Title 雄性生殖行動に関わる神経機構に関する研究( 本文(Fulltext) ) Author(s) 室井, 喜景 Report No.(Doctoral Degree) 博士(獣医学) 甲第218号 Issue Date 2007-03-13 Type 博士論文 Version author URL http://hdl.handle.net/20.500.12099/21401 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
雄性生殖行動に関わる神経機構
に関する研究
2006年
岐阜大学大学院連合獣医学研究科
(帯広畜産大学)
室 井 喜 景
目次 和文要旨・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・1 英文要旨・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・4 序論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・7 第 1 章 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・10 材料および方法・・・・・・・・・・・・・・・・・・・・・・・・・11 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・14 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・16 第 2 章 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・19 材料および方法・・・・・・・・・・・・・・・・・・・・・・・・・20 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・23 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・25 第 3 章 緒言・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・30 材料および方法・・・・・・・・・・・・・・・・・・・・・・・・・31 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・34 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・37 結論・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・40 謝辞・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・41 参考文献・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・42 図表・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・50
和文要旨 嗅覚は動物個体間の情報交換において重要な働きをしている。雄性生殖 行動においても嗅覚は重要な働きをしており, 雌の匂いは雄の生殖行動 が惹起されるために必須の因子である。本研究の目的は, 雄マウスが嗅覚 を介して雌マウスを認識する機構を明らかにしていくことである。 第 1 章では, 雌マウスの揮発性の匂いに対する雄マウスの嗜好性を調 べた。雄マウスが雌マウスに接触できない条件下で, 卵巣摘出した雌マウ スの匂いと, 偽手術を施した雌マウスの匂いに対する嗜好性を調べた。正 常雄マウスが, 発情前期, 発情期, 発情後期にある雌マウスの匂いを嗅 ぐ時間は, 卵巣摘出した雌マウスの匂いを嗅ぐ時間より長かった。これに 対し, 去勢雄マウスが, 偽手術を施した雌マウスの匂いを嗅ぐ時間と卵 巣摘出した雌マウスの匂いを嗅ぐ時間との間には, 有意な差は認められ なかった。これらの結果から, 雌マウスの匂いに対する雄マウスの嗜好性 が性ホルモンによる調節を受けている可能性を仮定し, 性ホルモンの投 与の影響を調べた。去勢雄マウスを 17ß -エストラジオール(E)で処置し たところ, 偽手術を施した雌マウスの匂いを嗅ぐ時間と卵巣摘出した雌 マウスの匂いを嗅ぐ時間との間には, 有意な差は認められなかった。これ に対し, 去勢雄マウスをジヒドロテストステロン(DHT)で処置したとこ ろ, 正常雄マウスとは対照的に, 発情周期に関係なく, 偽手術を施した 雌マウスの匂いを嗅ぐ時間は卵巣摘出した雌マウスの匂いを嗅ぐ時間よ り有意に短かった。去勢雄マウスを E と DHT の両方で処置したところ, 発 情前期, および発情期の雌マウスの匂いを嗅ぐ時間は, 卵巣摘出した雌 マウスの匂いを嗅ぐ時間より有意に短かった。以上の結果から, 雌マウス の揮発性の匂いに対する正常雄マウスの嗜好性は, アンドロジェンとエ ストロジェンによる調節を受けていると判断した。また, アンドロジェン が, 正常雄マウスの匂いに比べて, 卵巣摘出した雌マウスの匂いに対す る雄マウスの嗜好性を高めることがわかった。 第 2 章において, 雌マウスの揮発性の匂いの受容に対する主嗅覚系と 副嗅覚系の関与を調べた。実験では神経の活性化の指標として用いられる c-Fos の発現を調べた。雌マウスの揮発性の匂いを雄マウスに提示した 場合と, 匂いを提示しなかった場合との間では, 主嗅球顆粒細胞層にお ける c-Fos 陽性細胞数に変化は認められなかった。この結果と対照的に, 雌マウスの揮発性の匂いを提示しなかった場合に比べ, 雄マウスに揮発 性の匂いを提示した場合は, 副嗅球顆粒細胞層における c-Fos 陽性細胞 数が有意に増加した。この結果から, 雄マウスが雌マウスの揮発性の匂い
を副嗅覚系を介して受容していると判断した。第 1 章において, 雌マウス の匂いに対する嗜好性が性ホルモンによって調節されていると判断され たことから, 性ホルモンの投与が雌マウスの匂いの受容過程におよぼす 影響を調べた。ホルモン処置の有無に関わらず, 雌マウスの揮発性の匂い を雄マウスに提示した場合, 副嗅球顆粒細胞層における c-Fos 陽性細胞 数は有意に増加した。この場合, 正常雄マウスでは副嗅球顆粒細胞層の前 部より後部で c-Fos 陽性細胞数が多かった。去勢から 3 日後の雄マウス では, 正常雄マウスと同様に, 前部より後部で c-Fos 陽性細胞数が多か ったが, 去勢から 7 日後の雄マウスでは, 前部と後部の c-Fos 陽性細胞 数に差は認められなかった。この結果から, 副嗅球において, 去勢の影響 が去勢処置の 3 日以降に現れ, 雌マウスの匂いの処理に関係するシナプ ス伝達が性ホルモンによる調節を受けていることが示唆された。そこで, 去勢雄を E で処置したところ, 副嗅球顆粒細胞層前部と後部における c-Fos 陽性細胞数に差は認められなかったが, 去勢雄を DHT で処置した ところ, 正常雄と同様に, 副嗅球顆粒細胞層の前部より後部で c-Fos 陽 性細胞数が多かった。一方, 去勢雄を E と DHT の両方で処置したところ, 副嗅球顆粒細胞層の前部と後部で c-Fos 陽性細胞数に差は認められなか った。主嗅球においては, c-Fos 陽性細胞の発現にホルモン処置の影響 は認められなかった。以上の結果から, 副嗅球における雌マウスの揮発性 の匂いの受容過程に E と DHT の両方が関与し, E が DHT の働きを阻害し ていると判断した。 第 3 章において, 最初に発情期の雌マウスと卵巣摘出した雌マウスが それぞれ居住するケージ内の床敷きの匂いに対する雄マウスの嗜好性を 調べた。雄マウスが床敷きに接触できない条件下で調べたところ, 第 1 章で雌マウスを用いた場合と異なり, 床敷きの匂いを嗅ぐ時間に有意な 差は認められなかった。この結果は, 床敷き中には放出しない匂いを雌マ ウスが放出していることを示唆している。この可能性を検討するため, 副 嗅球における c-Fos の発現を調べたところ, 発情期の雌マウスの匂いを 提示した場合と対照的に, 床敷きの匂いを提示した場合は, c-Fos 陽性 細胞数に有意な増加は認められなかった。この結果から, 雌マウスは床敷 き中に放出しない匂いを放出していると判断した。雄マウスの匂いを提示 した場合, c-Fos 陽性細胞数の増加は認められなかった。また, 卵巣摘 出した雌マウスの匂いを提示した場合, 副嗅球顆粒細胞層の後部におい てのみ c-Fos 陽性細胞数が増加した。以上の結果から, 雌マウスに接触 できない条件下で雄マウスの副嗅覚系で受容される匂いは, 雌マウスに 特異的であり, 卵巣機能に依存して放出されると判断した。次に, この匂
いを介して雄マウスがどのような情報を得ているのかを調べるため, habituation-dishabituation 試験を行った。偽手術を施した雄マウ スと対照的に, 鋤鼻器摘出した雄マウスは, 雌と接触できない条件下で, 発情期の雌マウスと卵巣摘出した雌マウスの匂いを区別できなかった。こ の結果から, 雄マウスは副嗅覚系を介して雌マウスの卵巣の有無を識別 していると判断した。 以上を要するに, 本研究の結果から, 雄マウスは副嗅覚系の神経機構 を介して雌の繁殖に関する情報を受容し, 雌マウスに特異的な揮発性の 匂いを処理すると判断した。また, この情報伝達に関わるシナプス伝達は, 副嗅球において性ホルモンの調節を受けると判定した。さらに性ホルモン は, 雌マウスの揮発性の匂いに対する雄マウスの嗜好性も調節している ことを認めた。以上より, 雄マウスが雌マウスの匂いを受容し, 処理し, 応答する一連の過程は性ホルモンの調節下にあると判定した。
英文要旨
Olfaction has an important role in animal communication. Olfactory cues affect many behaviors, including male sexual behavior. The purpose of the present study is to clear the mechanism for males recognizing females via olfaction.
