侵入害虫マメハモグリバエに対する生物的防除資材としての土着寄生蜂類の利用技術に関する研究
103
0
0
全文
(2) (和文題目) 侵入害虫マメハモグリバエに対する生物的防除資 としての土着寄生蜂類の利用技術に関する研究. 近畿大学大学院. 農学研究科. 農学専攻 本藤智雄. (杉本毅教授). (英文題目) Us eofNa t i v eS p e c i e so fE u l o p h i dP a r a s i t o i d sa sThe B i o l o g i c a lC o n t r o lAgentA g a i n s tL i r i o m y z at r i f o l i i. TomooHondo. March, 2006. G r a d u a t eS c h o o l, K i n k iU n i v e r s i t y D i v i s i o nofA g r i c u l t u a lS c i e n c e M a j o r :Entomology ( A d v i s o r :P r o 王T u y o s iS u g i m o t o ).
(3) 目次. 第 1章 緒 言 ・ ・. 第 2章 第 l節. マメハモグリバエ及び寄生蜂類・・・・・・・・・・・・・・・・・ 5 マメハモグリバェ・・・・・・・・・・・・・・・・・・・・・・ 5. 分布 形態及び生態 本種による被害 殺虫剤抵抗性 第 2節. ハモグリパエ寄生蜂類・・・・・・・・・・・・・・・・・・・・ 9. 寄生戦略 繁殖 繁寄主体液摂取 ハモグリパエ類殺傷寄生蜂の寄主利用戦略 ハモグリバエ類寄生蜂の寄主探索. 第 3章. マメハモグリバエに対する天敵資材としての土着寄生蜂 7種の温度耐. 性 の 比 較 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 1 2 EE-A. 今4. ••. -. 第 2節 実 験 材 料 と 方 法 ・ ・. •••. ・. 第 1節 は じ め に ・ ・. • 1 4. マメハモグリバエ類 寄生蜂類 実験装置 温度と発育の関係、 雌成虫の寿命,寄生及び寄主体液摂取に及ぼす温度の影響 第 3節. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・ 1 6.
(4) 温度と発育の関係、 雌成虫の寿命,寄生及び寄主体液摂取に及ぼす温度の影響 第 4節. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・ 2 2. 土着寄生蜂の温度耐性 放飼と大量生産の各段階における捕食寄生蜂の有効性の総合 的な評価. 第 4章. ハモグリミドリヒメコバチの大量増殖法の確立・・・・・・・・・ 27. 第 1節. はじめに・・・・・・・・・・・・・・・・・・・・・・・・・ 27. 第 2節. 実験の準備・・・・・・・・・・・・・・・・・・・・・・・・ 27. 大量生産工程設計の概要 産卵装置の設計 羽化成虫回収装置の設計 梱包資材 第 3節. 実験方法・・・・・・・・・・・・・・・・・・・・・・・・・ 37. 産卵装置における成虫回収 羽化成虫回収装置における成虫回収率 剤型・保蔵が寄生蜂におよぼす影響 最適放飼密度の決定 第 4節. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・ 3 9. 産卵装置における成虫回収 羽化成虫回収装置における成虫回収率 剤型・保蔵が寄生蜂におよぼす影響 最適放飼密度の決定 第 5節. 第 5章. 考察・・・・・・・・・・・・・・・・・・・・・・・・・・・ 44. ハモグリミドリヒメコバチの生物的防除資材としての有効性とバンカ.
(5) 一プラント法の検討・. • 48. 第 l節 は じ め に ・ ・. • 48. 第 2節 材 料 と 方 法 ・. • 49. 供試植物 供試昆虫. 実験の準備 マメハモグリバエ幼虫高密度時における寄生蜂放飼の防除効果 マメハモグリバエ幼虫低密度時における寄生蜂放飼の防除効果 ハモグリミドリヒメコパチの寄主選好性の検討. 第 3節. 結果・・・・・・・・・・・・・・・・・・・・・・・・・・・ 55. マメハモグリバエ幼虫高密度時における寄生蜂放飼の防除効果 マメハモグリバエ幼虫低密度時における寄生蜂種放飼の防除効果 ハモグリミドリヒメコバチの寄主選好性の検討. 第 4節.考察・・・・. .6 0. ハモグリミドリヒメコバチの放飼密度および放飼時期 代替寄主キツネノボタンハモグリバエ,バンカー植物ラナンキュラスを用 いたパンカー植物法の可能性. 第 6章. 総合考察・・・・・・・・・・・・・・・・・・・・・・・・・・ 6 4. 謝 辞 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 75. 要 約 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 76. Summary・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 79. 引 用 文 献 ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ 82.
(6) 第 1章 緒 言. わが国における施設栽培は 1 9 6 5年には栽培延面積 6387haだ、ったが,その後. 9 9 9年では 70525haと 1 0倍を超え,現在,わが国の園芸にお 急速に拡大し, 1 いて重要な位置を占めている((社)日本施設園芸協会, h t t p : / / w w w . j g h a . c o m l ) . これら施設栽培を含めた戦後の農業技術の向上は,作物の品種改良とともに合 成農薬の急速な発達に負うところが大きかった.しかし,合成農薬の大量使用. C a r s o n, により,生態系へのリスクおよび人間や家畜への健康被害がもたらされ ( 1 9 6 2 ; 根本, 1 9 9 7 ),また,誘導多発生(リサージェンス)や天敵の減少した圃 場に新たに害虫が侵入し. v a nD r i e s c h eand 大発生をしばしば引き起こした (. 9 9 6 ).さらに,害虫の薬剤抵抗性の獲得により従来の薬剤がほとんど B e l l o w s,1 9 9 8 a, b ) . 近年の交 効果を示さない殺虫剤抵抗性害虫系統が出現した (WHO, 1 通網の整備や航空輸送など流通技術の進歩により農産物の物流が飛躍的に高速 化かっ広域化したため,輸送農産物と共に薬剤抵抗性を獲得した害虫も急速に. u r g e s sも , 拡散した.本研究で取り上げるマメハモグリバエ L i r i o m y z at r i f o l i iB 1 9 9 0年にわが国に侵入する以前に,既に強度の殺虫剤抵抗性を獲得していたた め化学的防除が非常に困難で、あった. 近年,マメハモグリバエのように高度の薬剤抵抗性害虫の出現,環境保全, 食の安全を求める消費者ニーズ、の高まりなどを受けて,合成農薬に対する過度 の依存には限界があり,その打開策のーっとして天敵を用いた生物的防除が注 目を集めるようになった. 天敵とは他の生物から栄養を摂取して生活し,その生物を死亡させたり,繁 殖能力を低下させたりする捕食者,寄生者,病原微生物を指す(広瀬, 1979; 矢野, 1 9 9 5 ) . 天敵昆虫は一般的に栄養摂取の過程で他の昆虫を殺害するので, 自然界において昆虫個体群の密度制御の重要な担い手である.天敵昆虫は,人 為的に制御したり,利用したりし易かったので,古来から害虫防除に利用され てきた.古くは l世紀頃に中国で柑橘害虫を駆除するためにツムギアリを利用.
(7) したとし寸記録もあるが(斉藤ら, 1 9 8 6 ),近代における生物的防除の画期的な. 8 8 8年,カリフォルニアにおけるベダリアテントウによるイ 成功例としては, 1 セリアカイガラムシ防除の成功が有名であり,これを機に世界的に天敵利用が 試みられた ( Debach,1 9 6 4 ).わが国においては,ベダリアテントウによるイセ リアカイガラムシ防除,シルベストリコバチによるミカントゲコナジラミ防除, ワタムシヤドリコバチによるリンゴワタムシの防除(安松, 1970;森・村上,. 1 9 8 1;広瀬, 1 9 8 7 ),さらに近年においてはヤノネカイガラムシ ( F u r u h a s h iand 9 8 3;古橋・西野, 1984;高木, 1983;高木・氏家, 1 9 8 6 ) やクリタ N i s h i n o,1 M o r i y ae ta . l1 9 8 9,1 9 9 0;村上, 1 9 9 7 ) などで成功を収めた.これらの マパチ ( C l a s s i c a lb i o l o g i c a l 天敵の利用方法は永続的利用法または伝統的生物的防除法 ( c o n t r o O とよばれ,主に果樹を中心とした永年作物の侵入固着害虫に対して効 C l a u s e n, 1978;Debacha n dRosen,1 9 9 1 ),土着害虫や周期的に 果を上げたが ( 作物が除去されることにより生息環境が大きく変動する l年生作物ではあまり 効果が得られなかった. しかし,近年,生物的防除法は急速に改善が進み,下 記の 2タイプに大別される.第 1は,害虫の発生時に,直接的,即時的防除効 果を期待して天敵を大量放飼する大量放飼法一生物農薬的利用法とよばれてい る.第 2は,比較的少量の天敵を放飼して後世代における持続的防除効果を期 待する接種的放飼法である.どちらも化学農薬のように害虫の発生時に合わせ て天敵を逐次放飼するので,土着天敵のいない,あるいは低密度のためにそれ らによる防除が期待できない施設閏場,永続的利用法ではあまり効果が得られ なかった一年生作物閏場においても有効であった.天敵放飼の例としては,鱗 題目害虫に対するタマゴバチ類 ( L i,1 9 9 4 ;S m i t h,1 9 9 6 ),ハダニ類に対するチリ カブリダニ(森・真梶, 1977;井上, 1993;森 , 1993;江原・真梶, 1 9 9 6 ),オ ンシツコナジラミに対するオンシツツヤコパチ(矢野, 1 9 7 9,1 9 8 8 ),アザミウ. 9 9 4 :R i u d a v e t s,1 9 9 5; Yano,1 9 9 6 ;C a s t a n e マ類に対するヒメハナカメムシ(永井, 1 e ta , . l 1 9 9 9 ),アブラムシ類に対するコレマンアブラバチ(山下・藤富, 1 9 9 8 ), そして,強度の殺虫剤抵抗性を獲得し,化学的防除が困難であったマメハモグ. 2.
