NII-Electronic Library Service J.CropRes,
55
:33-38
==
Research
Article
(2010)
Main
Effects
efIndiyidual
QTLs
andInteractions
between
Tvve
QTLs
Controlling
Grain
Length
ofRice
underNear-Isogenic
Lines
Tsuneo
Kato
Faculty
ofBiology-Oriented
Scienceand 7lechnologMKInkiUnivensity<Kinokawa,
Wakayama,649
-
6493,
Japan)Summary:
Tlie
objectiyes ofthis
study were to evaluatethe
main effects and interactionsDf individualquantitativetraitloci
(QTLs)
forgrain
lengthand theircombinationsin
rice.Fiveprimarynear-isogenic lines(NILs),
each of which hadan alleleforlonggrainonly atone of fiveQTLs
controlling grainlengthof alarge-graincultivar,
`BG
1',and an allele forshort grainlengthat the other fourQTLs,
were constructed with a common geneticbackgroundof a small-grain cultivar,`Gimbozu'.
Illaddition, an NIL'involvinga major genefor
long
grainof `Fusayoshi',L.of,
inthesamebackground
as above was developed.From intercrossingof theseprimaryNILs,phenetypic selection,and progenytests,15advanced NILs,each of which haddifferentalleles for
longgrainonly atone of
the
10
combinations offiveQTLs,
and one of thefivecombinations of fiveQTLs
andLkXwere furtherconstmcted. Themain effects
(additive
effects) and interactions(epistatic
interactions)ofQTLs
s t
and Lefon grainlengthwere estimated fromthemean grainlengthsof theseNILs and Gimbozu cultivatedover two years.The main effect of LigCwasthelargest,foIIowedbythosethe
QTLs
showing a ]argeand signMcant variation among them. T:hecorrelation coefTicient betweenthetwo yearsforthemain effects was highand significant(r=O.95"),
suggesting thattheQTLs'
main efEects were consistently expressed attheir
own magnitudes forthetwo differentyears.Theinteractions
between
twoQTLs
were abo'uthalf
the
strengthofthe main effects intheirmagnitudes and aisoshowed a large,significant variation amongQTLs.
The
year-to-year corre!ation forthe interactionswas, however,not significant(r=O.63),
Theseresults suggested thatQTL-by-Q[[IL
interactionscould contribute totheinstabilityof character expression and could playan importantrole ingenetic
systems controllingquantitative
traits.Key words: grainlength,
interaction,
rnain effect, Oryzasativa L.,QTL
lntroduction
Quantitative
traitsare geneticallycontrolled by multiplequantkatiyetraitloci
(QTLs),
each of whichgenerally
shows a smaller effect comparecl with non-genetic factorsand acts ina cumulative manner with otherQTLs
cencerned with thesarnetrait.These characteristicsof quantitativetraitshaye made it
difficult
to estimate indiyidualgenotypesfrom
observedphenotypes,which show a clear contrast to qualitatiyetraits contrelled by major genes.Tlierefore,several statistical
parameters,variance, covariance, mean, etc., hewebeenadopted as indispensableprocedures to analyze and deal with
quantitativetraitsinbreeding
(Kearsey
and Pooni1996),
The discoveryand developmentof various molecular markers, rnost of which are based on polymorphisms innucleetide sequences, gavearevolutionary impacton theabove situation: theindividualgenotypes concerned with aquantitativetraithave beenidentifiedclearIy, atleastforone ofthemultiple
QTLs
forthistrait,basedon closely linkedmolecular markers thatbehave
Acccepted:March15,2010
Correspending author :TsuneoKato
([email protected])
as typicalqualitativetraits.Ihisenables us toposition
QTLs
individuallyon an appropriate molecular linkagemap and to estimate theirgene actions and interactionsthroughseveral
algorithms
(Lander
and Botstein1989,
Thriksley1993,
Tbnksley and Fulton2007).