In the first chapter, in order to compare the attraction of male mice to volatile female odors, I measured the period during which a male mouse spent sniffing volatile odors from a sham-operated female mouse or an ovariectomized mouse without visual or tactile contact. Intact, but not castrated, male mice spent more time sniffing volatile odors from proestrous, estrous, or metestrous females than from ovariectomized females. To investigate the involvement of sexual hormones in this behavior, castrated male mice were treated with 17ß-estradiol (E), or dihydrotestosterone (DHT), or both. E-treatment did not affect sniffing behavior when male mice were exposed to sham-operated and ovariectomized females. Regardless of the estrous stage, DHT-treatment induced less sniffing behavior of the volatile odors from sham-operated than from ovariectomized female mice. Both E- and DHT-treatment induced less sniffing behavior during exposure to proestrous or estrous females than to ovariectomized females. These results suggest that neither androgen nor estrogen are sufficient for reproducing male attraction to volatile female mouse odors, and that androgen signaling has a competitive effect against the attraction.
In the second chapter, I investigated c-Fos expression in the major olfactory bulb (MOB) and the accessory olfactory bulb (AOB) after exposure to volatile female odors without physical contact. Volatile female odors increased c-Fos positive cells in the granule cell layer (GCL) of the AOB, but not in the GCL of the MOB, suggesting that male mice detect volatile female odors via the
accessory olfactory system. Next, I investigated hormonal modulation of signals from volatile female odors. In intact male mice, there were more c-Fos positive cells in the caudal GCL than in the rostral GCL. This effect was observed 3 d but not 7 d after castration, suggesting that hormonal deficiency causes the reorganization of the AOB after 3 d. There was no difference in the number of c-Fos positive cells between the rostral and caudal GCL of castrated male mice treated with E. In contrast, there were more c-Fos positive cells in the caudal GCL than in the rostral GCL of castrated male mice treated with DHT. In both E- and DHT–treated castrated male mice, there was no difference in the number of c-Fos positive cells between the rostral GCL and caudal GCL. Hormonal treatments did not affect c-Fos expression in the MOB. These findings suggest that E disrupts the effect of DHT in the AOB, and that androgen is required for maintaining the intact neuronal network of the AOB. Sexual hormones modulate the signals from volatile female odors in the AOB of male mice.
In the final chapter, I investigated more features of volatile female odors, and what information male mice receive via the volatile female odors. There was no difference in the total time spent sniffing volatile odors from soiled bedding collected from the cages of estrous or ovariectomized female mice, suggesting that female mice emit volatile odors which are not excreted into bedding. To test this possibility, I investigated c-Fos expression in the AOB after exposure of male mice to volatile odors without physical contact. In contrast to volatile female odors, volatile odors from estrous bedding did not increase the number of c-Fos positive cells in the GCL of the AOB. Volatile odors from a male mouse did not increase the number of c-Fos positive cells. Volatile odors from an ovariectomized female mouse increased the number of c-Fos positive cells only in the caudal GCL of the AOB. These results suggest that female mice emit the female-specific
volatile odors which are not excreted into bedding, and which are modulated by ovarian functions. To characterize the female-specific volatile odors, we conducted habituation-dishabituation tests. Whereas sham-operated male mice discriminated between volatile odors of estrous and ovariectomized female mice, vomeronasal organ-removed male mice did not. These results suggest that male mice discriminated whether or not female mice were ovariectomized, by volatile odors via the accessory olfactory system.
