平成 28 年度厚生労働科学研究費補助金(食品の安全確保推進研究事業)
「ウイルスを原因とする食品媒介性疾患の制御に関する研究」
研究協力報告
カキからのノロウイルス検出について
研究協力者 研究分担者 研究協力者 研究協力者 研究協力者 研究協力者
山本 美和子 野田 衛 則常 浩太 藤井 慶樹 八島 加八 松室 信宏
広島市衛生研究所
国立医薬品食品衛生研究所 広島市衛生研究所
広島市衛生研究所 広島市衛生研究所 広島市衛生研究所
研究要旨
2016/17 シーズンに採取されたカキのノロウイルス(NoV)浸淫状況を把握す るとともに,ヒトから検出された NoV 遺伝子型との関連性を調べた。カキか らは GⅡ.17 のみが 3 ロットから検出され、それらは Hu/GⅡ/JP/2015/G
Ⅱ.P17-GⅡ.17/Kawasaki308 に近縁な株であった。ヒトからは,全国的に流行 している GⅡ.2 が最も多く 5 株検出された。RdRp 領域の解析ができた 2 株は いずれも GⅡ.P16-GⅡ.2 であり、それらは過去に検出された株とは違うクラ スターを形成しており,ORF1-2 JCT 領域で組換えが起きた可能性が示唆され た。
カキからの NoV 検出法の改良を試みた。阻害物質の除去を目的にアセトン を添加する方法を実施したところ、NoV 検出感度は従来法と同等あるいは若干 高い結果であった。
A. 研究目的
カキ等の二枚貝はノロウイルス(NoV) による食中毒の主要な原因食品となって いる。一方、カキが関連する食中毒事例 から検出される NoV の遺伝子型は GI が比 較的多いなど、必ずしも、ヒトで流行し ている遺伝子型や二枚貝以外の食中毒事 例や集団感染事例から検出される遺伝子 型と一致するわけではない。そのため、
二枚貝から検出される NoV 遺伝子型とヒ
トから検出される遺伝子型に関するデー タを蓄積することはカキ関連食中毒の予 防や発生要因の解明に重要である。そこ で,2016/17 シーズンに採取されたカキの NoV 浸淫状況を調べるとともに,ヒトから 検出された NoV 遺伝子型と比較し、両者 の関連性を明らかにすることを目的とし た。
一方、カキからのウイルス検出検査に おいて、カキに含まれるグリコーゲン等
の阻害物質が検出感度を下げていると考 えられる。そこで,検査法を検討し,検 出感度の向上を試みた。
B. 研究方法 1. 材料
2016 年 6 月から 2016 年 9 月までに A 県 10 海域で採取したカキ 4 ロット,2016 年 10 月から 2017 年 1 月に採取された市 販カキ(A 県海域)7 ロットおよび 2016 年 9 月から 12 月までに NoV が検出された糞便 9 検体を用いた。
2. 方法
1) カキからの NoV 検出法
カキは中腸腺 1~2g を 1 検体とし実施 した。
(1)ウイルス濃縮
①アミラーゼ・ポリエチレングリコール (PEG)沈殿法
中腸腺を PBS(-)で 10%乳剤とし,α-ア ミラーゼ(和光純薬製)を添加した。37℃,
1 時間消化した後,4℃,10,000rpm,20 分間遠心し,上清を回収した。上清に PEG 6000 を 8%,NaCl を 2.1g/100ml となるよ うに添加した。4℃で一夜静置し,4℃,
10,000rpm,20 分間遠心した。沈渣に DDW を 500μl(沈渣が多い場合は 1ml)添加し,
再浮遊させた後,12,000rpm,5 分間遠心 した。
②アミラーゼ・アセトン法
中腸腺をバイオマッシャーに入れ,等 量の PBS(-)とα-アミラーゼを添加し粉 砕・撹拌した。37℃,1 時間消化した後,
PBS(-)と等量のアセトンを添加し,撹拌 した。4℃,3,000rpm,5 分間遠心し,3 層の場合は中間層,2 層の場合は上層の下
部を 500μl 分取した。
(2)NoV 検出および遺伝子型別