On theother hand,thedetectionofQTLs
strongly depends upon the levelof the threshold(LOD
threshold,etc.) which can be determinedrather arbitrarily by experimentei's,
This
is
aseriousproblem,particularlyfor
QTLs
with small effectsand
QTLs
linkedcloselytoeach other(Yano
and Sasaki199b.
Thcrrefore,the existence of
QTLs
should be confirrnedthroughother proceduresand materjals, such as near-isogenic
lines
(NILs)
(Lin
etat,2003),
chromosome segment substitutionlines
(Kubo
et at.2002),
etc, Sefar,alarge number ofQTLs
havebeendissectedindividuallyusing theseadvanced materials and identifiedas a definiteregion inthe genome invarieus organisms
(e.g.,
Fan et al.2006,
Song et al.2007),
However, such dissectionof rnultipleQTLs
intoasingle factorinevitably makes itimpessibletoeyaluate anyQTL-by-QTL
interactions, which are another importantcharacteristic ofQ[[Ls.
Therefore,pyramidingof thesedissected
QTLs
forthesame traitintoanCopyrighturasMlop・
veptWfftk(Tlte
Seciety of CropScience and Breeding inKinki,Japan)The Society of Crop Science and Breeding in Kinki, Japan
TheSociety ofCrop Science and Breeding in Kinki, Japan
J,
Crop
Res,55(2010)
NIL
is
necessary toanalyze theinteractionsamongQTLs,
Previousreports by the
present
author havedealt
with the grainlengthof rice(Oryza
sativa L.)as a typical quantitative traitand havetriedto dissectfiveQTI.s
controlling grainlength of a 1arge-graincultivar `BG1'
(Kato
1998,
Kato2005).
Consequentiy,a series ef fiveNILs with a cornnion genetic
background of a cultivar 'Gimbozu' haye already been
developed:each of thefiveNILs has an aileleforlonggrain
derivedfrom `BG
1'
at only ene of thefive
QTLs
andhas
the counterpart alleles forshort grainderivedfrom`Gimbozu'
atallfouTother
QTLs
(Kato
2005).
From intercrossjngof theseprimaryNILs
for
rice grainlength,thepresentstudy further constmcted aseriesof10
advanced NILs, each ofwhich had twoalleles forlonggrainatonlyone combination oftwo
QTLs
andhad thecounterpart alleles fbrshort grainat the threeother
Q[['Ls,
The objectives of the presentstudy were to investigatethe main effects
(additive
efiiects)of individualQTLs
for
grainlengthand theinteractionsbetweentwo
QTI.s
among thefive, using theabove primaryand advanced NILs, inaddition, the main effect of amajor geneforgrainlength,LdyConchromosome3
(Takeda
and Saito1980)
and the interactionsbetween theindividual
QTLs
and Lkfwere alsoevaluated using NILs ofthe
same kind.Several
novel characteristicsfor
the
QTLs'
main effects andQTL-by-QTL
interactions
for
rice
grainlength
arediscussed.
Materials
andMethods
Five
primaryNILs
for
rice grainlength
were eonstructed through theproceduresdescribed
previously(Kato
1998,
Kato2005).
Inthisprocess,recombinantinbred
lines(RILs)
weredevelopedatfirstfromacross between`BG 1'and
'Koshihikarj',
a cultivar with short grainlength,Each of fiyeRILs showingt
t;tt
differentgrarnlengthswas crossed with Gimbozu , another
cultivar withshert grain length.Segregantswith thelongest
graln length in each F2 populationwere successively
backcrossedwith therecurrent parent,`Gimbozu', as agenetic
background.Consequently,fourpopulationsof BCsF2and one
BC4F2,each of which showed a typical segregation forgrain
lengthas a single Mendelianfactor,were obtained
(Kato
2005).FiveNILs were established as true-breeding linesfrem the
respective ]ongest-grainsegregants inthe F2populations,and
designated
asNIL-a,
NIL-b,
NIL-c,
NIL-d,
andNIL-e,
Fromiso-allelismtestsforallpossiblecombinations
(data
not shown), alleles forlonggraininthesefiveNILs were confirmed toercist atdifferent
QTLs.