The present study suggests that male mice obtain the information about the reproduction of female mice by detecting the female-specific volatile odors via the accessory olfactory system, and that this neuronal system is modulated by sexual hormones. Sexual hormones also regulate male attraction to volatile female odors. Thus, sexual hormones regulate a series of male sexual behavior (i.e. from processing of odor information to individual response).
序論 動物は外界の変化に対応するため, 外界の様々な情報を取り入れ生命を維持 している。外界の情報は五感を介して受容されるが, そのうち嗅覚は重要な働 きをしている。環境中の様々な匂いを受容することにより, 食餌の探索や危険 の回避といった働きのほか, 同種, 異種の個体間の匂い情報の交換を介して, 種の区別, 縄張り, 性の区別, 個体認識などにも関っている(7, 8, 12, 50, 57, 58)。 匂いの受容, 処理を行う嗅覚系は, 多くの動物において主嗅覚系と副嗅覚系 の 2 つにより構成されている(12, 15)。主嗅覚系は食餌などの匂いに加え, フ ェロモン(同種の個体間で情報交換を担う物質(8, 25, 58))を受容する(7, 8, 19,57)。主嗅覚系は受容細胞である嗅上皮に始まり, 主嗅球を経て, the anterior olfactory nucleus, the olfactory tubercles, the lateral amygdala, the piriform cortex といった脳領域に入力する (17)。これに対し, 副嗅覚系はフェロモンを受容する経路としての役割を果た している(7, 8, 12, 58)。副嗅覚系は受容組織である鋤鼻器の鋤鼻神経に始 まり副嗅球を経て, the bed nucleus of the accessory olfactory tract, the medial amygdaloid nucleus, the posteromedial cortical amygdaloid nucleus, the bed nucleus of the stria terminalis に入力する(7, 8, 70)。主嗅覚系と副嗅覚系はそれぞれ 異なる神経核により構成されるが, いくつかの脳領域でそれら 2 つの系 からの入力が合流する可能性が示唆されており(32, 39), 受容された情 報が統合, 処理され, 個体反応が惹起されると考えられる。 雄性生殖行動は嗅覚系による調節を受けており, 調節機構に関して多 くの研究がなされている(5, 9, 10, 22, 26, 27, 34, 49, 54, 61, 62)。生殖行動に関わる匂いとして複数のフェロモンが同定されており, 雌雄の情報交換に重要な働きをしている(7, 8, 12, 58)。 また, 雄性生殖行動が内分泌系による調節を受けていることもよく知 られている(45)。このことは精子形成だけではなく, 雌を妊娠させるま での一連の反応が内分泌系により調節されることを意味し, 脳の高次機 能が内分泌系による支配を受けていることを表している。 以上の知見を踏まえ, 本研究では雄性生殖行動と嗅覚の関わり, 特に 雄が嗅覚により雌を認識する機構を明らかにすることを目的とする。第 1 章では, 雌マウスの揮発性の匂いに対する雄マウスの嗜好性と性ホルモンの関 係を調べた。第 2 章では, 雌マウスの揮発性の匂いの受容過程への性ホルモン の関与を調べた。第 3 章では, 雌マウスの揮発性の匂いの性質を検討し, この
第 1 章
緒言 雄性生殖行動は「欲求行動」と「完了行動」の二つに区分される(4)。完了行 動はマウンティングに始まり射精に至るまでの生殖行動を意味する。欲求行動 は完了行動以前にみられる行動を意味し, そのひとつに嗅覚を介した雌の存在 の認識がある。完了行動は内分泌系により調節を受けていることから(45), 欲 求行動も内分泌系により調節を受けている可能性がある。 嗅覚により雄が雌の存在を認識する機構と性ホルモンの関係に関して従来か ら多くの研究がなされている(10, 26, 48, 54, 71)。しかしこれらの研究 は, 雄が匂いの源と直接接触できる条件下で行われている。このため, 雄は嗅 覚だけでなく視覚や触覚による情報も受容できる環境下にあり, 嗅覚による認 識機構に対する性ホルモンの働きを限定的に研究しているとは言い難い。また 多くの研究では匂いの源として雌の尿や居住するケージ内の床敷きが用いられ ている(10, 22, 26, 27, 47, 48)。雌の尿中には雄を誘引する物質が多く 含まれているが(10), 唾液(6), 皮脂(23), 涙(31)の中にフェロモンを出す 動物もいることから, 雌が雄を誘引する物質を尿以外から出している可能性が あり, 嗅覚による認識機構を研究するために用いる匂いの源として尿や床敷き は不十分だと考えられる。また, 匂いの源として麻酔して不動化した雌を用い た研究があるが(34), 麻酔薬は性腺刺激ホルモンや性ホルモンの分泌を阻害 することから(14, 36), 麻酔薬の投与が生殖に関係する匂いの放出に影響す る可能性がある。 そこで, 本研究では, 従来の研究の問題点を踏まえ, 雌の情報を嗅覚によっ てしか受容できない条件下で, かつ匂いの源として自由行動下の雌マウスを用 いて, 雄マウスの生殖行動を観察した。
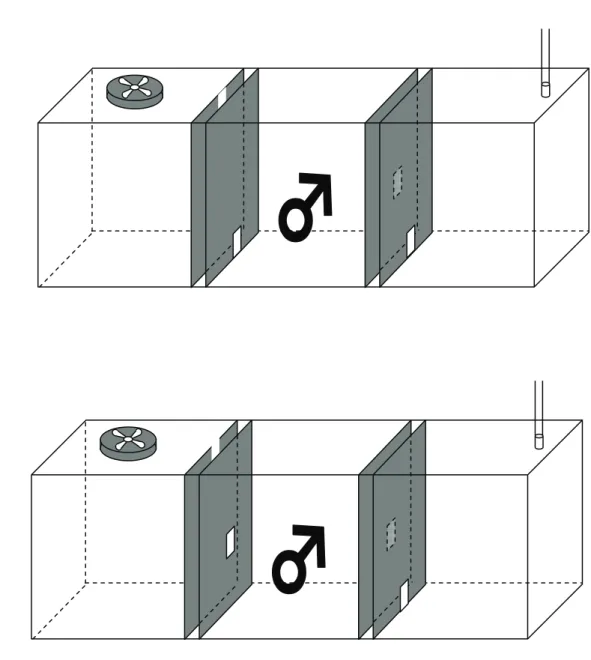
材料および方法 動物 日本エスエルシー(浜松)から購入した雌雄ともに 7 週齢から 15 週齢の交尾 経験のない ddY 系マウスを用いた。マウスはアクリル製ケージ(奥行き 250 mm, 幅 180 mm, 高さ 120 mm)内で, 飼料(固型:日本クレア, 東京)と水を自由に 摂取できる条件下で単独飼育した。飼育室内は温度 22±2℃, 湿度 35±5 %に 維持し, 午前 6 時に明期が始まる明暗 12 時間周期で管理した。 去勢手術およびホルモン投与 行動測定を開始する 1 週間前に, アバチン(360 mg/kg;Sigma, 米国)の腹 腔内投与による麻酔下で去勢手術を行った。ホルモンを投与する場合は, 結晶 状のジヒドロテストステロン(DHT), または 17ß-エストラジオール(E)(和光 純薬工業, 大阪)を長さ 1cm のシリコーン製チューブ(内径 1 mm, 外径 2 mm, 三商, 東京)に詰め, チューブの両端をペースト状シリコーン(信越化学工業, 東京)で塞いだものを, 去勢手術時に肩甲骨間皮下に移植した(5)。なお, ホル モンを詰めたチューブは移植する 24 時間前から 37℃の生理的食塩水の中で加 温しておいた。 匂いの源として用いた雌マウス 匂いの源として用いる雌マウスには, 用いる 10 日前までにアバチン麻酔下 で卵巣摘出または偽手術を施した。個体差による影響を軽減するため, 同一の 雄に提示する雌マウスは同腹子とし, 偽手術と卵巣摘出手術を同じ日に行った。 偽手術を施した雌の発情周期の判定は, 16 時から 18 時の間に膣スメアを採取 し, Nothnick(52)の基準に従って判定した。発情周期は発情前期, 発情期, 発情後期, 発情間期に分類し, 4 日間で発情周期を回帰する雌のみを匂いの源 に用いた。不動化した雌を匂いの源として用いる場合は, ペントバルビタール ナトリウム(60 mg/kg; ナカライテクス, 京都)の腹腔内投与により麻酔した。 行動測定装置 新規に開発した 2 種類(タイプ 1, タイプ 2)のアクリル製装置を用いて行動 測定を行った(Fig.1a, 2)。両装置は密閉された直方体(奥行き 200 mm, 幅 572 mm, 高さ 200 mm)の形をしており, 2 つの仕切りにより 3 等分されてい る(風上の部屋, 中央の部屋, 風下の部屋)。それぞれの仕切りは不透明な 2 枚 の板により構成されており, 金網で仕切られた正方形の窓(30x30 mm)が異な る高さに設置されている。タイプ 1 の装置では風上の部屋から順に, 床と同じ
