C a n ωr 20: 7ト77 (2011) C a rcinolo gical Society 0 1 Japan
ヤシガ二の初期生活史研究- 飼育によるアプ口ーチ
Early life history o f coconut crabs inferred f r o m culture experiments浜崎活幸
l Katsuyuki H a m a s a k i . は じめに ヤシガニ資源を保全し持続的に利用するために は,その資源維持機構に関する理解が不可欠であ る. ヤシガニは普段は陸に棲むが,海岸でふ化した 幼生は海でプランクトン生活を送り,陸に返ってく る (Drew et al., 2010) そのため,プランクトン生 活期が唯一生息域を分散 ・拡大できる時期であり, その分散・拡大能力はプランクトン生活期の長さに 依存する. プランクトン生活を送ったヤシガニは, 陸を目指して接岸し 上陸する. 上陸場所がなけれ ば,生活史はそこで分断され,個体群は早晩消滅す ることになる . このように,ヤシガニ個体群が存続 するためには,ふ化から上陸までの時期をいかにう まく切り抜けるかにかかっている . 換言すれば,プ ランク トン生活期,上陸時期や場所などの初期生活 史に関する情報は,ヤシガニの資源維持機構に対す る理解を促す また,初期生活史に関する情報に加 え,島々聞のヤシガニ個体群の遺伝的集団構造が明 らかになれば,遺伝子流動解明による幼生分散の実 態が浮き彫りになり,その保全・利用に向けた新た な視点を持つことができるであろう . し か し ヤ シ ガニの初期生活史に関する情報は少なく,また琉球 列島 内のヤシガニ個体群の遺伝的集団構造は明らか にされていない そこで,私たち東京海洋大学海洋 生物資源学科の増殖生態学研究室では,ヤシガニを 飼 育 し 初 期 生 活 史 を 調 べ て い る 飼育環境は野外 l 東京海洋大学海洋生物資源学科 〒108-8477 東京都港区港南 4-5-7Department of Marine Biosciences, Tokyo U niversity of M arine Science and Technology, 4-5-7 K onan,
Minato, Tokyo 108-8477, Japan E-mail: hamak@ kaiyodai.acj.p
とは異なるものの,小さな幼生が様々な環境条件に 対して示す反応を直接検出することができる利点が あ り,野外調査と併せると生活史研究の強力なツー ル となる. また,このような飼育研究で得られる情 報は,ヤシガニの人工繁殖技術を進展させ,人工種 苗 を 用 い た supportive breeding あ る い は creative conservation と呼ばれる保護増殖事業の展開をも可 能にするであろう. さらには ,私たちは与那国島, 鳩間島,石垣島,宮古島,沖縄本島,北大東島,与 論島,グアム,パラオ,インドネシアからヤシガニ のサンプルを得て, ミトコン ドリアD N A の塩基配 列に基づく遺伝的変異性と集団構造を明らかにしつ つある ここでは,主にふ化から上陸期までのヤシ ガニの初期生活史に関する飼育研究の概要を紹介し たい 圃 抱 卵 雌 の 飼 育 幼生の飼育実験には,飼育環境下で雌に産卵させ るか,野外で捕獲した抱卵雌を飼育し. 幼生をふ化 させる必要がある 東京海洋大学の前に所属してい た栽培漁業センターでは,エピ ・カニ類を広い水槽 で大規模に飼育し,産卵 ・ふ化させていたが,大学 の狭い研究室では無理なので,当研究室ではヤシガ ニの産卵期 (Sato
&
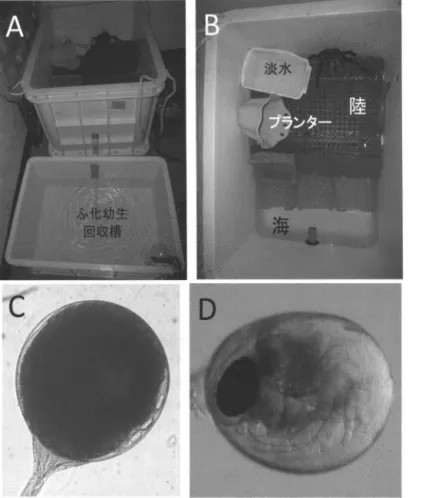
Y oseda, 2008) に八重山諸島に 出かけ,数尾の抱卵雌を捕獲し,東京まで空輸して 飼育している (H a m asaki et al., 2011 ). 搬入した抱 卵 雌 は 図 l,A B に示すような陸と海( 人工海水) を 設 け た 飼 育 容 器 に 個 別 に 収 容 し ふ化 に備える 容器内には, プランターをシェルターとして設置 し飲み水として淡水も置いている 飼育容器内の 海水は隣接したふ化幼生回収槽とつながっており, 小型のフィルタ ー付きポンプで海水を循環ろ過して 日本甲殻類学会 S抑 制 剖u m R
