③カレニア等有害鞭毛藻の生理・生態特性の把握による発生機構の解明
イ
. 現場ミクロコズムを用いたカレニア等有害赤潮鞭毛藻の動態と環境条件との関
係の解明
水産研究・教育機構 瀬戸内海区水産研究所 紫加田知幸,鬼塚剛,北辻さほ,中山奈津子,坂本節子,阿部和雄,松原賢 大分県農林水産研究指導センター 水産研究部 大竹周作,井口大輝 埼玉大学大学院 理工学研究科 西山佳孝 1 全体計画 (1)目的近年,西日本の沿岸域において,Karenia mikimotoi や Chattonella 等鞭毛藻による有害赤潮
が頻発しており,甚大な被害が報告されている。これまで直接赤潮を駆除する技術が実用化 された例はほとんどなく,現在のところ,発生状況を見ながら,筏の移動,餌止め,出荷等 を計画的に実施する「事前対策」が最も有効な被害軽減策である。しかしながら,効率よく 事前対策を実施するためには,各海域において赤潮の発達あるいは衰退が起こる際にキーと なる環境条件を事前に特定しておく必要がある。そこで本課題では,現場垂下式のミクロコ ズム実験法や生理活性の定量法(光合成活性および細胞周期に関与する mRNA やタンパク 質の発現量など)を改変または開発し,それらを用いて,近年甚大な被害が発生している豊 後水道および八代海において,Karenia 等有害赤潮鞭毛藻の現場個体群の増殖,他生物群と の共存・競合,捕食-被捕食関係とそれらに及ぼす諸環境条件との関係を定量的に明らかに する。これにより,当該海域における赤潮発生機構がより明瞭になることはもちろん,複数 の環境条件の相乗効果や捕食圧など室内実験では取得困難なパラメータを取得できるので, 現存の赤潮シミュレーションモデルを大幅に補強することにもつながる。なお,本課題では 有害赤潮鞭毛藻およびそれらの競合種(優占珪藻種)を研究対象とする。 (2)試験等の方法 平成29 年度は,平成 28 年度までに得られた情報から赤潮の衰退過程に重要と考えられた K. mikimotoi 等赤潮藻類の生残,鉛直分布および光合成の日変化に及ぼす光や栄養塩の影響 を調査船よる昼夜連続観測および室内実験により詳細に明らかにする(担当:水産研究・教 育機構 瀬戸内海区水産研究所,埼玉大学)。また,従属栄養性プランクトンによる捕食や ウイルス感染に及ぼす光や栄養塩条件などの影響を室内実験により明らかにする(担当:水 産研究・教育機構 瀬戸内海区水産研究所)。さらに,K. mikimotoi の頻発海域である豊後水 道の本種の発生と気象等との関係を整理し,統計的手法を用いて解析する。最後に,平成 28 年度までの成果を整理し,K. mikimotoi 赤潮の発達あるいは衰退が起こる際にキーとなる 環境条件を提案する。 2 平成29 年度計画および結果
(1)目的 全体計画と同じ。 (2)試験等の方法 全体計画と同じ。平成29 年度の実施内容および方法は以下の通りである。 1) K. mikimotoi 赤潮衰退期の鉛直移動および環境条件の昼夜連続観測 平成28 年度までの現場調査により,K. mikimotoi の赤潮衰退と環境条件との関係について 情報を蓄積してきた。平成29 年度は,K. mikimotoi 赤潮衰退時における細胞の鉛直分布,光 合成活性および環境条件の日変化について特徴を見出すことを目的とし,八代海において調 査船しらふじ丸による昼夜連続観測を行った。まず,平成29 年 7 月 23 日に,K. mikimotoi の高密度海域を探索した。その結果,K. mikimotoi の細胞密度が最も高かった楠浦湾 Stn. B (水深23.1~27.6 m,32-23.2N,130-13.7E)を調査定点として選定し,同日にその周辺の湾 内2 点(Stns. A, YK-22)の細胞密度を調査して周辺の細胞分布を把握した(図1)。その 後,7 月 24 日 8:00~25 日 8:00 に昼夜連続観測を実施した。2 時間おきに採水(表層から海 底まで 2 m 深ピッチ,クロロフィル極大層)および多項目水質計(AAQ-RINKO, JFE
Advantech Co., Ltd., Hyogo, Japan)や日射計(LI190S, LI-COR, Inc., Lincoln, Nebraska, USA)
による環境計測を行った。また,表層およびK. mikimotoi 細胞極大層の海水試料については 光合成活性を測定した。採取した海水を直ちにクーラーボックスに入れて遮光し,10~20 分後にWater-PAM(Waltz 社製)を用いて,Fv/Fm(光化学系 II 最大量子収率)を計測した。 ただし,クロロフィル蛍光値が極めて低い場合は計測しなかった。 2)K. mikimotoi 以外の有害赤潮生物の生残および光合成に及ぼす貧栄養および強光の影響 平成27,28 年度の室内実験結果から,強光と貧栄養の 2 つの条件が重なると,K. mikimotoi の光合成および生残が阻害されることが明らかとなった。平成29 年度は同様の阻害現象が 他の有害赤潮生物においても起こるか否かを明らかにすることを目的として室内実験を行 った。実験には,八代海より分離したChattonella antiqua の無菌クローン株を用い,通常培 養条件(12hL:12hD,明期 6:00~18:00,100 μmol m-2 s-1,白色光)で継代培養した。培地と して,完全培地(改変SWM-3 培地)および栄養塩無添加海水(トリスを添加して pH7.8 に 調整後,ろ過滅菌した海水)を用いた。対数増殖期のC. antiqua を滅菌ろ過海水(ろ過滅菌 した海水)で洗浄後,細胞密度が104 cells mL-1となるように各培地に接種した。その後,継 代培養と同じ温度および明暗周期に設定し,通常光(100 μmol m-2 s-1)あるいは強光(10:00 ~14:00 に強光 1,000 μmol m-2 s-1を照射,それ以外の明期は100 μmol m-2 s-1)条件で培養し, 毎日,光学顕微鏡下で細胞を計数して細胞密度を追跡した。次に,完全培地で継代培養した C. antiqua をそのまま(対象区),あるいは栄養塩無添加海水に移して 0, 1, 2 日間培養後, 一部の培養液を取り出して強光(2,000 μmol m-2 s-1)を照射した。30 分おきに 120 分間,ク ロロフィル蛍光測定装置PAM-2500(Waltz 社製)を用いて光合成活性 Fv/Fm を追跡した。 3)塩分が Noctiluca scintillans の鉛直分布に及ぼす影響 既往知見(Nakamura 1994)およびこれまでの本課題における調査実験結果より,従属性 渦鞭毛藻類N. scintillans は有害渦鞭毛藻類 K. mikimotoi の捕食者としてその動態に影響を与
える可能性が示唆されている。どのような環境下で両者の遭遇率が高まるかを解明するため に,塩分がN. scintillans の鉛直分布に及ぼす影響を明らかにすることを目的とした。実験に は,広島湾で採集し,Heterocapsa sp.を餌として,改変 SWM-3 培地で 25℃,12hL:12hD(明 期6:00~18:00,100 μmol m-2 s-1)の条件下で2 週間維持した N. scintillans 細胞を用いた。ま ず,25℃の恒温室内に設置した 10 cm 間隔に採水口を有するアクリル製円柱水槽(水深 85 cm)に,低塩分(10, 20, 33)の培地を上層に, 高塩分(33)の培地を下層に等量注入した。 次に,予め24 時間絶食させた約 1,000 cells の N. scintillans が含まれる懸濁液 50 mL を注射 器で各水槽の底層から接種し,24 時間培養した。1~6 時間おきに,各採水口から注射器で 3 mL サンプリングし,実体顕微鏡下で正常な形態を有する N. scintillans 細胞(生細胞)お よび異常な形態を有する細胞(死細胞)を計数した。サンプリング後,表層および底層から 各塩分の培地を15 mL ずつ補填した。また,実験終了時には円柱水槽内の N. scintillans を目 合い 75 μm のネット上に捕集し,生細胞と死細胞を計数した。各層の塩分は実験開始時, 15 時間後,実験終了時の計 3 回測定した。 4)K. mikimotoi へのウイルス感染に及ぼす光照射の影響 K. mikimotoi は赤潮衰退前に表層に浮上することが知られている(宮村 2016)。また, K. mikimotoi に感染するウイルスが存在する可能性も指摘されている(Onjj et al. 2003)。表