(8) リバエに対してもイサエアヒメコバチとハモグリコマユバチといったヒメコバ チ科や,コマユパチ科の寄生蜂が実用化されている ( M i n k e n b e r ga n dvan. 9 9 0 ;P a r r e l l ae ta , . 11 9 8 7 ;H e i n t ze ta , . 11 9 9 0 ; 小津, 2001 )• L e n t e r e n,1 こうした天敵昆虫の利用には生きた昆虫そのものを利用するが故の技術的な 困難を伴うが,反面,害虫が天敵に対して抵抗性を獲得する危険性が極めて低 いこと(安松, 1 9 7 0 ),合成農薬のような人体,特に薬剤散布者の健康への影響 が無いこと,天敵成虫のボトル梱包や踊を貼り付けたカード等,軽量な剤型梱 包が可能で,省力的化につながることなど多くの利点がある.また欧米におい て天敵生産を専門に扱う企業が現れ,天敵を商品化して販売を始めたことによ り,天敵昆虫が生物農薬として安定して供給されるようになったことも天敵普 及のきっかけとなった.ところで,オーストラリア,タスマニア島において送 粉見虫として導入されたセイヨウオオマルハナパチが野生化し急速に拡大した. ( H i n g s t o n,1 9 9 7 ) . わが国においても,施設内の送粉昆虫として放飼した同種 9 9 7 a, b ),在来種との聞に雑種が が野外へ分散,定着し(保全生態学研究会, 1 M i t s u h a t a& Ono,1 9 9 6 ) . 導入天敵についても対象害虫だ 形成されるに至った ( けでなく,対象外の土着生物をも攻撃し,絶滅または絶滅寸前に追い込んでい. H o w a r t h,1 9 9 1 ),天敵も含めた外来昆虫導入にともなう固 ることが指摘され ( 有生態系へのリスクが世界的関心事となった ( H i r o s e,1 9 9 4,1 9 9 9;日本生態学. e n t e r e n ,e ta 1 .2 0 0 3 ) . 会 , 2002;vanL F A0 ) (http://www.fao.orglindex_en.htm)は「外来 1 9 9 5年,国連食糧農業機関 ( の生物的防除素材の輸入と放飼のための取り扱い規約」を出し,各国政府に遵 守を求めた.わが国においても, 1 9 9 9年 3月に環境庁水質保全局が環境影響評 価ガイドラインを示し(環境庁水質保全局, 1 9 9 9 ),土着天敵を含め,天敵昆虫 の導入に際して環境影響調査が行われる事となった.こうした背景から,導入 天敵に代わり,地域個体群聞の交雑など一定の環境リスクはあるものの,導入 天敵と比べリスクの低い土着天敵を用いた防除法確立の必要性が高まっている. そこで,本研究で、は侵入害虫マメハモグリパエに対する生物的防除資材とし. 3.
(9) て導入されているイサエアヒメコパチ導入系統と日本未記録である(小西,. 1 9 9 8 ) ハモグリコマユパチに代わる防除資材としての土着寄生蜂の選抜とその 大量生産技術の確立および施設内放飼のための基礎資料を得ることを試みた. 初めに,現在わが国の施設内において,マメハモグリバエに寄生することが確 認された土着寄生蜂 4科 , 27種より(小西, 1 9 9 8 ),西日本のハモグリバエ寄 生蜂群において優占的なヒメコバチ科寄生蜂類のうち 7種を選び,それらの温 度特性を明らかにした後,その温度特性に基づいて温度変化の激しい西日本に 最適な寄生蜂種を選抜した.続いて,選抜種の大量増殖のための技術確立を行 った.最後に,選抜した寄生蜂を実際に温室に放飼して,防除効果を検討した. さらに,近年生物的防除の補強法として期待されているバンカープラント法に ついて多少検討を試みた.. 4.
(10) 第 2章 マ メ ハ モ グ リ パ エ 及 び 寄 生 蜂 類. 第 1節 マ メ ハ モ グ リ パ エ i n e r )は,幼虫が植物の柔組織内や表皮組織内に潜り食害するも 潜葉虫 (Leafm H e r i n g,1 9 5 1 ),こ のの,表皮組織や外壁を無傷で残す見虫のグループであり (. れら潜葉虫は分類学上,鱗麹目,双麹目,甲虫日,膜麹目の 4つの目に属して いる.ハモグリバエ類 (Agromyzidae)は,幼虫は主に葉や茎に潜孔するが,中に S p e n c e r ,1 9 9 0 ).こ は茎に穿入するものや根や花頭を食害するものも含まれる (. れらの分布域は広く,グリーンランド北部から南米末端,ニュージーランドの南 極に近い島々にまで分布し,それらの寄主植物はゼニゴケ類から被子植物に及 6 0科と広範に及ぶものの,寄主特異性が極 び,現存 380科の被子植物のうち 1 6種のみである ( S p e n c e 巳1 9 9 0 ) . めて発達しているため真の広食性は 1. 日本において 200種以上のハモグリパエが確認されており (Sasakawa,1 9 6 1 ), これらの幼虫が葉に潜ることによって生じる食害痕のくねくねとした形状から, 日本では俗にエカキムシと呼ばれている.これらのハモグリパエ類は,農業上 問題となる種も少なくなく(笹川, 1966a, b ),マメハモグリバエ L i r i o m y z at r i f o l i i ( B u r g e s s )はこうした潜葉性害虫の l種である.. 分布. 本種の起源はフロリダ地方を含む北アメリカ東部やカリブ海諸国とさ. れている ( S p e n c e r1 9 6 5,1 9 7 3,1 9 9 0 ) . 近年,航空機などの輸送手段の急速な発 達によって,植物の輸送が高速かっ世界的な規模に広域化されたことにより, 本種に寄生された植物が北アメリカ大陸東部から輸送され,本種はアメリカ国 P a r r e l l ae ta l .1 9 8 1, Trumble1 9 81),さらにはアフリカ,ヨーロッパそし 内全域に (. てアジアへと伝搬し,その生息範囲を拡大していった (Minkenbergandvan L e n t e r e n1986, Minkenberg,1 9 8 8 ) .. 本種については, 日本では 1949年に北海道のマメ科植物からの採集報告があ ったが (Sasakawa1 9 6 1 ),後に L .c ong 目 , 的 B eckerと訂正された (Sasakawa1 9 9 2 ).. 5.
(11) 本種は 1990年に静岡県と愛知県で初めて侵入が確認さ れ ( 西東, 1992a),その 後,加害植物 の流通に伴い急速に分布域を拡大 し,全国的に発生が確認される に 至 っ た (小津 2000).. 形態及び生態. マメハモグリバエ成虫の体長は約 2mmで,雌がやや大きい .. 複眼はプラ ム レッド,頭部,胸部側板および脚の大部分が黄色で,その他 の部分 Minkenberg,1 9 9 0) ( 図 1).雌成虫は腹部に発達した産卵管を持 は黒色である ( . 1 5" " 'O. 3m mの穴を開け,内部組織にゼリー状で半透明の卵 ち,葉の表皮に 0 ( 0 . 2 3xO. 12mm) を産卵し,その穴よりにじみ出た汁液をしばしば摂食する ( Spe n c e r,1 9 7 3). その際に生じた穴は,葉に白い斑点状の痕として残る(図 2).. 図1.マメハモグリパエ雌成虫 雌は摂食または産卵行動のために l日に 100回以上も葉に穴を開けるが,それ ら痕跡 の多くは摂食痕である ( P a r r e l l ae ta . l1 9 8 3,P a r r e l l a1984,S m i t hand Hardman1 9 8 6).雄は産卵管を持たないため単独で摂食す ることはできず,雌成. 虫が作った摂食, 産卵痕を利用して吸汁すると思われる.成虫は,数時間で 100m 程度の移動能力がある ( vandeV r i eandD i r k s e1 9 8 2).本種の活動のピークは 1 2: 00" " '1 6: 00であり,夜間にはほぼ活動を休止する ( C h a n d l e r1 9 8 5).また,成 Z e h n d e randTrumble,1984), この性質を利 虫は黄色に誘 引 される特性があり (. 6.
(12) 用して発生調査法として黄色粘着 トラップが使用される(小津ら, 1999b) . 本種幼虫は淡い黄色をした無脚のワジであり, ~I降、化 後は葉内に潜り鎌状の口 鈎で内部組織を摂食し,この際にくねくねとした食害痕が葉に残る(図 2 ).糞 は孔道内に連なり ,幼虫が体の向きを変えるたびに糞粒は側面に沿って交互に 2列になる .幼虫は 3齢期で成熟し,体長 3mm程度に成長する .成熟した幼虫. は葉の表皮を破って脱出し,葉上や土の表面や隙聞に潜り,数時間後に 2mm 程度の囲踊となる ( S p e n c e r ,1 9 7 3;西東, 1 9 9 2 a,1 9 9 2 b) .本種の発育は,温度や寄 9 9 5 a;小津ら, 1 9 9 9 a),発育零点はお 主植物種により異なるものの(西東ら, 1 0C前後であり, 3 5o C以上では高温障害が生じる(西東, 1992b, 1994). よそ 1 0. また,地域個体群によって発育零点,有効積算温度が多少異なる(西東ら, 1 9 9 5 a;小津, 2 0 0 1 ;坂巻ら, 2003;徳丸 ・阿部, 20 0 3).. マメハモグリパエ幼虫 雌成虫による産卵または 摂食痕. 図2 .マメハモグリパエ雌成虫による産卵および摂食痕と,幼虫による摂食痕. 7.
(13) 本種は,海外では 2 1科 1 2 0種以上の植物に寄生することが確認さており. ( M i n k e n b e r gandvanL e n t e r e n, 1 9 8 6 ), 日本においてもマメ科,ウリ科,ナス科,キク 2科 50種以上の植物で寄生が確認されているが(西 科,セリ科,アブラナ科など 1 9 9 7 a ),産卵数は寄主作物種により大きく異なる(西東ら, 1995a;小津ら, 東1 1 9 9 9 a;小津, 2001 ) .. 本種による被害. 本種による被害は,成虫による摂食・産卵,および幼虫によ. る摂食により引き起こされる.セルリーやチンゲンサイなどの葉菜類,キク, ガーベラなどの花井作物は,産卵痕および食害痕が生じただけで商品価値が低 下する.特に花井類は深刻で,作物に僅かな摂食痕や産卵痕が生じただけでそ の商品価値は著しく低下する.また,幼虫によって葉内部組織が摂食されると 光合成が阻害され(Jo n s o ne ta , . 11 9 8 3 ) 植物体の生育不良が起こり,その結果,. T r u m b l ee ta , . 11 9 8 5, 西野・内田 収量の減少や収穫期の遅延を引き起こす ( 1 9 9 9 ).幼虫による食害が深刻な場合には,表皮組織だけを残して葉の内部組織 が食い尽くされ,植物体が枯死することもある.また,わが国では確認されて. i c h o r i iを媒介したり, いないものの,成虫がキクの斑点細菌病菌 Pseudomonasc P r i c ea n d 成虫の摂食痕,産卵痕がその感染口となっているとの報告もある ( 9 8 1 ;M a t e o n ia n dB r o a d b e n t,1 9 8 8 ;B r o a d b e n ta n dM a t t e o n i,1 9 9 0 ) . Harbaugh,1. 殺虫剤抵抗性. 1 9 4 0年代には L i r i o m y z α属が殺虫剤抵抗性を獲得することが確. 認され (Genung,1 9 5 7 ;W o l f e n b a r g e r ,1 9 5 8 ),その中でも本種の殺虫剤抵抗性の. P a r r e l l aandK e i l,1 9 8 4 ;P a r r e l l a,1 9 8 7 ),ペルメトリンに対する感 発達速度は高く ( 受性が 2年で 20倍に低下した事例もある ( P a r r e l l aa n dK e i l,1 9 8 4 ).日本に侵入 したマメハモグリバエについても高い殺虫剤抵抗性が確認され,侵入後合成ピ レスロイド剤によるリサージェンスが頻発した(西東ら, 1 9 9 2 a, b ; 西東ら, 1 9 9 3,. 1 9 9 6 ) .. 8.