,
句) OrtI
73図 1. ヤシガニ抱卵雌の飼育容器 (A,S ),脹体形成卵 (C,長径0.75 m m) およびふ化直前卵 (0,長径 0.95 m m). いる. ヤシガニが逃亡しないように飼育容器には厳 重に蓋をしているが,それでもちょっとした隙間が あれば,歩脚をその隙聞にかけて,蓋をこじ開けて 逃亡することもあか朝飼育室に入ると,壁に張り 付いているヤシガニを発見することもある . 何年か 前のニュースで,日1 崎でヤシガニ発見,ペットが1 逃亡かj と聞いた時には肝を冷やした記憶がある 品)11 から多摩川の陸橋を渡 ってい ったのか? 幸い研 究室のヤシカゃニで、はなかった. なお,ふ化が終わっ たヤシガニはすべて空輸し,現地に放している . 卵 発 生 と ふ 化 幼生がいつふ化するのかが予めわかっていれば, 幼生に与える餌の培養や栄養強化などを計画的 ・効 率的に行うことができ,飼育実験を行う者にとっ 74
I
C an回r20 (2011) て,ストレスなく準備にあたることができる . 雌が 抱いている卵をピンセットでつまんで顕微鏡で観察 すると,発生がある程度進んだ卵では,成長した妊 体を透明な部位としてみることができる( 図 lC). 匹体は卵黄を吸収しながら付属肢を順次形成 ・成長 させて大きくなり,その聞に眼点が出現し,心臓が 拍動する (図l D). また,脹体に色素が発達するこ とから,雌が抱える卵塊はオレンジ色から次第に黒 褐色へと変化する. 卵を真横から見た場合の卵の側 面積に対する匹体の側面積を求めて距体成長指数と し,ふ化までの毎日の気温を積算した値( 積算温 度) との関係を調べたところ,いくつかの変曲点を 持ちながらも,) j f 体成長指数が大きくなるにともな い積算温度は直線的に減少する関係を見出した( 村 上ら,日本甲殻類学会第 46 回大会講演要旨集). こ れによ って,ふ化日をある程度予測することができi.1'"一一一・・・・帽- ・...,. . 園 田1"111ーー一一一一一一ー= 四u ・.. . . 1 1司 ...-"" ・ るようになった また,野外で捕獲したヤシガニの 卵をみることによって,捕獲した個体の産卵時期や ふ化時期をある程度推測することも可能で、ある . の行動を観察していると(山下ら,未発表),雌は 18時をすぎて暗くなると活発に動き出し ,陸域と 海域を行き来するようになる. そして陸域と海域の 浅瀬の部分で静止 し,卵のグル ー ミングを行う. ふ 化の際には海に降り,飼育容器内の水深の深い部分 へ入っていき ,全身を水の中へ入れ,潜ったまま約 10分間放幼する. 放幼が終わると,雌は陸へ上が り腹部のグルーミングを行い,放幼行動を終了す る琉球石灰岩の波打ち際で卵を抱えた腹部を海水 に濡らして放幼する ヤシガニの写真を見た記憶があ るが,私たちが観察した放幼行動とは違うようだ. これが,実験室内での特別な行動なのか,今後野外 での観察なども含めてさらに検討したいと考えてい る. . 幼生の分散と接岸 ヤシガニ幼生の分散能力を評価するためにはプラ ンク トン生活の期間を知る必要がある 一般に,十 脚類幼生の発育期間には水温が大きな影響を及ぼ し,低水温では長く,高水温では短く なる (A nger, 2001 ) . これまでにヤシガニのゾエア幼生を飼育し た研究として 2 事例が報告されていたが (Reese & Kinzie, 1968; W a n g et al., 2007),水温別に飼育した 研究はなかった. そこで,ヤシガニ幼生を 6 段階の 異 な る 水 温 (平 均 18. 9,21. 3, 24.6, 27. 