層は光の波長構成が中層と異なり,紫外線も多く含まれるが,紫外線によってウイルス感染 が促進されるとの知見もある(Weinbauer 1997)。また,一般的に宿主 DNA に組み込まれ た状態で存在するファージ(細菌に感染するウイルス)を溶菌サイクルに誘導する場合にも 紫外線照射が使われることから,紫外線によるウイルス感染の誘発がK. mikimotoi 赤潮の終 息に寄与する可能性を考えた。そこで,本研究では紫外線がK. mikimotoi のウイルス感染に 及ぼす影響について検討した。まず,24 穴プレートに 800 μL の K. mikimotoi KmH6 株の培 養液を注入し,K. mikimotoi に感染するウイルス KmDNAV5 懸濁液(ウイルス接種区)ある いは改変SWM-3 培地(対照区)を 100 μL ずつ加え,約 1,000 cells mL-1に調製した。その後, 12hL:12hD(明期 6:00~18:00,50 μmol m-2 s-1,白色蛍光灯),25℃の条件で培養した。光照 射については,明期中に1時間,岡崎大型スペクトログラフを用いて,強度50 μmol m-2 s-1 の異なる波長の単色光を鉛直上方向から追加照射した。光波長は UV~可視域の 320, 360, 440, 600 nm に設定し,追加照射しなかったものを通常光とした。試験開始から 3 日後まで 毎日,K. mikimotoi の細胞密度を光学顕微鏡下で計数した。また,培養開始時および 3 日後 にMPN 法を用いてウイルスの力価(感染力)を測定した。 5)豊後水道における K. mikimotoi の発生と環境条件との関係 宇和島湾および佐伯湾における2006~2017 年 5~8 月の K. mikimotoi 細胞密度データを用 いて,本種の発生規模と環境条件との関係を解析した。ただし,2011 年の宇和島湾につい ては極端にデータ数が少なかったため,解析対象から外した。宇和島および佐伯の気象デー タ ( 降 水 量 , 日 照 時 間 ) は 気 象 庁 の ウ ェ ブ サ イ ト (http://www.data.jma.go.jp/obd/stats/etrn/index.php)より得た。統計解析は
IBM SPSS Statistics
(3) 結果および考察 1)八代海における K. mikimotoi 赤潮衰退期の細胞および環境条件の鉛直分布の日変化 楠浦湾付近では,本調査終了後すぐにK. mikimotoi 赤潮は衰退したことから(データ非表 示),調査時は赤潮衰退直前の時期にあったと推察される。調査期間中,K. mikimotoi の細 胞極大層深度は0~3 m 深で推移し,一貫して表層付近に集積していた(図 2)。過去に, 同湾のほぼ同じ定点においてK. mikimotoi が表層~底層を 20 m 程度日周鉛直移動したとい う報告がある(Shikata et al. 2017)。よって,本種は環境条件によって,同じ海域でも鉛直 分布パターンが変化すると考えられる。調査時,水温は24.1~29.5℃,塩分は 30.8~33.0(図 3),DIN および DIP 濃度はそれぞれ 0.14~31.1 µM および 0.02~0.72 µM の範囲で推移し た(図4)。栄養塩濃度は K. mikimotoi が比較的多く分布した表層付近では極めて低濃度で あり,細胞密度極大層におけるDIN および DIP 濃度はそれぞれ 0.18~2.02 µM および 0.04 ~0.20 µM で(図 4),ほとんどの時間帯において増殖に必要な濃度(半飽和定数,DIN: 0.78 µM,DIP:0.14 µM,山口 1994)より低かった。日射量(PAR,5 分ピッチで観測)は 最高で1,413 μmol m-2 s-1,水中の光量子束密度(1 時間平均値)は最高で 778 μmol m-2 s-1(図 5)で,日中時折,細胞は強光に曝されていた。細胞密度極大層における光量子束密度(2 時間平均値)は最高で320 μmol m-2 s-1であった(図6)。表層および細胞極大層における光 合成活性(Fv/Fm)はそれぞれ 0.44~0.57 および 0.45~0.57 の範囲で変動した(図 6)。Fv/Fm は,日中,光量子束密度が高まった7 月 24 日 12:00 および 16:00 は低い値(<5)を示し, 光阻害が起こっていたことが推察された。平成27, 28 年度の室内実験により,貧栄養塩環境 下でK. mikimotoi は日周鉛直移動を停止し,特に窒素欠乏条件下では昼夜一貫して表層に集 積すること,栄養塩欠乏状態で強光に曝されると細胞死が起こることが明らかとなっている。 上述の調査結果はこの室内で認められた現象が現場においても起こり得ることを示す。 2)K. mikimotoi 以外の有害赤潮生物の生残および光合成に及ぼす貧栄養および強光の影響 培養光の強度にかかわらず,C. antiqua の細胞密度は栄養塩無添加海水で培養開始 3 日後 から低下し始めた(図7)。また,栄養塩無添加海水での培養日数に関わらず,強光下での 光合成活性の低下についても相対的な差異は認められなかった(図8)。これらの結果から, K. mikimotoi とは異なり,C. antiqua は栄養塩の欠乏条件下で短期間内に死滅すること,強光 による相加相乗効果は明確でないこと,栄養塩欠乏状態で強光耐性は明確に低下しないこと が分かった。K. mikimotoi と C. antiqua の頻発する海域や時期は概ね異なることが知られて いるが,今回明らかとなった栄養塩欠乏や強光に対する耐性の差異が関与している可能性が ある。 3)塩分が N. scintillans の鉛直分布に及ぼす影響 異なる塩分条件におけるN. scintillans の鉛直分布の経時変化を図 9 に示す。N. scintillans の細胞極大層は,上下層とも塩分が33 の場合は実験開始 9 時間後に,上層塩分が 20 の場合 は16~18 時間後に表層に達した。N. scintillans 細胞は通常海水よりも比重が軽いために浮き やすいことが報告されており(Kiørboe 1998),本結果を支持する。一方,上層塩分が 10
の場合,正常な形態を有するN. scintillans 細胞は塩分躍層の下部にのみ分布したが,異常形
態を有するN. scintillans 細胞は全層にわたって観察された。Storm et al. (2013) は異なる強度
の塩分勾配下で有害赤潮ラフィド藻 Heterosigma akashiwo の捕食者である渦鞭毛藻類
Oxyrrhis marina,Gyrodinium dominans,有鐘繊毛虫類 Favella ehrenbergii の行動を観察した。
その結果,渦鞭毛藻類O. marina および G. dominans は塩分 5 および 10 の層においても分布
したが,有鐘繊毛虫類F. ehrenbergii は塩分 15 以上の層にのみ分布した。この既往知見を踏
まえると,実環境中において,N. scintillans は O. marina および G. dominans といった他の従
属栄養性渦鞭毛藻類に比べてより高塩分層に分布する可能性がある。また,K. mikimotoi は 10 以下の塩分層には進入せず,その直下の層に集積することが知られている(Shikata et al. 2014)。このことは,仮に現場海域において塩分 10 以下の水塊が上層に存在していた場合, N. scintillans と K. mikimotoi の分布深度が重なり,捕食圧が増す可能性が推察される。しか しながら,実験終了時点でN. scintillans 細胞の生残率は上下層の塩分がともに 33 の場合, ならびに上層塩分が20 で下層塩分が 30 の場合,それぞれ 86%,および 76%であったが, 上層塩分が10 の場合は 24%と低かった。また,上下層の塩分が 33 および上層塩分が 20 の 場合,異常形態を有するN. scintillans 細胞はほとんど認められなかったが,上層塩分の場合, 多くの細胞に収縮や破裂が認められた(図 10)。これらのことから,一部の細胞は上層に
