短
報
日本哺乳類学会熊本県におけるクロホオヒゲコウモリMyotis pruinosusの生息初記録と
外部形態・頭骨およびミトコンドリアCyt b遺伝子の変異について
船越 公威
1,坂田 拓司
2,河合久仁子
3,荒井 秋晴
4 1鹿児島国際大学国際文化学部生物学研究室 2熊本市立千原台高等学校 3北海道大学北方生物圏フィールド科学センター 4九州歯科大学総合教育学分野 摘 要 クロホオヒゲコウモリ Myotis pruinosus の九州におけ る生息分布は,これまで宮崎県綾町照葉樹林における捕 獲記録だけであった.今回,熊本県でも生息が確認された ので報告する.確認場所は熊本県東部の山都町内大臣渓 谷にある隧道トンネルの天井の窪みで 2007 年 9 月 30 日, 2008 年 7 月 27 日および 8 月 24 日に成獣雄各 1 頭が確認 された.また,2011 年 11 月 27 日に同トンネルで成獣雌 1 頭が確認された.背面の体毛は灰黒褐色または黒褐色 でクロホオヒゲコウモリの特徴である差し毛に銀色の光 沢がない個体もみられた.しかし,側膜がモモジロコウ モリ M. macrodactylus と違って外足指の付け根に着いて いること,九州ではヒメホオヒゲコウモリ M. ikonnikovi が分布せず尾膜の血管走行が曲線型でなかったことを考 慮して,クロホオヒゲコウモリと判定した.また,熊本 県産 2 個体のミトコンドリア DNA(cytochrome b 遺伝子 1140 bp)を解析した結果,クロホオヒゲコウモリである ことが支持されたが,種内の遺伝的変異が比較的大きく 地理的変異があることが認められた.前腕長や下腿長は 九州産の方が本州・四国産よりも大きかった.頭骨の形 状についても,九州産では本州・四国産のものに比べて 頭骨基底全長が短く,眼窩間幅や脳函幅が広かった.頭 骨計測 8 項目を基に主成分分析を行った結果,九州産は 本州・四国産との間で明瞭に分離された. は じ め に クロホオヒゲコウモリ Myotis pruinosus は落葉広葉樹 林や照葉樹林に生息する種として本州や四国に分布して いるが,捕獲記録は点在しており,分布域は分断化され ていると考えられている.九州における生息記録は,こ れまで宮崎県綾町の照葉樹林に限られていた(前田 2001;Kawai 2009b).今回,熊本県において新たに生息 が確認されたので報告する.また,本種における九州産 個体と本州・四国産個体との違いを,宮崎県産個体も加 えて,外部形態,頭骨形状および DNA 分析によって考 察した. 方 法 調査地は熊本県上益城郡山都町内大臣渓谷の隧道トン ネルで,2007 ~ 2011 年にかけて定期的な捕虫網による 捕獲調査を行った(Fig. 1).また,宮崎県東諸県郡綾町 川中の照葉樹林内では,2003 年 9 月 1 ~ 3 日,2006 年 8 月 16 ~ 17 日および 2007 年 10 月 5 ~ 7 日にカスミ網によ る調査を行った. 捕獲した個体については,齢・性や繁殖状態を調べ, 電子体重計(ハンディミニ 1476,タニタ,東京,最小目 盛 0.1 g)で体重を測定し,ノギス(KSM-15,中村製作 所,東京,最小目盛 0.05 mm)で前腕長,頭胴長,尾長, 下腿長,後足長,耳介長および耳珠長を測定した後,前 腕部に標識用リングを着けて捕獲場所で放獣した.また, 熊本県産 2 個体(B, C: Table 1)から尾膜の組織片を採取 して DNA 分析用に,熊本県産 2 個体(B, D: Table 1)と 宮崎県産 2 個体(G, I: Table 1)は頭骨形態の測定用に供 された.また,齢については,前肢の指骨関節の化骨の 程度や腹部の体毛,雄ではペニスや精巣のサイズ,雌で は乳頭や乳腺の発達程度をチェックして,幼獣,亜成獣 および成獣の区分を行った.幼獣は指骨関節部位に軟骨が残っていることや腹部の体毛が薄く灰黒色であるこ と,亜成獣雄は精巣が未発達であること,亜成獣雌では 乳腺や乳頭がみられないこと,成獣雄は精巣が発達し(秋 季には長径 5 mm 以上),成獣雌では乳腺の発達や乳頭の 肥大がみられることで識別した. 得られた外部形態と頭骨形態の測定値は,九州外の他 地域産の同種と比較検討した.