, IκB-α のリ ン酸化分解に変化がないかをウェスタンブロット法によって調査した (Figure 18). リン酸化 IKK の量は TNF-α での刺激を行わないとき, gonytolide A 処理 によって増加したが, TNF-α 刺激 5 分後では gonytolide A 処理により減少し, 15, 30, 60 分後でも同様であった. 続いて IκB-α のリン酸化状態を調べたとこ ろ, TNF-α で刺激を行わない場合, gonytolide A で処理しても変化は認められな かった. TNF-α で刺激した時のリン酸化 IκB-α の量は, IKK の場合と同様に gonytolide A 処理によっていずれの時間も少なくなった. 一方, 細胞中での全 IκB-α 量は減少していることから IκB-α のユビキチン化とプロテアソームで の分解は起きていると考えられた. そこで, プロテアソーム阻害剤である MG132 を用いて IκB-α のリン酸化体 の検出を試みた (Figure 19). その結果, MG132 処理群では, gonytolide A 単独処 理時に IκB-α の増加が認められた. また, TNF-α 刺激下では, MG132 処理時に gonytolide A を作用させると IKK と同様にリン酸化 IκB-α の量が減少した.
31
Figure 18. IKK, IκB-α のリン酸化状態
DMSO (-), gonytolide A 10 µg/ml (+) で HUVEC を 1.5 時間前処理した後, TNF-α 1 ng/ml で刺激 し, 0, 5, 15, 30, 60 分後の IKK, phospho-IKK (P-IKK), IκB-α (IκB), phospho-IκB-α (P-IκB), 及び actin を検出した.
Figure 19. MG132 処理時の IκB-α のリン酸化状態
MG132 10 µg/ml で 1.5 時間前処理した HUVEC を DMSO (-), gonytolide A 10 µg/ml (+) で 1.5 時間処理した後, TNF-α 1 ng/ml で刺激し, 0, 15 分後の IκB-α (IκB), phospho-IκB-α (P-IκB), 及 び actin を検出した.
第二項 核抽出物の NF-κB 活性に対する作用の評価
Gonytolide A は単独処理で, IKK, IκB-α をリン酸化したため, TNF 経路の転写 因子である NF-κB の活性が gonytolide A 処理により活性化されるかどうかを 検討した. HUVEC を 10 µg/ml の gonytolide A で 1.5 時間処理した後, TNF-α で刺激 0, 1, 4 時間後の細胞を抗 NF-κB p65 抗体を用いて免疫染色した (Figure 20a). その結果, NF-κB は無刺激時には細胞質に存在し, TNF-α 刺激によって核へと 移行したが, いずれの時間においても, gonytolide A 処理群と非処理群で変化は 認められなかった. 細胞を細胞質画分と核画分に分画したところ, 免疫染色の ときと同様に NF-κB は TNF-α 刺激により細胞質画分から核画分へと移行し たが, gonytolide A 処理の有無で変化は見られなかった (Figure 20b). 続いて, 得 られた核画分中の NF-κB 活性を測定した結果, TNF-α 処理 0, 15 分後いずれ においても NF-κB の活性は gonytolide A の処理の有無でほとんど差がなかっ た (Figure 20c).
33
Figure 20. Gonytolide A の NF-κB に対する作用
a) 抗 NF-κB p65 抗体を用いた免疫染色. DMSO (-), gonytolide A 10 µg/ml (+) で HUVEC を 前処理し, TNF-α 1 ng/ml で 0, 1, 4 時間刺激した後, 細胞を抗 NF-κB p65 抗体を用いて免疫 染色をした. 緑色のシグナルは NF-κB p65 を, 青色のシグナルは細胞核を示す. b) NF-κB の 細胞内局在への作用. DMSO (-), gonytolide A 10 µg/ml (+) で HUVEC を 1.5 時間処理した後 TNF-α 1 ng/ml で 0, 15 分刺激した後, 核画分 (Nuc.), 及び細胞質画分 (Cyto.) に分画し, NF-κB (p65) を検出した. c) NF-κB 転写活性に対する作用. DMSO (-), gonytolide A 10 µg/ml (+) で HUVEC を前処理し, TNF-α 1 ng/ml で 0, 15 分刺激した後, 核画分を抽出し, NF-κB 転写活性を測定した. 縦軸は TNF-α 刺激後 15 分後の NF-κB 転写活性を 1 とした時の NF-κB 転写活性.
