1〒 010–0673 秋田県男鹿市戸賀塩浜字壺ヶ沢 93 男鹿水族館 2〒 905–0206 沖縄県本部町石川 424 沖縄美ら海水族館 3〒 905–0206 沖縄県本部町字石川 888 沖縄美ら島財団 総合研究センター 4〒 903–0213 沖縄県西原町千原 1 琉球大学理学部海洋自然科学科生物系 (2015 年 11 月 18 日受付;2016 年 2 月 18 日改訂;2016 年 2 月 22 日受理) キーワード:タナゴモドキ,絶滅危惧種,ミトコンドリア DNA,遺伝的集団構造,遺伝子流動 Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2016
Shumpei Tomita*, Shohei Matsuzaki, Shin-ichiro Oka, Minoru Toda and Hideyuki Imai. 2016. High levels of genetic diversity and gene flow in the endangered goby Hypseleotris
cyprinoides on Okinawajima Island, Ishigakijima Island and Luzon Island. Japan. J.
Ichthyol., 63(1): 27–32.
Abstract The amphidromous goby Hypseleotris cyprinoides is the most widely distributed
Hypseleotris species in the Indo-Western Pacific Ocean. Recently, the species has declined
around Japan because of environmental destruction, overexploitation, water pollution and the effects of invasive fish species, and has been designated as an “endangered species in Japan”. Because captive breeding for the preservation of endangered species is considered an indispensable course of action in conservation biology, the Japanese Association of Zoos and Aquariums (JAZA) is currently attempting such for H. cyprinoides. Furthermore, there exists an urgent need for comprehensive studies on the genetic population structures and diversity levels of the species for the promotion of effective conservation and management activities. The present study examined the genetic population structure, genetic diversity and gene flow of H. cyprinoides from the Philippines to southern Japan, covering the Kuroshio Current region of the western North Pacific. Eight hundred and thirty-five bp of the mtDNA control region were analyzed in 140 specimens taken from three localities: Okinawajima Island and Ishigakijima Island, Ryukyu Archipelago, southern Japan and Luzon Island, Philippines. A total of 113 haplotypes were detected, four to eight shared haplotypes occurring at all localities, which had high levels of genetic diversity (range: h = 0.9930–0.9984; π = 0.0133–0.0153). AMOVA, pairwise FST, and Nm