外部形態の測定値の比較 では,岩手・愛媛県産(Yoshiyuki 1989),広島県産(上 野ほか 2002)および栃木県産(安井・上條 1999)を合わ せた平均値と比較した.外部測定値の有意差検定では, Mann-Whitneyの U-test を採用した. 頭骨形態の測定値については,デジタルノギス(CD-10CX,ミツトヨ,川崎,最小目盛 0.01 mm)を使用し て,頭骨基底全長(前顎骨前端―後頭顆後縁:CBL),頬 骨弓幅(左右頬骨弓間の最大幅:ZW),眼窩間幅(左右 眼窩間部の最狭部幅:IOW),脳函幅(脳函の最大幅: BCW),脳函高(脳函の最大高:BCH),上顎臼歯間幅 (上顎 M3 の外側間の幅:UCW),上顎歯列長(上顎 C― M3:UTL)および下顎骨長(下顎骨前端―関節突起先 端:MdL)の 8 項目を採用して測定した(Table 2).他地 域産については,国立科学博物館所蔵のタイプ標本 M14842(E1400:岩手県産)と青森県産 1 個体 M28245 を 測定した.その他の岩手・愛媛県産の 8 個体について は,本研究で測定したタイプ標本の測定値が Yoshiyuki (1989)における同一個体の計測値とほぼ同じであること が確認できたため,Yoshiyuki(1989)の計測値を採用し た.以上の九州産と他地域産の頭骨形状を比較するため, 上記の 8 変量を用いて相関行列による主成分分析を行っ た(統計ソフト:SPSS Version 11.0 使用).本州産のク ロホオヒゲコウモリ M. pruinosus と同属近縁のヒメホオ ヒゲコウモリ M. ikonnikovi の頭骨の形状観察について は,それらが所蔵されている国立科学博物館で行った. ヒメホオヒゲコウモリの分布や分類については Kawai (2009a)に従った.
尾膜の組織片からキアゲン社の DNeasy Blood and
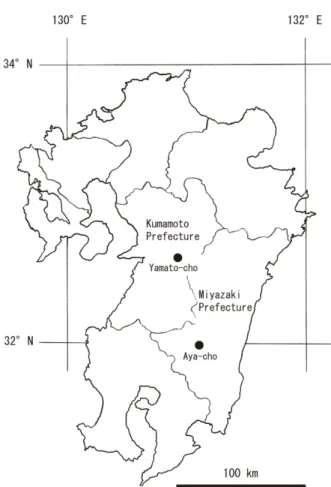
Fig. 1. Map showing the location of the study area.
Table 1. External measurements (mm) of Myotis pruinosus from Kumamoto and Miyazaki Prefectures
Individ. No. Date Sex Age HB Ta Fa Ti Hfcu E Tr Yamato-cho, Kumamoto Pref.
A 2007. 9.30 ♂ Sa 41.6 32.0 32.4 — — 11.9 6.1 B 2008. 7.28 ♂ A 41.1 34.2 33.1 15.0 8.3 13.3 5.3 C 2008. 8.24 ♂ A 38.1 31.8 31.9 15.0 9.0 12.0 5.9 D 2011.11.27 ♀ A 38.0 32.0 34.2 16.0 8.5 11.7 6.3 Aya-cho, Miyazaki Pref.
E 2003. 9. 2 ♀ A 34.0 38.0 32.3 15.0 8.0 12.5 5.5 F 2003. 9. 2 ♀ A 44.0 34.0 32.5 15.5 9.0 11.4 5.6 G 2003. 9. 2 ♀ A 39.0 36.0 35.0 15.5 9.6 10.7 5.5 H 2003. 9. 2 ♀ A 38.5 36.0 32.8 15.5 8.5 11.5 5.0 I 2006. 8.16 ♂ A 39.5 33.5 32.5 15.7 9.5 11.5 6.0 J 2006. 8.16 ♀ A 46.0 36.0 33.8 15.5 11.1 11.8 5.8 K 2006 .8.16 ♀ A 46.9 37.0 33.2 14.8 8.5 11.2 6.0 Sa: Subadult, A: Adult, HB: Head and body length, Ta: Tail length, Fa: Forearm length, Ti: Tibia length, Hfcu: Hind foot length with claw, E: Ear length, Tr: Tragus length.