前処理した検体を QIAamp Viral RNA Mini Kit(QIAGEN)を用いて RNA 抽出した。
逆 転 写 反 応 は , High Capacity cDNA Reverse Transcription Kit(ライフテクノロジー ズジャパン)および Oligo(dT)Primer(ライフテク ノロジーズジャパン)を用いて行った。
増幅用プライマーは,Capsid N/S 領 域:1st primer COG1F/G1SKR,COG2F /G2SKR,2nd primer G1SKF/G1SKR,G2SKF /G2SKR を用い,semi nested PCR により 増 幅 し , 増 幅 産 物 を QIAquick Gel Extraction kit(QIAGEN)を用いて精製し た 。 BigDyeTerminator v3.1 Cycle Seqencing Kit(ABI)でサイクルシークエ ン ス を 行 っ た 後 , BigDye Xterminator Purification Kit(ライフテクノロジーズジャパン) で精製し,ダイレクトシークエンスを行 い,遺伝子配列を解析し,遺伝子型を決 定 し た 。 系 統 樹 は MEGA5.05 を 用 い , Neighbor-joining 法で作成した。
(3)α-アミラーゼの検出感度の比較 販売中止となった和光純薬製のα-ア ミラーゼ(015‐03731)(以下、旧アミラー ゼ)とその後販売されたα-アミラーゼ (017‐23671)(以下、新アミラーゼ)との 検出感度をアミラーゼ・PEG 沈殿法で濃縮 し、得られた NoV cDNA の実測値コピー数 および中腸腺 1g あたりのコピー数に換算 し比較した。
2) 糞便から検出された NoV 遺伝子型別 糞便は DDW で 10%乳剤とし,12,000rpm,
5 分間遠心した後,カキからの NoV 検出法 と同様に実施した。増幅用プライマーは,
Capsid N/S 領 域 :COG1F/G1SKR ,
COG2F/G2SKR,RdRp 領域:P1/G2SKR を用い た。
(倫理面への配慮)
本研究では,特定の研究対象者は存在 せず,倫理面への配慮は不要である。
C. 研究結果 1. NoV 検出状況
カキからの NoV 検出状況を表1に示し た。2016 年 6 月以降,6 月に GⅡ.6,7 月 に GⅡ.4 Sydney2012,11 月,12 月および 2017 年 1 月から GⅡ.17 が各 1 ロットから 検出された。
2016/17 シーズンにおけるヒトから検 出された NoV 遺伝子型を表 2 に示した。G
Ⅱ.2 が 5 株,GⅡ.4 が 2 株,GⅡ.3 および GⅡ.17 が各 1 株検出された。集団発生事 例は 2 事例のみであったが,いずれも G
Ⅱ.2 が検出された。RdRp 領域の解析がで きた 2 株の GⅡ.2 はいずれも GⅡ.P16-G
Ⅱ.2 であった。
VP1 領域(291nt)における NoV GⅡ.17 の系統樹を図1に示した。3 ロットのカキ および糞便1検体から検出された NoV は い ず れ も Hu/G Ⅱ /JP/2015/G Ⅱ .P17-G
Ⅱ.17/Kawasaki308 に近縁な株であった。
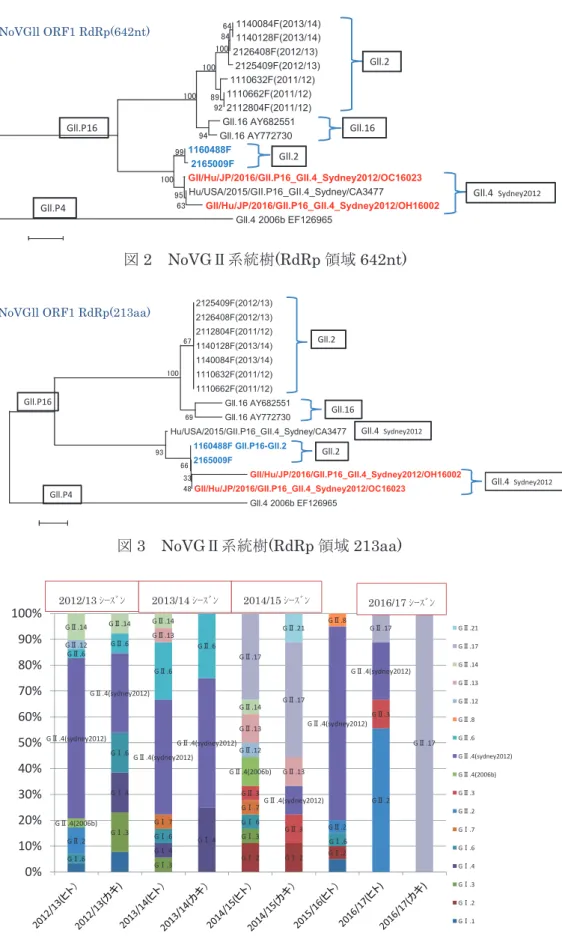
RdRp 領域(642nt,213aa)における NoV G
Ⅱ系統樹を図 2,3 に示した。2016/17 シ ーズンに検出された 2 株は,広島市にお いて 2011/12 シーズンから 2013/14 シー ズンまでに検出された 7 株とは異なるク ラスターに分類されており,入谷らが報 告 (IASR Vol.37p.136-138) し た G
Ⅱ.P16-GⅡ.4 Sydney2012 と同じクラス ターに分類された。