Tliesealleleswere recessive against thealleles
for
shortgrainin
al1
QTLs.
TheseQTLs
were tentatively designated as qGin, gGth, qGhe, qGLd. and qGLe forNIL-a,NIL-b,NIL-c,NE.-d,and NIL-e,respcctively. Inthesame way,
a large-graincultivar `Fusayeshi', which hasa major
gene
for
long
ginin,
LkA hasbeensuccessivelybackcrossedinfivetimestl
tt
tt
with Gimbozu, resulting m an NIL wnh Lifinthebaekground of 'Gimbozu'
(Kato
1997),
TT)isNILwas designatedasNIL-ETbcarry out pyramidingof these
Q
[Ls,thefiveprimaryNILswere intercrossedinal1
10
combinations without reciprocals. in each of theirF2populations(about
150plaants),the sixlongest-grain segregants were selected and theirhomozygosity confirmed
for
thelong
grain
length
inF3progenies(l2
plants per line).From these progenies,true-breeding lineswere established as advanced NILs,illwhich alleles forlonggrain exist inevery combination of twoQTLs
out of five.These 10NILs
for
10QTL
combinations were designatedasNIL-al)toNIL-de. According tothesame
procedure
asabove, NIL-Fwas cressed with each of thefiveprimaryNILs.FiveNILs withUar and an allele forlonggrainat only one difTerentQTL
were established, ThesefiveNILsweredesignated
asNIL-Fa
to NIL-Fe.In
2007,
al1of these2],
NILs and threecultivars, `Gimbozu', `BG1'
and `Fusayoshi', were seeded innursery boxeson16
May and transplantedina paddyfieldef theFacultyof
Biology-OrientedScienceand Technology,KinkiUniversity,Japan,on
13
June,
In2008,
15NILs(excluding
NIL-F
andNIL-Fa
toNll.-Fe)
and'Gimbozu'
were seeded on14
May
and transplanted on10
Junein
thesamefield.
Inboth
years,theplantingdensity
was30
cm inter-rowand15
cm inter-hi11with asingle plantperhill.Eachlineconsisted of sixhills.Fertilizerswere applied as
basaldressingat the rate of 6 g m'2 each
for
nitrogen,phosphorous,and potassium.Other
field
managements weredone
according tostandard cultivation practicesforthisreglon.Afterrnaturing. apanicleon thelongestculm of every plant was harvestedand air-dried, Fivegrainson proximalpartsof
primaryrachis-branches along thetopof arachis were collected
fromindividual
panicles,
Ihesegrains
were measured fortheirgrain
length(between
thebottomand thetopof alemma exceptfor an awn) using a video-micrometersystem
(VM-50N,
Olympus Corp., Japan) equipped with a macro-zoom microscope
(SZHIO,
Olympus Corp,,Japan).An averagelength
forfivegrainswas regarded thegrainlengthof an individual
plant.
Based on themean
grain
lengthof Gimbozu and thelengths of individualNILplants,
themain effect(additive
efTect) of theith
QTL(or
of Lkf),Ai,was estimated as a fixed effect using thefo11owingequation:
Xik=C+Ai+ Eik
(Eq,
1),
where Xikisthegrainlengthof the kthplantinaNIL with a
long-grainallele atthemb
QTL,
Cisacommon background(the
mea" grainlength of Gimbozu),and sikisa residual randomNII-Electronic Library Service
Main
EffectsofIndividualQTLs
and InteractionsbetweentwoQTLs
ControllingGrainLengthof Riceunder Near-IsogenicLinesvariable drawnfromN
(O,
a,Z).Afterestimating
A,
and4 ofthe
twoQTLs
(or
aQ[[L
andzaD
fromEq.