生残不可(塩分10 以下,Lee and Hirayama 1992)の低塩分水が存在しても上層に浮上し,
死滅することが明らかとなった。今後,本研究と同様に様々な塩分下で実験を行い,N. scintillans の K. mikimotoi 捕食に及ぼす塩分の影響ついて,より定量的な解析を進めたい。 4)K. mikimotoi へのウイルス感染に及ぼす光照射の影響 K. mikimotoi にウイルスを接種したあるいはしなかった培養液について,異なる波長の光 を照射し,細胞密度を追跡した結果を図11 に示す。UV 領域である 320 nm(UV-B)照射区 では,ウイルスを接種した区でのみ,細胞密度が試験開始直後から減少し,3 日後には 35 cells mL-1となった。同様に,360 nm(UV-A)照射区では,ウイルスを接種した区でのみ,細胞 密度が3 日後には 65 cells mL-1まで減少した。一方で,440(青色領域), 600 nm(橙色領域) 照射区では,ウイルス接種区において細胞密度の減少は認められず,単色光を照射せず,ウ イルスを接種しなかった区(対象区)では,3 日後までに細胞密度が 1.5~2 倍に増加した。 実験開始3 日後のウイルス力価は 320 nm 照射区では 102から104 infectious unit ml-1へ,360 nm 照射区では102から103 infectious unit ml-1へ増加した。一方,440 と 600nm 照射区および通 常光下ではウイルス力価に変化は認められなかった。以上の結果より,ウイルスKmDNAV5 のK. mikimotoi への感染は紫外線により促進される可能性が示唆された。今後は,紫外線照 射強度あるいは量を実際に起こる値に調整するなど,実験系を改良するとともに,紫外線照 射がウイルス感染に与える影響について,生理的な側面からのアプローチも必要となるであ ろう。 5)豊後水道における K. mikimotoi の動態と環境条件との関係 宇和島湾および佐伯湾における2006~2017 年 5~8 月における K. mikimotoi の最高細胞密
度の変化を図12 に示す。宇和島湾および佐伯湾において,それぞれ 2007, 2009, 2012, 2014, 2015, 2016, 2017 年および 2007, 2008, 2009, 2011, 2012, 2013, 2017 年に最高細胞密度が 5,000 cells mL-1を超え,赤潮が発生した。また,宇和島湾では2007, 2014, 2015, 2016 年に,105 cells mL-1を超え,高密度化した。12 年間全ての年を対象として,最高細胞密度に達する 10 日前 から到達日までの降水量および日照時間の平均値を算出し,散布図を描いたところ(図13), 宇和島湾において日照時間が短い年に最高細胞密度が高い傾向が認められた。さらに,最高 細胞密度と降水量および日照時間について相関分析を行った。その結果,宇和島湾の場合, 最高細胞密度は日照時間と5%水準で有意な負の相関(Pearson の相関係数:-0.724,有意確 率:0.012)が認められたが,降水量とは有意な相関はなかった(相関係数:0.469,有意確 率:0.145)。一方で,佐伯湾の場合,最高細胞密度は降水量と 1%水準で有意な正の相関が 認められたが(相関係数:0.716,有意確率:0.009),日照時間とは有意な相関は検出され なかった(相関係数:-0.36,有意確率:0.25)。以上のことより,海域によって差異がある が,K. mikimotoi の発達に対して日照(光)は負の効果,降水量(出水,塩分,栄養塩濃度) は正の効果があることが分かった。これまでの本課題における室内外の実験より,貧栄養環 境下において強光はK. mikimotoi の生残に負の効果があることを見出してきた。今回の統計 解析結果はそのことを支持するものであり,光および栄養塩環境が本種の動態に多大な影響 を及ぼすことを強く示唆する。 3 5 ヵ年のまとめ 【背景】 近年,西日本においてK. mikimotoi 赤潮による甚大な被害が頻発している。本種赤潮は長 期化することも多く,魚介類のへい死という直接的な被害だけでなく,餌止めなどの対策を 講じることによって生じる「間接的な損失」も無視できない。そのため,現場から赤潮衰退 予測手法の確立が求められている。衰退の見通しが立てば,対策を最小限に留め,被害を減 じることができる。これまでK. mikimotoi について,増殖好適条件はある程度明らかとなっ ており,発生メカニズムも整理されている(山口 1994)。一方で,死滅条件や衰退メカニ ズムについては知見が少なく,体系化されているとは言い難い。本研究課題では,主に本種 赤潮の衰退過程に関与する環境条件を見出すことを目的として,室内外の調査実験を実施し た。 【Karenia mikimotoi の発生環境と衰退要因】 宇和島湾の 2006~2017 年のデータを解析したところ,日照時間が短いほど各年の K. mikimotoi 最高細胞密度は高くなる傾向が認められた。他種と比して非常に低い光量子束密 度で増殖可能で(山口 2014),日周鉛直移動を持続できる(Shikata et al. 2015)という本種 の生理特性はこの傾向と矛盾しない。また,K. mikimotoi 培養株を用いた室内研究結果によ り,本種は貧栄養(窒素欠乏)かつ強光条件下において,光阻害が誘導され,死滅すること を明らかにした。この現象は同じ赤潮鞭毛藻類C. antiqua では認められず,本種の生理特性 の一つであると考えられた。佐伯湾において実施した高日照時の現場ミクロコズム実験にお いても,3 m 層と比べて表層に係留した場合に増殖が低下し,死滅しやすいことをおさえた。 さらに,宇和島湾において大規模赤潮が発生した2014 年に高頻度の調査を実施したところ,
栄養塩濃度が低下し,梅雨明けして日照が急激に回復するタイミングでK. mikimotoi は急速 に衰退する過程を観測することに成功した。
【赤潮衰退時の日周鉛直移動】
K. mikimotoi は日周鉛直移動を行い,1 日で 20~25 m を往復できる(Koizumi et al. 1996)。
また,本種はいつも昼間に表層まで上昇するわけではなく,光合成や増殖に好適な環境(塩
分:Shikata et al. 2014,光:Shikata et al. 2017)などが存在する深度層に集積する,いわゆる
中層定位を行うことが知られている。このことから,実環境中で本種が長時間強光に曝され ることがあるのかどうか,疑わしい。しかしながら,本課題において複数の海域で昼夜連続 観測を重ね,本種は衰退直前の時期に鉛直移動を停止し,高日照下でも一日中表層に定位す ることがあることを突き止めた。さらに,室内実験によって,窒素欠乏が表層定位を誘導す る要因の一つであることを見出した。日周鉛直移動や中層定位の消失は,栄養塩や(適量の) 光の獲得を困難にするだけでなく,珪藻類などの競合生物,捕食動物や殺藻微生物などとの 接触頻度・時間を上昇させ得る。実際に現場観測および室内実験においてK. mikimotoi を捕 食するN. scintillans は昼夜問わず一途に浮上する,あるいは表層に分布する様子を観察し, さらに,表層にのみ存在する紫外線の照射でウイルス感染が誘発される可能性を室内実験で 見出した。 【結論】 本研究により,光および栄養塩環境はK. mikimotoi の衰退に深く関与していること,日周 鉛直移動の不調は衰退の重要なトリガーとなることが明らかとなった。よって,光,栄養塩 濃度,K. mikimotoi の鉛直分布パターンが本種の衰退を予知するための重要なパラメータで あることを提案する。今後,より定量的な解析を進め,赤潮予測技術の確立を目指す予定で ある。 謝辞 宇和島湾におけるK. mikimotoi の細胞密度データを提供して頂いた愛媛県水産研究センタ ーに感謝の意を表する。 引用文献
Koizumi, Y., Uchida, T., & Honjo, T. (1996) Diurnal vertical migration of Gymnodinium mikimotoi during a red tide in Hoketsu Bay, Japan. J. Plankton Res 18: 289-294.