第三項 MAP kinase に対する作用
TNF-α 刺激では NF-κB への経路以外にも MAP kinase ファミリーの因子も 活性化されることが知られている (Figure 21). これらの因子の中から JNK, phospho-JNK, c-Jun, phospho-c-Jun に対する gonytolide A の作用について特異的 抗体を用いて解析した. その結果, gonytolide A 単独処理時にリン酸化 c-Jun の 量が増加した (Figure 22).
35
Figure 22. JNK, c-Jun に対する作用
a) JNK, c-Jun のリン酸化状態. DMSO (-), gonytolide A 10 µg/ml (+) で HUVEC を 1.5 時間処理 し, TNF-α 刺激 0.5, 1, 2, 4 時間後の JNK, phospho-JNK (P-JNK), c-Jun (cJun), phospho-c-Jun (P-cJun), 及び actin を検出した.
37
第三節 考察
本章では, gonytolide A のヒトの自然免疫に対する作用を検討した. まず, 第 一節では, TNF-α 刺激により誘導される IL-8 の産生量を指標に, gonytolide A の TNF 経路に対する作用を調べた. その結果, gonytolide A は HUVEC におい て 1 µg/ml 以上で濃度依存的に, TNF-α 刺激後 16 時間から IL-8 の産生量を 増加させた. さらに gonytolide A による IL-8 産生の増加が TNF-α の刺激に依 存しているかを調査するために, リガンドとして IL-1β, および LPS 用いたと きの IL-8 産生量を測定した. その結果, TNF-α では gonytolide A による IL-8 の産 生 増加 が認め ら れる の に対し , LPS あるいは IL-1β で刺激した場合, gonytolide A の処理で IL-8 産生量が減少した. したがって, gonytolide A は TNF 経路の下流のシグナル伝達は正に制御するが, TLR 経路及び IL-1R 経路 に対しては抑制的に働くことが示唆された. リガンドの違いによる IL-8 産生 量の変化がどのようなメカニズムで生じるかは現在調査中であるが, gonytolide A により産生誘導される遺伝子の中に TNF-α 刺激下では IL-8 の産生を正に 制御するが, IL-1β, および LPS 刺激下では IL-8 の産生を負に制御する遺伝子 が 含 ま れ る 可 能 性 が 考 え ら れ た . IL-8 量 は TNF-α 刺 激 1 時 間 後 で は gonytolide A の有無で変化は認められなかったが, 4 時間後では gonytolide A の処理で発現量が 2 倍に増加したことから, gonytolide A は TNF 経路におい て IL-8 のダウンレギュレーションに関わる因子を抑制し発現量を増加させた と考えた. Gonytolide A が TNF 経路によって発現誘導される他の炎症関連因子 に対してどのように作用するかを調べるために, IL-6, COX-2, ICAM1 の発現量 を調べた. IL-6 は gonytolide A による発現誘導が見られなかったが, COX-2, 及 び ICAM1 は そ れ ぞ れ TNF-α 刺 激 0, 1 時 間 後 あ る い は 4 時 間 後 で , gonytolide A により発現が増加した.続いて, gonytolide A の TNF 経路に対する作用を解析するために, リン酸化 特異的抗体を用いて gonytolide A の TNF 経路のシグナル伝達因子への作用を 検討した. その結果, gonytolide A は単独で IKK, IkB-α を活性化することが明 らかとなった. したがって, gonytolide A の処理で IKK のリン酸化状態が変 化することから, IKK よりも上流の因子に対して作用することが示唆された. 一方, TNF-α 刺激下では IKK, および IκB-α のリン酸化量が減少していた. ま た, MAP kinase 関連経路に関しては, gonytolide A 処理で c-Jun がリン酸化され
た. 続いて TNF 経路の最下流に存在する NF-κB に対する作用を調べたとこ ろ, gonytolide A で処理しても NF-κB の細胞内局在及び転写活性は変化しな かった.