1,
theinteraction(epistatic
interaction)
between
the
ithandJ'th
QTLs
(or
between
theithQTL
andop,
Iij,wasalso
estimated as afixedeffect bythefollowingequation:Xpt = C + Ai+ Aj+ Iij・+ sijk
(Eq.
2),
where Xijkisthegrainlengthof thekthplant
in
aNIL withlong-grainallelesat theithand
jth
QTLs
(er
the ithQTL
aiid Ljij). and Eijkisalsoaresidual random variable as Eik.Results
andDiscussion
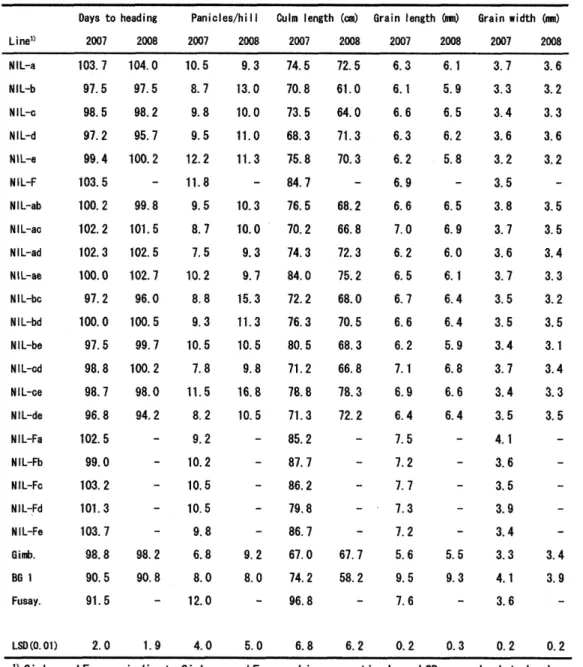
[[bble1 shows seyeral agronomic traitsof
the
presentmaterials
in
2007 and2008,
Large,
significantvariationswerefoundinbothyearsamong NILsand alsobetweenNILsand the
t t
backgroundcultivar, Gimbozu, inngt en!y grainlengthbut also most other traits.TheseTesults suggested thepresenceof some pleiotropiceffects of grain-]ength
QILs
on other traits,Fig,
1
shows themain effbcts of grain-lengthQTLs
in
both
yearsand a major gene,Lofin
2007,
estimated fromthedatainThb]e
1
using Eq.I.
Significantvariations amongQILs
weredetectedinthemain effect jnbothyears
(P<O,Ol).
ThelargestTable1Agronomic traitsottheNILsand thelrparentsused inthepresentstudy
Linei)
Daysto heediog
2co7 2oesPeniclesfhill2co1 2coSCulm
length 2oo7(on)
Grainlengthkem) Grainvldth im}
20(IS 2oo7 2oog 2oo7 2ooS
NIL-aNILthNlL-cN1L-dNIL-eNIL-FNiL-ebNIL-scNIL-adNiL7afiNlL-bcNIL-・bdNILmbeNtL-edNIL-eeNIL-deML-FaNIL-thNIL-FeNIL-FdNIL-・FeGimb.Bfl 1Ftisay. I03.797.598,597.2g9,4I03.51eo, 2le2.2102.3100, O 97.2100.0 97.5 98,8 98.7 96,SI02.5 99.0le3.2tOl,3le3. 7 gs,g 90.5 9L5 I04,O97.59S,295.7loo. 2
--99.SIOL5102.5102,7 96.G1oo, 5 g9.71oo2 98.e 94.2 98.
2
ge.s le.58.7 9.S 9,512.2ll.S 9.5 a.7 7.510, 2 8.8 9,3le. 5 7,811.5 S.29.2le.