(14) 第 2節 ハ モ グ リ パ エ 寄 生 蜂 類 p a r a s i t o i d) には,世界で双麹目 植食性昆虫の重要天敵としての捕食寄生者 ( ( 8科,約 14925種以上),鞘麹目 ( 2科 , 2400種),撚麹目(10種),脈麹目 ( 50. 種)など多岐にわたるが ( God合ay ,1994)3種数,個体数共に最も多いのは, 1 3 上科, 46科にわたり 1 5万種をはるかに超える膜麹目寄生蜂類である ( L a S a l l e 9 9 1).日本の土着ノ¥モ グ リパエ類も多種にわたる寄生蜂の攻撃にさ andGauld,1. らされており ( TakadaandKamijo, 1979,Sugimotoe ta . l,1983a,Kato,1994),侵入 国虫マメハモグリパエの生物防除資材として大きな関心が寄せ られている.. 寄生戦略. As kewandShaw( 1 9 8 6 )によると,寄主利用戦略に基づき,寄生蜂を. 2つのカテゴリ ー に大別できる.すなわち,寄主を生かしたまま寄生し,発育 Koinobiont) と,害主に産卵管を挿入 を完了すると寄主 を殺す飼殺し寄生種 (. して毒液を注入して殺害したり,永久麻庫させて寄生する殺傷寄生種 ( I d i o b i o n t) である.また,苓生様式は,寄主の体内に産卵する内部寄生 ( e n d o p a r a s i t i s m ) ( 図 3a) と害主の体表面やその周辺に産卵する 外部寄 ( e ct p a r a s i t i s m)( 図 3b)に分けられ,さらに l寄主 に l卵を産む単寄生と ( s o l i t a r y g r e g a r i o u sp a r as i t i s m), 寄主自体に寄生を行 p a r a s i t i s m),複数卵を生む多寄生 (. う一次寄生 ( primaryp a r a s i t i s m) と寄主に苓生している寄生者に寄生する高次 hyperp a r a si t i s m) に分類される . さらに,寄生対象となる寄主の発育段 寄生 (. . 内部寄生 ( a ) と外部寄生 ( b ) 図3. 9.
(15) 階により,卵寄生 ( e g gp a r a s i t i s m ),幼虫寄生(Ia r v a lp a r a s i t i s m ),踊寄生 ( p u p a l p a r a s i t i s m ),卵・幼虫寄生 ( e g g l a r v a lp a r a s i t i z u m ) や幼虫・踊寄生 O a r v a l p u p a l p a r a s i t i s m ) などに分類される.こうした寄生様式の多様性は,同一寄主種を攻. 撃する複数の寄生蜂の種間競争を緩和していると思われる.. 繁殖. 寄生蜂を含む膜麹目は,原則として半数倍数性 ( h a p l o d i p l o i d y ) といっ. た特殊な性決定様式を持つ.膜麹目では,雄が未受精卵から発生するので半数 体であり,その精子は,体細胞突然変異を除き全て遺伝的に同一で,雄ゲノム. 1 2を持つ. を全て持っている.一方,雌は倍数体であり,各卵子は雌ゲノムの 1 母蜂は,交尾によって得た精子を受精嚢に貯蔵し,産卵時に精子を少しずつ出 して受精させる.受精の成否は母蜂次第であり,母蜂は雌雄を産み分けること o l t o n,1 9 8 8;Godfray ,1 9 4 4 ) . 未受精卵から雄が生 ができる(例えば GauldandB a r r h e n o t o k y ) と呼ぶ.一方,主に生殖細胞がリケ まれる現象を産雄単為生殖 (. ッ チ ア 肋l b a c h i aに感染された寄生蜂では,未受精卵からも雌が生じる,いわ t h e l y t o k y ) を行う .W o l b a c h i aは無脊椎動物の精巣や卵巣か ゆる産雌単為生殖 (. ら発見され,感染種には天敵も多く含まれている(例えば, S t e e v e n sa n dWade, 1 9 9 0 ;S t o u t h a m e randLuck ,1 9 9 3 ;Werren,1 9 9 7 ) .. 寄主体液摂取. 寄生蜂類の中には,雌成虫が,寄主の排推する甘露,花粉,蜜. などを餌とする他に,産卵管や口器を用いて寄主体壁に孔をあけ,体液を摂取 する種がし 1る.この行動は寄主体液摂取 ( h o s t -f e e d i n g ) と呼ばれ,特に幼虫寄 生蜂ではよく見られる現象であり ( C l a u s e n,1 9 4 0 ),1 8科 1 4 2種の寄生蜂で寄主 体液摂取行動が確認されている(Je r v i sandKidd,1 9 8 6 ).寄主体液摂取は雌成虫 p r o o v i g e n i c ) のみが行い,卵形成が産卵開始に先行して完了する斉一成熟性 (. 寄生蜂では寄主体液摂取はほとんど見られないが,卵形成が産卵と平行して行 われる逐次成熟性 ( s y n o v i g e n i c ) 寄生蜂ではよく見られ,主に卵形成のための タンパク質確保 L 雌成虫のエネルギー補充のために行われると考えられてい 1 0.
(16) る ( F r a n d e r s,1 9 3 5 ;Chana n dG o d f r a y ,1 9 9 3 ) . さらに,後者についてみると,ヒ メコバチ科寄生蜂 C h r y s o c h a r i sp e n t h e u s( W a l k e r )の羽化直後の雌成虫に 3日間寄 主を与えた後,蜂蜜または水のみを与えて飼育すると,卵巣内の成熟卵はいっ たん増加した後減少したことから,タンパクの供給が断たれると,いったん形. S u g i m o t oe ta , . l 成した卵を分解,再吸収し,生存のためのエネルギー源にする ( 1 9 8 3 b ) . 卵再吸収は体液中のタンパク量の低下が引き金となっておこる ( K i n g a n dR i c h a r d s, 1 9 6 8 ) .. ハモグリパエ類殺傷寄生蜂の寄主利用戦略. スイカズラハモグリバエ野外個. 体群の調査において,ヒメコバチ類の寄生により死亡した寄主のうち,大型. K a t o,1 9 8 5 ).また,c. 寄主から雌蜂が,小型寄主から雄蜂が多く羽化した ( p e n t h e u sを用いた実験において,小型の若齢寄主を寄主体液摂取に,大型の寄 S u g i m o t oa n dI s h i i,1 9 7 9 ). 主を産卵に使い分けることが確認された (. ハモグリパエ類寄生蜂の寄主探索. 寄主であるハモグリパエ幼虫は生息場所. に連続的に存在しているのではなく,葉や株を単位(パッチ)として不連続に. u g i m o t oe ta l( 1986,1 9 8 7, 1 9 8 8a , b, c ) は,不連続的に分布する 分布している. S a p s i l a r t h r ar u f i v e n t r i s( N e e s )を 寄主の効率的な探索行動についてコマユバチ科 D 用いて検討した.その結果,寄生蜂は単一の手がかりによって寄主探索を行う のではなく,まず匂い物質を,続いて寄主幼虫の摂食によって生じた食害痕, さらに寄主幼虫が植物をかじる際に生じる摂食音を順次手がかりにして寄主探 索を効率的に行うことが明らかにされた.さらにその探索時間は寄主密度が高 いほど長くなり,雌蜂が寄主密度に比例的に探索時間を配分する最適戦略を採. S u g i m o t oe ta , . l1 9 8 7 ). っている (. 1 1.
(17) 第 3章. マメハモグリパエに対する天敵資材としての土着寄生蜂 7種の温度耐. 性の比較. 第1 節. はじめに. マメハモグリバエは,わが国に侵入する前に欧米において既に高い薬剤抵抗 性を獲得しており,侵入後リサージェンスがしばしば発生し,化学農薬による防 除には問題があることが明らかになった(第 2章).現在,. 日本では,本種の生物. αs i b i r i c a 的防除を目的としてヨーロツノ《からハモグリコマユバチ Dacnus. T e l e n g a とイサエアヒメコバチ Dig か phusi s a e a( W a l k e r )の 2種寄生蜂が輸入さ 9 9 7年 1 2月に農薬登録され,利用されている(小津, 1 9 9 9 ;小津ら, 1999c; れ , 1 西東ら, 1995b; 市川ら, 1996; 柴尾ら, 1996; 松村ら, 2001).一方,. S a s a k a w a ,1 9 6 1 ),研 我が国には 200種以上のハモグリバエ類が生息しているが ( T a k a d aandKamijo, 1 9 7 9 ),キツネノボタン 究,調査の進んでいるナモグリパエ ( S u g i m o t oe ta , . l1 9 8 3 a ),スイカズラハモグリパエ ( K a t o,1 9 9 4 )は ハモグリバエ ( 多種の寄生蜂類の攻撃にさらされていることが実証された.マメハモグリバエ の侵入後しばらくして行われたハウス内の調査によると,土着ハモグリバエ類 の寄生蜂種として記録されている 4科 27種が寄生することが確認された(小西. 1 9 9 8 ) . 本章後半で述べるが,西日本の温室は外気温と比べ年間を通じて激しく 変動するため,生物的防除資材にはその温度変化への耐性が求められる.そこ で本章では,西日本の土着ノ¥モグリバエ寄生蜂群において優勢なヒメコバチ科. tal . , 1983a;Kato, 寄生蜂類 (TakadaandKamijo,1979; Sugimotoe 1994; 1996a, b ) より 7種を選び ( T a b l e1 , F i g .4 ),それらの発育,雌成虫の寿 命,繁殖能力に対する温度耐性を明らかにした.さらにその結果をもとにマメ ハモグリバエの生物的防除資材として最適な寄生蜂を選抜するために,それら の防除効率を包括的に評価した.ところで,本実験に用いたヒメコバチ科寄生 蜂は,殺傷寄生,単寄生性の寄生様式をとる(第 2章).また E u l o p h i n a eの R. k a t o n i sI s h i i, H .v a r i c o r n i s, D .i s a e a,D .m i n o e u s, D .p u s z t e n s i sは,殺害した寄主幼 1 2.
(18) b. F i g.4.E u l o p h i dp a r a s i t o i d se x a m i n e di nt h i ss 同d y .a ,P k a t o n i s ;b,H v a r i c o r n i s ;c ,D. i s a e a;d, D.minoe u s;e , D. pus z t e n s i s; 仁C.p e n t he u s; g, N formosa( c f . T a b l e1 ). 1 3.
(19) 虫の表皮上やその周辺に産卵する外部寄生蜂であり, E n t e d o n t i n a eの仁 p e n t h e u s,. N e o c h r y s o c h a r i sformosaは寄主体内に産卵する内部寄生蜂である.また,本研 究で用いた寄生蜂は,. o l b a c h i aに感染されて産雌単為生殖を行う リケッチア W. Nformosa( A r a k a k ia n dK i 吋0,1 9 9 8 )を除き,全て半数倍数性のため産雄単為生殖 を行う(第 2章)•. T a b l e1 .L i s tofe u l o p h i dp a r a s i t o i d sexaminedi nt h i ss t u d y Bodyl e n g t h P a r a s i t i z a t i o n (mm). S p e c i e s E u l o p h i n a e P n i g a l i ok a t o n i sI s h i i Heη m 1 科 申 { 伊 p α t ω ,r s e n Di恕 'g~か yphu ω IS i s αeα(Wa 叫l k e r )(イサエアヒメコノバ〈チ) n a t i v es t r a i n(土着系統) i n t r o d u c e ds t r a i n(導入系統) D .minoeus( W a l k e r ) D .p u s z t e n s i s( E r d o s& N o v i c k y ). -1 .7 1 .3 1 .1 ・ 1 .9. E c t E c t. 0 . 9 -1 .6. E c t. 1 .0 -1 .6 0 . 9・ 1 .5. E c t E c t. E n t e d o n t i n a e C h r y s o c h a r i sp e n t h e u s( W a l k e r ) 0 . 8・1 . 5 N e o c h r y s o c h a r i sformos α(Westwood) 0 . 8・ 1 .6 (ノ¥モグリミドリヒメコノミチ). End End. aE ct :E c t o p a r a s i t o i d, E n d :E n d o p a r a s i t o i d. 第 2節 実 験 材 料 と 方 法. 14LlOD に維持した近畿大学農学部昆虫飼育 飼育および実験は全て, 70%RH, 室内で行い,温度は必要に応じて種々に設定した.. マメハモグリパエ類. 9 9 2年に マメハモグリパエは,静岡県農業試験場から 1. 譲り受け,恒温室内 ( 2 50C)で,インゲンマメ P h a s e o l u sv u l g a r i s( 品種:つるなし すじなし黒菜豆)の初生葉を用いて累代飼育し,実験に供試した. 寄生蜂類. H .v a r i c o r n i sと Nformosa以外の寄生蜂については, 1 9 9 8から 2 0 0 0. 年の春に近畿大学農学部実験圃場に栽培されたエンドウ PisumsativumL .か. 1 4.