Kiørboe T and Titelman J. (1998) Feeding, prey selection and prey encounter mechanisms in the heterotrophic dinoflagellate Noctiluca scintillans. J Plank Res, 20(8): 1615–1636. Lee, J., K. & Hirayama, K. (1992) Effects of salinity, food level and temperature on the population
growth of Noctiluca scintillans (Macartney). Bull. Fac. Fish. Nagasaki Univ. 71: 163-168.
宮村和良(2016)Karenia mikimotoi の赤潮動態と発生予察・対策.有害有毒プランクトンの
科学(今井一郎,山口峰生,松岡數充 編)pp. 191–200,恒星社厚生閣,東京.
Nakamura, Y. (1998) Biomass, feeding and production of Noctiluca scintillans in the Seto Inland Sea, Japan. J Plankton Res 20: 2213-2222.
Onji, M., Nakano, S. I., & Suzuki, S. (2003) Virus-like particles suppress growth of the
red-tide-forming marine dinoflagellate Gymnodinium mikimotoi. Mar Biotech 5: 435-442 Shikata, T., Onitsuka, G., Abe, K., Kitatsuji, S., Yufu, K., Yoshikawa, Y., Honjo, T., & Miyamura,
K. (2017) Relationships between light environment and subsurface accumulation during the daytime in the red-tide dinoflagellate Karenia mikimotoi. Mar Biol 164: 18.
Shikata et al. (2015) Diurnal vertical migration rhythms and their photoresponse in four phytoflagellates causing harmful algal blooms. Limnol Oceangr 60: 1251–1264. Shikata, T., Sakamoto, S., Onitsuka, G., Aoki, K., & Yamaguchi, M. (2014) Effects of salinity on
diel vertical migration behavior in two red-tide algae, Chattonella antiqua and Karenia
mikimotoi. Plankton Benthos Res 9(1): 42–50.
Storm, S., L., Harvey, E., L., Fredrickson, K., A., & Menden-Deuer, S. (2013) Broad salinity tolerance as a refuge from predation in the harmful raphidophyte alga Heterosigma
akashiwo (Raphidophyceae). J Phycol 49: 20–31.
山口峰生 (1994) Gymnodinium nagasakiense の赤潮発生機構と発生予知に関する生理生態的 研究. 南西海区水産研究所研究報告 27: 251-394.
A
B
YK-22
図1.八代海楠浦湾における調査定点. 103 102 10 1 (cells/mL) 0800 1200 1600 2000 2400 0400 0800 7/24 7/25 図2.八代海楠浦湾における K. mikimotoi の鉛直分布(コンター図)および 細胞極大層深度(折れ線グラフ)の日変化.Temperature Salinity Sigma-t (c) Chlorophyll Irradiance (μg/L) 0800 1200 1600 2000 2400 0400 0800 7/24 7/25 (kg/m3) 図3. 八代海楠浦湾における水温,塩分,Sigma-t,クロロフィル濃度の鉛直分布の日変化 (μM) 0800 1200 1600 2000 2400 0400 0800 7/24 7/25 DIN DIP N ut rie nt con cen trat ion ( µM ) 0 1 2 3 DIN DIP 図4.八代海楠浦湾における栄養塩濃度(DIN,DIP)の鉛直分布および K. mikimotoi 細胞極大層における栄養塩濃度の日変化.
0 500 1000 1500 a) (μmol/m2/s) 0800 1200 1600 2000 2400 0400 0800 7/24 7/25 103 102 10 1 b) 0 100 200 300 400 PA R (μ mo lm − 2 s − 1) c) 図5. 八代海楠浦湾における日射量(a),水中光量子束密度(b), K. mikimotoi 細胞極大層における平均光量子束密度(c)の日変化. 0.4 0.45 0.5 0.55 0.6 0 m 0.4 0.5 0.6 細胞極大層 8:00 12:00 16:00 20:00 0:00 4:00 8:00 Fv /Fm 図6. 八代海楠浦湾における表層および細胞極大層における光合成活性の日変化.
0 1 2 3 4 5 Incubation days Cell de ns ity (c ell s mL ‐1) 105 104 103
Complete medium, strong light Complete medium, weak light
Filtrated seawater, strong light Filtrated seawater, weak light
図7. C. antiqua の生残に及ぼす貧栄養塩および強光の影響. 0 20 40 60 80 100 0 30 60 90 120 Fv /Fm ( % ) time (min) 図8. C. antiqua の強光耐性に及ぼす貧栄養下での培養期間の影響. 判例は図7 と同じ.
0 20 40 60 80 0 10 20 30 0 20 40 60 80 Wa te r de pt h (c m) 0 20 40 60 80 0 2 4 6 8 10 12 14 16 18 20 22 24 Time (h) R e la ti ve c e ll den si ty of No ct ilu ca sc in tilla ns (% ) (a) (b) (c) (d) (e) (f) Salinity 図 9 . N. scintillans の鉛直分布に及ぼす塩分勾配の影響(a-c:塩分,d-f:相対細胞密度). a) b) c) 図 10.異なる塩分条件下(a:30,b:20,c:10)で観察された N. scintillans.
320 km 320 km+V5 360 km 360 km+V5 400 km 400 km+V5 600 km 600 km+V5 通常光 km 通常光 km+V5 K . m ik im oto i (c e lls /m L ) Incubation day 103 104 102 101 K m DNA V 5 t ite r (i n fe c ti o u s u n it /m L ) 103 104 102 101 0 0 Not measured 0 1 2 3 通常光 320 360 440 600 図 11. 光条件およびウイルス接種の有無が K. mikimotoi の生残増殖およびウイルス力価に及ぼす影響. 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 0 2 4 6 5/1 5/31 6/30 7/30 8/29 L o g n u m b e rs o f c e ll d e n s it y (c e lls m L -1) 2006 2007 2008 2009 2010 2011 2012 2013 2014 2015 2016 2017
Uwajima Bay Saiki Bay
図 12. 2006~2017 年の宇和島湾および佐伯湾における K. mikimotoi の動態 (宇和島湾データは愛媛県水産研究センターより提供).
0 2 4 6 8 10 12 0 100 200 300 0 2 4 6 8 10 12 0 100 200 300 M a xi m u m ce ll d e n si ty (ce lls m L -1 )
Duration of bright sunshine (hours) Precipitation (mm) Uwajima Bay Saiki Bay Uwajima Bay Saiki Bay 103 104 102 101 1 105 106 103 104 102 101 0 105 106 103 104 102 101 1 105 106 103 104 102 101 0 105 106 図 13. 宇和島湾および佐伯湾における K. mikimotoi の年間最高細胞密度(Y 軸)と 日照時間(X 軸左)および降水量(X 軸右)の関係.