Tissue Kitを用いてDNAの抽出を行った.ミトコンドリア
DNAのチトクロームb遺伝子(Cyt b)の領域全長(1140 bp)
を PCR 法によって増幅した後,配列決定をおこなった (AB855787).方法は Kawai et al.(2003)に準じた.標本 の Cyt b 遺伝子配列と GenBank 上に登録されている種と の相同性を確かめるため,GenBank 上に登録されてい るヒメホオヒゲコウモリの塩基配列(アクセッション 番号 AB106594 ~ AB106598),クロホオヒゲコウモリ (AB106607 および AB0857737),クロホオヒゲコウモリの 近縁種とされるヤンバルホオヒゲコウモリM. yanbarensis
(Kawai et al. 2003)(AB106610)との配列比較およびアラ
イメントをおこなった.Clustal W 1.82 を用いてアライ メントを行った後,さらに目視による確認も行った. また,外群としてヒメヒナコウモリ Vespertilio murinus (AF376834)を用いた. 種内および種間の平均遺伝的距離の計算および近隣結 合法による系統樹の作成は,木村の 2 パラメータ法を塩 基置換モデルとし,MEGA5.01(Tamura et al. 2011)を用 いて行った.また,最尤法およびベイズ法による系統樹 作成に際し,MEGA5.01 によってモデルテストを行い, 最適な進化モデルを探索した.この結果,赤池情報量規 準 AIC が最も小さくなる GTR + I が最適モデルとされた ため,最尤法およびベイズ法による系統樹探索にはこの モデルを用いた.最尤法による系統樹探索には RAxML (Stamatakis 2006)を用い.ベイズ法による系統樹探索に は MrBayes v.3.2.1(Ronquist et al. 2012)を用いた.解析 ではマルコフ連鎖を 2,500,000 世代繰り返し,1000 世代 ごとにサンプリングを行い Burn-in は 100,000 世代(全世 代数の 25%)に設定した. なお,本研究は環境省九州地方環境事務所から鳥獣捕 獲許可証(平成 18 年度第 0001 号,平成 19 年度第 0001 号,平成 20 年度第 0232 号,平成 21 年度第 0001 号,平成 21 年度第 0001 号,平成 22 年度第 10-0001号および平成23年度第10-0036号)を得て行われた. 結果および考察 熊本県産の個体では,4 個体中 2 個体で背面の体毛が 黒褐色または灰黒褐色で,棘毛の先端に銀白色の金属光 沢がなかった(A, B: Table 1).一方,宮崎県産の個体で は捕獲されたすべての個体において,体毛が黒色で棘毛 の先端が銀色の金属光沢が認められた.外部計測値 (Table 1)について,熊本県産と宮崎県産を合わせて(以 下,九州産とする),雌雄差をみると性差が認められな かった(Z=-0.924, P=0.356).そこで,両性をあわせ た前腕長の平均値は 33.1±0.93 mm(平均±SD, n=11) であった.これを本州・四国産(Yoshiyuki 1989;向山 1989;安井・上條 1999;上野ほか 2002;木村ほか 2003) の平均値 31.7±0.87 mm(n=15)と比較すると,有意差 が認められた(Z=-3.095,P<0.001).また,九州産 の下腿長は 15.4±0.93 mm(n=10)で,本州・四国産 (Yoshiyuki 1989;安井・上條 1999;上野ほか 2002;木村 ほか 2003)の 13.8±0.75 mm(n=12)に比べて有意に 大きかった(Z=-3.479,P<0.001). 