(20) ら寄生蜂に寄生されたナモグリパエ Chromatomyi ahorticola(Goureau) 幼 虫を含んだ葉を採集し,実験室に持ち帰り羽化させた • H .v a r i c o r n i sは ,. 1996. に静岡県農業試験場より成虫を譲り受けた • N .f ormosaは , 1998年に鹿児島県. 農業試験場より寄生されたマメハモ グリパエ幼虫含んだスイカの葉を譲り受け, それらより回収した.さらに比較として用いた生物的防除資材として用いられ ている D.i s a e a( 以後導入系統とよぶ)は株式会社 トー メン TOMEN( 現,株式 会社ア リスタライフサイエンス)より提供を受けた MINEXより選別して供試 した . これら全ての寄生蜂は,同 一条件下で、 マメハモグリバエ幼虫を寄主にし て累代飼育し,各実験に供試した .. 実験装置. 全ての実験は,上面にガーゼを張った直径 10cm,高さ 20cmの透明. F i g .5 ) . 初生葉 2枚 のついたインゲンマ なプラスチック円筒を用いて行なった ( 5 0 r nl ) に差し,これを綿で固定して円筒内に静 メ株を ,水 を入れた広口ピン (. 置し,その中へ寄主母虫を放飼し,インゲンマメ株へ 24時間産卵させ,その後 幼虫が 2齢後期から 3齢中期に成長したものを全ての実験に用いた.. F i g .5 . T r a n s p a r e n tp l a s t i cc yl i n de ru s edi nt h i ss t u d y( 2 0cmh i g h,1 0cmw i d e ). 1 5.
(21) 温度と発育の関係. 2 00Cにおいて実験装置内に最低 6 0匹のマメハモグリバエ. Nformosaは 5匹,その他の寄生蜂に 幼虫に加害されたインゲンマメ株を設置し, おいては 5対を放し, 2 4時間寄生させた.その後,この株を 1 5, 2 0, 2 5, 3 0, 3 30C, に設定した恒温室へ移し,寄生蜂成虫が羽化するまでの発育所要日数を調べた.. 30C で高温障害による発育遅延が認められなかった種については, さらに, 3 3 5o cと 37o cについても実験を行った.その際寄生させた寄生蜂幼虫がし、るイ ンゲンマメ株は,蜂の羽化が完了するまで各温度を維持した.. 雌成虫の寿命,寄生及び寄主体液摂取に及ぼす温度の影響. 4時間以 羽化後 2. 内の Nformosaは雌成虫 l匹,その他の寄生蜂種は雌成虫 1匹と雄成虫 3匹を. 5,2 0,2 5,3 00C の恒温室に設置した実験装置内に放飼し,死 ランダムに選び, 1 0匹以上の 2齢後期から 3齢前期のハモグリパエ幼虫に加 亡するまで毎日最低 6 害された株を与えて飼育した.その際. 寄主幼虫のいる株は毎日新しい株と交. 0Cの恒温室に移し,寄生蜂成虫が羽化するまで保管し, 換し,寄生させた株は 2 0. 雌雄を確認した.回収した株にいる死亡した寄主幼虫については,双眼実体顕 微鏡を用いて寄生または寄主体液摂取のいずれによる死亡かを毎日確認した. 仁. p e n t h e u sと Nformos αを除く外部寄生蜂については,死亡したハエ幼虫の体. 表やその付近に寄生蜂の卵が確認された場合に寄生による死亡と認定し,内部 寄生蜂については,卵の確認が困難だ、ったため,寄生 3日後から検鏡して,解 化した寄生蜂幼虫が確認された場合に寄生とみなした.寄主体液摂取による寄 主幼虫殺害数は,殺害総寄主幼虫数から寄生による殺害寄主幼虫数を引くこと によって求めた.. 第 3節 結 果. 温度と発育の関係 a b l e2に示した. 1 50Cでは D .i s a e aの両 各種土着寄生蜂の平均発育日数を T. 1 6.
(22) T a b l e2 .E釘e c to f t e m p e r a t u r e supond e v e l o p m e n t a lp e r i o d( d )合omo v i p o s i t i o nt oa d u l te c l o s i o no fs e v e ne u l o p h i dp a r a s i t o i d so nム立並立t ( m e a n : ! :SD) 1 50C. 2 50C. 0 20 C. 3 00C. 3 30C. n. n. n. n. n. 47 3 2 . 7士 2 . 3c d. 42 1 6 . 0 士1.9 c e. 5 2 11 .4 土1.3 b. 4 8 9 . 6: ! :1 .0 b. 1 1. 8 . 5士0 . 8 c d. . 9c 1 9 1 7 . 6 士1.2 d . 8士0 1 3 9 9. 9 9. 7.3士1.2 d. 3 70C. 3 50C n. n. 49 7.5士 0 . 6a. 4 3 7 . 6 : ! :0 . 9a. F e m a l e P .k a t o n i s H .v a r i c o r n i s. C .p e n t h 四. S. 1 1 64 6 . 7士 8 . 3b. 1 2 01 6 . 1 士1.7 d e. 6 93 5 . 2士 2 . 3c. 6 91 9 . 9 士1.8 b. F 正form 目 。. 3 25 2 . 5: !3 . 9a. D .m i n o e u s. 2 52 8 . 5士4 . 9e f. D .p u s z t e n s i s. 2 63 2 . 2士 0 . 9c d e. D .i s a e a( n a t i v e ) D .i s a e a( i n 仕o d u c e d ). 1 1 72 8 . 8: ! :3 . 2f 3 8 31 .4 士 3 . 7d e f. 6 91 4 . 1: !1 . 7a. 7 91 2 . 0: ! :1 . 1 a. 1 3 71 4 . 0 士1.6 a. 9 9 11 .9 : ! :1 . 3 a. 1 81 7. 1: ! :2 . 9c d. .6 士 0 . 9b 2 8 11. A l ld i e db. 1 5 1 7 . 5: ! :2 . 8c. 2 0 11.2士1.7 b. 1 3 7 21 .4 士1.4 a. 1 7 81 5 . 5: ! :1 .6 e 5 21 5 . 9 士1.7 d e. . 4士 0 . 6b c 1 7 9. . 1 c . 8: ! :1 1 7 01 0 . 1: !1 . 2c 1 3 6 8. 3 6 11 .0 士1.1 b. . 7b . 6士 0 4 3 8. 2 1 4 . 5士0 . 5 a b 7 1 9.3土 0 . 7 7 5 11.3土1.6 a. 1 0 5 9 . 1: ! :0 . 7. A l ld i e db 2 7 11 .7 士1.6 a b 34 1 0 . 6士 2 . 6 b 3 5. 9 . 1士0 . 5 c d. . . t ・ ・. 1. 民1 a l e. P .k a t o n i s. 5 93 0 . 4: !3 . 2d e. 6 61 5 . 6: ! :0 . 9d. .6 士 3 . 6b 5 2 11. 8 2 8 . 9士0 . 9b. 1 5. 8 . 1士0 . 7 c d. 1 2 3. 6 . 9 士1.1 d. H .v a r i c o r n i s. 1 0 24 3 . 7 士 8. 3 a. 1 5 51 4 . 9 士1.4 e. . 2士0 . 9d 1 4 6 7 . 1士0 . 9d 1 5 3 9. C .p e n t h e u s. 9 83 3 . 8士 3 . 1b. 1 4 41 8 . 8 士1.4 a. 3 4 11 . 5: ! :1 . 3 a 1 3 21 3 . 7: !1 .4 a 1. D .m i n o e u s. 2 32 6 . 9 士1.2 e. 2 61 6 . 4士 2 . 1b. 3 5 11 . 3: !0 . 6b. 3 11 . 7: ! :0 . 9a. A l ld i e db. D .p u s z t e n s i s. 1 9 31 .4 : !1 . 2b cd. 20 1 6 . 4: ! :1 . 2a c d. 1 5 11 .5 士 2 . 7b. 1 4 9 . 0士 0 . 7b c. 1 8 1 0.3士 0 . 9 b. D .i s a e a( n a t i v e ) D .i s a e a( i n t r o d u c e d ). 1 4 42 8 . 3士 3 . 5c e 7 6 29. 3: ! 2. 3 c d e. 2 5 5 1 4 . 9 士1.5 b c 7 81 5 . 0 士1.1 bcd. 1 9 21 0 . 0: !1 .2 c 2 48 8. 3: ! :1 . 1 c. 8 9. 9.4土 2. 3 b. . 6c 6 1 8.3士 0. 5 3. 8 . 5: ! :0 . 8 c. 8 01 0 . 0 士1.0 c. s U ¥ ,f o l l o w e dbysameI e t t e r sw e r en o ts i g n i 白c a n t l yd i f f e r e n ta tp< 0 . 0 5( T u k e y -Kr amert e s t ) Meansi nt h esamec o l u 目. aS i g n i f i c a n t l ys h o r t e ra tpく 0 . 0 5comparedw i t hN .formosa( t t e s t ). bA l ld i e dp r i o rt oa d u l te c l o s i o n. 7 1 5 . 1 土 3. 4 a. 5 0 6 . 9士 0 . 6. 47 6 . 9士 0 . 6.