2)カレニア等有害赤潮の高度モニタリング技術開発,発生機構解明,予察技術開発
および被害防止対策
④カレニア等有害赤潮鞭毛藻による魚介類斃死機構の解明と斃死防止技術の開発
ア
. カレニア等有害赤潮鞭毛藻による斃死原因の特定とアッセイ系の構築
水産研究・教育機構 瀬戸内海区水産研究所 外丸裕司,大久保信幸,持田和彦 高知大学 山口晴生 1 全体計画 (1)目的 近年,西日本の海域で有害 Karenia 属等鞭毛藻による赤潮が頻発し,養殖魚類を斃死させ るなどの深刻な被害をもたらしている。このような赤潮による漁業被害を未然防止および軽 減するためには,Karenia 属等による魚類へ毒性を評価するとともに,その斃死機構を解明 する必要がある。しかしながら,Karenia 属等鞭毛藻による魚介類の斃死現象を室内で安定 的に再現することは,これまで様々な理由によりきわめて困難であった。そのため,有害赤 潮鞭毛藻による魚類に対する毒性評価法は未だ確立されていない。本課題では,天然赤潮に 依存しない Karenia 属等鞭毛藻の培養細胞を用いた,安定かつ再現性の高い魚類斃死試験法 ならびに魚毒性評価法の確立を目指す。 (2)試験等の方法 1)Karenia 属等の培養株の確立と簡易大量培養法の開発 西日本沿岸を中心に発生する,Karenia 属等の培養株を確立するとともに,それらの増殖 等諸性状に基づいた簡易大量培養手法の開発を行う。その上で選抜無菌株を用いた魚介類 ならびに魚類鰓培養細胞等への曝露試験によって,魚介類斃死能を有する無菌培養株の確 立,ならびにそれを安定かつ大量供給可能なシステムの構築を図る。 2)鰓培養細胞を用いた魚毒性評価手法の開発 マダイ鰓培養細胞等を用い,Karenia 属を含む有害赤潮藻類等の培養株の毒性を評価する。 さらに,マダイ以外の魚種に合わせた鰓細胞培養法の開発に向け,鰓細胞の無菌的調整法, 培養温度,ならびに培養プレートの種類等を検討する。最終的に,これまでに得られた研 究成果のとりまとめ,本開発に関する今後の実用化に向けた総合考察を行う。 2 平成29 年度計画及び結果 (1)目的 全体計画と同じ (2)試験等の方法 1)Karenia 属等の培養株の確立と簡易大量培養法の開発 ①本邦沿岸における推定 Kareniaceae 藻の培養株確立2017 年の夏季,高知県浦ノ内湾において一見すると Karenia 属と思われる渦鞭毛藻が 発生した。当該藻の魚類斃死能を明らかにする一環として,分離培養に成功した数株の rDNA 部分配列を決定し,系統学的な位置を推定した。 ②Karenia papilionacea 新奇系統の大量培養系確立ならびに魚類斃死能 2012 年に高知県野見湾より分離された K. papilionacea 新奇系統の有菌クローン株 KspNOM1H を実験に供した。平成 25 年度の本事業で確立した方法により当該株を大量 培養した。曝気速度は 7.8 mL/s とした。この培養期間中,経時的に細胞密度・クロロフ ィル蛍光量を測定した。また,同様に Karenia mikimotoi および K. papilionacea 原記載種
系統を培養し,得られた各種細胞を含む培養液(104 cells/mL)ならびに IMK 培地をそ れぞれガラス製透明水槽に移し,そこに体長 10~13 cm のマアジ(Trachurus japonicus) を水槽当たり 5 尾曝露した。その後,供試魚の行動・斃死状況を観察した。 2)鰓培養細胞を用いた毒性評価法の開発 ①Karenia mikimotoi の毒性が変化する培養条件の探索 K. mikimotoi KmURN6Y 株の毒性が培養液中の栄養塩濃度で変化するか調べるため, 通常の SWM-3 培地(通常区),窒素濃度を 1/100 にした窒素欠乏 SWM-3 培地(N 欠区), リン濃度を 1/65 としたリン欠乏 SWM-3 培地(P 欠区)でそれぞれ培養し,同じ培養フ ラスコから培養 17 日目,24 日目,32 日目,38 日目の赤潮細胞をマダイ鰓培養細胞に それぞれ曝露し,鰓細胞の生残率を指標としてその毒性を比較した。統計解析は一元配 置分散分析を行い,その後,Bonferroni 法による多重比較検定を実施した。また,デー タの等分散が仮定できない場合はクラスカル=ウォリス検定を行い,その後,Steel 法に よる多重比較検定を実施した。 ②新奇有害赤潮藻の毒性評価 新奇有害赤潮藻 Takayama 培養株の鰓細胞への毒性の有無について明らかにするため, SP2(13.5 × 104 cells/mL),SP5(13.8 × 104 cells/mL),C4(11.4× 104 cells/mL)の 3 株を マダイ鰓培養細胞にそれぞれ曝露し,その毒性を評価した。また,K. papilionacea KpURN9Y 株について,KmURN6Y 株と同様に培養ステージで毒性が変化するか調べる ため,同じ培養フラスコから培養 12 日目(2.0 × 104 cells/mL),19 日目(2.6 × 104 cells/mL), 25 日目(3.0 × 104 cells/mL),32 日目(2.2 × 104 cells/mL)の赤潮細胞をマダイ鰓培養細 胞にそれぞれ曝露し,その毒性を評価した。(統計解析法は①と同様)。なお,KpURN9Y 株の培養温度は 25℃,マダイ鰓細胞の培養温度は 20℃のため,曝露試験は両温度で実 施した。 ③マダイ以外の魚種に合わせた鰓細胞培養法の開発 マダイ鰓培養細胞作製法(Ohkubo et al. 2017)を基本とし,マグロ類のスマ幼魚 4 尾 (平均全長 15.7 cm)を用いて鰓培養細胞の作製法を検討した。今年度は細菌の混入を 防ぐため,紫外線殺菌装置(REI-SEA UVF-1000)を取り付けた陸上水槽でスマ幼魚を 3 日間畜養した後,培養に用いた。また,培養鰓細胞の基礎的な性質の確認として,培養 マダイ鰓細胞が生体の鰓細胞と形態的にどの程度同じなのか,走査型電子顕微鏡での観 察,および多糖染色(アルシアンブルー液・コールドシッフ試薬重染色)を実施し,粘 液細胞の有無についても検討した。 ④赤潮発生地域での毒性評価に向けた検討
赤潮発生地域での毒性評価を目指し,鰓培養細胞の輸送試験を行った。温度変化を避 けるため,鰓細胞を培養している容器をクーラーボックスに入れ,瀬戸内水研から自動 車およびフェリーで愛媛大学南予水産研究センターまで輸送した。その後,20℃のイン キュベータに収納し,翌日,凍結保存した K. mikimotoi 株を用いて先方の実験室内で曝 露試験を実施した。 (3)結果及び考察 1)本邦産 Karenia papilionacea 系統の増殖特性と魚毒性 ①本邦沿岸における推定 Kareniaceae 藻の培養株確立
rDNA 配列に基づいた系統解析の結果,分離株の多くが Takayama xiamenensis の原記 載種(TPXM,Gu et al. 2013)の配列と完全に一致する LSU rDNA を有していた(図 1)。 また一株は Takayama tasmanica と同系統に位置し,少なくとも Takayama 属二種が本邦 沿岸域に分布していることを明らかにした。