高さ, 床から高さ 7 cm, 床と同じ高さ, 天井の高さに各々の窓が設置されて いる(Fig.2a)。タイプ 2 の装置では風上の部屋から順に, 床と同じ高さ, 床 から高さ 7 cm, 床から高さ 7 cm, 天井の高さに各々の窓は設置されている (Fig.2b)。実験装置の風上の部屋の天井には実験室の外に繋がっているホース が接続されており, 風下の部屋の天井につけてあるファンにより空気を排出す ることにより, 実験室内と温度, 湿度を同じに調節した空気を実験室の外から 吸引してきた。これにより, 中央の部屋を通って風上の部屋から風下の部屋に 向けて空気が一方向にのみ流れる仕組みになっており, 匂いも風上から風下に 向けて一方向にのみ流れる仕組みになっている。風量は 0.035 m3/min に調節 した。雄マウスが立ち上がって高さ 7 cm の位置にある窓に触る行動を匂いを嗅 ぐ行動と見なした(Fig.1b)。 行動測定の方法 行動測定は, 18 時から 23 時までの暗期に弱赤色光下で行った。 タイプ 1 の装置を用いた測定 1 日 1 回 6 日間にわたり同一の雄を測定に用いた。測定をする際には, まず 中央の部屋に雄を入れ 30 分間環境に慣らした。1 日目の測定では, その後風上 の部屋に雌を入れずに雄が風上の窓に触る時間を 30 分間測定した。2 日目以降 の測定では, 30 分間の慣らす操作の後, 偽手術または卵巣摘出した雌を風上の 部屋に入れ, 雄が風上の窓に触る時間を 30 分間測定した。偽手術を施した雌は 4 日間連続して使用し, 卵巣摘出した雌は偽手術を施した雌を用いた前日また は後日に使用した。雌の匂いをかがせる順番の影響を考慮して, 2 日目に用い る雌の発情ステージは雄により変えた。 麻酔して不動化した発情期の雌を匂いの源として用いる場合, 同様な手順に 従って, 雄が風上の窓にタッチする時間を 30 分間測定した。 タイプ 2 の装置を用いた測定 匂いの源として発情期の雌を用いた。中央の部屋に雄を入れ 30 分間環境に慣 らす操作をした後, 風上の部屋または風下の部屋に雌を入れて, 雄が風上また は風下側の窓に触る時間を 30 分間測定した。 マウンティングの観察 上記の測定が終了した後, マウンティング行動を観察した。雌の発情を促す ため, 測定に用いる少なくとも 1 週間前までに卵巣摘出した雌に試験の 48 時間 前と 24 時間前に E(0.02 mg)を, 4 時間前にプロジェステロン(0.5 mg; 和 光純薬, 大阪)を皮下投与した。測定に用いる 1 時間前に交尾経験のある雄と同 居させ雄への受容性があることを確認した。雌が逃避したり振り向いて攻撃す
ることなく腰部周囲に雄が上腕を回すことを許した場合, 雌に受容性があると 判定した。受容性の確認後すぐに雌を別のケージに移し単飼した。被験雄と発 情雌を同居させ, 20 分以内に腰部の前後運動を伴うマウンティング行動が認め られるかを観察した。 統計処理 測定装置内で雄マウスが雌マウスの存在を認識しているかを調べる実験と, 聴覚の関与を調べる実験のデータ解析には F 検定の後, スチューデントの t 検 定を用いた。雌の発情周期の違いによる雄マウスの嗜好性の変化をみる実験の データ解析にはバートレット検定の後, 一元配置分散分析法を用いた。ポスト ホック検定としてダネット検定を用いた。マウンティングの回数の比較には, フィッシャーの直接確率計算法を用いた。
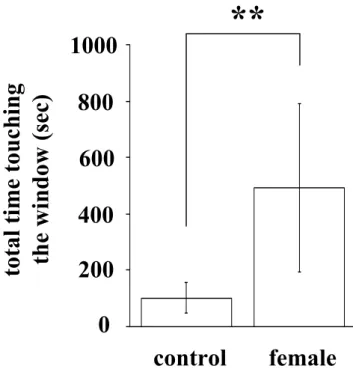
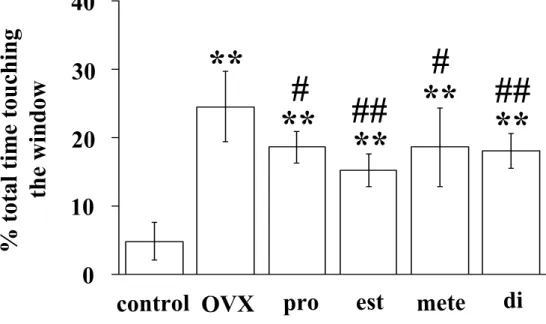
結果 雌マウスの匂いに対する雄マウスの行動 最初に, 雌マウスの揮発性の匂いに対する雄マウスの行動を調べるため, タ イプ 1 の測定装置を用いて実験を行った。風上の部屋に雌がいない場合に比べ て, 発情期の雌が風上の部屋にいる場合には, 雄マウスが風上の窓にタッチす る時間が有意に長かった(いない場合, 102.0±54.7 sec; いる場合, 493.2±298.2 sec; Fig.3)。この結果は, 風上の部屋に雌マウスがいるこ とを雄マウスが認識していることを示している。 嗅覚を介した雌マウスの認識 この測定条件では雄マウスが雌マウスの存在を聴覚により認識している可能 性がある。そこで, この可能性についてタイプ 2 の装置を用いて検討した。も し雄マウスが雌マウスを聴覚により認識しているのであれば, 風下の部屋に雌 マウスがいる場合に風下の窓にタッチする時間と, 風上の部屋に雌マウスがい る場合に風上の窓にタッチする時間との間に差は認められないはずである。風 下の部屋に雌がいる場合に風下の窓にタッチする時間は, 風上の部屋に雌がい る場合に風上の窓にタッチする時間に比べて有意に短かった(Fig.4a)。また, 風下の部屋に雌がいる場合に風下の窓にタッチする時間と, 風下の部屋と風上 の部屋のどちらにも雌がいない場合に風下の窓にタッチする時間との間に有意 な差は認められなかった(Fig.4a)。さらに, 物音や鳴き声を発することを できなくするために, 麻酔し不動化した発情期の雌マウスをタイプ 1 の 測定装置の風上の部屋に入れて窓にタッチする時間を測定した。雌マウス を麻酔した場合と麻酔しなかった場合の間で, 雄マウスが風上の窓にタ ッチする時間に有意な差は認められなかった(無麻酔の雌を用いた場合, 468.3 ± 325.9 秒; 麻酔をした雌を用いた場合, 569.2 ± 231.6 秒; Fig.4b)。以上の結果は, 雄マウスが風上の雌マウスを聴覚により 認識しているのではなく, 嗅覚により認識していることを示している。 嗅覚による雌マウスの認識に対する性ホルモンの影響 タイプ1の装置を用いて, 発情前期, 発情期, 発情後期, 発情間期の各ステ ージにある雌マウスの匂い, および卵巣摘出した雌マウスの匂いに対する雄マ ウスの嗜好性を調べた。雄の去勢やホルモン処置, 雌の卵巣の有無に関わらず, 風上に雌がいる場合は風上に雌がいない場合に比べて窓にタッチする時間は有 意に長かった(Fig.5, 6, 7, 8, 9)。この結果は, 去勢やホルモン処置に関 わらず, 雄は風上の部屋にいる雌を認識していることを示している。
雌マウスの匂いに対する正常雄マウスの嗜好性 正常雄マウスでは, 正常雌の発情周期の違いによって窓にタッチする時間に 有意な差は認められなかった(Fig.5)。しかし, 卵巣摘出した雌が風上の部屋 にいる場合に比べて, 発情前期, 発情期, 発情後期の雌が風上の部屋にいる場 合では, 正常雄マウスが窓にタッチする時間は有意に長かった(Fig.5)。これ らの結果は, 卵巣摘出雌マウスの揮発性の匂いよりも, 発情前期, 発情期, 発 情後期の正常雌マウスの揮発性の匂いに対して, 雄マウスは高い嗜好性を示す ことを示している。正常雄マウスが雌マウスにマウンティングするか否かを調 べたところ, すべての正常雄マウスはマウンティング行動を示した(Table 1)。 雌マウスの匂いに対する去勢雄マウスの嗜好性 去勢した雄マウスのマウンティング行動を調べたところ, すべての去勢雄マ ウスは雌マウスにマウンティング行動を示さなかった(Table 1)。そこで, 雌 マウスの揮発性の匂いに対する去勢雄の嗜好性を調べた。正常雄と異なり去勢 雄では, 卵巣摘出した雌が風上の部屋にいる場合と, 正常雌が風上の部屋にい る場合とで, 窓にタッチする時間に有意な差は認められなかった(Fig.6)。以 上の結果は, 正常雄マウスに認められた雌マウスの揮発性の匂いに対する嗜好 性とマウンティングする能力が去勢によって失われたことを示している。 E で処置した去勢雄マウスにおける雌マウスの匂いに対する嗜好性 精巣由来のテストステロンはアンドロジェン受容体を活性化する。また, エ ストロジェンに変換されてエストロジェン受容体も活性化する。雌にマウンテ ィングするためにはアンドロジェンよりも, むしろエストロジェンが必要であ るとの報告がある(45)。そこで, E で処置した去勢雄の雌マウスの揮発性の匂 いに対する嗜好性を調べた。従来の報告どおり, E で処置した去勢雄は, マウ ンティングする能力を有意に回復した(Table 1)。これに対し, 去勢雄と同様 に, 卵巣摘出した雌が風上の部屋にいる場合と, 正常雌が風上の部屋にいる場 合とで, 窓にタッチする時間に有意な差は認められなかった(Fig.7)。以上の 結果から, E はマウンティングする能力を回復するが, 雌マウスの揮発性の匂 いに対する嗜好性を回復しないことがわかった。 DHT で処置した去勢雄マウスにおける雌マウスの匂いに対する嗜好性 雄性生殖行動ではアンドロジェンが重要な働きをする (68)。そこで, エス トロジェンに変換されないアンドロジェンである DHT で処置した去勢雄を用い て実験を行った。去勢雄と同様に, すべての雄はマウンティング行動を示さな