210.5le.5 9.8 6.8 B.O12.0 9.3l3.0IO,Oll.O".3 le.3fe.o 9,3 9.715.311.310.5 9.816.ele.5 9.2 s.o 74.57e.873.568,375,884.77G.E70,274,3a4.o12.276.3SO.571.2IB.g71,385,287,786.279.886.767.014.2g6.8?2.561.064.'O71.370,3 6S.266.872.315.268.070.56g.366. S7S.312.2 67,758.2 6.36,1G.66.36.26.96.67.06.26.56.76.66,27.16.96.4Z57,2].7?.37.2S,69,57.66.15.96.56.25.8 6.56.96.06.16.46.45,96.Sg.66, 4-5.59.3
3,73,33.4・3,63.23.53.S3.73,63.73,53.53.43.73.43.54.I3.63.53.93,4 3.34.1 3.6 3.63.23.33.63.2 3.53.S3.43.33.23.53.13.43.33.5-3,43.9
LsD(o.el}2,O1.g4.e5.06.86.2e.2O.3O.2e,2t)
G}mb.
andFusay.
indlcate
Gimbezu
andFusayoshi,
respeetively.Flsher' s
F-pretected
Lso
at theO.
Ol
probability Ievel.`- '
meens that thematerlals were not available inthls year.
LSD
was ca1cu1ated usingThe Society of Crop Science and Breeding in Kinki, Japan
TheSociety ofCrop Science and Breeding iri Kinki, Japan
J.CropRes.
55(2010)
ggEg
{
tsgs
Fig.I Aggggs
i:f,l
l・ aal'
2cxn'.
2eosMaineftectsofindividual
QTLs
and a major gene,Lkf;on ricegraiRlengthovertwoyears.
Main effects with a common letterwere not significantly diherentat theO.Olprobabilityleve],
using Fisher'sF-prctectedLSD,
main effectwas obtained with themajor geneLdy1Arnongthe
five
QILs,
eGLc showed thelargestmain effect compared tothe effect of Llj though therewas asignificantdifferencebetweeneGLc and LijZ The orders inthemagnitudes of themain effects among the
QTLs
were very similar forthetwo differentyears,
A highlysignificant correlation coerncient was obtained between thetwoyearsforthemain effects(r=O.95,
P<O.05),although theinteractionbetween the yearsand
QTLs
by ANOYA was significant(data
not shown). This findingsuggested thatthe!t
t
QTLs
main effects were consistently expressed at theirown magnitudes forthedifferenttwo years.
Song
et al.(2007)
identified
one ofthe
Q[ILs
controlling grain size of a1arge-grain
japonica
cultivar,`WY3', anddesignated
it as GVV2,ThisGIV2
encoded anopen readmgframe
inyolying
the
domainof RING-typeE3ubiquitin ligase.Interestingly,thesame authors showed thatanother 1arge-graincultivar, `Oochikara', also
l
+
hasthesame Gwa. Oochikara was derivedfrom a
・cross
ofSachiminorVInabawaselfBG
1,
and BG1
is
thesource of al1fiveallelesforlonggrainat thefivegrain-length
QILs
as above.Consequently,GW2 would correspond toone of thefive
QTLs
forgrainlengthexarnined inthisstudy,
Zhang et al,
(2009)
analyzed thevariation inthe number of spikelets inaricepanicle.identifiedfourQTLs
forthistrait,anddissected
these
QTLs
intosingleMendeliall factorsfor
each of fburNILs,just
1ikethepresentstudyfor
ricegrainlength. Theyalso
demonstrated1argedifferencesinthe rnain effects amongthescfour
QTLs:
QTLs
with smaller effects could not be expressed suMciently dependingen thegrowingenvironments,Fig.
2
shows theinteractionsbetweentwoQTLs
forgrainlengthinbothyears,estirnatedwith theEq.2.[Ihevalues of the
interactienswere mostly negative and showed a largesignificant variation among
QTL
combinations(P<O.Ol).