(23) 系統と D .m i n o e u sの発育所要日数が他の寄生蜂種と比べて短く,一方 H .. v a r i c o r n i sとN .f ormosaが長かった.特に N .f ormosaは,D .m i n o e u sの約 2倍長 かった ( T u k e y -Kr amerの多重比較検定, p < 0 . 0 5 ) . 200C以上では, C .p e n t h e u s とN .f ormos αが他の寄生蜂よりも発育所要日数が長かった ( P < 0 . 0 5 ).全ての種 で 250C まで、は温度の上昇に伴って,発育所要日数が短くなった.しかし, 300C において ,D .m i n o e u sの雌成虫では供試虫数が 50匹以上であったにもかかわら ず羽化個体数が 0匹であり,雄についても羽化成虫数が 3匹と少なく,それらの 発育所要日数は 250Cと比べ遅延した .P .katonis,両 系 統 の D.isaea, D.. minoeus, D.pusztensisお よ び C.pentheusは , 3 30C において発育途中 で多くの幼虫が死亡し,かつ,羽化した場合も発育所要日数が遅延した. 3 30C において,高温障害による発育遅延が見られなかった H .v a r i c o r n i sとN .f ormos α については,さらに 3 50C,370Cで実験を試みた. 3 50C以上で H .v a r i c o n i sが 雌雄ともに多少の発育遅延が見られたが ,N .f ormos αでは見られなかった.. D .m i n o e u sでは 1 5o cから 250C 他種の寄生蜂は 1 5o cから 300Cの発育遅延 の見られなかった温度範囲で得られた結果 ( T a b l e2 ) から,発育零点 ( t o )および 有効積算温度(めを有効積算温度の法則(( t-t o ) D=K. t : 飼育温度, D: 発育所. T a b l e3 ) . その結果,発育零点は雌雄ともに仁 p e n t h e u s 要日数)をもとに求めた ( Table3 .R e g r e s s i o neq 田 t i o n sofr e c i p r o c a l s( y )ofd e v e l o p m e n t a lp e r i o d( d ) . ont e m p e r a t u r e s(X,oC ) , andt h r e s h o l dt e m p e r a t u r e sf o r development(C)andt h e r m a lc o n s t a n t s( d a y d e g r e e )ofs e v e ne u l o p h i dp a r a s i t o i d sonL .t r i f o l i i T h r e s h o l dt e m p e r a t u r e s o C) f o rdevelopment(. R e g r e s s i o ne q u a t i o n. Thermalc o n s t a n t s ( d a y d e g r e e ). Female. P .k a t o n i s Hv a r i c o r n i s 仁p e n t h e u s N .f ormosa D .m i n o e u s D .p u s z t e n s i s D .i s a e a( n a t i v e ) D .i s a e a( i n t r o d u c e d ). Y 0 . 0 0 4 9X -0.0395( R2 =0.986, F =71 .6, P=0 . 0 7 5 ) Y=0.0074X -0.0872( R2 =0 . 9 9 2, F =1 2 0 . 9, P 0 . 0 5 8 ) Yニ 0 . 0 0 3 7X -0 . 0 2 5 1( R2 =0 . 9 8 2, F =5 , 4 4 .P=0.086) Y 0 . 0 0 4 4X -0.0439( R2 =0 . 9 7 7, F =8 4 . 4, P=0 . 0 1 2 ) Y=0 . 0 0 5 1X -0 . 0 4 2 6( R2 =0 . 9 9 8, F =42, 5 4 .P 0.031) Y=0.0052X -0.0460( R2=0.987, F =15 , 6 5 .P=0.006) Y=0.0054X-0.0444( R2 =0.977, F =84, 3 . P=0 . 0 1 2 ) Y=0.0057X -0.0518( R2 0 . 9 9 8, F =1 1 4 9 . 9, P=0 . 0 0 1 ) ニ. ニ. ニ. ニ. 二. Male. P .k a t o n i s H .v a r i c o r n i s C .p e n t h e u s D .m i n o e u s D .p u s z t e n s i s n a t i v e ) D .i s a e a( D .i s a e a( i n t r o d u c e d ). , 4 . , 4 .. . 0 0 5 2X -0.0437( R2 =0.996 日F =450 P=0 . 0 0 2 ) Y 0 Y=0 . 0 0 7 9X -0 . 0 9 2 6(R2 =0 . 9 9 5 F =397 Pニ 0 . 0 0 3 ) 二. ,. Y=0 . 0 0 3 9X -0 . 0 2 6 1(R2 =0 . 9 8 8, F =1 6 6 . 6, P 0 . 0 0 6 ) Y=0 . 0 0 5 1X -0 . 0 4 0 4( R2 0 . 9 9 8,F =581 . 1 , P=0 . 0 2 6 ) Yニ 0 . 0 0 5 3X -0.0458( R2 =0.998, Fニ 1 07 2 .6, P=0 . 0 0 1 ) ニ. ニ. , , 5 . 0 . 9 9 1, F =23, 2 4 .P=0.004). Y=0 . 0 0 5 8X -0 . 0 4 9 0(R2 =0 . 9 8 5 F =1 9 3 P=0 . 0 0 5 ). . 0 5 2 1( R2 Y=0 . 0 0 5 9X -0. ニ. 1 8. 8 . 1 .8 11. 2 0 4 . 1 1 3 5 . 1. 6 . 8 9 . 7. 2 7 0 . 3 3 227. 1 9 6 . 1. 8 5 .. 8 . 8. 1 9 2 . 3 1 8 5 . 2. 9 . 1. 1 7 5. 8 4 .. 8 4 . 1 1 . 7 6 . 7 7 . 9 8 . 6. 8 4 . 8 . 8. 4 .. 1 9 2 . 3 1 2 6 . 6 256 1 9 6 . 1. 4 .. 1 8 8 . 7 7 2 .4 1 1 6 9 . 5.
(24) が最も低く,逆に H .v a r i c o r n i sが最も高かった.有効積算温度についても,仁 .v a r i c o r n i sが最も小さかった. p e n t h e u s最も大きく ,H. 雌成虫の寿命,寄生及び寄主体液接種に及ぼす温度の影響 雌成虫の寿命は, 7種類とも高温になるにつれて短くなった ( T a b l e4 ) . H. v a r i c o r n i s, N formosa ,お よ びD .i s a e aの両系統は,全ての温度で,寿命は長く, 逆に D .m i n o e u s, D .p u s z t e n s i s, および仁 p e n t h e u sは,全ての温度区で寿命は短か った • P .k a t o n i sは ,. 1 50C で、は寿命が長かったものの, 300Cでは非常に短くな. った. 全ての温度区で寄生による殺害寄主幼虫数( T a b l e4 )が多かったのは ,D.isaea (土着系統)で,特に 1 50C と 200Cでの寄生数は標準偏差が大きいものの他種 と比べて極めて多かった.しかし, 200C をピークにそれ以上の温度では寄生数 が減少した .D .i s a e a( 導入系統)は D .i s a e a( 土着系統)と同様の傾向を示し たが,その殺害数は土着系統と比べて明らかに少なかった.仁 p e n t h e u sは , 1 50C, 200Cで,寄生による死亡寄主幼虫数が比較的多かったが, 250C以上では低下. し , 30 0 C ではほとんど寄生が見られなかった • P .k a t o n i sも , 1 50Cから 250C で は比較的多かったが,30 o C ではほんど寄生が見られなかった • D .pusztensis. と D.minoeusは 全 て の 温 度 区 で 他 の 寄 生 蜂 と 比 べ て 寄 生 に よ る 寄 主幼虫殺害数は非常に少なかった.特に D .m i n o e u sは全ての温度区でほ とんど寄生が見られなかった • H .v a r i c o r n i sとNformosaは 1 50C では寄生数が. .i s a e a( 土 少なかったものの,温度の上昇とともに寄生数は増加し, 30oC では D 着系統)と同程度の寄生数となった.この 2種は高温に適応していることが分 .v a r i c o r n i sとD .i s a e a,N かった.寄主体液摂取数による寄主幼虫殺害数は ,H. formos αが実験した全ての温度区で他の寄生蜂種よりも多く,特に D .i s a e a( 導 入系統)は 250C においてもっとも多くの寄主を寄主体液摂取によって殺害した. ( T a b l e4 ) .一方,これら 3種を除く士着寄生蜂種では全ての温度区で寄主体液 摂取数は少なかった.. 1 9.
(25) Table4 .E f f e c t so f t e m p e r a t u r e suponf e m a 1 e1 0 n g e v i t y( d ), numberso f h o s t sk i l l e dbyp a r a s i t i z a t i o nandbyh o s t f e e d i n gp e rf e m a 1 et h r o u g h o u tl i f e t i m e(mean土 SD) 1 50C n. 。. 2 1 nツ ti OAutiku,、“. 。 。 。 。. ,・ 0c d a d a d cGa b 36848454. 土土士士士土士士. 7 9 . 2土 1 0 6 . 0cd 1 3 5 . 5土 8 0 . 5bc .7d 3 5 . 8 土 31 2 0 1 . 2士 5 8 . 7ab 0 . 1土 0 . 3d 3d 3 2 . 9 士 33. 4 . 9a 2 5 3 . 0土 7 1 61 .8 土 8 3 . 3bc. 1 0 1 0 1 0 1 0 1 0 1 0 1 0 1 0. 1 .0 土1.8b 1 51 . 3 土 9 8 . 7a 3 . 2土 1 0 . 1b 1 7 2 . 3土 7 0 . 0a . 8b 0 . 5土 0 4 . 8士 8 . 1b 4土 8 5 . 0a 1 7 4. .1土 7 6 . 1a 1 01. 3 4 . 1土 1 51 .6 土 7 . 3土 l : 2 1 8 . 1: 5 . 9土 7 . 6土 1 21 .5土 2 3 4 . 0土. 1 0 1 0 1 0 1 0 1 0 1 0 1 0 1 0. 3 . 5土 1 5 7 . 8土 2 . 1土 1 7 7 .0 土 4 . 8土 6 . 1土 1 3 8 . 6士 1 3 8 . 8士. Meansi nt h esameco1umn, f ol 1owedbysame1 e t t e r sweren o ts i g n i f i c a n t 1yd i f f e r e n ta tpく 0 . 0 5( T u k e y -Kr amert e s t ) .. 4 4 . 9 cd 4 8 . 7a b 5. 4 d 1 1 4 . 8a 7 . 7 d 6 . 2 d 9 8 . 7 bc 37. 4a. 5 . 3b 9 0 . 7a 4 . 2b 9 6 . 8a 5. 4b 5 . 6b 7 8 . 2a 9 9 . 0a. CN. り. A U n y A U n U A ζJnu lA Ul i----. nvnunynuoyAUF、MAU l -----. 4 5 . 2土 3 1 . 2c . 3bc 1 1 7 . 0 土 51 2 5 . 9土 1 6 . 9c 5 . 9bc 1 4 0 . 3土 7 1 5 . 6土 1 9 . 7c 2 . 2c 2 8 . 7土 2 2 3 8 . 9土 2 0 5 . 1b 1 3 . 0a 4 6 5 . 6土 2. 1 0 1 0 9. 1 4 . 0土 1 3. 4ad 2 0 . 7土 7 . 0bc 4 . 3abd 1 0 . 2士 23. 4 : l : : 9 . 2a b c 2 . 3 士1.5d 7 . 3土 5 . 0ad 5 . 6c 1 9 . 5土 1 6 . 0土 5. 4c. 1211. 5 8 . 1土 9 0 . 0b 8 0 . 1土 4 6 . 0b 4土 129. 4b 1 2 8. 1 41 .9 土 9 3 . 2b 5. 4土 7 . 1b 3 9 . 5 士 38. 4b 2 9 . 5a 3 3 4 . 5土 2 1 5 7 . 1土 1 0 4. 4b. nununUF、MAU l ----. AYAUζJnu lAU--. 9. 1 7 . 5cd 7. 4a b 1 0 . 0bcd 1 0 . 0a b c 3 . 6d 9 . 1cd 1 0. 4a 1 0. 1a. tJ4Uウ&戸コ勺jLO 五u n v 今& 4 U ウ ム n u t - 3 今. ' E且. 6 . 2b 3 2 . 8土 3 7 6 . 8土 5 3 . 7a b 1 5 . 1土 7 . 6b 3 2 . 3士 2 9 . 0b 2 6 . 5士 2 2 . 7b 6 . 0b 6 . 1士 8 2 . 3: l : 8 8 . 7a 1 0 9 . 7土 1 2 0 . 3a. --且. No.o f h o s t sk i l l e dbyh o s t f e e d i n g P .k a t o n i s 10 H .v a r i c o r n i s 1 0 C .p e n t h e u s 1 0 N .f ormosa 10 D .m i n o e u s 1 0 D .p u s z t e n s i s 1 0 D .i s a e a( n a t i v e ) 20 D.i s a e a( i n 仕o d u c e d ) 10. nり AU. No.o f h o s t sk i l l e dbyp a r a s i t i z a t i o n P .k a t o n i s 10 7 9 . 6 土 81 .8b H .v a r i c o r n i s 10 3 7 . 1土 3 0 . 7b C .p e n t h e u s 10 6 5 . 2士 6 6 . 0b N .f ormosa 10 4士 2 7 . 8b 1 7. D .m i n o e u s 10 1 3 . 0士 1 4 . 6b D .p u s z t e n s i s 10 2 . 1b 1 4 . 8土 2 D .i s a e a( n a t i v e ) 20 1 8 0 . 2土 1 3 8 . 2a s a e a( i n 位o d u c e d ) 10 D.i 6 6 . 9士 3 9 . 5a b. 1 7 . 2土 2 6 . 8土 1 7 . 2士 2 3 . 3: l : 5 . 3士 11 .5土 3 3 . 9土 31 .9 土. n u n u A U A U A U Aり AUAU. 2 5 . 7a b c 1 4 . 7a b c 11 . 7b c 1 6 . 6a b c 6 . 6c 7 . 2c 1 6 . 2a 1 8. 4ab. n. 11111111. 3 0 . 2土 .6 土 31 2 2 . 0士 2 9 . 0士 l : 1 2 . 6: 1 3. 4士 4 4 . 1土 3 9 . 5土. ounuQJAuounu,5nu l --1111. 10 1 0 10 10 10 1 0 20 1 0. 300C. n. n のunフハunヲハリ戸、νAV Iのりl ----. L o n g e v i t y( d ) P .k a t o n i s H .v a r i c o r n i s C .p e n t h e u s N .f ormosa D .m i n o e u s D .p u s z t e n s i s n a t i v e ) D .i s a e a( D .i s a e a( i n 仕o d u c e d ). 250C. 200C.