0, 29.8, 32.4t ) で 飼 育 し て み た (H am asaki et al., 2009; Sugizaki et al., 2010). ふ化幼生を 30 尾ずつ 1 L の 白いポ リエチレンビ ーカーに入れ,餌としてアルテ ミアを与えてメガ ロパまで飼育したところ, 18.9 t では l 度も脱皮せず全滅した 生残率と発育・成長 は 27.0t と29.8t で良好であり,ゾエアは 4 回脱皮 してメガロパに変態し た ふ化からメガロパまでの 期間は,高水温で短縮され,2 1.3t で 54 日, 32.4t また,水温とメガロパ までの期間の関係に積算温度則を適用して求めた発 育臨界温度は 18t 程度であり,実際の飼育実験の ヤシガニの初期生活史研究 結果に一致した夏季の琉球列島近海の水温は,ヤ シガニ幼生の生残と発育に適した水温範囲内で推移 することから,ふ化からメガロパ までは 3 週間前後 を要すると考えられ. この期間 に海洋を分散できる ことになる それでは,ヤシガニのゾエア幼生は海洋のどこで 生活しているのだろうか. これに答えるためには, 海洋でのプランク トンネッ トによる採集調査が必要 であるが,すぐには実現できそうにない. 十脚類幼 生の光や重力に対する走性を調べ,海洋における幼 生の鉛直分布や深浅移動のメカニズムを理解しよう とする研究分野がある (A n ger,2001 ) そこで,飼 育 し た ヤ シ ガ ニ 幼 生 の 光 量 (0,0.0031 , 0.031, 0.3,1 3.,1 31 お よ び 310μmo1S-Im →) や 光 波 長 n mの範囲で 20 n m) に対する走性を調 べてみた (杉本ら,日本甲殻類学会第 46 回大会講 演要旨集). その結果,ヤシガニ幼生は光に対して 強い正の走性を示したが,齢期が進むほど弱い光量 (0.031 S-lm-2) に対する正の走性は弱 くなる傾向を示した光量別実験では,ソーェア幼生 は ど の 波 長 に お い て も 正 の 走 性 を示したが, 62 0 n m以上の波長( 赤い光) ではやや弱い正の走 性を示す傾向がみられた. メガロパ幼生の波長に対 する反応はゾエアと 同様であったが,正の走性は弱 くなる傾向がみられた. 以上の ことから,少なくと もゾエア期には長波長光の影響が及ばない程度の表 層に分布しているものと考えられる . メガロパになると,陸を目指して遊泳・接岸し, 上陸しなければならない. 明暗周期 (12 時間明期 . 12時間暗期) を与えた小型容器にメガロパを個体 別に収容してビデオカメラで 24 時間撮影し,その 行動を調べてみると (石山ら,未発表),メガロパ の遊泳行動は当初は活発であったが,遊泳頻度は次 第に低下し ,4日目以降にはあ ま りみられなくなっ た 一方,遊泳頻度の低下とともに,メガロパが貝 に入 って歩く行動が活発になり, 5 日目以降にはす べての観察個体が員に入り歩行 した. また,遊泳と 歩行のリズムは概潮汐 リズムと 関連 しているように みえ,遊泳行動は上げ潮時に,貝に入ってからの歩 行は下げ潮時に活発になる傾向があった. このよう に , ヤ シ ガ ニ メ ガ ロ パ 幼 生 は 選 択 的 潮 汐 移 動 (A nger,2001 ) によって接岸している可能性が示唆
Ca
ncer
2 0 (2011)I
75されている . メ ガ 口 パ の 上 陸 接岸したメガロパはいつ上陸するのであろうか. 変態当日のメガロパを様々な環境条件下 (陸の有 無,員の有無,湿度の高低,シェルターの有無) で 飼育し (図2). 第 2 齢稚ガニごろまでの行動を調 べてみた (H am asakiet al., 201 1). 親や幼生に与え た餌の栄養価によってメガロパの行動に違いがみら れたが,メガロパは早い場合は5日程度で貝殻を背 負 い 上 陸 し そ の 行 動 が 安 定 す る の は 10日ほど経 過してからであ った また,湿度が低いと上陸頻度 は減か湿度が低く,貝殻がないと潜砂頻度が増え る傾向がみられた さらに , シェルターと貝殻があ ると上陸頻度が高まり , シェルターの下に入る個体 図 2. メガロパの 上陸 実験に用いた飼 育容器 (A ) (8 c m 幅. 2 0 c m長. 6.5 c m高) と貝殻 (殻高 は4 m m程度) に入 っ て 上 陸 し た メ ガ ロ パ (8 ) 容器は人工海水を入れたバ ッ トに置き, 多数の小さな穴を通して海水が入 ってくるが, 容器の奥半分側には砂を高く盛り ,陸地とな っ ている. 7 6