In2007,the values of allinteractionswere significantly differentfromzeroCP<O.Ol),
whereas in2008thevalues of six outof10
interactionss
s
g
g
Fig.2-ggsi・gg
・gg
g・
s
gFig.3
al "a4'gx2・a34L4as",fi.-(t7e,s-agl-..It'I'"
lts'Ll''l
2ecrreg.
k ts L.ve 2ooe'y
eetele
-I・ Oexb ;,ll ttangil,:・・:
i
'li:uelI-b,taI1l・ vaetu
l,1Io,op
/-l.f.rmdw...[1, ; de・
t
tslnteractionsbetween two QTLs on rice grain
lengthintwo years.
a xb, for example, [ndicatesthe interaction betweenqGLa and qGLb.
Interactionswith a common letterwere not significantiy dlfferentatthe O,OlprobabilityIevel, fromFisher'sFprotectedLSD, and those with ' and "
were significantly difterentfromzero at the
O.05and O.Olprobabilitylevels,respectiveiy,
qeL-#Lkf
,suT
Irmim'
m.ts} lga l-mm
.L lnteractions qGLa × LkL Nothelnteractions ditferentprobability
gGLbwhptxca'i"'nqtgcakca
..A.
pt"Nttaf'A.
.・
g
・t
e
・
between grain
maior gene,Lkf,on ricegrain
forexamp[e,
'
betweenqGLa and Lkf,
signMcant differences interactions at the O.05
with "and *t from zero at the
levels,respectively. t'
.t.--t lengthQTLs and a length.indicates theinteraction
were detected among
probabilityIevel. were significantly O.05 and O.Ol
were not significantly different
(PIO.05).
The order of magnitudes oftheinteractionsamongQIIL
combinations changedinthetwe different
years,
altheugh highmagn'itude interactionswere consistentlyfoundin
both
yearsinthesame combination(e.g,,
qGLa xqGLd),lrhecorrelation coefficient betweenyearsfor theinteractienswas not significarit(r=O.63,
P)O,05)
and lowerthanthecoefficient forthesame kindformain effects. This result meant thatthe expression of thetwo-QTL interactionswas relatively unstable indifferentyears,which showed a clear centrast with thestable expression of
QTL
main effects, Thisinstahilityintheexpression inthe two-QTL interactionsmight
centribute to theinstabilityof the character expression of rice
grainlength,Theinteractionsbetweena
QTL
and Llj"areshownNII-Electronic Library Service MainEffectsofIndividual
QTLs
and interactionsbetweenTkvoQTLs
ControllingGrainLengthofRiceunder Near-IsogenicLinessmalI compared with two-QTL interactionsinFig.
2,
and net significantlydifferent
among them(Pk
O,05).
Thevalues of twointeractionsoutof fivewere fiotsignificantly differentfromzero
(P)
O.05).
Linet at,
(2000)
demonstratedepistaticinteractionsamongthreemajor
QTLs
controllingheadingdateofrice,Hdl,Ild2,andHd3,usingaseriesofnear-isogenic Iines,inwhich allofthe
two-QTL
combinations of thesethreeQTLs
were irrvolved,They revealedthe
non-additive manner of theseQTL
actions onheading
datefromthese
advancedrnaterials.Ina statistical analysis forquantitativevariation beforethe
popularizationof molecular markers, many geneticmodels for
geneactions
generally
regarded epistatic actions tobenegligible, mainly because of the complexity ef the calculations, and concentrated on the analysis of additive and dominance gene effects(Kearsey
and Pooni1996).
Howeveg thepresentresultsforrice grainlengthartd alsotheresults forriceheadingdate
(Lin
etal.2000)
obviouslydemonstrated
thatinteractions
betweentwoQTLs,
and also aQTL
and a majorgene,
concerned with thesame traitshould not
be
negligiblefactors
ingenetic
models fbrquantitative
traits.Ineweryyear
of thepresentstudy,themeanabsolutevalues across allinteractionswere
.about
halfof thoseofthe
main effects, Moreover,theQTL-by-QTL
interactionsmighthavesome conuibution totheinstabilityofcharacter expression,
This impliesthe interactionsamong
QTLs
should play animportantand fundamentalrole intheregulation of quantitative traits.Interactionsinhigherorders among more thantwo
QTLs,
as well as
QIL-by-QTL-by-environment
interactions,should beevaluated using more advanced NrLs with long-grain
alIeles
in comblnations ofmore thantweQTLs,
by
furtherpyramidingofQTLs.