2012/13 シーズン以降のカキおよびヒ トからの NoV 遺伝子型別検出割合を図 4 に示した。2015/16 シーズンを除く過去 3 シーズンは,カキおよびヒトから最も多 く検出された遺伝子型が一致していたが,
2016/17 シーズン(2017 年 1 月 10 日現在) は一致しなかった。
2. アミラーゼ・アセトン法による NoV G
Ⅱ検出
同一ロットのカキ A,B,E を用いて行 った旧および新アミラーゼ・PEG 沈殿法と 新アミラーゼ・アセトン法の実測値コピ ー数および中腸腺 1g あたりに換算したコ ピー数の比較を図 5,6 に示した。
新アミラーゼを使用したアミラーゼ・
PEG 沈殿法とアミラーゼ・アセトン法を比 較すると,実測値コピー数,中腸腺 1g あ たりに換算したコピー数のいずれも同一 ロット間でのコピー数の差は認められる ものの,全体的にアミラーゼ・アセトン 法が同等あるいは若干高い結果となった。
同一ロットのカキ A,B を用いて行った 新アミラーゼ・PEG 沈殿法,新アミラー ゼ・アセトン法および旧アミラーゼ・PEG 沈殿法の実測値および中腸腺 1g あたりに 換算したコピー数の比較では、旧アミラ ーゼ・PEG 沈殿法が最も高い結果となった。
D. 考察
1.NoV 検出状況
2016/17 シーズンは,ヒトから検出され た遺伝子型は GⅡ.2 が 5 株と最も多かっ たが,カキからは GⅡ.17 のみが 3 ロット から検出された。ヒトから検出された遺 伝子型とカキから検出された遺伝子型が 異なっており,GⅡ.17 は患者としては把
握されていないものの,軽症者や不顕性 感染者は存在していることが推測された。
RdRp 領域の解析ができた 2 株の GⅡ.2 はいずれも GⅡ.P16 であった。過去に検 出された株とは異なるクラスターを形成 しており,入谷らが報告した GⅡ.P16-G
Ⅱ.4 Sydney2012 と同じクラスターに分 類された。NoVはORF1-2 JCT領域で頻 繁に組換えが起こることが知られている。
解析できた 2 株も遺伝子組換えを起こし た可能性が示唆された。
2.アミラーゼ・アセトン法による NoV G
Ⅱ検出
新アミラーゼ・アセトン法と新アミラ ーゼ・PEG 沈法を比較した場合,同一ロッ ト間でのコピー数の差は認められるもの の,全体的に同等あるいは若干高い結果 となった。旧アミラーゼ・アセトン法と 比較するといずれも低い結果となったが,
旧アミラーゼはすでに販売中止となって おり,新アミラーゼを使わざるを得ない 状況となっている。今回,新アミラーゼ・
アセトン法を検討し,検出感度はアミラ ーゼ・PEG 沈法と同等あるいは若干高い結 果であったが,アミラーゼ・アセトン法 はアミラーゼ・PEG 沈殿法に比べ,検査時 間が短いことや,高速遠心機を使用しな くても検査が可能などのメリットが得ら れた。
E. 結論
2016/17 シーズンのカキからは NoV G
Ⅱ.17 が 3 ロットから検出された。いずれ も Hu/GⅡ/JP/2015/GⅡ.P17-GⅡ.17 /Kawasaki308 に近縁な株であった。ヒト からは GⅡ.2 が 5 株と最も多く検出され た。RdRp 領域の解析ができた 2 株はいず れも GⅡ.P16-GⅡ.2 であった。それらは,
過去に検出された株とは異なり,入谷ら が報告した GⅡ.P16-GⅡ.4 Sydney2012 と 同じクラスターに分類されたことから,
ORF1-2 JCT 領域で組換えが起きた可能性 が示唆された。
カキからの NoV 検出法として検討した アミラーゼ・アセトン法の検出感度は,
従来法であるアミラーゼ・PEG 沈法と同等 あるいは若干高い結果であったが,検査 時間の短縮が図れる等のメリットが得ら れた。
F. 研究発表 1. 論文発表:なし 2. 学会発表:なし