以上の外部形態から,特に熊本県産の個体について, 大型で体毛が黒褐色また灰黒褐色で銀白色の棘毛を欠い ているタイプでは,類似の体毛を持ち外部測定値が近似 している同属のモモジロコウモリ M. macrodactylus やヒ メホオヒゲコウモリに酷似していて判別が困難であっ た.しかし,モモジロコウモリでは側膜が踵または下腿 下部に付いているのに対して,クロホオヒゲコウモリで は側膜が外足指の付け根に付いていた.近縁のヒメホオ ヒゲコウモリの特徴として北海道産の個体においては尾 膜の血管が「くの字型」の曲線走行(Kondo and Sasaki 2005;Kawai 2009a)を示している.これは,本州産も含 めて共通した特徴と予想される.しかし,クロホオヒゲ コウモリの尾膜の血管走行についての記載はないが,本 個体では直線的であった.以上の外部形質を考慮して, 本個体をクロホオヒゲコウモリと判別した.したがって,
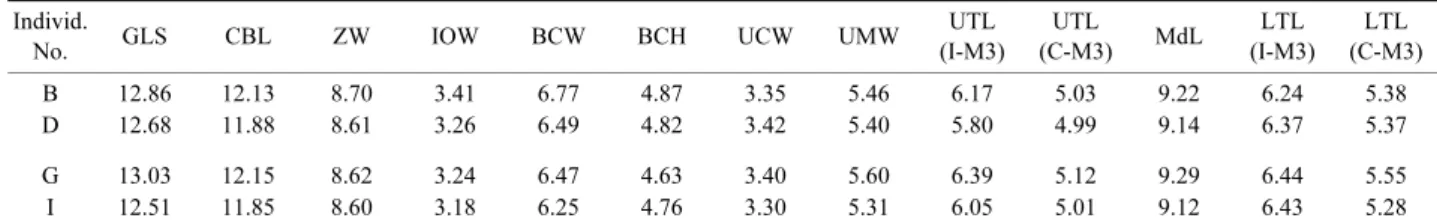
Table 2. Cranial measurements of Myotis pruinosus from Kumamoto (B, D) and Miyazaki (G, I) Prefectures Individ.
No. GLS CBL ZW IOW BCW BCH UCW UMW UTL (I-M3) UTL (C-M3) MdL LTL (I-M3) LTL (C-M3) B 12.86 12.13 8.70 3.41 6.77 4.87 3.35 5.46 6.17 5.03 9.22 6.24 5.38 D 12.68 11.88 8.61 3.26 6.49 4.82 3.42 5.40 5.80 4.99 9.14 6.37 5.37 G 13.03 12.15 8.62 3.24 6.47 4.63 3.40 5.60 6.39 5.12 9.29 6.44 5.55 I 12.51 11.85 8.60 3.18 6.25 4.76 3.30 5.31 6.05 5.01 9.12 6.43 5.28 GLS: Greatest length of skull, CBL: Condylobasal length, ZW: Zygomatic width, IOW: Interorbitral width, BCW: Width of brain case, BCH: Height of braincase at the level of median line without auditory bullae, UCW: Width across upper canines, UMW: Width across upper molars, UTL (I-M3): Length of upper tooth row from incisor to molar, UTL (C-M3): Length of upper tooth row from canine to molar, MdL: Mandible length from alveole of incisor to articular process, LTL (I-M3): Length of lower tooth row from incisor to molar, LTL (C-M3): Length of lower tooth row from canine to molar.