Gonytolide A は単独では自然免疫関連因子である IKK, IkB-α, JNK, c-Jun を リン 酸化 する が , 単独処理時には IL-8 の発現増加は認められない. 一方, TNF-α 刺激下では gonytolide A の処理で IL-8 の産生が増加するが, IKK 及び IkB-α のリン酸化量は減少する. さらに, gonytolide A は NF-κB に対して活性 化作用を示さない. 以上の結果から, gonytolide A 単独処理で活性化した因子が, TNF-α 刺激下で TNF 経路を負に制御することが示唆された. Gonytolide A の処理がどの経路のシグナル伝達に関連しているのか確かめる ために, 次章では gonytolide A により発現が変化する遺伝子の網羅的な解析を 試みた.
39
第三章 Gonytolide A による遺伝子発現変動の網羅的解析
前章で, gonytolide A は単独で TNF 経路のシグナル伝達因子である IKK, IκB-α をリン酸化すること, また, TNF-α の刺激に応答し IL-8 の産生を増加す るこ と を 明 ら かと し た. しかし, IKK, および IκB-α のリン酸化体の量は TNF-α 刺激下では gonytolide A 処理で減少した. 通常, TNF 経路が活性化され ると, IKK や IκB-α のリン酸化が起こり, 続いて NF-κB が活性化され核内へ 移行する. しかし, NF-κB の活性について調査したところ, gonytolide A の処理 でも NF-κB 活性の上昇は見られなかったため, gonytolide A の作用は TNF 経 路に直接的に作用して生じるものではないことが示唆された. Gonytolide A がどの経路に作用し IL-8 の産生を増加するかを調べるために, 本 章 で は マ イ ク ロ ア レ イ に よ る 遺 伝 子 発 現 変 動 の 網 羅 的 解 析 を 試 み た . Gonytolide A 単独で発現が増加した遺伝子について, 実際に RNA 量が増加す るか RT-PCR で確認した. 続いて, gonytolide A により発現が増加した遺伝子 をノックダウンしたときの IL-8 産生量を評価し, gonytolide A の作用との関連 を調査した. また, 各遺伝子の発現に対する相互作用を RT-PCR で検討した. さらに, TNF-α 刺激で誘導される遺伝子に対する gonytolide A の影響を調べた.第一節 Gonytolide A 単独処理で発現が変化する遺伝子
第一項 Gonytolide A 単独処理時の遺伝子発現変動 第二章での解析から, gonytolide A による IL8 の発現増強は単純に TNF 経路 の活性化による NF-κB の活性の上昇によるものではなく, 他の因子の制御を 受けている可能性が考えられた. そこで, gonytolide A 単独で発現が変化する遺 伝子を調べるため, HUVEC を gonytolide A で処理したときの遺伝子発現を網羅 的に解析した.HUVEC を DMSO あるいは 10 µg/ml の gonytolide A で 1.5 時間処理した ときの遺伝子の発現量をマイクロアレイで解析し, gonytolide A の処理で発現量 が 4 倍以上増加あるいは 4 分の 1 以下に減少する遺伝子を抽出した (Figure 23). その結果, 増加する遺伝子は 24 個, 減少する遺伝子は 20 個存在した. 発 現が変化した遺伝子について KEGG パスウェイ解析をしたが, 傾向は見られ なかった. Gonytolide A 単独で発現が変化する遺伝子は, 増加する遺伝子のうち増加率 が大きい遺伝子に着目し解析を進めたところ, DDIT3 (DNA damage-inducible transcript 3), DDIT4 (DNA damage -inducible transcript 4), KLF4 (krü ppel-like factor 4 (gut)), NR4A2 (nuclear receptor subfamily 4, group A, member 2), ATF3 (activating transcription factor 3) が含まれていた. パスウェイ解析では該当しなかったが, これらの遺伝子はストレス応答により発現誘導されることが報告されてい る.53)-57) DDIT3, DDIT4, KLF4, NR4A2, ATF3 はそれぞれ, gonytolide A で 21.4, 9.5,
8.7, 4.6, 4.4 倍に発現が増加した.