G. 知的財産権の出願・登録状況 1. 特許取得:なし
2. 実用新案登録:なし 3. その他:なし
Hu/GII.17/HUN5737/2015/HUN Hu/GII.P17_GII.17/KR/2015/CAU‐289 3160010oy4
Hu/GII.P.17/GII.17/LNOV_PA16009/BRAZIL/2016 GII/Hu/VNM/2015/GII.P17‐GII.17/NVN15.349.
Hu/GII/JP/2014/GII.P17_GII.17/Kawasaki323 Hu/GII.17/18‐1712/GD‐JM/2014‐12‐26
Hu/GII.17/41621/Guangzhou/2014/CHN Hu/GII.17/1548/GD‐MM/2014
Hu/GII/JP/2015/GII.P17_GII.17/Kawasaki308 3170001oy4
3160012oy3
isolate 14‐BW‐3/2014/GII.P17/GII.17 1160506F
3160012oy3 3170001oy6 3170001oy1
Hu/NoV/Katrina‐17/2005/US GII/Hu/JP/2002/GII.P16_GII.17/Saitama/T87
Hu/NoV/CS‐E1/2002/USA 73
67 94
14 10 15 19 47 9 13 35
51
7 6 11
0.005
図1 NoVGⅡ.17系統樹(VP1領域291nt) 表1 カキからのNoV検出状況 採取日又は加工日
又は消費期限 採取海域 用途 結果および遺伝子型
2016/06/18 A県10海域 - GⅡ.6
2016/07/03 A県10海域 - GⅡ.4 Sydney2012
2016/08/14 A県10海域 - -
2016/09/10 A県10海域 - -
2016/10/10 A県10海域 加熱調理用 -
2016/10/28 A県14海域 加熱調理用 -
2016/11/05 A県20海域 加熱調理用 -
2016/11/25 A県10海域 生食用 GⅡ.17
2016/12/03 A県12海域 生食用 -
2016/12/22 A県10海域 加熱調理用 GⅡ.17
2017/01/10 A県10海域 加熱調理用 GⅡ.17
表2 ヒトから検出されたNoV遺伝子型(2016/17シーズン) 採取日 散発・集団の別 遺伝子型
2016/09/12 散発 GⅡ.2
2016/09/14 散発 GⅡ.3
2016/10/17 集団 GⅡ.P16-GⅡ.2
2016/10/18 散発 GⅡ.4 Sydney2012
2016/10/28 散発 GⅡ.4 Sydney2012
2016/11/10 散発 GⅡ.2
2016/11/24 散発 GⅡ.P16-GⅡ.2
2016/12/01 散発 GⅡ.17
2016/12/13 集団 GⅡ.2
NoVGll.17 ORF2 VP1(291nt)
GⅠ.2 GⅠ.2 GⅠ.2 GⅠ.3
GⅠ.3
GⅠ.3 GⅠ.4
GⅠ.4 GⅠ.4 GⅠ.6
GⅠ.6
GⅠ.6
GⅠ.6
GⅠ.6 GⅠ.7
GⅠ.7
GⅡ.2
GⅡ.2 GⅡ.3 GⅡ.2
GⅡ.3
GⅡ.3
GⅡ.4(2006b)
GⅡ.4(2006b) GⅡ.4(sydney2012)
GⅡ.4(sydney2012)
GⅡ.4(sydney2012) GⅡ.4(sydney2012)
GⅡ.4(sydney2012) GⅡ.4(sydney2012)
GⅡ.4(sydney2012)
GⅡ.6 GⅡ.6
GⅡ.6 GⅡ.6
GⅡ.8
GⅡ.12
GⅡ.12 GⅡ.13
GⅡ.13
GⅡ.13
GⅡ.14 GⅡ.14 GⅡ.14
GⅡ.14 GⅡ.17
GⅡ.17
GⅡ.17
GⅡ.17 GⅡ.21