I
C a n曲 r 2 0 (2011) が多くみられた . このようにヤ シガニのメガロパ は,湿度が高い場所を好む傾向が強く,そのような 環境が上陸を促した. . ヤ シガニの初期生活史 まだまだ不十分であるが,私たちが進めてきたヤ シガニの飼育研究に基づき,想像を交えながら,そ の初期生活史を類推してみよう 夏の繁殖期にふ化 した幼生は海洋の表層付近で発育し. 4 齢のゾエア を経て3週間前後でメガロパへ変態する 仮にゾエ アが黒潮本流に乗 った場合,その流速は毎秒2 m 以上にも達することから. 3 週間で琉球列島全域は 無論のこと, 日本本土へも容易に分散することがで きる したがって,琉球列島の島々に棲むヤシガニ 個体群は遺伝的には大きく異ならないことが予想さ れるが,現在までの私たちの分析ではそれを支持す る結果が得られている. メガロパは上げ潮を 利用して遊泳 ・接岸すると考 えられ,変態後数日経つ と海中で貝殻に入り歩行で きるようになる 。沖合で変態したメガロ パがいつ頃 接岸するかは分からないが,接岸したメガロパはた だちに上陸し隠れた方が潮間帯付近での被食の危険 性を回避できるように思われる 。貝殻を背負い歩行 する行動が安定し,上陸頻度が高くなるのはおよそ 10 日日ごろからであった したが って,ヤシガニ はメガロパ変態後 10 日程度かけて接岸し,貝殻を 背負い,上陸し湿度の高い海岸の転石下や植物の 堆積物の中 に潜んでいるものと想像される . 実際 に,貝殻に入ったヤシガニのメガロパや稚ガニがそ のような野外環境で発見されているが (Drew et al.,
2010; 藤田. 2011 ).発見事例は少なく ,上陸お よ び初期の生息場として重要な微細環境は明らかでは ない. 圃 お わ り に ヤシガニのゾエア幼生を飼育することは比較的容 易で,その大量飼育も可 能であ る (Sugizaki et al., 20 10) また,シェルターを与えることで,上陸後 のメガロパの大量飼育も可能であり( 浜崎ら,未発 表). 近い将来,野外において貝殻に入ったヤシガニのメガロパを放ち,その行動や移動・ 分散状況を 調査することを試みたい. また ,いったん海に出た ゾエアがメガロパとして陸に回帰するには,陸の存 在 を知る必要があるが,カニ類のメガロパでは岩礁 域の「音」 を感知して陸を知り,同種の成体 などか ら出される化学物質などを手掛かりに微細な環境 を 選択し着底するものと考え られている (Stanbleyet al.,2009) 先の人工種苗を用いた放流再捕実験や飼 育環境下で上陸場所の選択性に ついて調査すること で,不明な 点の多い上陸期とそれに続く初期稚ガニ 期の生態を解明した い と考 えてい る このような試 みの過程で,ヤ シガニの人工繁殖技術が向上し,諸 喜田 (2011) も提案 した,また私たちも保全の観点 から目標としている 「ヤシガニ牧場j の実現も夢で はないかもしれない. . 謝 辞 ここで紹介した研究は,著者が2004年7月に東 京海洋大学 に異動して以来,北田修一教授, 独立行 政法人水産総合研究センターの圏 重樹氏,およ び ヤシガニとオカヤドカリ類を研究テーマと した歴代 の研究室学生,杉崎みお,杉本斐香,村上友羽, 尾 島明日香,安藤友祐,岡部和紀, 石川琢真,山下朔 太,石山尚樹,小木曽陽介,加藤沙織,服回 宙,
i
度透知博の諸氏と取 り組んだ ものであ る ここに記 して感謝の意 を表する また , ここでの研究の一部 は,科 学 研 究 費 補 助 金( 基 盤 研 究 C , 課 題 番 号 20580198,ヤ シガニ牧場 の構 築 に向けた 基礎 的 研 究) によ った ヤシガニの初期生活史研究 圃 文 献Anger, K., 2001 . The biology of decapod crustacean larvae, Crustacean lssues, V ol. 14. A. A Balkema Publishers, Rotterdam, 419 pp