revealed high gene flow among the three localities, suggesting overall genetic homogeneity and a single shared gene pool, due to high dispersal ability during the oceanic larval stage and likely influence of the Kuroshio Current. Conservation and management directives should recognize the potential of these areas for sustainable recruitment of the species.
*Corresponding author: Oga Aquarium Co., Ltd, 93 Toga Shiohama, Oga, Akita 010– 0673, Japan (e-mail: [email protected])
タ
ナゴモドキ- Hypseleotris cyprinoides(Valenciennes, 1837)は,タナゴモドキ属のハゼ亜目魚類で あり,インド洋西端のレユニオン島から日本やオー ストラリアを含むインド・西太平洋地域に広く分 布している.タナゴモドキ属は外観がその名前の とおりタナゴに似ており,遊泳生活を送るなどハ ゼ科魚類の中では特異な形態,生態を示すことが 知られている(道津ほか,2000).本種の分布北 限は琉球列島とされており,国内では奄美大島, 沖縄島,久米島,宮古島,石垣島,西表島などか ら報告されている(立原,2005).タナゴモドキ の仔魚は高い塩分耐性を持っており,生まれた仔 魚は海に下って,3ヶ月ほど海での生活を送った 後, 川に遡上する両側回遊を行う(道津ほか,絶滅危惧種タナゴモドキ集団の高い遺伝的多様性と
遺伝的均一性
富田峻平
1・松崎章平
2・岡 慎一郎
3・戸田 実
3・今井秀行
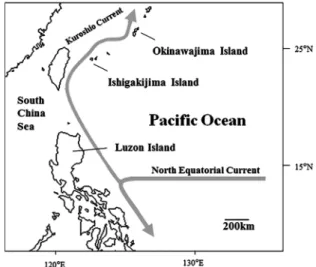
41998).和歌山(山本,1997)や高知(木下ほか, 1990)などでも南方からの偶来種としての報告が あり,海域生活期において広い範囲に分散すると 考えられる.河川に定着後は水生植物が繁茂し, 比較的流れが緩やかな淡水域から汽水域上端を好 んで生息する(立原,2005). 近年,タナゴモドキは湿地の埋め立てや護岸の コンクリート化,堰堤やダムによる河川の分断, 水質汚染,河川への土砂流入,外来種との競合な どさまざまな人為的影響のために日本国内で減少 している(立原,2005).現在,環境省からは絶 滅危惧 IB 類(環境省,2014),鹿児島県では絶滅 危惧 I 類(米沢,2003),沖縄県では絶滅危惧 IB 類(立原,2005),日本動物園水族館協会からは 繁殖対象種(日本動物園水族館協会,1995),日 本水産資源保護協会からは危急種(鈴木,1996) に指定されており,本種の保護・管理対策が急務 となっている.絶滅危惧種には,人工繁殖により 野生個体群の回復を図る場合もある.このような 保全策をとる場合,繁殖単位とすべき遺伝的に独 立した集団の識別や,遺伝的多様性に基づく集団 構造や歴史的な集団動態,地域間の遺伝的交流の 有 無 を 明 ら か に す る 必 要 が あ る( 大 仲 ほ か, 1999;向井・杉本,2006). 本研究では,進化速度が大きく,個体変異を蓄 積しやすいミトコンドリア DNA 調節領域(control region)の塩基配列分析によって,沖縄諸島内お よびフィリピンルソン島に生息するタナゴモドキ の遺伝的多様性や固有性を評価した. 材 料 と 方 法 サンプル採集 DNA 分析に供したサンプルは 2010 年から 2011 年に沖縄島で釣りやタモ網によっ て採集した 42 個体(宜野湾市田芋畑用水路;n = 1,世冨慶川;n = 1,大浦川;n = 6,源河川;n = 17,満名川;n = 4,大井川;n = 1,与那川;n = 12),石垣島の名蔵川で採集した 50 個体およびル ソン島で採集した 48 個体の計 3 地域 140 個体と した(Fig. 1).本研究では大仲ほか(1999)に従い, 個 体 へ の 影 響 を 最 小 限 に と ど め る た め 臀 鰭 を DNA 分析用の試料とした.採集した個体は,原 則として臀鰭を切除後,採集地へ放流した. 飼育実験 DNA 分析用に行った臀鰭切除の影 響を確かめるために 2010 年から 2011 年に沖縄島 で採集した個体のうち,琉球大学の施設内で 7 個 体,沖縄美ら海水族館で 16 個体のタナゴモドキ を 1 年間飼育した.飼育環境は,大学施設内では 45cm 規格水槽でエアーリフト式底面フィルター を使用した.水槽に 7 個体収容し,水温調節はせ ず外気温で管理した(約 20–25˚C).2 日に 1 度冷 凍の赤虫を給餌した.美ら海水族館では 60cm 規 格水槽でエアーリフト式スポンジフィルターを使 用した.1 水槽に約 10 個体収容し,水温調整は せず外気温で管理した(水温約 23–29˚C).