かった(Table 1)。一方, 窓にタッチする時間は発情周期に関係なく, 正常雌 が風上の部屋にいる場合に比べて, 卵巣摘出した雌が風上の部屋にいる場合の 方が長かった(Fig.8)。以上の結果から, DHT は去勢雄のマウンティングする 能力を回復せず, 正常雄とは対照的に, 正常雌マウスよりも卵巣摘出した雌マ ウスの匂いに対する嗜好性を高めることがわかった。 E と DHT で同時処置した去勢雄マウスにおける雌マウスの匂いに対する嗜好性 去勢雄の雄性生殖行動を完全に回復させるためにはアンドロジェンとエスト ロジェンの両方が必要である (68)。そこで, E と DHT の両方で処置した去勢 雄を用いて実験を行った。従来の報告どおり (45), E と DHT の同時処置によ り, 去勢雄のマウンティング能力は有意に回復した(Table 1)。これに対して, 窓にタッチする時間は, 卵巣摘出した雌が風上の部屋にいる場合は, 発情前期, および発情期の雌が風上の部屋にいる場合より有意に長かった(Fig.9)。 発情 後期, および発情間期の雌が風上の部屋にいる場合には有意な差は認められな かった。以上の結果から, E と DHT の同時処置により, マウンティングする能 力は回復するが, 正常雄とは対照的に, 発情前期, および発情期の雌の匂いよ り卵巣摘出した雌の匂いに対する嗜好性を高めることがわかった。
考察 雄マウスでは, 卵巣摘出した雌マウスの揮発性の匂いを嗅ぐ時間より, 発情 前期, 発情期, 発情後期にある正常雌マウスの揮発性の匂いを嗅ぐ時間の方が 長かった(Fig.5)。この結果は, 雄マウスが卵巣摘出した雌マウスの匂いより, 発情前期, 発情期, 発情後期にある正常雌マウスの匂いに嗜好性を示すことを 示している。雄ラットも同様に卵巣摘出した雌ラットの匂いより, 発情期の雌 ラットの匂いに嗜好性を示すことが知られている(34)。従って, このような雄 の嗜好性は雌の繁殖能力に基づいている可能性がある。ラットを用いたこの研 究では, ペントバルビタールにより麻酔した雌を用いて同様な実験が行われた。 しかし, 麻酔薬は性腺刺激ホルモンの分泌を阻害することから (14, 36), 雌 の匂いの放出に影響する可能性を考え, 本研究で嗜好性を調べる場合は無麻酔 の雌を用いた。先の研究(34)で麻酔した雌を用いても本研究と同様に, 卵巣摘 出した雌の匂いより発情期の雌の匂いに嗜好性を示す結果が得られたのは, エ ストラジオール ベンゾエートとプロジェステロンの投与により雌の発情を人 工的に誘起したため, 麻酔薬による性腺刺激ホルモンの分泌阻害の影響を受け なかったためだと考えられる。 雌の尿や居住しているケージ内の床敷きを用いた場合も, 卵巣摘出した雌の 匂いより発情期の雌の匂いに対して嗜好性を示す(10, 22, 26, 54)。これら の研究は, 尿や床敷きに雄が直接接触できる条件下で行われており, 雄は揮発 性の匂いに加え不揮発性の匂いも受容することができる。さらに視覚や触覚と いった嗅覚以外からの情報も受容できる。本研究では雄マウスが視覚や触覚に よって雌マウスの情報を受容することを制限し, 嗅覚を介した揮発性の匂いの 受容のみを可能にしている。このことから, 本研究の結果は, 正常雄マウスが 卵巣摘出した雌と正常雌の揮発性の匂いを区別し, それぞれに異なる嗜好性を 示すことを表している。雄性生殖行動は内分泌系により調節されている(45)。 去勢した雄マウスでは正常雌と卵巣摘出した雌の匂いに対する嗜好性に差は認 められなかった(Fig.6)。同様に, E で処置した去勢雄は正常雌と卵巣摘出し た雌の匂いに対する嗜好性に差を示さなかった(Fig.7)。DHT で処置した去勢 雄は, 正常雄とは対照的に, 発情周期に関係なく, 正常雌マウスの匂いよりも 卵巣摘出した雌マウスの匂いに対して嗜好性を示した(Fig.8)。E と DHT の両 方で処置した去勢雄は, 発情前期, 発情期にある正常雌の匂いよりも卵巣摘出 した雌の匂いに対して嗜好性を示した(Fig.9)。以上の結果から, アンドロジ ェンとエストロジェンは雌の匂いに対する雄の嗜好性の決定に関与することが 示された。また, この嗜好性に関して, エストロジェンがアンドロジェンの働 きを阻害していることが示された。
雄性生殖行動は欲求行動と完了行動に区別され, 欲求行動に始まり完了行動 に終わる(4)。雄による雌の匂いの認識は欲求行動に, マウンティングは完了行 動に含まれる。E で処置した去勢雄はマウンティング能力を回復したが, 雌の 匂いに対する嗜好性に関しては去勢雄と同様であった。これに対し, DHT で処 置した去勢雄はマウンティングする能力を回復しなかったが, 雌の匂いに対す る嗜好性は正常雄とは対照的であった。これらの結果は, 一連の生殖行動であ っても欲求行動と完了行動が内分泌系により異なる調節を受けていることを示 している。オピオイドは欲求行動と完了行動のそれぞれに対して異なる調節を 行っている (69)。アンドロジェンやエストロジェンはオピオイド系を介して生 殖行動を調節している可能性がある。 嗅覚系は主嗅覚系と副嗅覚系の二つから構成される(12, 15)。扁桃体内側部 は副嗅覚系を構成する神経核であり(15), 主嗅覚系によっても活性化される (27, 39)。雄ラットは卵巣摘出した雌の匂いより, 発情雌の匂いに対して強い 嗜好性を示すが, 扁桃体内側部を破壊することによりこの嗜好性は失われる (34)。扁桃体内側部はアンドロジェン受容体とエストロジェン受容体を発現し ており(64), 雄性生殖行動の調節に重要な働きをしている(55)。このことから, 雌の匂いに対する雄の嗜好性は扁桃体内側部の働きによって決まっていること が示唆される。 雄性生殖行動においてアンドロジェンは, 中枢機能よりも陰茎の勃起や射精 といった末梢機能に重要であるとされている(3)。その一方で, アンドロジェン 受容体は雄性生殖行動の制御に関わる脳領域に発現しており(2), アンドロジ ェンが鳴き声, 尿によるマーキング, 性の区別など中枢性の機能を調節してい ることが示唆される(43)。本研究の結果は, アンドロジェンが雌の揮発性の匂 いに対する嗜好性を調節していることを示しており, 末梢機能だけでなく中枢 性にも働いている可能性を示している。
緒言 匂いの源と接触できない条件下であっても, 主嗅覚系は動物の揮発性 の匂いにより活性化される(40)。これに対し, 副嗅覚系はたとえ動物の 匂いであっても, 匂いの源と直接接触しなければ活性化されないとされ ている(41)。このことは, 副嗅覚系が揮発性の匂いによって活性化され ない可能性を示している。しかし, 鋤鼻器のスライス標本を用いた実験や, マウスの行動解析を用いた実験から, 雄の尿中にみられる α-または ß-farnesene や 2-heptanone のような揮発性フェロモンが, 副嗅覚系 を活性化することが知られている(38, 67)。したがって, 主嗅覚系と同 様に匂いの種類によっては, 非接触条件下であっても副嗅覚系が活性化 される可能性がある。 本章では, まず雌マウスの揮発性の匂いの受容過程への主嗅覚系と副嗅 覚系の関与を調べた。次に, 第 1 章において, 雌マウスの揮発性の匂いに 対する雄マウスの嗜好性がアンドロジェンやエストロジェンによって調節され ることが示されたことから, これらのホルモンによる主嗅覚系と副嗅覚系の調 節機構を調べた。
材料および方法 動物 日本エスエルシー(浜松)から購入した雌雄ともに 7 週齢から 15 週齢の交尾 経験のない ddY 系マウスを用いた。マウスはアクリル製ケージ(奥行き 250 mm, 幅 180 mm, 高さ 120 mm)内で, 飼料(固型:日本クレア, 東京)と水を自由に 摂取できる条件下で単独飼育した。飼育室内は温度 22±2℃, 湿度 35±5%に維 持し, 午前 6 時に明期が始まる明暗 12 時間周期で管理した。行動測定や脳のサ ンプリングは 18 時から 23 時までの暗期に弱赤色光下で行った。 鼻腔内への硫酸亜鉛水溶液の灌流 嗅上皮を特異的に破壊するため, McBride K.らの方法 (44)に従い, 行動 測定を開始する 24 時間前に, ジエチルエーテル麻酔下で, 5% 硫酸亜鉛水溶液, または対照として生理的食塩水を左右両方の鼻腔に 50 μl ずつ鼻鏡側から灌流 した。誤嚥を防ぐため, 頭側をやや低くして仰臥位に固定し, アスピレーター で喉頭側から余分な溶液を吸引除去しながら, 吸気にあわせて水溶液を灌流さ せた。 鋤鼻器摘出手術 Saito らの方法 (62)に従い, アバチン(360 mg/kg;Sigma, 米国)の腹 腔内投与による麻酔下で, 鋤鼻器摘出手術または偽手術を実験の 2 週間前に行 った。鋤鼻器が完全に摘出されているかを確認するため, すべての測定が終了 後に c-Fos の染色と同様の手順(以下参照)で biotinylated soybean agglutinin(1:10000)(J-Oil Mills, 東京)を用いて染色した (53)。 soybean agglutinin は鋤鼻神経に特異的に結合する性質を持つ (29)。こ のため, 両方の副嗅球の glomerular cell layer における染色性が完全に 欠如している場合に鋤鼻器が完全に摘出されたと判定し, データとして用いた (Fig.13c, d)。 匂いの源として用いる雌の準備 用いる雌マウスは実験に供する少なくとも 1 週間前から単独飼育した。実験 に供する日の 16 時から 18 時の間に膣スメアを採取し, Nothnick (52)の基 準に従って発情期の雌のみを選別し, 実験に用いた。 雌マウスの匂いを嗅ぐ時間の測定 5% 硫酸亜鉛水溶液, または生理的食塩水を鼻腔内に灌流した雄マウスを,