DDIT3, DDIT4, KLF4, NR4A2, ATF3 の遺伝子産物はいずれも転写因子として 働くため, これらの遺伝子との関連を解析することは gonytolide A により生じ る遺伝子発現を明らかにする手がかりとなることが期待された.
DDIT3 は, 転写因子 CEBPB (CCAAT enhancer-binding protein) と二量体を形 成し, 抗アポトーシスタンパク質である Bcl-2 の発現を抑制し, アポトーシス を誘導する.58)
DDIT4 は, 低酸素血症時に発現誘導される遺伝子として同定され, T 細胞の増殖を担う mTOR (mammalian target of rapamycin) を抑制する役割を持
41
症作用を示す. 61) ATF3 は TLR 経路のネガティブレギュレーターとして働き,
過剰な炎症反応を抑制することが知られている. 62)
Gonytolide A 処理でこれらの遺伝子の発現が増加するか否か再度調べるため に, RT-PCR を行い mRNA 量を測定した (Figure 24). その結果, gonytolide A の処理で, DDIT3, DDIT4, KLF4, NR4A2, ATF3 はそれぞれ 10, 4, 2.5, 1.5, 7 倍 増加し, マイクロアレイの結果と同様 gonytolide A による発現増加が認められ た.
Figure 23. Gonytolide A による遺伝子発現変動の網羅的解析
43
Figure 24. マイクロアレイの結果の確認
HUVEC を 12 時間培養し, DMSO (-), gonytolide A 10 µg/ml (+) で 1.5 時間処理した時の DDIT3,
第二項 遺伝子のノックダウンによる gonytolide A の作用の評価
前項で, gonytolide A が DDIT3, DDIT4, KLF4, NR4A2, ATF3 の発現を増加させ ることを明らかにした. Gonytolide A の作用がこれらの遺伝子を介しているの かを確認するために, siRNA を用いて各遺伝子をノックダウンしたときの IL-8 産生量を調査した. HUVEC に siRNA を導入して 8 時間後に gonytolide A で 1.5 時間処理し, TNF-α あるいは LPS 刺激 16 時間後の IL-8 産生量を測定し た (Figure 25). TNF-α 刺激下では gonytolide A 処理で IL-8 の産生量が増加する が, gonytolide A による IL-8 の産生量の増加に関わる遺伝子をノックダウンし た場合, gonytolide A で処理しても IL-8 産生量が増加しなくなると考えられる. それぞれの遺伝子をノックダウンした時の IL-8 の産生量を調査した結果, い ず れ の 場 合 に お い て も gonytolide A に よ っ て IL-8 産 生 量 は 増 加 し た . Gonytolide A による IL-8 の増加率は, ノックダウンを行わない時 1.28 倍, Negative Control を用いた場合, 1.25 倍であり, DDIT3, DDIT4, KLF4, NR4A2,
ATF3 をノックダウンした際は, それぞれ 1.80, 1.77, 1.62, 1.94, 1.49 倍となり, いずれの遺伝子のノックダウンの場合も TNF-α 刺激下での gonytolide A によ る IL-8 の産生量の増加には影響を与えなかった. この結果から, これらの遺伝 子群は gonytolide A による TNF 経路を介した IL-8 の産生に関与しない可能 性が示唆された. 続いて, LPS 刺激下での gonytolide A による IL-8 の産生に対する各遺伝子 の関連を調べた. その結果, gonytolide A によりノックダウンを行わない場合 0.68 倍, Negative Control を用いた場合, 0.75 倍であり, DDIT4, KLF4, NR4A2 を ノックダウンした場合はそれぞれ 0.76, 0.68, 0.50 倍に IL-8 産生量が減少した が, DDIT3 のノックダウンにより gonytolide A による抑制が減弱した. したが って, LPS 刺激下の IL-8 産生の抑制には, DDIT3 が関わっている可能性が考え られた.