飼料 には冷凍赤虫と乾燥オキアミを使用し,1 日 2 度 適量を給餌した. DNA 分析 試料から SDS・フェノール・クロ ロフォルム法とジエチルエーテルとエタノール沈 殿 に よ っ て 粗 全 DNA を 抽 出 し た(Imai et al., 2004).粗全 DNA から PCR 法によってミトコン ドリア DNA 調節領域を含む 835 bp の増幅を行っ た.用いたプライマーは,12SAR-H(5ʼ-ATAGTG GGGTATCTAATCCCAGTT-3ʼ)(Palumbi et al., 1991) と Pro-goby(5ʼ-CCACCBCTRRCTCCCAAAGC-3ʼ) である.Pro-goby プライマーは,タナゴモドキと 近縁のジャノメハゼ Bostrychus sinensis とチチブ モ ド キ Eleotris acanthopoma の 塩 基 配 列(DDBJ/ EMBL/GenBank Accession No.: AP004455, HM117667)を参照にして,tRNAPro領域に設計し た.PCR 反応溶液は,KAPATaq DNA ポリメラー ゼ(Kapa Biosystems)5 unit,鋳型 DNA 2 µl,各プ ライマー25 pmol,5 × KAPATaq Extra Buffer(Mg2+ free)10 µl,25 mM MgCl2 3.5 µl,KAPA dNTP Mix(10 mM)1.5 µl に滅菌蒸留水 30.75 µl を加え,全量を 50 µl とした.反応には,GeneAmp® 9700(Applied Biosystems) を 用 い,94˚C で 2 分 の 熱 変 性 後, 94˚C で 60 秒の熱変性,48–54˚C で 60 秒のアニー
リング,72˚C で 90 秒の伸張反応のサイクルを 30 回繰り返し,最後に 72˚C で 7 分間の伸張反応を 行った.増幅した PCR 産物は,PCR Product Pre– Sequencing Kit(USB Co.) を 用 い て 精 製 後, BigDye Terminator Cycle Sequencing Kit ver. 3.1 (Applied Biosystems)を用いてサイクルシークエ ン シ ン グ 反 応 を 行 い,ABI 3700 Genetic Analyzer (Applied Biosystems)を用いて塩基配列を決定した.
決 定 し た 塩 基 配 列 は 日 本 DNA デ ー タ バ ン ク (DDBJ) に 登 録 し た(Accession No.: AB989437–
AB989549).
データ解析 得られた塩基配列は,Clustal X (Thompson et al., 1997)でアライメント後,ハプ
ロタイプを決定した.ハプロタイプ間の類縁関係 を示すハプロタイプネットワークを,NETWORK ver. 4.6.1.1(Bandelt et al., 1999)を用いて作成した. 遺伝的多様性の指標であるハプロタイプ多様度 h (Nei,1987)と塩基多様度 π(Tajima,1983)は,
ARLEQUIN 3.11(Excoffier et al., 2005)を用いて算 出した.同様に,ARLEQUIN を用いて分子分散 分析(AMOVA),集団間の遺伝的分化を評価する ペアワイズ FSTの算出を行った.ペアワイズ FST における多重検定の補正には sequential Bonferroni 法(Rice, 1989)を用いた.世代ごとの移住個体 数 を 示 す Nm は Nm = 1/2[(1/ FST )-1] 式 に FST値 を 代入して算出した(Hudson et al., 1992).また,集
Table 1. Sampling locations, date and number of Hypseleotris cyprinoides samples used for mtDNA analysis
Sampling sites Date n Number of haplotypes h ± S.D. π ± S.D. Tajima’s D Fu’s FS
Okinawajima Island May, 2010– August, 2011 42 38 (30) 0.9930 ± 0.0073 0.0136 ± 0.0070 -0.56138 -22.91213* Ishigakijima Island September, 2011 50 48 (40) 0.9984 ± 0.0044 0.0153 ± 0.0078 -0.39188 -24.39838* Luzon Island December, 2012 48 43 (32) 0.9938 ± 0.0063 0.0133 ± 0.0068 -0.51481 -24.53167