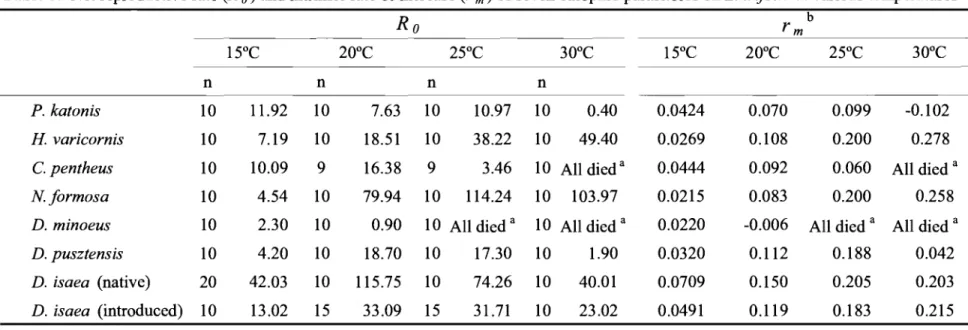
(26) Table5 .Netr e p r o d u c t i v er a t e( R0 )andi n t r i n s i cr a t eo fi n c r e a s e( rm)o fs e v e ne u l o p h i dp a r a s i t o i d sonL .t r i f o l i ia tv a r i o u st e m p e r a 加r e s. 。. b. R 1 50C n. 200C. rm 2 50C. 300C. n. n. n. 1 50C. 200C. 250C. 300C 0. 102. P .k a t o n i s. 1 0. 11 .9 2. 1 0. 7 . 6 3 1 0. 1 0 . 9 7 1 0. 0. 4 0. 0 . 0 4 2 4. 0 . 0 7 0. 0 . 0 9 9. H.v a r i c o r n i s. 1 0. 7 . 1 9. 1 0. 1 8 . 5 1 1 0. 3 8 . 2 2 1 0. 4 0 49.. 0 . 0 2 6 9. 0 . 1 0 8. 0 . 2 0 0. C .p e n t h e u s. 1 0. 1 0 . 0 9. 9. 1 6 . 3 8. 3. 46 1 0A l ld i e da. 0 . 0 4 4 4. 0 . 0 9 2. 0 . 0 6 0 A l ld i e da. Nformosa. 1 0. 4 . 5 4. 1 0. 0 7 9 . 9 4 1. 1 1 4 . 2 4 1 0 1 0 3 . 9 7. 0 . 0 2 1 5. 0 . 0 8 3. 0 . 2 0 0. D .m i n o e u s. 1 0. 30 2.. 1 0. D .p u s z t e n ぽi s. 1 0. 20 4.. 1 0. 0 1 8 . 7 0 1. 1 7 . 3 0 1 0. 1 .9 0. 0 . 0 3 2 0. 0 . 1 1 2. 0 . 1 8 8. 0 . 0 4 2. D .i s a e a( n a t i v e ). 20. 4 2 . 0 3. 1 0. 1 1 5 . 7 5 1 0. 7 4 . 2 6 1 0. 4 0 . 0 1. 0 . 0 7 0 9. 0 . 1 5 0. 20 5 0.. 0 . 2 0 3. D .i s a e a( i n t r o d u c e d ) 1 0. 1 3 . 0 2. 1 5. 5 3 3 . 0 9 1. 31 .7 1 1 0. 2 3 . 0 2. 0 . 0 4 9 1. 0. 11 9. 0. 18 3. 0 . 2 1 5. 9. 0 . 9 0 1 0A l ld i e da 1 0A l ld i e da. aA l lp r o g e n yhadd i e dp r i o rt oa d u l te c l o s i o nd u et oh i g ht e m p e r a t u r e s . bN o s .o f r e p l i c a t i o n si nrm a r et h esamea si nR玖. r e s p e c t i v e l y .. 0 . 0 2 2 0. 司. 0 . 2 7 8 0 . 2 5 8. 0 . 0 0 6 A l ld i e da A l ld i e da. ・ ,4 N.
(27) 本実験結果に基づく ( T a b l e4 ) 7種の寄生蜂の雌成虫の日齢別生存率( ι ),日齢 別産卵数(m から求めた純増加率 ( R o )と内的自然増加率は ( r m )( B i r c h,1 9 4 8 ) ) x. (T=. I :xl爪/玄 lxmx'. R o. 玄lxmx'. I :e-rmlxmx=1)(Table5),D.isaea(土. 着系統)が 1 50C,200C で純増加率が最も高く,特に 1 50C では,他の寄生蜂と比 べ 3倍以上となったが, 250C, 30 0 C で、は平均的だ、った • D .i s a e a( 導入系統) も土着系統とほぼ同様の温度傾向となった.内的自然増加率も D .i s a e a( 土着・ 導入系統)が 1 50C,20 0C で他種と比べて非常に高かった • H .l仰 ・i c o r n i sと N. 50C では低いが,温度の上昇に伴 formosaは純増加率,内的自然増加率ともに 1 って高くなった.特に Nformosaの純増加率は 300C で他の寄生蜂の 2倍以上の 値となった.一方,P k a t o n i s, C .p e n t h e u s,D .m i n o e u s,D .p u s z t e n s i sは,純増 0 加率,内的自然増加率ともに 20 C以上の温度では,上記 4種と比べて極めて低. い値で推移した.特に D .minoeus は 200Cで内的自然増加率がマイナスとな り , 2 50C以上の温度では雌成虫が羽化しなかった.. 第 4節 考 察. 土着寄生蜂の温度耐性. vanL e n t e r e n( 19 8 6 )は,外来の天敵から生物的防除資材選抜するにあたり, 8 つの事前評価基準を示した.それらの基準のうち 6つの基準は,温室における 接種的放飼法による生物的防除と密接に関連している.そのうちの 3つの基準 だけが土着天敵の有用性に関連する.すなわち適した飼育方法と同様に 際 立 っ た 繁 殖 能 力 と 密 度 反 応 性 で あ る (WaldeandMurdoch,1988). これらは特に温度と密接に関連する.西日本の温室は外気温と異なり 1年を 通じて激しく変動する;例えば,静岡農事試験場における 1 9 9 5年から 1 9 9 7 年までの記録によると,平均気温は冬季で 1 5 . 6o C( 6 . 0' " ' ' 31 .6 O C ),夏季は平 均2 8o C( 19 . 0' " ' '4 4 . 9O C ) であった(小津博士,私信).したがって,そのよ うな温度環境で用いられる生物的防除資材は,極端な温度変化に対する高い適. 22.
(28) 応性が求められ,それは v a nL e n t e r e nの導入天敵の事前評価における 3番目の 基準である「気候への適応 J に相当する.ゆえに,施設内で接種的放飼するた めの生物的防除資材を土着寄生蜂類から選抜するにあたり,温度耐性,特にそ れらの発育と寿命および繁殖能力に関する評価は必須である. 本研究では,西日本における有力なハモグリバエ類寄生蜂類で、あり ( Takada. andKamij0,1 9 7 9 ;Sugimotoe ta , . 1 1 983a ;Kato,1994,1996a,b ), かつ わが国の施設聞場においてマメハモグリバエに寄生が確認された(小西,. 1998)7種のヒメコバチ科寄生蜂に注目し,マメハモグリパエの生物的防除資 材として最適な寄生蜂種を選抜するために,それらの発育および繁殖能力にお ける温度耐性を実験的に比較,検討した.その結果,それらは高温適応的な 2 種,H v a r i c o r n i s,N formosaおよび低温適応的な他の種に大別することができ た• D .i s a e a( 土着系統)の寄生による寄主の死亡率が. 2 5C以上で大きく減少 0. したものの,全ての温度区において 7種の中で最も高かったことは注目される. ( T a b l e4 ) . 逆に ,D.minoeusのそれは概して非常に低いものの,試験されたすべ 5 oCで最も高かった.わが国では ,D.m i n o e u sは高地におけ ての温度区では 1. K a t o, る捕食寄生蜂群で優勢種であるものの,低地では劣勢であることから ( 1 9 9 6 a, b ),D.minoeusは,より低温に適応的な種であるようである. 杉本(19 9 8 ) は,わが国に生息するハモグリバエ類と,それらの土着寄生蜂 類に関して各々の世界的な分布域の聞には明白な対応関係、があり,それぞれ南 方型と北方型に大別できる事,そして,その類型化は個々の種の温度特性と密 接な関係があることを明らかにした.本研究において解明された温度特性に基 づく 2つのカテゴリーは,杉本の説と良く一致した.. 放飼と大量生産の各段階における捕食寄生蝉の有効性の総合的な 評価 生物的防除資材として有効な捕食寄生蜂は,その寄主と同等ま rm)を 持 つ べ き で あ る こ と は し ば し ば 述 たはより大きい繁殖能力 (. 23.