わが国では Gymnodinium sp. ‘84-K として認識されていた渦鞭毛藻が,その形態学的 特徴から,Takayama pulchella として転属・種記載されている(de Salsa et al. 2003)。本 種は 1984 年および 1985 年に鹿児島湾にて大発生したことがあり,その時の細胞密度は
2.7 × 105 cells/mL に達したと記録されている(福与ら 1990)。それに伴い漁業被害も報
告されていることから,本種は有害種であるものと考えられている。しかしそれ以降,
Takayama 属の発生はほとんど公表されていない。先に開催された EAST HAB では,フ
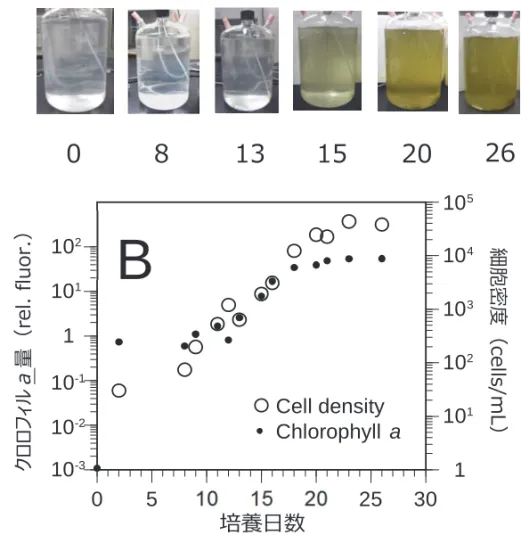
ィリピンにおける T. xiamenensis のブルームとそれに伴う漁業被害が Benico 氏らにより 報告されており,本種を有害種として認識する必要があるかもしれない。このことを受 けて,本研究により T. xiamenensis および T. tasmanica を見出し,今後それらの魚類斃死 能の解明に必要となる培養株を確立できたことは極めて意義深い。近く,確立された培 養株の魚類斃死能を試験する必要がある。 ②Karenia papilionacea 新奇系統の大量培養系確立ならびに魚類斃死能 2015 年度の事業で開発した培養系により K. papilionacea 新奇系統も大量培養できる ことが明らかとなった。KspNOM1H 有菌株を接種した培地は,培養 15 日目から黄緑色 に淡く色づきはじめ,培養 25 日目には濃い緑褐色に色付いた(図 2)。本藻は,最大 1.0 division/day のスピードで増殖し,培養 23 日目には最大収量 4.3 × 104 cells/mL に達した (図 2)。昨年度,同新奇系統の大量培養には予備的に成功していたが,定量的な結果を 得るには至っていなかった。今回の結果を踏まえると,この大量培養系を用いることで, 従 来 の 対 象 種 K. mikimotoi お よ び K. papilionacea 原 記 載 種 系 統 だ け で な く , K. papilionacea 新奇系統をも赤潮状態まで大増殖可能なことが実証されたと言える。なお 新奇系統の無菌化には未だ成功していないものの,有菌状態であっても,十分な細胞収 量まで培養可能であった。とは言え今後,コンタミのリスクを軽減するためにも無菌化 を行うことは重要である。 本培養系により,K. mikimotoi および K. papilionacea 原記載種系統・新奇系統を大量培 養し,魚類へ曝露しようとした。しかしながら,陽性試料として位置付けられる K. mikimotoi については,早期にフロック(細胞凝集体)が発生したことで,大量の細胞を 他藻と同期的に調製・確保することができなかった。このような同期的確保にあたって
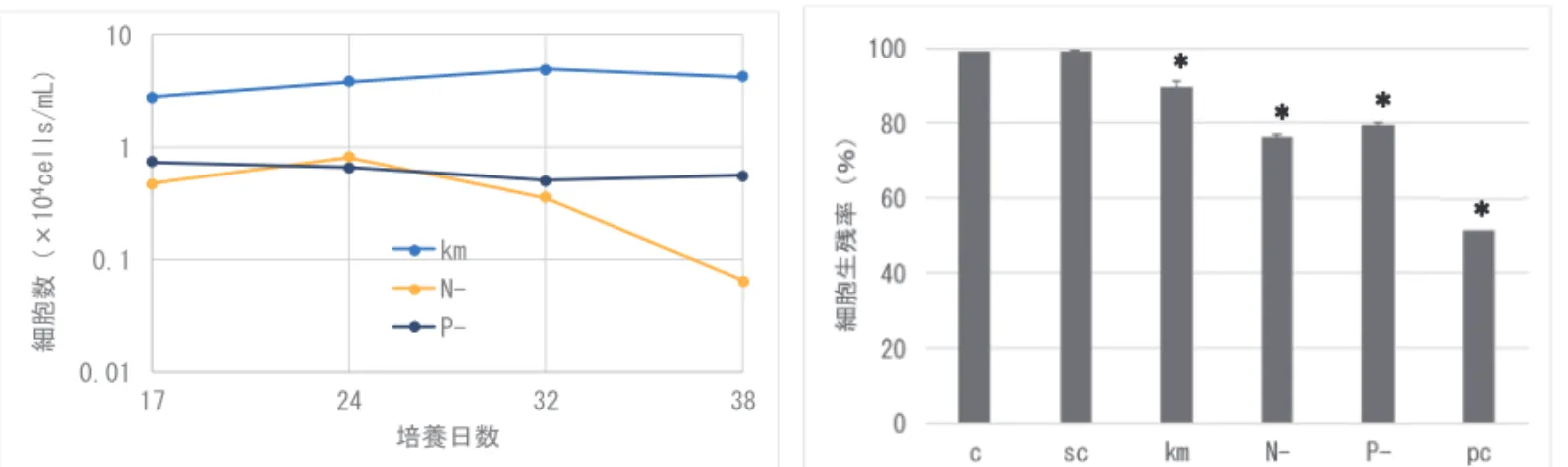
は,まずコンタミの影響を軽減するために無菌株を用いることが望ましく,さらに培養 状態を制御することが重要である。今回のようなケースでは,曝気速度を低下させるこ とで,K. mikimotoi の増殖を抑えることが有効であったかもしれない。このような措置 を施すために,今後,Karenia 属の細胞状態を簡易に評価可能な手法を確立することが 求められる。 今回の魚類曝露試験では,K. papilionacea 二系統いずれも斃死魚が認められなかった (図 3)。陽性試料であった K. mikimotoi を同期的に確保できなかったため,試験結果を 評価することは難しいものの,なんらかの原因で供試株の魚類斃死能がかなり弱いこと, あるいは魚類に毒性耐性が備わっていた可能性が考えられる。また,新奇系統の無菌株 を確立できていないことも考慮する必要がある。再度試料を調製し,これら二系統の毒 性を評価する必要がある。 2)マダイ鰓由来の培養細胞を用いた毒性評価法の開発 ①Karenia mikimotoi の毒性が変化する培養条件の探索 K. mikimotoi 株の細胞数は,通常区では培養 32 日目にかけ 4.9 × 104 cells/mL まで増加 した後,ゆるやかに減少した(図 4 左)。N 欠区では 24 日目にかけ 8.2 × 103 cells/mL ま で増加した後,減少し,38 日目には 7.0× 102 cells/mL となった。また,P 欠区では特に ピークは認められず 7.3~5.1× 103 cells/mL で推移した。N 欠区や P 欠区で培養した K. mikimotoi 株は,細胞密度は通常区で培養したものに比べて低く,例えば 32 日目では N 欠区が 1/14,P 欠区では 1/10 であったが,培養液全体の毒性は同程度か,より強くなっ ており(図 4 右),1 細胞あたりの毒性が強く発現していたことが示唆された。 ②新奇有害赤潮藻の毒性評価 Takayama 培養株のうち,C4 株では鰓細胞の生残率が 86.4%に有意に低下し(p=0.03), 若干の毒性が認められた(図 5)。このことから本種について高密度化した場合には注 意が必要と考えられた。K. papilionacea 株の細胞数は培養 25 日目にかけてゆるやかに 増加した後,減少した。一方,培養液全体の毒性は,培養 25 日目までは検出されず, その後,最終盤の 32 日目に 20℃での曝露試験で,わずかに鰓細胞の生残率の減少(96.5%) が観察された(図 6)。このことから本種については鰓細胞への毒性はほとんど無く, 以前検出された魚への毒性は他の要因によるものではないかと推察された。 ③マダイ以外の魚種に合わせた鰓細胞培養法の開発 スマ幼魚 4 尾(図 7)を用いて鰓細胞の培養を試みたところ,細菌の混入もなく十分 量の鰓細胞の単離,培養に成功した。鰓細胞は順調に増殖し,1 週間以内にコンフルエ ント(培養容器を覆いつくす状態)となった。また,この初代培養細胞に K. mikimotoi 株を曝露したところ,マダイ鰓細胞と同様に感受性を示した。しかし,スマ鰓細胞の継 代培養はできなかった(表 1)。今後,継代培養法を確立し,毒性アッセイを安定的に 実施するには,培養方法や継代時の酵素による細胞分散方法について引き続き試行錯誤 が必要と思われる。マダイ培養鰓細胞の電子顕微鏡観察では,マイクロリッジ等の構造 が観察され(図 8),典型的な魚類鰓上皮細胞像を示した。また,多糖染色で濃染され た中性粘液塊も観察され,粘液細胞の存在が示唆された。これらの結果,本鰓細胞培養 系が魚類鰓組織へのダメージを調べるモデル試験系として有用であることが再確認さ