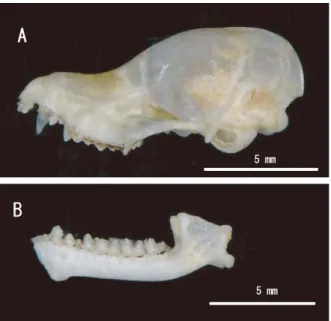
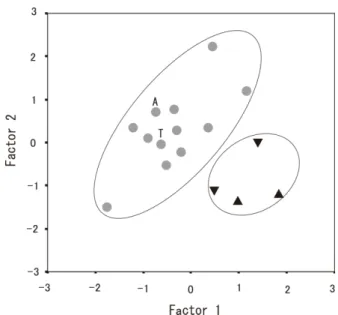
体毛の特徴や種間で重複する外部計測だけからの判別に は注意を要する. 頭骨の形状をみると,クロホオヒゲコウモリの特徴と して吻が長く側面からみた背面の輪郭は眼窩前縁よりも かなり後方(第 3 大臼歯上方)から上昇している(阿部 2007)が,熊本県産では比較的に吻が短く,側面からみ た 背 面 の 輪 郭 は 眼 窩 前 縁 の 上 方 で 上 昇 し て い て (Fig. 2A),むしろヒメホオヒゲコウモリの特徴(阿部 2007)に近い.また,クロホオヒゲコウモリの下顎の第 2 番目の小臼歯(P3)は第 1 番目の小臼歯(P2)に比べ て顕著に小さい(阿部 2007)とされているが,九州産の クロホオヒゲコウモリではヒメホオヒゲコウモリと同程 度の P3 をもつものもあり(歯冠高:0.23 ~ 0.45 mm), 変異が大きかった(Figs. 2B, 3B). 九州産の頭骨基底全長は 12.0±0.16 mm(n=4)で, 本州・四国産 12.4±0.24 mm(n=10)と比較して短かっ た.他方,眼窩間幅 8.6±0.05 mm(n=4)や脳函幅 6.5±0.21 mm(n=4)は,本州・四国産のそれら 8.2± 0.19 mm(n=10),6.2±0.11 mm(n=10)と比較して広 かった(Table 2, Figs. 3A and 4A, B).こうした頭骨の相 対的な幅広い形状は,ヒメホオヒゲコウモリ(Fig. 4C) のそれと類似している.そこで,九州産の頭骨の形態的 特徴をつかむため,主成分分析を行い,主成分得点を Table 3に示し,第 1 主成分と第 2 主成分の散布図を作成 した(Fig. 5).固有ベクトルの絶対値は第 1 主成分では ZW,次いで BCH,BCW および UMW が,第 2 主成分では CBLと MdL が大きかった.
Fig. 2. Lateral views of cranium (A) and mandible (B) of Myotis pruinosus in Kumamoto Prefecture.
Fig. 3. Ventral view of cranium (A) and dorsal view of mandible (B) of Myotis pruinosus in Kumamoto Prefecture. P2: lower second premolar, P3: lower third premolar.
Fig. 4. Dorsal view of cranium of Myotis pruinosus in Kumamoto Prefecture (A), that of the type specimen in Iwate Prefecture (B), and that of Myotis ikonnikovi in Yamanashi Prefecture (C).
これらの結果から,第 1 主成分は主に頭骨の幅広さ, 第 2 主成分は主に頭骨の長さを示していて,九州産の頭 骨は幅広で脳函高が高いが,長さが比較的短く,いわば 「ずんぐり」した頭骨の形状を示していた(Figs. 3, 4).そ れらを反映して,散布図(Fig. 5)から九州産と本州・四 国産の間で分離が可能で,特に熊本県産はより離れて明 瞭であった. DNA抽出による塩基配列について,熊本県産標本 2 個 体(B および C 個体:Table 1)のミトコンドリア DNACytb 塩基配列は,互いに完全に一致した.GenBank に登録の あったクロホオヒゲコウモリの塩基配列 AB106607(石 川県白山市市ノ瀬産;Kawai et al. 2003)とは 98.7%, AB085737(宮城県仙台市産;Sakai et al. 2003)とは 99.4% の相同性があったことから,熊本県産 2 個体はクロホオ ヒゲコウモリである可能性が高いと考えられた(Fig. 6). 加えて,近隣結合法,最尤法,ベイズ法による系統樹は 全て種ごとの高い単系統性を支持しており,また熊本県 産標本がクロホオヒゲコウモリであることを支持してい た(Fig. 6).
Table 3. Eigenvectors for the first two principal compnents based on eight skull characters (CBL, ZW, IOW, BCW, BCH, MdL, UMW and UTL (C-M3) in Myotis pruinosus
Characters PC 1 PC 2 CBL −0.053 0.839 ZW 0.947 −0.115 IOW 0.589 −0.386 BCW 0.805 −0.212 BCH 0.859 −0.463 MdL 0.680 0.674 UMW 0.779 0.537 UTL (C-M3) 0.503 0.144
Fig. 5. Scatter plots for the first and second principal components extracted from principal component analysis of eight skull characters of Myotis pruinosus. ●: Honshu and Shikoku, ▲: Kumamoto Prefec-ture, ▼: Miyazaki Prefecture. A: specimen M28245 in Aomori Prefec-ture, T: type specimen in Iwate Prefecture.