(29) べられる. 本研究に供試した全ての寄生蜂は,寄生だけでなく寄主体液摂 取によっても寄主を殺害した.寄主体液摂取による寄主殺害は, 害 虫 の 防 除 に お い て 有 効 で あ る (YamamuraandYano,1988;Kidd. andJ e r v i s,1989). そ れ ゆ え , 寄 生 蜂 の 効 率 的 評 価 の た め に は , 寄 生 だ け で な く 寄 主 体 液 摂 取 に も 基 づ く べ き で あ る (vanLenteren,. 1986). し か し , 寄 主 体 液 摂 取 に よ っ て 寄 主 を 殺 さ な い 寄 生 蜂 と 異 なり,寄主体液摂取によって寄主殺害を行う寄生蜂の評価は複雑 である.すなわち圃場での「寄生蜂放飼段階」においては寄主体 液 摂 取 に よ る 寄 主 殺 害 に よ っ て , 高 い 防 除 効 果 を 得 ら れ る . しか し.寄主体液摂取により殺害された寄主は,次世代虫生産に直接に は寄与しないので,寄主体液摂取による寄主殺害は「大量増殖段 階j においては寄主の無駄な利用ということになる.その結果,寄 主体液摂取による寄主殺害の生物的防除全体に及ぼす効果は,生物的防除の「放 飼段階」と「大量増殖段階」の 2段階間で相反することになる.したがって, 生 物 的 防 除 資 材 と し て の 寄 生 蜂 の 効 率 評 価 は , こ れ ら の 2段 階 に おける効率を総合的に考慮、して行われるべきである.. Hondoe ta1 .(2006) は ,. 生物的防除資材としての有効性を増殖段階から. 放飼段階まで通して総合評価するため,生物的防除効率 B i o l o g i c a lC o n t r o l. E f f i c i e n c y (BCE) を提案した • BCEは以下の式で求められる.. BCE=(Nov+N h f)aRNov/(N o v+Nh f). この方程式は. 1匹の母蜂によって寄生のために殺害された寄主. 数 を Nov ,寄 主 体 液 摂 取 に よ っ て 殺 害 さ れ た 寄 主 数 を Nhj,同 様 に 娘 蜂 に よ っ て 寄 生 の た め に 殺 害 さ れ 寄 主 数 を Nい , 寄 主 体 液 摂 取 の た め に 殺 害 さ れ た 寄 主 数 を N'hjと す る . さ ら に 寄 主 1匹 あ た り. 24.
(30) の 平 均 産 卵 数 を α, 次 世 代 雌 成 虫 比 を R とする .α RN o vは , 母 蜂 の 純 増 加 率 (Ro). にほぼ等しい •. ( N ' o v+N ' h f ) /(N h f )は , 母 蜂 の ν +N o. 「増殖段階」における温度と娘蜂の「放飼段階Jにおける温度の 組 み 合 わ せ に よ り 変 化 す る (Table4 ) . また,生物的防除行程における 2 段階間の総寄主殺害比率を示すだけである (Table 5). し た が っ て BCE はんに依存し,特に「増殖段階」と「放飼段階」の温度が等しい とんに等しい.娘蜂の寄主体液摂取による寄主殺害の利益により母蜂の寄 主体液摂取による寄主殺害の損害が相殺されるため ,BCEは寄主体液摂取に直 接影響されない.上記の方程式において,なんらかの方法によって Nhfを 減 ら す こ と が で き れ ば ,BCEは 増 大 す る . Ueno( 1997年)は効. 率的な大量生産のために,メスに寄主の代わりに人工飼料を給餌 して寄主体液摂取の割合を減らすことを提案したが,それは人工 飼料の生産コストが寄主生産コストより安い場合のみ当てはまる. 0. Table6は , 20Cで 生 産 さ れ た 7種 の 寄 生 蜂 の 娘 蜂 が , 種 々 の 温 度 区 に 放 飼 さ れ た と 仮 定 し て 計 算 さ れ た BCE値 で あ る . N. formosα 0. (産雌単為生殖系統)の BCEは , 25C以 上 で 供 試 寄 生 蜂 の な か で 最 も 高 く , 逆 に D. isaea (土 着 系 統 ) の そ れ は ,. 0. 0. 1 5C と 20Cで 最 も 高. か っ た . ま た , こ れ ら の 2種 の BCEは 供 試 さ れ た す べ て の 種 の 間 で 一 貫 し て 高 か っ た .D. isaeaの 導 入 系 統 は , 士 着 系 統 と 比 べ て 寄 主 体 液 摂 取 に よ る 寄 主 殺 害 数 が 多 い の で , 導 入 系 統 の BCEは 土 着 系統より劣った. これらの結果より,この研究で実験に供試されたどの寄生蜂種 も,温度の変化の激しい西日本の施設閏場において年間を通した 一貫使用には適さないことが分かった • BCEの 優 れ た D.i s a eα(土. 着系統)と N.formosa(産 雌 単 為 生 殖 系 統 ) に つ い て み る と ,D. isaea ( 土 着 系 統 ) が 相 対 的 に 広 い 温 度 範 囲 で 適 用 可 能 で あ る が , BCE 0. は 30Cで 減 少 し た . 一 方 , 高 温 耐 性 の 高 N.formosaでは, 300C以上の. 2 5.
(31) 高温で高い防除効果が期待できるが,反面 200C以下では期待できない.した. がって,西日本の施設圃場では,平均気温 20から 2SCを目安として,秋の後 0. 半から春にかけた低温季には D. isaea (土着系統)を,高温季には高温耐性 産雌単為生殖系統)を使い分けるのが最適と思われた. の高い Nformosa (. 30yooζJ-. 戸、︼司. ni---. ﹁. 司 ζυ J a 1 3 3. :ht 一 30592917 一 剖ゆ一 0 9 0 8 0 2 3 2一 d142961e-u 一de-一. H 況. ロ. 戸旦一一. 3 -3-j 76886003一 a 時一叱一3 9 5フ 2 1 后ρ 一 位己 5-86480151f ' 2 2 2 1 1 17 2 - 一 一一-. 一一. 一一一. 一 m. 沼氏一一 3d. £U. ah. む. o se -一-一 一 一 同 一 59m 益畑一一 ℃ 一 rb5399フフ mm一 a n 一間一O 7 一 8 6 9 0 8 5 3一 p u - 2 2 1 1 7 1 1 3一. 江一一一. 盟T 一 一 一. 制問韮一一 g n. ・胃. Y. 托一. 問削一. m o u -. 町 一. AU一. 5 AtL12. 弘 一 ,1一&仏1oa 1ζJ 一 ・. ωωη" 見一. mH一一一 別問一℃一却刈ロ 戸し曹、--. A U一 FU 江. a. M剖一)﹄ 川 川 一 u d -E CG-c-. 堕. 刊 一 山一・ぽ. 一川・附仇仰悦政沼均一. 刊訂引. ss.t m ぽ zs.M. d 路一今川一 c h一 川 町 一 B. E C. ιm. ・削・刷一. R E 仁 爪Daaa一. JLEq7nsM 仙 LMmmaρNmm. U M一. 26.
(32) 第 4章 ハ モ グ リ ミ ド リ ヒ メ コ パ チ の 大 量 増 殖 法 の 確 立. 第 1節 は じ め に. 第 3章においてわが国のハモグリバエ類の優勢な寄生蜂群で、あるヒメコバチ 科から,温度特性と生物的防除効率 (BCE) に基づいてイサエアヒメコパチ D.. i s a e aとハモグリミドリヒメコバチ N .formosaの 2種をマメハモグリバエ(本章 において以後ハエと表記)に対する有望な生物的防除資材として選抜した. 寄生蜂を生物的防除資材として商品化するためには,効率の高いそして低コ ストな寄生蜂の大量生産工程の確立が必要である.わが国の高い人件費を考慮 すると,全作業工程の中でも最も人件費を要すると考えられる寄主および寄生 蜂の増殖工程の省力化・効率化が求められる.さらに,剤型,梱包,保蔵方法 といった品質管理についても検討が必要である.そこで,本章で、はまずノ¥エの 生物的防除資材として. ハモグリミドリヒメコバチ産雌単為生殖系統(本章に. おいて以後ノ¥チと表記)を採用することとし,その効率的な大量増殖工程を設 計した.ついで作業の省力化・効率化のための大量増殖装置の試作とその性能 評価,さらに剤型,梱包,保蔵について検討した.ところで,ハエおよびハチ ともに,寄主植物 1株あたり可能な限り多くの個体を生産することが求められ るが,ハエが寄主植物に過剰産卵すると,過密の弊害が生じてハエ幼虫の発育 不良や死亡率上昇を招き,さらには寄主植物の枯死を招く.また,ハチ成虫の 過剰放飼は寄生蜂の寄主体液摂取による寄主死亡率を高め,その結果寄生効率 の低下を招く(大野ら. 1 9 9 9 ) . そこで本研究では,寄主植物 l株あたりのハエ. とハチの最適放飼密度を実験的に検討した.. 第 2節 実 験 の 準 備. 大量生産工程設計の概要 図 6に本研究で設計したハエおよびハチの大量生産工程の概略を示した.. 2 7. , . ., . .. 、 ー 、 』 ー.
(33) に提案する大量増殖工程は,寄主植物栽培,ハエ生産,ハチ生産およびハチの 梱包・保蔵の 4工程からなる.このうち,ハチの増殖は,はじめの 3工程を経 る必要がある.生産コストを抑えつつ天敵の安定供給を行うためには,第 1に 生物的防除資材として性能が高く,かっ生産効率が高い寄生蜂種の選抜,第 2 に栽培容易な寄主植物と増殖容易な寄主種の選定 ( F i n n e yandF i s h e r ,1 9 6 4 ),第. 3に寄主および寄生蜂の生理,生態的特性を考慮した各作業工程における各種 装置の作成とその稼動時間の設定などが必要である.本研究では,候補寄生蜂 種として,温度特性を加味した基準である生物的防除効率 (BCE) によって選 抜したイサエアヒメコバチとハモグリミドリヒメコパチのうち,増殖および管. b a c h i αに感染さ 理が容易な寄生蜂ノ¥モグリミドリヒメコバチの共生微生物恥 l れた ( S t o u t h a m e randLuck,1 9 9 3 ) 産雌単為生殖系統 (Hondoe ta , . l 2006) を採 用した.本系統は,鹿児島県農業試験場の好意により入手した. 一般的に天敵生産における大幅なコスト低減と省力化のために代替寄主・人 工飼料の利用が求められるが ( S t i n n e r ,1 9 7 7 ),ハエ寄生蜂に関しては有望な人 工飼料は未開発であること,また代替寄主の使用によって寄主選好性に影響を 9 7 7 ),寄主としてハエ幼虫を 与える危険性があることから (MorisonandKing,1. 用い,寄主植物としてはインゲンマメ ( P h a s e o l u sv u l g a r i sL.)の十分に展開し た初生葉を採用した.インゲンマメは他種寄主植物と比べて寄主幼虫の発育が 9 9 9 a ) 食草として優れ 速く,繁殖能力も高いなど(西東ら, 1995a;小津ら, 1 o. 0. ており,さらに栽培が容易で, 20C " " ' ' 2 5C下の温室では初生葉が実験に必要な 0日から 2週間程度と短し、(西東, 1993;1997a; 大きさに成育するまでに播種後 1 b ).. ハチの生物的防除効率 (BCE) は 25Cで最も高く (Hondoe ta , . l 2006),ハエ 0. 0. の増殖能力も 25Cが最も高かった(小津, 1 9 9 9 ) . これらの理由から,大量増 0. 殖工程の温度を 25Cに設定した. 0. ハエ幼虫は, 25C下でインゲンマメを寄主植物として飼育すると,卵期間は 3 . 1土0 . 2日 (mean 土SD 以下同様),幼虫期間は 4 . 0 土0 . 2日で,産卵後平均 7 . 1日. 2 8.