れた。 ④赤潮発生地域での毒性評価に向けた検討 鰓細胞の輸送時間は 6 時間,温度変化は+3℃以内であった。輸送した鰓細胞および 凍結 K. mikimotoi 株を用いた曝露試験は特に問題無く実施可能であった(図 9)。 スマ幼魚の供与,および鰓細胞の輸送試験にご協力いただいた愛媛大学南予水産研究 センター 松原孝博教授に感謝いたします。 3 5ヵ年の成果 (1)Karenia 属をはじめとする Kareniaceae の培養株を確立 西日本沿岸域においては,K. mikimotoi のみならず K. papilionacea が広域にわたって分布し ており,海域によっては分布を共にすることも明らかにした。また,本邦において発生した Takayama 属を分離することに成功し,これらの培養株を確立することができた。 (2)室内で魚類を斃死させる Karenia 属藻の培養株を確立 赤潮原因藻の魚類斃死機構を解明するためには,厳密に制御可能な室内環境において当該 藻を「魚類斃死能を維持したまま」培養することが重要である。本課題では,要件を満たし た K. mikimotoi 株を確立し,当該株の魚類斃死作用とともにエラ細胞への壊死作用も矛盾な く見出すことができた。このように課題の中核的な目的を達成できたことは,今後の魚類斃 死機構の解明を実現する上で極めて意義深いと成果と言えよう。 (3)Karenia 属の大量培養系確立 最終年度をもって Karenia 属三種の大量培養系を構築することができた。天然熟成された 海洋深層水を培地のベースに用いることで,多大な労力を有する海水採取のみならず数か月 にわたる熟成操作も不要であった。さらには海水が清浄性に富んでいるため,煩雑極まりな い海水のろ過操作も不要となり,従来よりも遥かに高効率な大量培養系を実現できた。さら に各藻の増殖に及ぼす水温,塩分,光条件を明らかにし,増殖予測に欠かせない増殖速度・ 最大収量などの増殖動力学パラメーターも得ることができた。これにより従来よりも格段に 効率よく,かつ計画的に,大量の Karenia 属藻を調製することが可能となった。 (4)海産魚類の鰓細胞培養法の確立と高度化,簡便化 飼育管理の容易さ,培養に使う魚のコンディションの均一化等のため,マダイ 0 歳魚を用 いた鰓細胞培養法を確立した(H25)。さらに,均質な鰓細胞を多数準備するための継代培 養法の開発を確立するとともに,凍結保存法についても検討した(H26)。なお,培養した 鰓細胞は上皮細胞や粘液細胞を保持しており,生体の鰓細胞組織をかなり再現していること が示唆された(H29)。 また,他の養殖対象魚を用いた鰓細胞培養法の開発としてブリ(H26),ヒラメの代替と してホシガレイ(H27-28),近年養殖量が急増しているマグロ類(クロマグロ(H26),ス マ(H27-29))について検討した。養殖魚から培養細胞を作製する際は細菌の混入が大き な問題となったが,事前に減菌した海水で畜養し,小型の個体を複数尾使う等の改良により, ホシガレイおよびスマでは初代培養が,ブリでは継代培養(3 代)まで可能となった。なお, マグロ類の鰓細胞培養については特に難易度が高く,培養法について引き続き検討が必要で ある。 (5)鰓培養細胞を用いた赤潮株の毒性評価法の開発
本課題での毒性評価は,海域での K. mikimotoi 赤潮により養殖魚の鰓組織が破壊される被 害を想定し,鰓細胞に有害赤潮藻類を曝露した際,鰓細胞が死ぬ,あるいは培養容器から剥 がれることを指標とした。初年度は曝露後に生き残った鰓細胞数をカウントして毒性の指標 としたが,2 年目からはより客観的で,作業者に負担の少ない方法として赤潮藻を曝露後, 生き残った鰓細胞を写真撮影して生細胞が占める面積を画像解析で求める方法を確立した。 この方法で K. mikimotoi 株のほか,7 種の新奇赤潮培養株(K. papilionacea,K. cf. papilionacea,
Cochlodinium polykrikoides,Heterosigma akashiwo(H26), Dictyocha(H27), K. digitata(H28), Takayama (H29) )の毒性を評価した。その結果,K. digitata 株は K. mikimotoi 株と同程度の
強い毒性を示し,K. cf. papilionacea 株と Takayama 株では比較的弱い毒性を持つことが示さ れた。 (6)K. mikimotoi 株の鰓細胞への毒性作用機序解明に向けた検討 (5)で確立した鰓細胞毒性評価法を用い,K. mikimotoi 株の毒性作用機序解明に向けて以下 を検討した。はじめに毒性発現部位の探索として,培養した K. mikimotoi 細胞と培養液を分 けて鰓細胞にそれぞれ曝露したところ,K. mikimotoi 細胞のみで毒性が認められた(H26)。 なお,この現象は K. cf. papilionacea 株,および K. digitata 株でも観察された。さらに,K. mikimotoi 細胞をホモジナイズし,遠心分離で細胞器官を粗分画したところ,細胞壁等を含 む沈殿部分に毒性が最も強く認められた(H27)。これらのことから,K. mikimotoi 細胞と鰓 細胞が接触することで毒性が発現することが強く示唆された。なお,K. mikimotoi 細胞のホ モジナイズを-80℃で凍結後,解凍したものは毒性を保持するが,100℃で加熱すると毒性を 失うことも示され,毒性の原因物質は比較的不安定であることが示唆された。 次に K. mikimotoi 株の毒性が変化する培養条件の探索として,塩分濃度(H26),培養ステ ージ(H28),栄養塩濃度(H29)について検討した。SWM-3 培地の塩分を 20 と 30 に調整 し,培養しても毒性の差は認められず,培養ステージでは終期に細胞密度増加と相まって毒 性が強まることが示唆された。また,窒素やリンの濃度を下げた SWM-3 培地で培養した K. mikimotoi 株は,細胞密度は通常の SWM-3 培地で培養したものに比べて低いが,鰓細胞 への毒性は同程度か,より強く発現することが示された。 (総括) 本邦では今なお,K. mikimotoi などの有害赤潮が頻発しており,それに伴う漁業被害はあ とを絶たない。また近年は,海外の K. papilionacea(原記載種系統)培養株から,魚介類等 を死に至らしめる神経毒“ブレベトキシン”が検出されており(Fowler et al. 2015),それと 同系統のものが西日本にも分布していることが明らかとなった。漁業被害の軽減・防止策の 構築のため,Karenia 属の赤潮と魚類の斃死との関係を明らかにすることは喫緊である。し かし従来までは,自然環境で赤潮が発生するのを待ち構えるのが常であり,魚類に対する赤 潮の斃死作用の評価は遅々として進んでいなかった。このような背景の下,我々は 5 ヵ年に わたる研究活動を通じて,本邦 Karenia 属の魚類斃死の解明に関わる基盤的知見とともに新 技術を整備することができた。これにより,従来よりも計画的に,かつ周年にわたって魚類 斃死アッセイならびに赤潮藻の毒性評価を実施することが可能となり,赤潮と魚類斃死現象 を関連付けて理解することに大きく貢献するものと期待される。
4 引用文献
de Salas MF, Bolch CJS, Botes L, Nash G, Wright SW, Hallegraeff GM. Takayama gen. nov. (Gymnodiniales, Dinophyceae), a new genus of unarmored dinoflagellates with sigmoid apical grooves, including the description of two new species. J. Phycol., 39, 1233–1246, 2003.