45
Figure 25. siRNA を用いたときの IL-8 産生量
HUVEC に siRNA をトランスフェクトし 8 時間培養した後, DMSO (-), gonytolide A 10 µg/ml (+) で 1.5 時間処理し, a) TNF-α 1 ng/ml あるいは b) LPS 100 ng/ml で 16 時間刺激した時の IL-8 量を ELISA で測定した. siRNA を無添加, あるいは Negative Control (Nega), DDIT3,
第二節 TNF-α 刺激下での gonytolide A による遺伝子発現変動
TNF-α 刺激時の遺伝子発現変動を解析するため, gonytolide A 処理細胞を TNF-α で刺激し, 1, 4 時間後の遺伝子の発現量をマイクロアレイを用いて網羅 的に解析した. 主に, TNF-α の刺激に対する gonytolide A の作用を検討するた めに無刺激時の発現量に対して, TNF-α 刺激で発現が 4 倍以上増加あるいは 4 分の 1 以下に減少する遺伝子を抜き出し, gonytolide A の影響を調べた (Figure 26). 前章の実験で, TNF-α 刺激下で gonytolide A による発現増加が確認されたIL-8, ICAM1 遺伝子の発現は gonytolide A によって増加しており, RT-PCR と一
致する結果が得られた. 全体として, TNF-α 刺激により発現が増加あるいは減少 する遺伝子のうち半数以上に対して, gonytolide A は TNF-α の作用を減弱させ ることが明らかとなった.
パスウェイ解析の結果, TNF-α 刺激 1 時間で発現が増加する遺伝子のうち, 免疫・炎症に関わる IL-6, IL-8, IκB-α (NFKBIA), CCL20, CXCL1, CXCL2, CXCL5, が含まれていたが, IL-6, NFKBIA, CCL20, CXCL2 は gonytolide A 処理で発現が さらに増加し, CXCL5 は減少していた. 一方, TNF-α 刺激 1 時間で抑制される 遺伝子は細胞接着に関わる CD58, cldn18, HLA-DRB4, NEO1, PTPRM が含まれ, いずれも gonytolide A 処理でそれぞれ 4.8, 1.9, 6.4, 8.0, 7.5 倍発現量が増加す ることが分かった.
刺激 4 時間後では, TNF-α 刺激で免疫・炎症に関わる CCL20, CXCL1, CXCL2,
CXCL5, Cx3cl, IL-15, IL-8, LTB, TNFRSF9 の発現が増加した. Gonytolide A 処理後
に 発 現 量 が さ ら に 増 加 し た 遺 伝 子 は CCL20, CXCL1, CXCL2, IL-15, IL-8,
TNFRSF9 でそれぞれ 2.8, 1.3, 1.9, 1.7, 1.8, 1.2 倍になった. LTB 及び cx3cl の発
現は gonytolide A 処理で 0.4, 0.75 倍に減少した. また, 刺激下で発現が抑制さ れる遺伝子は, TGF-β 経路の SMAD6, SMAD7, SMURF1, DCN, id1, ID2, id4, nog であり, SMAD6, SMAD7, DCN の発現が gonytolide A 処理でそれぞれ 3.3, 4.3, 11.8 倍に増加した.
47
Figure 26. TNF-α 刺激 1, 4 時間後の遺伝子発現変動.
無刺激時の発現量の平均値と比較し, TNF-α 刺激 a) 1, b) 4 時間の時, 発現量が 4 倍増加, ある いは 4 分の 1 以下に減少した遺伝子を抜き出し, ヒートマップを作成した.