(34) 寄主植物 生産工程. l. l. ハエ生産工程. E. 梱包・保蔵工程. ハチ生産工程. ハチ母虫回収 産卵装置. 羽化成虫 回収装置 ノ、チ育成 ( 7日間). ハエ育成 ( 5日間). ノ、チ:卵 幼虫・踊 ハエ:卵 ~2齢. ハエ:踊. ハエ・ハチ回収 ( 10日間). 、 。 N. ハチ.踊・羽化 I I ハエ:踊・羽化. II. ・・・・圃圃______.1. 1 li-. 収一-. 長一-. 一 -. エ回一ハ虫一-. 一 -. 11-. ﹂-. -. 羽化成虫回収装置. .放飼 保蔵・輸送.. 梱包 (網付紙箱). 図6 .放飼寄生蜂ハモグリミドリヒメコパチの大量増殖工程.
(35) 目に踊化のために寄主植物を脱出する(西東ら, 1 9 9 5 a ) . トマトを寄主植物と. 9 9 5 a ), トマトの場合に すると若干発育が遅れるといわれているが(西東ら, 1 は産卵後平均 5.5日でハチの寄生適齢期である 3齢期まで発育したことから. ( M i n k e n b e r g,1 9 9 0 ),これらを参考にハエ増殖工程においてハエ産卵期間を 1 日間,そしてハエ幼虫の育成期間を産卵後 5日間と設定した.一方,ハエの寿 命,生涯産卵数は,飼育環境や採集地域によって変動するが(西東ら, 1 9 9 5 a ; 小津ら, 1 9 9 9 a ;徳丸・阿部, 2 0 0 3 ),本研究で用いた系統の 2 5Cにおける平均 0. . 9 土2 . 0日であり,生残寄主成虫の日当たり産卵数は羽化後 8日まで比 寿命は 7 較的多かった(図 7 ) . そこで,寄主生産工程においては,活発な産卵を持続す るために羽化直後のハエ成虫を母虫として 7 日ごとに補充することとした.. 40. 1 6 0. 35. 1 4 0. 30. 1 2 0. ~ 25. 1 0 0. 警 20. l 四 童 幽 F む 80 理. z 恒不. 明 到. 5 1 5. B 長 60 40. 1 0. 20. 。. 。 2. 4. 6. 9. 1 0. 1 1. 生残成虫の羽化後日齢(日). ーヨ五日志向 +累積由主7 図7 . マメハモグリパエ生残成虫 l匹による日齢ごとの平均産卵数および累 積産卵数の推移(本藤. 1 9 9 9 ). . 9 士0 . 7日 ( m e a n : l :S D ),踊 2 5Cにおけるハチの産卵後踊化までの発育期間は 5 0. 期間は 8 . 1土 1 .7日,雌成虫寿命は 2 3. 4 士9 .2日であった ( H o n d oe ta , . l2 0 0 6 ).こ れらの結果をもとに,寄生蜂生産工程においてハチの寄生のための産卵期間を. 1日間,そして育成期間を寄生後 7日間に設定した.そして,成虫剤型の場合. 3 0.
(36) ( 図6 )の羽化成虫回収期間を発育期間の個体差を考慮し 1 0日間と幅を持たせ た.生残ハチ成虫の日当たりの寄生数は,羽化後 2週間までは比較的多かった ので(図 8 ),活発な寄生を持続するために羽化直後のハチ成虫を母虫として 2 週間ごとに補充することとした.ハチによる寄生と寄主体液摂取による殺害を 免れたハエ幼虫は,ハチ育成期間中にインゲンマメ葉より脱出して多くは床面 で踊化する.これらのハエ踊は回収し,ハチ母虫回収用の羽化成虫回収装置(図 6 )へ放飼することとした.つぎに,ハエの平均発育全期間は,前述した卵期間,. 幼虫期間と踊期間 9 . 7 土0 . 5日を合わせ 1 6 . 8日(西東ら, 1 9 9 5 a ) なので,ハエ生 産工程において,たとえばハエ産卵(I日間)終了間際に産卵されたハエのう ち幼虫期に寄生を免れて葉上で踊化した個体は,ハチ羽化成虫回収装置内に移 して 4日後に羽化することになる.一方,ハチについてみると,産卵後,羽化. 4 . 0 士 1 .6日 ( H o n d oe ta , . 12 0 0 6 ) であった.したがって上記 までの発育全期間は 1 において寄生されたハエ幼虫についてみると,ハチ寄生(I日)開始直後に寄 生したハチは,羽化成虫回収装置に移して 6日後に羽化することになる. した がってもっとも遅れて羽化するハエと,もっとも早く羽化するハチの羽化予定. 200 2 5 . 0". 1 5 0. 2 0 . 0 凶 巳. 語 ま E. ". 制 1 0 0理 事 B 長. 書 非“ 霊 住 性. l l i H I : : ! r n U I I U I I. 1 0 . 0. l. 5 0. 島周囲. 。. 12 3 4 5 6 7 8 91 01 11 21 31 41 51 61 71 81 9202 122232 42 5262 72 82 93 03 13 23 3343536373 8 生残成虫の羽化後日齢(日). │圏平均産卵数二千票積産卵数. l. 図 8 . ハモグリミドリヒメコパチ生残成虫 1匹による日齢ごとの平均寄生. Hondoe ta , . l2 0 0 6より) 数および累積産卵数の推移( 3 1.
(37) 日には 2 日程度のずれが生じることとなり,羽化したハチ成虫にハエ成虫が混 在して回収される可能性はほとんど無いと考えられる.しかし,成虫剤型の場 合には,ハエ成虫の混入を 1匹たりとも防がなければならないので,羽化成虫 回収装置の回収部底面に lmm目の網を貼り,ハチ成虫だけが移動可能とした. 現在,導入寄生蜂ハモグリコマユバチとイサエアヒメコパチは,植物検疫上 の理由から剤型が成虫態に限られているが(農薬登録情報,. h t t p : / / w w w . a c i s . g o . j p / ),成虫剤型ではその保蔵および輸送過程でハチ成虫の寿命 の損耗,さらに温度環境の変化や栄養不足による品質低下を引き起こす.特に,. s y n o v i g e n i c ) であるので,成虫の ヒメコバチ類のほとんどの種は逐次成熟性 ( 生存や卵形成に必要なたんぱく質を得るために寄主体液摂取をするため. ( F l a n d e r s,1935;J e r v i sa n dKidd,1 9 8 6 ),成虫保蔵は寿命や繁殖能力に大きく S u g i m o t oe ta , . l1 9 8 3 b ) . 一方,踊剤型は設置したハ 影響を与える恐れがある ( ウスにおいて寄生蜂は羽化後直ちに寄主攻撃が可能であるので,保蔵・輸送過 程における成虫寿命の損耗,栄養不足などによる品質低下の恐れが少ない.本 研究では成虫剤型と踊剤型について品質を検討した.. 産卵装置の設計 従来の実験において一般的に行なわれてきた吸虫管による成虫の回収作業は,. P a r r e l l ae ta , . l1 9 8 9 ),より省力的な回収装 全生産工程の中でも特に労力を要し ( 置の作成が求められる.巌崎ら(19 9 9 ) は連結した 2つの飼育箱を用い,正走 光性を利用して産卵を終えたハエ成虫および寄生蜂成虫を効率よく回収する産 卵装置を考案した.他方,正走光性に加え,負走地性も利用した回収装置も考 案された(小津ら, 1998;大野, 1998;大野ら, 1 9 9 9 ) . 本研究では,これらを 参考に,以下の 2通りの産卵装置を試作した. まず,正走光性を利用して成虫を隣りの産卵区画へ移動させる横移動式産卵 装置を試作した.この装置は木製フレーム(横 70cm,高さ 25cm,奥行き 46cm) の内側側面にアクリル板(厚さ O. 5mm),底面はベニヤ板,天井面は通気性を. 3 2.
(38) 考慮、してさらし布を張り,上 ・下端に移動用開口部(縦 3cm,横 30cm) を開け た仕切り板を装置中央部に設置して装置を 2区画に分割した(図的 .増殖は 2 区画で交互に行うこととし,寄主生産工程ではまず,一方の区画にインゲンマメ 株を設置した後,ハエ成虫を放飼して産卵させた .産卵終了後,その区画を暗 幕で遮光して,正走光性により成虫が明るい他方の区画に移動するように した. 寄生蜂生産工程では,ハエ 3齢幼虫がいるインゲ、ンマメ株を 一方の区画に設置 した後,ハチを放飼して寄生させた.寄生終了後,ハエと同様にして他方の区 画に移動させた .. 図9 . 横移動式産卵装置(詳細は本文参照). つぎに,寄生終了後のハチ成虫の回収率の上昇を目指して,正走光性と負重 力走性を利用した縦移動式産卵装置を試作した . この装置は木製フレーム(幅. 3 5c m,高さ 3 0c m,奥行き 3 5c m ) の内側側面にアクリノレ板 ( 0.5mm),天井面 は通気性の確保とハチ脱出を防止するために O.lmm目の網を張り,天井面の中 央部に穴(縦 10cm,横 8 .5cm) を開け,上部を切断したペッ トボトルの開口部. 3 3.
(39) をこの穴に取り付けた(図 7).ハチの寄生については,成虫回収口にあたるベ ¥エ幼虫のいる ッ トボトルの注ぎ、 口をキャ ップで 閉めた後,この装置内に 3齢ノ イン ゲンマメ株を設置 し,ノ¥チを放飼して寄生させた .寄生終了後,キャ ップ を外し,あらかじめ用意したベットボ トルのキャ ップ 2つを両面粘着テ ープで 張り合わせて,そこ穴(直径 20mm) を開けたものをベッ トボトルに取り付け た.次いで,ペッ トボトノレ部を除く産卵装置全体を暗幕で遮光して,正走光性 を利用してハデを明るいペットボトル部に移動させた.この際,回収部を産卵 装置の天井面に設置したことにより負走地性の相乗効果による回収率の向上が 期待できる.. 図1 0 . 縦移動式産卵装置(詳細は本文参照). 34.
図



+2

Outline
関連したドキュメント
は,医師による生命に対する犯罪が問題である。医師の職責から派生する このような関係は,それ自体としては
告—欧米豪の法制度と対比においてー』 , 知的財産の適切な保護に関する調査研究 ,2008,II-1 頁による。.. え ,
● 生徒のキリスト教に関する理解の向上を目的とした活動を今年度も引き続き
具体的な取組の 状況とその効果 に対する評価.
造船に使用する原材料、半製品で、国内で生産されていないものについては輸入税を免除す
具体的な取組の 状況とその効果 に対する評価.
● 生徒のキリスト教に関する理解の向上を目的とした活動を今年度も引き続き
※1 廃棄物等の発生抑制(リデュース:Reduce,原材料を効率的に製品を長期間使用する等により廃棄物になる