Fowler N, Tomas C, Baden D, Campbell L, Bourdelais A. Chemical analysis of Karenia papilionacea. Toxicon, 101, 85–91, 2015.
福代康夫,高野秀昭,千原光雄,松岡數充.「日本の赤潮生物-写真と解説-」内田老鶴圃, 東京,1990.
Gu, H., Luo, Z., Zhang, X., Xu, B., Fang, Q. Morphology, ultrastructure and phylogeny of Takayama
xiamenensis sp. nov. (Gymnodiniales, Dinophyceae) from the east China sea. Phycologia 52, 256–
265, 2013.
Larsen, J. Unarmoured dinoflagellates from Australian waters II. Genus Gyrodinium (gymnodiniales, Dinophyceae). Phycologia 35, 342–349, 1996.
Ohkubo N, TomaruY, YamaguchiH, KitatsujiS, Mochida K. Development of a method to assess the ichthyotoxicity of the harmful marine microalgae Karenia spp. using gill cell cultures from red sea bream (Pagrus major). Fish. Physiol. Biochem. 43, 1603–1612, 2017.
Yamaguchi H, Hirano T, Yoshimatsu T, Tanimoto Y, Matsumoto T, Suzuki S, Hayashi Y, Urabe A, Miyamura K, Sakamoto S, Yamaguchi M, Tomaru Y. Occurrence of Karenia papilionacea (Dinophyceae) and its novel sister phylotype in Japanese coastal waters. Harmful Algae, 57, 59–68, 2016.
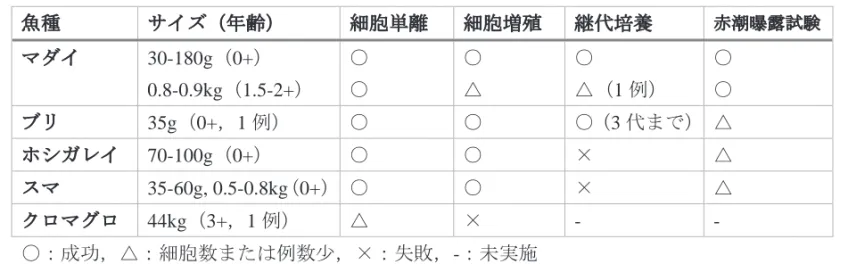
表 1 これまで瀬戸内水研で鰓細胞培養を実施した海産魚. 魚種 サイズ(年齢) 細胞単離 細胞増殖 継代培養 赤潮曝露試験 マダイ 30-180g(0+) 0.8-0.9kg(1.5-2+) ○ ○ ○ △ ○ △(1 例) ○ ○ ブリ 35g(0+,1 例) ○ ○ ○(3 代まで) △ ホシガレイ 70-100g(0+) ○ ○ × △ スマ 35-60g, 0.5-0.8kg(0+) ○ ○ × △ クロマグロ 44kg(3+,1 例) △ × - - 〇:成功,△:細胞数または例数少,×:失敗,-:未実施 図 1 浦ノ内湾産 Takayama 属培養株(太字英数字)の系統学的位置づけ.ML 系統樹の 分岐点にはブートストラップ値(n = 1000)が示されている.
図 2 Karenia papilionacea 新奇系統 KsNOM1H の大量培養時における培養状況(A,数値 は培養日数を示す)ならびに細胞密度・クロロフィル a 量の経時的推移(B). 図 3 浦ノ内湾産 Karenia papilionacea 二系統を曝露したときのマアジ生残数の推移.
培養時間(分)
0 30 60 90 120 150 180 0 1 2 3 4 5⽣残数(尾)
⾮曝露区
0 1 2 3 4 5曝露区
( 原記載種系統)曝露区
( 新奇系統)⽣残数(尾)
0
8
13
15
20
26
1 101 102 1 101 102 103 104 105 0 5 10 15 20 25 30 Chlorophyll a Cell density 10-2 10-1 10-3培養⽇数
細胞密度 (c ells/mL) クロロフィル a 量(rel. fluor.)A
B
図 4 Karenia mikimotoi 株の細胞数の変化 (左),32 日目の培養液をマダイ鰓細胞に曝露した 際の鰓細胞の生残率(右).km:SWM-3 培地で培養した KmURN6Y 株(4.9× 104 cells/mL), N-:窒素欠乏 SWM-3 培地で培養した KmURN6Y 株(0.4 × 104 cells/mL),P-:リン欠乏 SWM-3 培地で培養した KmURN6Y 株(0.5 × 104 cells/mL),c:L15 培地,sc:SWM-3 培地,pc:陽性 対照(凍結 KmURN6Y 株)。*:sc と曝露区の生残率との間に有意差あり(p<0.05).グラフ 上のバーは標準誤差. 図 5 Takayama 培養株 C4(11.4× 104 cells/mL), SP2(13.5 × 104 cells/mL),SP5(13.8 × 104 cells/mL)をマダイ鰓培養細胞にそれぞれ曝露 した際の鰓細胞の生残率.swm3:SWM-3 培 地,*:sc と曝露区の生残率との間に有意差 あり(p<0.05).グラフ上のバーは標準誤差. 図 6 Karenia papilionacea 株の細胞数の変化 (左),32 日目の培養液を 20℃でマダイ鰓細胞に 曝露した際の鰓細胞の生残率(右).kp:K. papilionacea 株(2.3 × 104 cells/mL),c:L15 培地, sc:F/2 培地,pc:陽性対照(凍結 KmURN6Y 株). 0 0.5 1 1.5 2 2.5 3 3.5 12 19 25 32 細胞数 (× 10 4ce lls /m L) 0 20 40 60 80 100 c sc kp pc 細 胞生残率(%) * * * * 0 20 40 60 80 100 swm3 c4 sp2 sp5 細胞 生残率 (%) * 0.01 0.1 1 10 17 24 32 38 細胞数( ×1 0 4cel ls /mL ) 培養日数 km N-