タイプ 1 の測定装置の中央の部屋に入れ, 30 分間の慣らしの操作の後, 風上に 発情期の雌を入れ, 30 分間風上の窓に触る時間を測定した。 去勢手術およびホルモン処置 第 1 章に示す方法に従って, 去勢手術やホルモン処置を行った。去勢した雄 は手術の 3 日後, または 7 日後に, ホルモンで処置した去勢雄は手術の 7 日後 に実験に用いた。 雄マウスへの匂いの提示と脳のサンプリング タイプ 1 の測定装置を用いて発情期の雌マウスの匂いを提示した。中央の部 屋に雄を入れ 30 分間の慣らしの操作の後, 風上の部屋に発情期の雌を入れ 60 分間匂いを提示した。その後, ジエチルエーテル麻酔下で脳を採取し, パラホ ルムアルデヒドで固定した。匂いを提示しない場合は 30 分間の慣らしの操作の 後, 風上の部屋に雌を入れず 60 分後に同様の手順で脳を採取した。 免疫染色
固定した脳は Microslicer(LINEARSLICER, PRO7, DOSAKAEM, 京都) を用いて 40 μm の厚さで矢状断した。それぞれの切片はリン酸バッファー(PBS) で洗浄後, 室温で 60 分間, 0.5% Triton X-100 加 PBS(PBST)中においた。 1%H2O2加 PBS で 20 分間処置した後, 2%ヤギ血清加 PBST でさらに 1 時間処置
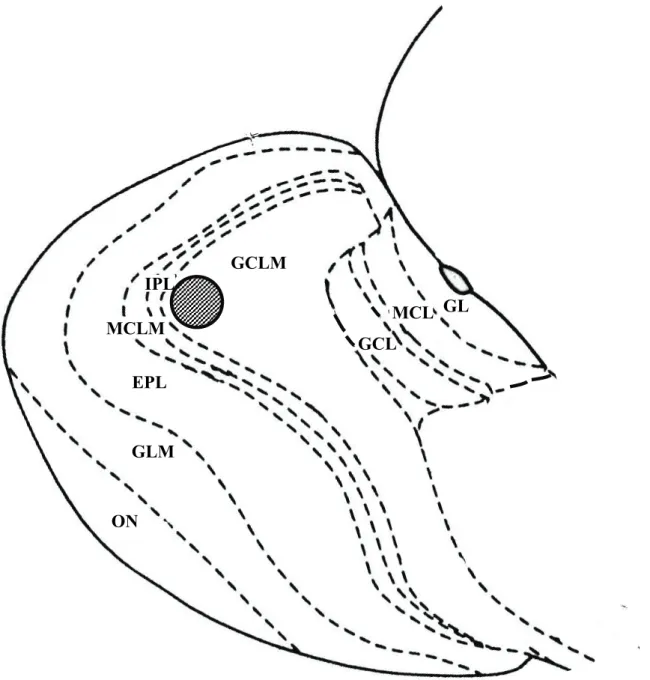
した後, PBST で 5000 倍希釈したウサギ由来 c-Fos 抗体(Ab-2; Calbiochem, 米国)を 24 時間, 4℃で処理した。PBST で洗浄後, ビオチン化ヤギ由来抗ウサ ギ抗体(1:200)(Vectastain elite ABC kit; Vector Laboratories, 米国)を 2 時間処置後, avidin-biotin peroxidase complex(1:200)で 2 時間処置した。嗅球の外側端から内側に向けて 600 μm から 720 μm の範囲 で 2 枚の切片を採取した。各個体の両側の嗅球から同様の手順で切片を選び出 し, 1 個体あたり合計 4 枚の切片を評価に用いた。主嗅球の顆粒細胞層, 副嗅 球の僧房細胞層と顆粒細胞層における c-Fos 陽性細胞数を評価した。主嗅球顆 粒細胞層では直径 400 μm の円の中の c-Fos 陽性細胞数を評価した(Fig.10)。 副嗅球では, 僧房細胞層と顆粒細胞層内の c-Fos 陽性細胞数を前部と後部にわ けて評価した。前部と後部は副嗅球の前端と後端の中央で区別した。4 枚の切片 のデータの平均を個体のデータとして用いた。 統計処理 雌の匂いを提示した雄と提示していない雄における, 主嗅球, および副嗅球 の c-Fos 陽性細胞数の比較と, 去勢やホルモンで処置した雄の間での c-Fos
陽性細胞数の比較には, 二元配置分散分析法を用いた。ポストホック検定とし て, ターキークレーマー検定を用いた。各群内の副嗅球前部と後部の間での c-Fos 陽性細胞数の比較には, 対応のある t 検定を用いた。鋤鼻器摘出群と偽 手術群における c-Fos 陽性細胞数の比較は F 検定の後, スチューデントの t 検 定を用いた。危険率 5%以下で有意差ありとみなした。
結果 主嗅覚系を介した雌マウスの匂いの受容 主嗅球顆粒細胞層における c-Fos 陽性細胞の数は, 雌の匂いを提示した場合 と提示しなかった場合の間で有意な差は認められなかった(Fig.11a, c, d)。 この結果は, 雄マウスが主嗅覚系を介して雌マウスの匂いを受容していない可 能性を示している。しかし, 鼻腔内に硫酸亜鉛水溶液を灌流し, 鋤鼻器の働き に影響を与えず, 嗅上皮のみを特異的に破壊 (44)した雄マウスに, 雌マウス の匂いを提示したところ, 匂いを嗅ぐ行動はほぼ完全に抑制され, 生理的食塩 水を灌流した雄マウスに比べて匂いを嗅ぐ時間は有意に減少した(Fig.11b)。 以上の結果は, c-Fos の発現によっては検出できないが, 雄マウスが主嗅覚系 を介して雌マウスの匂いを受容していることを示している。 副嗅覚系を介した雌マウスの匂いの受容 副嗅球では, 雌の匂いの提示により, 僧房細胞層における c-Fos 陽性細胞 数に有意な増加は認められなかったが, 顆粒細胞層における c-Fos 陽性細胞数 に有意な増加が認められた(Fig.12)。また顆粒細胞層前部と後部の c-Fos 陽 性細胞数を比較したところ, 前部より後部で c-Fos 陽性細胞数が多かった (Fig.12)。以上の結果は, 雄マウスが副嗅覚系を介して雌マウスの揮発性の匂 いを受容していることを示している。 雄マウスが雌マウスの揮発性の匂いを鋤鼻器で受容している可能性を検討す るため, 鋤鼻器摘出した雄マウスの副嗅球における c-Fos の発現を調べた。発 情期の雌マウスの匂いを提示した場合, 偽手術群と鋤鼻器摘出群の間で僧房細 胞層における c-Fos 陽性細胞数に有意な差は認められなかった(Fig.13a, e, f)。しかし顆粒細胞層では, 偽手術を施した雄マウスとは対照的に, 鋤鼻器摘 出した雄マウスでは c-Fos 陽性細胞はほとんど存在しなかった(Fig.13b, e, f)。以上の結果は, 雄マウスは鋤鼻器を介して発情期の雌マウスの揮発性の匂 いを受容していることを示している。 主嗅球における性ホルモンの影響 正常雄, 去勢手術から 3 日後の雄, 去勢手術から 7 日後の雄, DHT で処置し た去勢雄, E で処置した去勢雄, DHT と E の両方で処置した去勢雄の主嗅球顆 粒細胞層において, 匂いの提示の有無[F(1, 47)=3.60], ホルモンの処置 [F(5, 47)=2.21]に関して, c-Fos 陽性細胞数に有意な差は認められなかっ た。匂いの提示とホルモン処置の間で交互作用は認められなかった[F(5, 47)=0.28]。
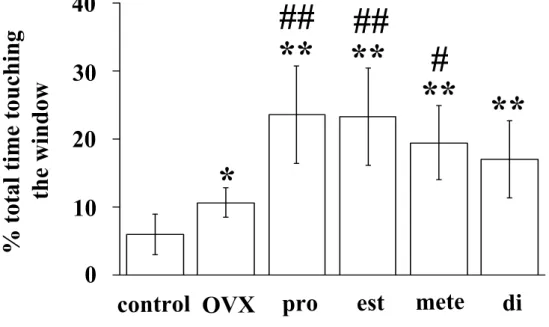
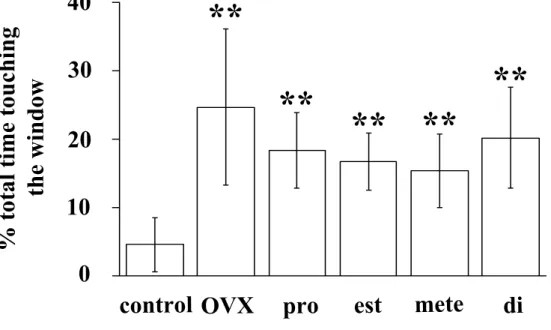
副嗅球における性ホルモン 発情期の雌マウスの匂いを提示していない場合, 僧房細胞層の前部と後部の 比 較 [F(1, 24)=8.35, p<0.01], ホ ル モ ン 処 置 に よ る 比 較 [F(5, 24)=3.49, p<0.05]で c-Fos 陽性細胞数に有意な差が認められた。僧房細胞 層 の 前 後 と ホ ル モ ン 処 置 の 間 で 交 互 作 用 は 認 め ら れ な か っ た [F(5, 24)=0.44]。ポストホック検定では有意な差は認められなかった(Fig.16a, 17a, c, e, g, i, k)。 発情期の雌マウスの匂いを提示した場合, 僧房細胞層の前部と後部の比較で は c-Fos 陽性細胞数に有意な差は認められなかったが[F(1, 72)=0.02], ホ ルモン処置による比較 [F(5, 72)=6.00, p<0.01]では有意な差が認められ た。僧房細胞層の前後とホルモン処置の間で交互作用は認められなかった[F(5, 72)=0.98]。ポストホック検定の結果, E で処置した去勢雄と DHT と E の両方 で処置した去勢雄は, 去勢から 7 日後の雄に比べて, 僧房細胞層後部の c-Fos 陽性細胞数が有意に少なかった(Fig.16c, 17f, j, l)。 発情期の雌マウスの匂いを提示した場合は匂いを提示しなかった場合に比べ て , 僧 房 細 胞 層 の 前 部 [F(1, 48)=11.87, p<0.01], 後 部 [F(1, 48)=22.41, p<0.01]のそれぞれにおいて c-Fos 陽性細胞数に有意な増加が 認められた。ポストホック検定の結果, 去勢から 7 日後の去勢雄の後部におい て, 匂いの提示による c-Fos 陽性細胞数の有意な増加が認められた(Fig.16a, c, 17e, f)。 発情期の雌マウスの匂いを提示していない場合, 顆粒細胞層の前部と後部の 比 較 [F(1, 24)=4.47, p<0.05], ホ ル モ ン 処 置 に よ る 比 較 [F(5, 24)=7.57, p<0.01]で有意な差が認められた。顆粒細胞層の前後とホルモン 処置の間で交互作用は認められなかった[F(5, 24)=0.81]。ポストホック検 定では有意な差は認められなかった(Fig.16b, 17a, c, e, g, i, k)。 発情期の雌マウスの匂いを提示した場合, 顆粒細胞層の前部と後部の比較 [F(1, 72)=41.33, p<0.01], ホルモン処置による比較 [F(5, 72)=4.64, p<0.01]でそれぞれ有意な差が認められた。顆粒細胞層の前後とホルモン処置 の間で交互作用が認められた[F(5, 72)=4.32, p<0.01]。ポストホック検定 の結果, 正常雄, 去勢から 3 日後の雄, DHT で処置した去勢雄では, 顆粒細胞 層の前部より後部のほうが c-Fos 陽性細胞数が有意に多かった(Fig.16d, 17b, d, h)。また, DHT で処置した去勢雄に比べて, DHT と E の両方で処置 した去勢雄では, 顆粒細胞層後部の c-Fos 陽性細胞数が有意に少なかった (Fig.16d, 17h, l)。 発情期の雌マウスの匂いを提示した場合と提示しなかった場合を比べて, 顆