**P < 0.05. Parentheses indicate number of unique haplotypes.
Fig. 2. Haplotype network tree presenting relationships among 113 haplotypes in Hypseleotris cyprinoides. Circle
団の拡大やボトルネックといった履歴を推定する ために,ARLEQUIN によって Mismatch Distribution 分析(Rogers and Harpending,1992;Rogers,1995) を行うとともに,DNA 変異の中立性を評価する 指標である Tajimaʼs D(Tajima,1989),Fuʼs FS(Fu, 1997)を算出した. 結 果 尻鰭をすべて切除した影響を調べるための飼育 実験を行った結果,遊泳や摂餌に影響はなく 2 週 間から 3 週間で尻鰭がほぼ再生した.また 23 個 体を飼育して 1 年間で死亡が確認されなかった. ミトコンドリア DNA 調節領域の解析において, 835 bp の塩基配列中の変異サイト数は 113 であり, 合計 113 種類のハプロタイプが検出された.各地 域におけるハプロタイプ出現数はそれぞれ,沖縄 島で 37,石垣島で 48, ルソン島で 43 であった. このうち 4 つのハプロタイプは 3 地域間で共有さ れており,4–8 個のハプロタイプは 2 地域間で共 有されていた(Fig. 2).各地域のハプロタイプ多 様 度 と 塩 基 多 様 度 は h = 0.9930–0.9984,π = 0.0133–0.0153 と高い値を示した(Table 1).ハプ ロタイプネットワークにおいて,地域ごとにクレー ドを形成する傾向は認められなかった(Fig. 2). AMOVA の結果においても,明確な遺伝的構造が 認められず(P > 0.05),地域集団間に起因する変 異がほとんどなく地域集団内に起因する変異が 100% であった.ペアワイズ FSTの値は沖縄島と 石垣島間では -0.0006,沖縄島とルソン島間では -0.0077,石垣島とルソン島間では -0.0027 を示し, 有意な分化は認められなかった.また,Nm の値 はすべての地域間において∞(無限)を示した. Mismatch Distribution 分析の結果,集団の安定を仮 定したモデルと一致した(Fig. 3). また,DNA 変異の中立性を評価する Tajimaʼs D と Fuʼs FSの値 はすべての地域で負の値を示した(Table 1). 考 察 絶滅危惧魚類からの非致死的な試料採集方法を 提唱した大仲ほか(1999)の鰭切法は,臀鰭を切 除したタナゴモドキにおいても,飼育実験の結果 から生存に影響を与えておらず,有効な手法であ ると判断された. ミトコンドリア DNA 調節領域の部分塩基配列分析 により,タナゴモドキ 3 地域における遺伝的多様性は, ボウズハゼ Sicyopterus japonicus(h = 1.000; Watanabe et al., 2006)や,Rhinogobius maculafasciatus(h = 0.9819; Cheng et al., 2005)などの両側回遊性ハゼ類と同等 に高い水準を有していた.本研究のタナゴモドキ 集団に見られた高いハプロタイプ多様度と塩基多 様度は,有効集団サイズが大きいことに起因する と考えられる.Mismatch Distribution 分析の結果か ら,これらの地域では,過去に集団の急激な拡大後に 安定した可能性が示唆された(Rogers and Harpending, 1992).また,Tajimaʼs D と Fuʼs FSの値がともに 3 地域で負の値をとり,なおかつ Fuʼs FSの値には, 中立進化からの有意な逸脱が認められたことから も,集団の急激な拡大が支持された.Fu(1997)は, Tajimaʼs D 検定より Fuʼs FS検定の方が高感度であ ることを報告しており,Fuʼs FSの値のみに有意性
Fig. 3. Observed pairwise differences (bars) and expected
mismatch distributions under the expansion model (solid line) of control region haplotypes in Hypseleotris
cyprinoides from Okinawajima Island, Ishigakijima Island
and Luzon Island locations. Model parameters for sudden expansion and goodness-of-fit test indicated: SSD sum of squared deviations; θ0 and θ1, pre-expansion and
post-expansion population sizes, respectively ; τ, time (number of generations elapsed) since sudden expansion episode.