D r e w, M . M ., Harzsch, S., Stensmyr, M ., Erland, S., &
Hansson, B. S., 2010. A review of the biology and ecology of the robber crab, Birgus latro (Linnaeus,
1767) ( A n o m町a: Coenobitidae). Zoologischer
Anzeiger, 249: 45-67.
藤 田喜久. 20日 小 さなヤシガニは 何処 に い る ? Cancer, 20: 79-82
Hamasaki, K.,Sugizaki, M ., D a n, S., & Kitada, S., 2009. Effect of temperaωre on survival and developmental period of coconut crab (Birgus latro) larvae reared in the laboratory. Aquaculture, 292・259-263.
Hamasaki, K., Sugizaki, M ., Sugimoto, A., Murakami, Y目,& Kitada, S., 2011 . Emigration behaviour during sea-to-land transition of the coconut crab Birgus latro: effects of gas甘opod shells, substrata, shelters and humidity
Journal of Experimental Marine Biology and Ecology, 403: 81 -89.
Reese, E . S., & Kinzie III, R. A., 1968. The larval develop-ment of the coconut or robber crab Birgus latro (L.) in the laboratory (Anomura, Paguridea). Crustaceana, Supplement, 2: 117-144
Sato, T., & Yoseda, K., 2008. Reproductive season and female maturity size of coconut crab Birgus latro in Hatoma lsland, southern part of Japan. Fisheries Science, 74: 1277-1282
諸喜 田茂 充. 2011 ヤシ ガニ資源 の保 全に 向けて Cancer, 20: 93-95
Stanley, J. A ., Radford, C. A., & Jeffs, A. G., 2009. lnduction of settlem巴nt in crab megalopae by ambient underwater
reef sound. Behavioural Ecology, 21: 1 13- 120 Sugizaki, M ., Hamasaki, K., D a n, S., & Kitada, S., 2010.
Growth and morphogenesis of larvae reared at different temperatures and mass culture of larvae in the coconut crab Birgus latro. Aquaculture Science, 58: 135ー142.
W a n g, F. -L., Hsieh, H. -L., & Chen, C. -P., 2007. Larval growth of the coconut crab Birgus latro with a discussion on the development mode of terrestrial herrnit crabs. Journal of Crustacean Biology, 27・ 616-625.