Fig. 6. Phylogenetic relationship of Myotis pruinosus based on cytochrome-b DNA sequence (1140 bp). Scale indicates genetic distance estimated by Kimura’s two parameter method on the NJ tree. Bootstrap values derived from 1000 replications for NJ tree; maximum likelihood probabilities values derived from 1000 replications; Bayesian probabilities are shown. Vespertilio murinus (AF376834) was used as the outgroup. AB106594: Yamanashi, AB106595: Tochigi, AB106596, AB106597 and AB106598: Hokkaido. AB106607: Ishikawa, AB085737: Miyagi.
熊本県産標本を含むクロホオヒゲコウモリ,形態が類 似しているとされたヒメホオヒゲコウモリの種内の平均 遺伝的距離は,前者が 0.010,後者が 0.003 となり,ク ロホオヒゲコウモリの方が種内の平均遺伝的距離が大き い傾向にあった.また,種間の平均遺伝的距離は 0.163 であり,種内の平均遺伝的距離よりも大きかった.クロホ オヒゲコウモリの種内平均遺伝的距離がより大きい傾向 にあったことについて,これは種内に地理的変異などに 起因する遺伝的変異が蓄積しているためと考えられた. 今回の調査で,特に熊本県産のクロホオヒゲコウモリ の外部形態や頭骨において,ヒメホオヒゲコウモリと類 似する点が多くみられた.こうした形状は生態的地位の 類似性を反映していると考えられる.九州にはヒメホオ ヒゲコウモリが生息していないので,熊本県産のクロホ オヒゲコウモリではヒメホオヒゲコウモリの特性も併せ 持った環境選択や行動(採食)習性がみられることが示 唆される.今後は,九州産クロホオヒゲコウモリの生態 的特性を明らかにし,同種他地域(本州・四国)のそれ らと比較考察してみたい. 謝 辞 本研究は宮崎県の本種の調査と熊本県が実施している 熊本県レッドデータブック補完調査の一環として実施さ れた調査結果に基づいている.宮崎県の調査では岩切環 境技術㈱の岩切康二氏の協力を得た.また,熊本県のコ ウモリ類調査では熊本県希少野生動植物検討委員会哺乳 類班調査員が中心となり,熊本野生生物研究会会員の協 力を得た.ここに感謝の意を表する.また,本研究を進 める上で,国立科学博物館所蔵の頭骨標本の計測等を行 う際に便宜をはかっていただいた国立科学博物館の川田 伸一郎博士,本論文を作成するにあたって適切なコメン トをいただいた 2 名のレフェリーに感謝申し上げる. 引 用 文 献 阿部 永.2007.日本産哺乳類頭骨図説(増補版).北海道大学 出版会,札幌市,290 pp.
Kawai, K. 2009a. Myotis ikonnikovi Ognev, 1912. In (S. D. Ohdachi, Y. Ishibashi, M. A. Iwasa and T. Saitoh, eds.) The Wild Mam-mals of Japan, pp. 98–100. Shoukadoh Book Sellers, Kyoto.
Kawai, K. 2009b. Myotis pruinosus Yoshiyuki, 1971. In (S. D. Ohdachi, Y. Ishibashi, M. A. Iwasa and T. Saitoh, eds.) The Wild Mammals of Japan, pp.108–109. Shoukadoh Book Sellers, Kyoto.
Kawai, K., Nikaido, M., Harada, M., Matsumura S., Lin, L. K., Wu, Y., Hasegawa, M. and Okada, N. 2003. The status of the Japa-nese and East Asian bats of the genus Myotis (Vespertilionidae) based on mitochondrial sequences. Molecular Phylogenetics and Evolution 28: 297–307.
木村吉幸・富樫祐美子・佐藤正幸.2003.福島県の翼手類IV. 福島生物 46: 29–35.
Kondo, N. and Sasaki, N. 2005. An external taxonomic character suit-able for separating live Myotis ikonnikovi and M. mystacinus. Mammal Study 30: 29–32.