粒細胞層の前部[F(1, 48)=103.93, p<0.01], 後部[F(1, 48)=146.13, p<0.01]のそれぞれにおいて有意な差が認められた。ポストホック検定の結果, すべての群の雄マウスにおいて, 発情期の雌マウスの匂いを提示した場合, c-Fos 陽性細胞数に有意な増加が認められた(Fig.16b, d, 17)。
考察 発情期の雌マウスの匂いを提示した場合と提示していない場合との間 で, 主嗅球顆粒細胞層における c-Fos 陽性細胞数に有意な差は認められ なかった(Fig.11, 14, 15)。同様な結果が雌の尿などを匂いの源とし て用いた他の研究でも報告されている(17, 54)。この結果は雄マウスが 主嗅覚系を介して雌マウスの揮発性の匂いを受容していない可能性を示 す。しかし, 生理的食塩水を鼻腔内に灌流した雄マウスとは対照的に, 硫 酸亜鉛水溶液を灌流し嗅上皮を特異的に破壊した(44)雄マウスは, 発情 期の雌マウスの匂いを嗅ぐ行動をほとんど示さなかった(Fig.11b)。こ の結果は, 雄マウスが主嗅覚系を介して雌マウスの匂いを受容している ことを示している。主嗅覚系はいくつかのフェロモンをはじめ, 餌の匂い など一般的な匂いの受容を担っている(12)。雄マウスは主嗅覚系を介し て環境中の様々な匂いを常に受容しているため, 雌の匂いを提示した場 合と提示しなかった場合とで, 主嗅球顆粒細胞層の c-Fos 陽性細胞数に 有意な差は認められなかったと考えられる。 これに対し, 雄マウスに発情期の雌マウスの匂いを提示した場合, 副 嗅球顆粒細胞層の前部と後部において c-Fos 陽性細胞数は有意に増加し た(Fig.12b, c, d)。しかし, 鋤鼻器摘出した雄マウスに発情期の雌マ ウスの匂いを提示しても, 副嗅球の顆粒細胞層における c-Fos 陽性細胞 数の有意な増加は認められなかった(Fig.13b, e, f)。これらの結果は, 発情期の雌マウスの揮発性の匂いを雄マウスは鋤鼻器を介して受容して いることを示している。 鋤鼻器摘出群と偽手術群に発情期の雌マウスの匂いを提示した場合, 顆粒細胞層における c-Fos 陽性細胞数に有意な差は認められたが, 僧房 細 胞 層 に お け る c-Fos 陽性細胞数に有意な差は認められなかった (Fig.13)。本研究の結果と同様に, 鋤鼻器摘出した雄ラットと偽手術を 施した雄ラットのそれぞれが雌ラットと交尾した場合, 顆粒細胞層にお ける c-Fos 陽性細胞数に有意な差が認められたが, 僧房細胞層における c-Fos 陽性細胞数に有意な差は認められなかった(35)。橋の青斑核を構 成するノルアドレナリン神経は軸索を副嗅球に伸ばし, 僧房細胞とシナ プスを形成している (46)。この青斑核からのノルアドレナリン神経は, 雌マウスが交尾した相手の雄マウスの匂いを記憶するために重要な働き をしている (59)。雄マウスにおいても青斑核ノルアドレナリン神経が僧 房細胞の働きを調節している可能性がある。 本研究の結果と異なり, Luo らは匂いの源と直接接触しなければ副嗅
覚系は活性化されないと報告した(41)。Luo らは僧房細胞層における神 経の活動を微小電極により記録しており, 雌マウスと接触しない状態で は, 雄マウスの僧房細胞が興奮しないことを示した。本研究の結果も, Luo らの報告に一致し, 雌の揮発性の匂いにより僧房細胞層における c-Fos 陽性細胞数に変化は認められなかった(Fig.12a, c, d)。僧房 細胞は顆粒細胞と相反性シナプスを形成しており(66), 先の報告から (16, 17, 54), 僧房細胞層と顆粒細胞層における c-Fos の発現は同調 すると予想した。雄ラットに 60 分間警戒フェロモンを提示した後, 副嗅 球における c-Fos の発現を調べた研究では, 本研究の結果と同様に, 顆 粒細胞層の c-Fos 陽性細胞数の有意な増加は認められたが, 僧房細胞層 の c-Fos 陽性細胞数に有意な変化は認められなかった(33)。同じ研究グ ループの他の報告では, 30 分間警戒フェロモンを提示した場合は, 僧房 細胞層と顆粒細胞層の両層において c-Fos 陽性細胞数の有意な増加が認 められた(30)。この結果から, 僧房細胞層における c-Fos の発現の違い はフェロモンを提示する時間の違いに起因する可能性がある。しかし本研 究では, 予備実験において, 発情期の雌マウスの匂いを 15,30 分間提示 した場合, 僧房細胞層と顆粒細胞層の両層において c-Fos 陽性細胞数の 有意な増加は認められなかった。このことから, 60 分間発情期の雌の匂 いを提示した場合に, 僧房細胞層における c-Fos 陽性細胞数が増加しな かったのは, 匂いの提示時間に原因があるとは考えにくい。正常雄マウス に発情期の雌マウスの匂いを提示した場合, 僧房細胞層における c-Fos 陽性細胞数に変化は認められなかったが, 去勢から 7 日後の雄マウスで は僧房細胞層後部における c-Fos 陽性細胞の数は有意に増加した(Fig.16, 17)。また, E で処置した去勢雄の僧房細胞層の後部や, DHT と E の両方で処 置した去勢雄の僧房細胞層前部と後部における c-Fos 陽性細胞数は, 去勢か ら 7 日後の雄に比べて有意に少なかった(Fig.16, 17)。この結果から, 雌の 揮発性の匂いによる僧房細胞の活性化が E や DHT によって抑制されている可能 性がある。顆粒細胞は γ-アミノ酪酸(GABA)を介して僧房細胞の働きを抑制す る(66)。雄の副嗅球において, テストステロンは GABA 合成酵素である glutamic acid decarboxylase(GAD)の活性を高める(51)。テストステロ ンはアンドロジェンとして, またはエストロジェンに変換されて働くことから, E や DHT の処置により GAD の活性が高まり, 正常雄やホルモンで処置した去勢 雄の顆粒細胞における GABA の合成, 放出が増加し, 去勢から 7 日目の雄マウ スに比べて, 僧房細胞の活性が抑制されている可能性がある。このために, 正 常雄マウスに発情雌マウスの匂いを提示した場合, 顆粒細胞層における c-Fos 陽性細胞数は増加するが, 僧房細胞層における c-Fos 陽性細胞数に変化は認め
られなかった可能性がある。 発情期の雌マウスの揮発性の匂いを提示した場合, 正常雄マウスの顆 粒細胞層における c-Fos 陽性細胞数は前部より後部で有意に多かった (Fig.16b, c, d)。僧房細胞層における結果と同様に, 去勢から 3 日後の雄 マウスの顆粒細胞層では c-Fos 陽性細胞数は前部より後部で有意に多く, 去勢 の効果は認められなかった。これに対し, 去勢から 7 日後の雄マウスの顆粒細 胞 層 で は 前 部 と 後 部 の 間 で c-Fos 陽 性 細 胞 数 に 差 は 認 め ら れ な か っ た (Fig.16d, 17b, d, f)。In vivo で性ホルモンが数分以内に効果を発揮す る機構が存在するが(56), 以上の結果は, 去勢から 3 日後以降に副嗅球にお ける神経伝達が変化している可能性を示している。DHT で処置した去勢雄マ ウスの副嗅球顆粒細胞層における c-Fos 陽性細胞数は前部より後部で有意 に多かった(Fig.16d, 17h)。DHT はエストロジェンに変換されないため, アンドロジェン受容体を介した情報伝達によって正常雄の神経伝達機構が維持 されている可能性がある。E だけ, または DHT と E の両方で処置した去勢雄マ ウスでは, 前部と後部の c-Fos 陽性細胞数に差は認められなかった(Fig.16d, 17j, l)。この結果は, E が DHT の働きを阻害していることを示している。他 の脳領域でみられるように, E はアンドロジェン受容体に作用することで(63), 副嗅球における神経伝達を調節している可能性がある。