Acknowledgements
The
author expresses special thankstoM.inoue
and S.Senami,Schoolof Bioresources,HiroshimaPrefecturalUniversity,and T.
Ybshida,A, ekamoto, T.Kakiuchi,and A.Horibata,Facultyof
Biology-OrientedScienceand [Ibchnology,KinkiUniversity,for
theirvaluable technicalassistance.
References
Fan,
C,,
YL
Xing,H.
Mao.
T.Lu,
B.HanC.
Xu, X LiandQ,
Zhang
(2006)
GS3.
amajorQTL
forgrainlengthand weightandminor
QTL
forgrainwidth and thicknessinrice,encodes aputativetransmembraneprotein.
Theor.
Appl.
Genet.
112
:1164
-
1171,Kato,T,
(1997)
Inheritanceofgrainsizein
rice.
6.
Development of near-isogenic 1inesformajor genescausing longgrain.BreedSci,
47
(Suppl,
2)
:232.
(In
Japanese)Kato,T.
(1998)
Isolationof individualquantitativetraitIecicausing longgraininrice.JARQ 32:233
-
238.Kato,
T.
(2005)
Isolation
of quantitatiyetraitloci
controllingrice
grainlengthas asingle Mendelianfactorinnear-isogenic lines,
Proc.
10th
Int.Cong.SABRAO :G-
2.
Kearsey,M, J.and H, S.Pooni
(1996)
Geneticalanalysis ofquantitative
'traits.
Chaprnan
& Hall,London, UK,p,378,
Kubo,T,,YL
Aida,
K,Nakamura,H.Tsunematsu,K.Doi and A,Ybshimura
(2oo2)
Reciprocalchromosome segment substi-tutionseries derivedfromjaponica
ancl indicacross of rice(Oryza
sativa L.),Breed.Sci,52:319-
325.
Lander,E.S.and D,Botstein
(1989)
Mapping Mendelianfactors
underlyingquantitative
traitsusing RFLP linkagemaps.Genetics
121
:185
-
199,
Li" H. X., T.Yarnfimoto,T. Sasakiand M, Ylano
(2000)
Characterizationand detectionof epistatic interactionsof
3
QTLs,
Hdi, Hd2, andHd3,
controllingheading
date
in
rice
using nearlyisogeniclines.Tlieor,Appl.Genet.
101
:1021
-1028.Song,
X,J,,WLHuang,M. Shi,M.Z.Zhuand H,X,Lin
(2007)
AQTL
forricegrainwidth and weight encodes a previously unknown RING-type E3 ubiquitinligase.
Nature
aenet.
39
:623
-
630.
Takeda,
K.
andK,
Saito
(1980)
A
major genecontrolling long grain size derivedfrom a ricevariety "Fusayoshi". Jpn.J.Breed.
30
(Suppl.
1):254-
255,
(I,n
Japanese)Thnksley,S.D,
(1993)
Mapping polygenes.Ann, Rev.Genet,27:205
-
233,
Tlanksley,S.D.and T.M. Fulton
(2007)
Dissectingquantitative traitvariation-examples from the tomato, Euphytica154 :365
-
370.
Yano,M, and r Sasalci
(1997)
Geneticand molecular dissection ofquantitativetraitsinrice. PlantMel.Biol.35
:145
-
153,
Zhang,YL,L. Luo,T:Liu,C,Xu andYLXing
(2009)
FourrjceQTLs
controlling numher of spikelets perpanicleexpressed the characteristics of single Mendelian gene innear isogenicbackgrounds.Theor,Appl.Genet.
118
:1035
-
1044.The Society of Crop Science and Breeding in Kinki, Japan
The Sooiety of Crop Soienoe and Breeding in Kinki
厂
JapanJ