前田喜四雄.2001.日本のコウモリ研究誌.東京大学出版会, 東京,203 pp.
向山 満.1989.青森県初記録の翼手目2種.Paulownia 21: 112. Ronquist, F., Teslenko M., van der Mark P., Ayres, D. L., Darling, A., Höhna, S., Larget, B., Liu, L., Suchard, M. A. and Huelsenbeck, J. P. 2012. MrBayes 3.2: efficient Bayesian phylogenetic infer-ence and model choice across a large model space. Systematic Biology 61: 539–542.
Sakai, T., Kikkawa, Y., Tsuchiya, K., Harada, M., Kanoe, M., Yoshiyuki, M. and Yonekawa, H. 2003. Molecular phylogeny of Japanese Rhinolophidae based on variations in the complete sequence of the mitochondrial cytochrome b gene. Genes & Genetic Systems 78: 179–189.
Stamatakis, A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22: 2688–2690.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. and Kumar, S. 2011. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution 28: 2731–2739.
Tamura, K. and Nei, M. 1993. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology and Evolution 9: 512–526.
上野吉雄・畑瀬 淳・小柴正記・河津 功・日比野政彦・向山 満・薄木奈津子・吉行瑞子.2002.西中国山地のコウモリ 相I.高原の自然史 7: 85–97.
安井さち子・上條隆志.1999.栃木県におけるクロホオヒゲコ ウモリMyotis pruinosus Yoshiyuki とカグヤコウモリMyotis frater Allenの初記録.栃木県立博物館研究紀要 16: 77–80. Yoshiyuki, M. 1989. A Systematic Study of the Japanese Chiroptera.
ABSTRACT
First record of the frosted Myotis, Myotis pruinosus, in Kumamoto Prefecture, Japan, with special reference to variations in their external character, skull and mitochondrial cytochrome b sequences
Kimitake Funakoshi1,*, Takuji Sakata2, Kuniko Kawai3 and Shusei Arai4
1Biological Laboratory, Faculty of Intercultural Studies, The International University of Kagoshima, Kagoshima 891-0197, Japan 2Kumamoto Municipal Chiharadai High School, Shimasaki, Kumamoto 860-0073, Japan
3Field Science Center for Northern Biosphere, Hokkaido University, Sapporo 060-0809, Japan 4General Education, Kyushu Dental University, Kitakyushu 803-8580, Japan
*E-mail: [email protected]
There has so far been no record of the frosted Myotis, Myotis pruinosus in Kumamoto Prefecture. We confirmed their inhab-itation by capturing these bats at their tunnels in Yamato-machi, Kumamoto Prefecture on 30 September 2007, 27 July and 24 August 2008, and on 27 November 2011. In some bats, the fur was not blackish but dark or grayish brown, and the tips of the guard hairs on the back had no silver metallic luster, unlike those of M. pruinosus. However, the plagiopatagium of the bats was inserted in the base of the first toe, unlike in M. macrodactylus. The bat’s blood vessels formed a straight line in the tail membrane, while those of the closely related M. ikonnikovi formed a curve. Therefore, we identified them as M. pruinosus. This was supported by phylogenetic analysis based on the mitochondrial gene (Cyt b) sequences, and genetic and geographical variations were sug-gested in this species. Forearm and tibia lengths in Kyushu were significantly larger than those in Honshu and Shikoku. In the skull characters in Kyushu, CBL was short, and ZW and BCW were large as compared with those in Honshu and Shikoku. These results were further supported by principal component analysis.
Key words: distribution, mitochondrial cyt b gene, Myotis pruinosus, Kumamoto Prefecture, intra-specific variation
受付日:2013 年 6 月 25 日,受理日:2013 年 9 月 22 日 著 者:船越公威,〒891-0197 鹿児島市坂之上8丁目34-1 鹿児島国際大学国際文化学部生物学研究室 [email protected] 坂田拓司,〒860-0073 熊本市西区島崎2-37-1 熊本市立千原台高等学校 河合久仁子,〒060-0809 札幌市北区北9条西9丁目 北海道大学北方生物圏フィールド科学センター 荒井秋晴,〒803-8580 北九州市小倉北区真鶴2-6-1 九州歯科大学総合教育学分野