が認められたのは検出力の違いに起因すると考え られた. ペアワイズ FSTの値は 3 地域間の遺伝的分化が 非常に小さい,あるいはほとんどないことを示唆 した.つまり沖縄島,石垣島,ルソン島の間には 遺伝子流動が十分にあると考えられた.沿岸性魚 類では,主に仔稚魚の浮遊期間に広域分散が起こ ることはよく知られている(Sale,1991).特に, タナゴモドキのような卵サイズが比較的小さなハ ゼ亜目魚類では,3ヶ月を超える長い浮遊期間が あると報告されている(道津 , 1979).タナゴモ ドキと同様に,琉球列島に生息するハゼ類(例え ば,スジクモハゼ Bathygobius cocosensis やチチブ モドキ Eleotris acanthopoma)においても地域間で 遺伝的分化が生じていないことが報告されている (向井 , 2010).琉球列島の西方沖には流れの強い 黒潮とそれに伴う反流が流れているため,長い浮 遊期間を有するタナゴモドキの仔稚魚は南方から 沖縄諸島への加入が十分考えられる.本種の分布 域の一つであるフィリピン諸島と沖縄諸島は直線 距離で約 1,700 km 離れているが,黒潮の流速は 5.5–9.25 km/h であり(Yoshimura et al., 1999),3ヶ 月という浮遊生活期間はタナゴモドキの仔稚魚が 黒潮に乗って沖縄諸島まで分散するのに十分であ る.したがって,沖縄諸島のタナゴモドキはフィ リピン諸島の沿岸地域で孵化した仔魚が海に下っ た後,黒潮に取り込まれ,沖縄島,石垣島沿岸に 到達後,定着したものも多く含まれていると考え られる. 本研究では,沖縄諸島内の沖縄島と石垣島およ びルソン島のタナゴモドキの地域間には大きな遺 伝子流動があるという結果を得た.国内の水族館 等で実施されているタナゴモドキの保護・繁殖の 現場において,少なくとも黒潮流域内の地域集団 を飼育する場合は,異なる集団の混在に特段の配 慮をしなくても遺伝的撹乱を起こす可能性が低い と考えられる.ただし,数少ない親魚から生産さ れた数万尾単位以上の人工種苗を野外に放流する 場合は,自然集団の遺伝的組成を変えてしまう可 能性があるため,人工孵化にはより多くの親魚を 用いる必要があると考えられる.また,日本産タ ナゴモドキ集団の保全策として,本種の仔稚魚の 加入を妨げる河口閉塞の解消,外来魚の侵入の防 止,本種が好む水生植物の繁茂する河岸や水田水 路(井口ほか,2002)の保全など良好な河川環境 を維持することが最も重要であると考えられる. 謝 辞 本研究を進めるにあたり,琉球大学理学部の立 原一憲博士,小枝圭太博士(現,鹿児島大学総合 博物館)および沖縄科学技術大学院大学の前田 健博士にはルソン島における標本の採集協力およ び有益な助言を頂いた.標本の採集に協力して頂 いた宮本 圭氏をはじめとする沖縄美ら島財団と 沖縄美ら海水族館の職員の方々に心より厚く御礼 申し上げる.琉球大学理学部今井研究室の岩本健 輔博士(現,WDB バイオ環境研究所),Muhamad Fadry Abdullah 博士(現,共栄製作所),常盤健治 郎氏には心から感謝の意を表す.本研究の一部は, 琉球大学理学部の「国際サンゴ礁研究教育ハブ形 成プロジェクト」の援助を受けた. 引 用 文 献
Bandelt, H. J., P. Forster and A. Rohl. 1999. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol., 16: 37–48.
Cheng, H.-L., S. Huang and S.-C. Lee. 2005. Phylogeography of the endemic goby, Rhinogobius
maculafasciatus (Pisces: Gobiidae), in Taiwan. Zool.
Stud., 44: 329–336. 道 津 喜 衛.1979. ハ ゼ 亜 目 魚 類 の 卵 と 仔, 稚 魚. 海洋科学,11: 111–116. 道津善衛・鈴木寿之・柳 昌之.1998.タナゴモ ドキ(ハゼ科魚類) の採卵, 卵内発生, 仔魚. 長崎生物学会誌 , 49: 15–21. 道津善衛・柳 昌之・乾 輝男.2000.オースト ラリア産ハゼ科タナゴモドキ属魚類 Hypseleotris compressus の採卵と仔魚の飼育.長崎大学水産学 部研究報告,81: 43–48.
Excoffier, L., G. Laval and S. Schneider. 2005. Arlequin ver. 3.0: an integrated software package for population genetics data analysis. Evol. Bioinform. Online, 1: 47– 50.
Fu, Y. X. 1997. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics, 147: 915–925.
Hudson, R. R, M. Slatkin and W. P. Maddison. 1992. Estimate of levels of gene flow from DNA sequence data. Genetics, 132: 583–589.
井口恵一郎・淀 太我・片野 修.2002.西表島 の水田用水系に出現する魚類の生活環境.魚類 学雑誌,50: 115–121.
Imai, H., J.-H. Cheng, K. Hamasaki and K. Numachi. 2004. Identification of four mud crab species (genus
Scylla) using ITS-1 and 16S rDNA markers. Aquat.