以上の結果は, DHT や E だけでは正常雄の神経伝達は維持できないことを示している。正常雄の神経伝 達を維持するために, アンドロジェンやエストロジェン以外の精巣由来の因子 が関与している可能性がある。 鋤鼻器には V1Rs (11)と V2Rs (18, 42, 60)の 2 つのファミリーの フェロモン受容体が発現している。鋤鼻器の頂端側の受容上皮は V1Rs を (60), 基底側の受容上皮は V2Rs を発現している (18)。頂端側の受容 上皮は副嗅球の前部に, 基底側の受容上皮は副嗅球の後部にそれぞれ軸 索を入力し(18), 僧房細胞層前部と後部を構成する僧房細胞にそれぞれ シナプスする(24)。この知見から, 副嗅球における c-Fos 陽性細胞の分 布は, 匂いの受容に携わる受容体のタイプを反映している可能性が示さ れる。 性ホルモンの処置によって顆粒細胞層における c-Fos 陽性細胞の分布が 異なるという結果から, 性ホルモンがフェロモン受容体の発現を調節し ている可能性がある。実際に, V2Rs ファミリーの 1 つである VR1 の発 現は性ホルモンにより調節されている(1)。発情雌マウスの揮発性の匂い を提示した場合, DHT は顆粒細胞層後部の c-Fos 陽性細胞数のみを変化 させた(Fig.16d)。この結果は, DHT が V2Rs ファミリーの発現を調節 している可能性を示している。しかし, V2Rs は V1Rs と異なり, 非常に
多様性に富む細胞外ドメインを N 末端側に持っており, このドメインを 介して低分子のペプチドを認識すると考えられている(37, 42)。したが って, 雌マウスの揮発性の匂いの受容は V2Rs を介していないと考えら れ, DHT の処置によって顆粒細胞層後部の c-Fos 陽性細胞数が変化する のは V2Rs の発現の変化の結果ではないと考えられる。顆粒細胞層におけ る c-Fos 陽性細胞の分布パターンが変化する機構に関しては, 更なる研究が 必要である。 第1章において, 雌マウスの揮発性の匂いに対する雄マウスの嗜好性 はアンドロジェンやエストロジェンにより調節されることを示した。アン ドロジェン受容体やエストロジェン受容体は多くの脳領域に発現してお り(64), 雌マウスの匂いに対する雄マウスの嗜好性は多くの脳領域の相 互作用により決まると考えられる。しかし, 副嗅球は副嗅覚系の神経伝達 において最初にシナプスを形成する場であり, 匂いの記憶形成といった 高度な脳機能の中枢でもある(21)。性ホルモンによる副嗅球における神 経伝達の調節は, 雌マウスの揮発性の匂いに関する神経伝達に大きく影 響すると考えられる。
緒言 第 1 章において, 性ホルモンが雌マウスの匂いに対する嗜好性を調節 することを示した。第 2 章において, 雄マウスは副嗅覚系を介して雌マウ スの揮発性の匂いを受容することを示した。さらに, 性ホルモンが雌マウ スの揮発性の匂いに関する神経伝達を調節することを示した。 本章では, 雄マウスの副嗅覚系を介して受容される雌マウスの揮発性 の匂いの性質をさらに検討し, この匂いにより雄マウスがどのような情報を 得ているのかを調べた。
材料および方法 動物 日本エスエルシー(浜松)から購入した雌雄ともに 7 週齢から 15 週齢の交尾 経験のない ddY 系マウスを用いた。マウスはアクリル製ケージ(奥行き 250 mm, 幅 180 mm, 高さ 120 mm)内で単独飼育した。室温 22±2℃, 湿度 35±5% に管理され, 午前 6 時に明期が始まる明暗 12 時間周期に管理された飼育室で動 物は飼育された。飼料(固型:日本クレア, 東京)と水は自由に摂取できる条件 にした。 提示する匂い 実験に供する少なくとも 10 日前までに, 雌マウスはアバチン(360 mg/kg; Sigma, 米国)の腹腔内投与による麻酔下で, 卵巣摘出手術, または偽手術を 施し, その後単独飼育した。偽手術を施した雌の発情周期を判定するため, 16 時から 18 時の間に膣スメアを採取し, Nothnick(52)の基準に従って発情期 の雌のみを選び出した。匂いの源として用いる雄マウスは, 実験に供するまで 単独飼育した。雌マウスが居住するケージ内の床敷きを採取するため, 20 g の ウッドチップを敷いたアクリルケージ内で, 発情前期から発情期にかけて偽手 術を施した雌マウスを, または卵巣摘出した雌マウスを, 実験開始の 24 時間 前から飼育した。実験の直前に 10 g の床敷きを採取し, 匂いの提示に用いた。 床敷きの匂いに対する雄の嗜好性の測定 第 1 章で用いたタイプ 1 の測定装置を用いて, 18 時から 23 時までの暗期に 弱赤色光下で行った。同一の雄を 1 日 1 回 3 日間にわたり測定に用いた。毎日 の測定では, 被験雄を中央の部屋入れて 30 分間環境に慣らした後, 風上の部 屋に 10 g の床敷きを入れ, 風上の窓にタッチする時間を 30 分間測定した。た だし, 1 日目の測定は床敷きの匂いを提示せずに行った。2 日目と 3 日目の測 定では, 発情期の雌が居住するケージの床敷き, または卵巣摘出した雌が居住 するケージの床敷きを提示した。2 日目の測定で発情期の雌が居住するケージ内 の床敷きを提示した場合は, 3 日目に卵巣摘出した雌が居住するケージの床敷 きを提示した。一方, 2 日目の測定で卵巣摘出した雌が居住するケージの床敷 きを提示した場合は, 3 日目に発情期の雌が居住するケージ内の床敷きを提示 した。 雄マウスへの匂いの提示 すべての実験は 18 時から 23 時までの暗期に弱赤色光下で行った。第 1 章で
用いたタイプ 1 の測定装置の中央の部屋に 30 分間被験雄を入れた後, 発情期の 雌, 卵巣摘出した雌, 雄, 発情前期から発情期にかけての雌が 24 時間居住し ているケージ内の床敷き 10 g のいずれかを風上の部屋に入れ, 60 分間匂いを 提示した。匂いを提示しない場合は, 中央の部屋に 30 分間被験雄を入れた後, 風上の部屋に匂いの源を入れず, 同様の手順で行った。発情前期から発情期に かけて雌が 24 時間居住しているケージ内の床敷きを, 直接接触できる条件下 で提示する場合は, 被験雄が居住するケージ内に 10 g の床敷きを入れた。匂 いの提示から 60 分後に, ジエチルエーテル麻酔下で被験雄の脳を採取し, パ ラホルムアルデヒドで固定した。 免疫染色 第 2 章に示す方法に従った。 鋤鼻器摘出手術 第 2 章で示す方法に従った。 Habituation-dishabituation 試験 Pankevich ら の 方 法 (53) に 一 部 変 更 を 加 え , habituation-dishabituation 試験を行った。発情前期から発情期にかけて 雌マウスが 24 時間居住しているケージ内の床敷き 10g, 発情期の雌マウス, 卵巣摘出した雌マウス, 雄マウスを匂いの源として用いた。匂いの源を入れ替 えた後に匂いが風上の部屋に残ることを防ぐため, 匂いの提示には円筒状のプ ラスチック製容器(直径 100 mm, 高さ 110 mm, Fig.21b)を用いた。容器の 天井を金網で覆い, 側壁の高さ 5 cm の位置に直径 5 mm の穴を 12 個開け, 容 器内の匂いが外に漏れるようにした。まず, タイプ 1 の測定装置の中央の部屋 に被験雄を入れ, 30 分間環境に慣らした。その後, 匂いの源の提示に用いる容 器のみを風上の部屋に入れ 2 分間提示した。その後, 容器を取り出して 1 分間 匂いのない状態にした後, 再び容器のみを風上の部屋に入れ 2 分間提示した。 更に, 容器を取り出して 1 分間匂いのない状態にし, 再び容器のみを風上の部 屋に入れて 2 分間提示した。次に容器を取り出した 1 分後に, 1 つ目の匂いの 源を容器に入れ, 同様な手順で 3 回の匂いの提示を行った。さらに容器を取り 出し1分後に, 2 つ目の匂いの源を容器に入れ, 同様な手順で 3 回の匂いの提 示を行った。空の容器の提示も含めた合計 9 回の匂いの提示の間に風上の窓に タッチする時間を測定した。1 組の匂いに関して, 提示する順番の影響を考慮 し, 提示する順番を逆にして, 同一の被験雄に対して別の日に同様の実験を行 った。2 回の測定の平均値をその雄のデータとして用いた。
統計処理 雌が居住するケージ内からの床敷きの匂いに対する雄の嗜好性を評価するた め, バートレット検定の後, 繰り返しのある一元配置分散分析法を用いた。ポ ストホック検定にはターキークレーマ検定を用いた。匂いを提示した雄の副嗅 球前部, 後部における c-Fos 陽性細胞数を比較するため, バートレット検定 の後, 一元配置分散分析法を用いた。ポストホック検定にはターキークレーマ 検定を用いた。各群内の副嗅球前部と後部の間での c-Fos 陽性細胞の数の比較 には, 対応のある t 検定を用いた。Habituation-dishabituation 試験の 評価には, 対応のある t 検定を用いた。危険率 5%以下で有意差ありとみなし た。