49
第三節 考察
本章では, gonytolide A の遺伝子発現に対する影響を検討した. まず第一節で は, gonytolide A 単独処理時および, TNF-α 刺激下での遺伝子発現の変化を網羅 的に解析した. 単独処理時には, 24 個の遺伝子が増加し, 20 個は減少した. Gonytolide A 単独処理で発現が大きく増加する遺伝子は, gonytolide A の作用と の関連が期待されるため, DDIT3, DDIT4, KLF4, NR4A2, ATF3 に着目し解析した. これらの遺伝子のうち, NR4A2 は酸化ストレスに, DDIT3, DDIT4, KLF4, ATF3 は 小胞体ストレスに応答し発現が誘導される. 実際に gonytolide A 処理でこれら の遺伝子の発現が増加するのかを確かめるために, RT-PCR により発現量を測 定した. その結果, DDIT3, DDIT4, KLF4, NR4A2, ATF3 はいずれも gontolide A により発現が増加することが確認された. 続いて, これらの遺伝子の発現と gonytolide A による自然免疫活性の関連を調べるために, ノックダウンによる gonytolide A の作用への影響を検討した. TNF-α 刺激下での IL-8 の産生増加に 関わる遺伝子をノックダウンした場合, gonytolide A で処理しても IL-8 産生量 は増加しないことが期待された. しかし,いずれの遺伝子もノックダウンすると gonytolide A による IL-8 の産生増加が確認された. したがって, これらの遺伝 子は gonytolide A による TNF 経路の活性化に関与しないことが示唆された. 一方, LPS 刺激下での gonytolide A の作用とこれらの遺伝子との関連を検討し た結果, DDIT3 のノックダウンで gonytolide A による IL-8 の抑制効果が減弱 した. この結果から, gonytolide A は DDIT3 を介して TLR 経路を制御する可 能性が考えられた. 続いて, TNF-α 刺激で, 発現量が 4 倍以上に増加, あるいは 4 分の 1 以下 に減少する遺伝子に対する gonytolide A の影響を検討した. Gonytolide A は TNF-α 刺激で発現が増加あるいは減少する半数以上の遺伝子に対し, その発現 の変化を抑制しているが, 対象となる遺伝子に傾向が見られなかった. TNF-α 刺激下での gonytolide A の作用が DDIT3, DDIT4, KLF4, NR4A2, ATF3 の発現 増加と関連があるのか, また, これらの発現を誘導するストレス応答と関連が あるのかを検討し, 今後 gonytolide A の作用機構を明らかにしていきたい.結語
自然免疫は病原微生物の感染に対して最前線で働く重要な生体防御機構で ある. かつて自然免疫は非特異的に働く原始的な免疫機構と見なされ, ほとん ど注目されなかった. しかし, 自然免疫が病原微生物の構成成分を認識し特異 的に働く高度に制御されたシステムであること, また, 自然免疫が獲得免疫の 活性化に必須であることが示され, 自然免疫は原始的な免疫機構という認識か ら, 感染防御の中心の役割を果たす防御機構であるという認識へと変わった. 一方, 自然免疫の分子機構はまだ十分解明されておらず, 当研究室では自然免 疫制御機構の詳細を明らかにするために, 自然免疫を制御する化合物を天然資 源に求めショウジョウバエを用いたスクリーニング系により探索を行った. そ の結果, 新規化合物 gonytolide A が自然免疫増強作用を有することが明らかと なった. しかし, gonytolide A の自然免疫増強への作用機構は不明であった. そ こで, 本研究では gonytolide A の詳細なメカニズムの解明を目的とした. まず第一章ではショウジョウバエに対する作用を検討した. Gonytolide A は ショウジョウバエ IMD 経路において, DAP 型ペプチドグリカン刺激に依存し,diptericin プロモーターの活性を増強した. また, gonytolide A は diptericin の
mRNA 量を増加することが明らかとなった. しかし, 同様に IMD 経路の制御 を受ける attacin の mRNA 量には変化を与えなかった. Gonytolide A による
diptericin の発現は DAP 型ペプチドグリカン刺激の 18 時間後に増加しており, diptericin プロモーター活性の上昇も 12 時間後から生じている. この結果から,
gonytolide A は IMD 経路のシグナル伝達因子に直接働くのではなく, IMD 経 路のネガティブレギュレーターとして働く因子を抑制する可能性が示唆された. ショウジョウバエとヒトの自然免疫機構は相同性が高いことが知られている ため, 続いて第二章では, HUVEC を用いてヒト TNF 経路に対する gonytolide A の作用を評価した. まず, TNF 経路の活性化で産生される IL-8 の産生量を 測定したところ, TNF-α 刺激下で gonytolide A は IL-8 の産生を有意に増加さ せた. IL-8 の発現量は, TNF-α 刺激 1 時間後では gonytolide A の有無で差が認 められないが, 4 時間後では gonytolide A 処理で発現が約 2 倍増加した. この 結果から, gonytolide A は TNF-α 刺激後の IL-8 発現機構に対しては影響を与