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
GⅡ.21 GⅡ.17 GⅡ.14 GⅡ.13 GⅡ.12 GⅡ.8 GⅡ.6 GⅡ.4(sydney2012) GⅡ.4(2006b) GⅡ.3 GⅡ.2 GⅠ.7 GⅠ.6 GⅠ.4 GⅠ.3 GⅠ.2 GⅠ.1
2012/13シーズン 2013/14シーズン 2014/15シーズン 2016/17シーズン
図4 カキおよびヒトからのNoV遺伝子型別検出割合
2125409F(2012/13) 2126408F(2012/13) 2112804F(2011/12) 1140128F(2013/14) 1140084F(2013/14) 1110632F(2011/12) 1110662F(2011/12)
Gll.16 AY682551 Gll.16 AY772730
Hu/USA/2015/GII.P16_GII.4_Sydney/CA3477 1160488F Gll.P16-Gll.2
2165009F
GII/Hu/JP/2016/GII.P16_GII.4_Sydney2012/OH16002 GII/Hu/JP/2016/GII.P16_GII.4_Sydney2012/OC16023
Gll.4 2006b EF126965 48
33 66 93
69 100
67
0.005
Gll.2
Gll.16
Gll.2
Gll.4 Sydney2012 Gll.4 Sydney2012
Gll.P16
Gll.P4
図3 NoVGⅡ系統樹(RdRp領域213aa)
1140084F(2013/14) 1140128F(2013/14) 2126408F(2012/13)
2125409F(2012/13) 1110632F(2011/12) 1110662F(2011/12) 2112804F(2011/12) Gll.16 AY682551 Gll.16 AY772730 1160488F
2165009F
GII/Hu/JP/2016/GII.P16_GII.4_Sydney2012/OC16023 Hu/USA/2015/GII.P16_GII.4_Sydney/CA3477
GII/Hu/JP/2016/GII.P16_GII.4_Sydney2012/OH16002 Gll.4 2006b EF126965
99
63 95 100
94 100
92 89 100
64 84 100
0.02 Gll.P16
Gll.P4
Gll.2
Gll.16 Gll.2
Gll.4 Sydney2012
NoVGll ORF1 RdRp(642nt)
NoVGll ORF1 RdRp(213aa)
図2 NoVGⅡ系統樹(RdRp領域642nt)
2000 3000 4000 5000 6000 7000 8000 9000 10000 11000
ロット E
0 10000 20000 30000 40000 50000 60000 70000
ロット B
●:新アミラーゼ/アセトン法
◆:新アミラーゼ/PEG沈法
▲:旧アミラーゼ/PEG沈法
図6 中腸腺1gあたりコピー数の比較
コピー数 コピー数 コピー数
0 1000 2000 3000 4000 5000 6000 7000 8000
ロット A
コピー数
30 40 50 60 70 80 90
1 1.2 1.4 1.6
中腸腺重量(g)
ロット E
●:新アミラーゼ/アセトン法
◆:新アミラーゼ/PEG沈法
▲:旧アミラーゼ/PEG沈法
図5 実測値コピー数の比較
0 200 400 600 800 1000 1200 1400 1600
1.2 1.7 2.2
中腸腺重量(g) 0
20 40 60 80 100 120 140 160 180 200
1.40 1.90 2.40 2.90
中腸腺重量(g)
コピー数 コピー数 コピー数
ロット B ロット A