Living Resour., 17: 31–34.
物-レッドデータブック-汽水・淡水魚類.自 然環境研究センター,東京.226 pp.
木下 泉・藤田真二・高橋勇夫・東 健作.1990. 陸圏と水圏の間 , とくに河口域の仔稚魚動態.魚 類学雑誌,36: 505–506.
Leis, J. M. 1991. The Pelagic Stage of Reef Fishes: The Larval Biology of Coral Reef Fishes. pp. 183–230 in P. F. Sale, ed. The ecology of fishes on coral reefs. Academic Press, San Diego.
向井貴彦.2010.比較系統地理学から見た琉球列 島の淡水魚類相の成立.渡辺勝敏・高橋 洋(編 著),pp. 169–185.淡水魚類地理の自然史―多様 性と分化をめぐって.北海道大学出版会,札幌. 向井貴彦・杉本真奈美.2006.日本産トビハゼの ミトコンドリア DNA 多型に基づく遺伝的集団構 造の解析.魚類学雑誌,53: 151–158.
Nei, M. 1987. Molecular evolutionary genetics. Columbia University Press, New York. 180 pp.
日本動物園水族館協会.1995.第 8 回種保存会議 経過報告並びに種保存委員会経過報告.日本動 物園水族館協会,東京.216 pp. 大 仲 知 樹・ 佐 々 木 裕 之・ 長 井 健 生・ 沼 知 健 一. 1999. 絶 滅 危 惧 種 ウ シ モ ツ ゴ 集 団 に 見 ら れ た mtDNA D ループ領域の著しい単型性.日本水産 学会誌,65: 1005–1009.
Palumbi S., A. Martin, S. Romano, W. O. MacMillan, L. Stice and G. Grabowski. 1991. The simple Foolʼs Guide to PCR, version 2.0, Department of Zoology and Kewalo Marine Laboratory, University of Hawaii, Honolulu. 46 pp.
Rice, W. R. 1989. Analyzing tables of statistical tests. Evolution, 43: 223–225.
Rogers, A. R. 1995. Genetic evidence for a Pleistocene population expantion. Evolution, 49: 608–615.
Rogers, A. R. and H. Harpending. 1992. Population growth makes waves in the distribution of pairwise differences.
Mol. Biol. Evol., 9: 552–569.
鈴木寿之.1996.タナゴモドキ.水産庁(編),日 本の希少な野生水生生物に関する基礎資料(III), pp. 215–221.日本水産資源保護協会,東京 . 立原一憲.2005.タナゴモドキ.沖縄県文化環境 部自然保護課(編),pp. 166–167.改訂・沖縄県 の絶滅のおそれのある野生生物動物編レッドデー タおきなわ.沖縄県文化環境部自然保護課,那 覇.
Tajima, F. 1983. Evolutionary relationship of DNA sequences in finite populations. Genetics, 105: 437–460. Tajima, F. 1989. Statistical method for testing the neutral
mutation hypothesis by DNA polymorphism. Genetics, 123: 585–595.
Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin and D. C. Higgins. 1997. The Clustal_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res., 25: 4876–4882.
Watanabe, S., M. Iida, Y. Kimura, E. Feunteun and K. Tsukamoto. 2006. Genetic diversity of Sicyopterus
japonicus as revealed by mitochondrial DNA
sequencing. Coast. Mar. Sci., 30: 473–479.
山本泰司・大田 満・荒賀忠一.1997.富田川(和 歌山)で採捕したタナゴモドキ . 南紀生物,39: 132–134. 米沢俊彦.2003.タナゴモドキ.鹿児島県環境生 活部環境保護課(編),pp. 126.鹿児島県の絶滅 のおそれのある野生動植物 動物編鹿児島県レッ ドデータブック.財団法人鹿児島県環境技術協 会,鹿児島.
Yoshimura, T., H. Yamakawa and E. Kozasa. 1999. Distribution of final stage phyllosoma larvae and free-swimming pueruli of Panulirus japonicus around the Kuroshio current off southern Kyushu, Japan. Mar. Biol., 133: 293–306.