神経堤細胞と先天性心疾患
山岸 敬幸慶應義塾大学医学部小児科
Cardiac Neural Crest and Congenital Heart Defects
Hiroyuki Yamagishi
Department of Pediatrics, Division of Pediatric Cardiology, Keio University School of Medicine, Tokyo, Japan
Congenital heart defects (CHD) result from abnormal morphogenesis of the embryonic cardiovascular system and usually involve defects in specific structural components of the developing heart and vessels. Decades of research in molecular embryology have suggested that multiple distinct cell lineages give rise to the cardiovascular system. Neural crest cells (NCC) are pluripotent cells that delaminate from the dorsal neural tube and migrate widely throughout the body as mesenchymal cells. A subregion of the cranial NCC originating between the otocyst and somite 3 is called the “cardiac NCC” that migrate into the third, fourth, and sixth pharyngeal arches and the cardiac outflow tract. They contribute to the remodeling of the six pairs of bilaterally symmetric pharyngeal arch arteries that eventually results in formation of the aortic arch system. In addition, they also form the septum of the outflow tract that divides the embryonic single truncus arteriosus into the aortic and the pulmonary trunk. Recently, a new population of myocardial precursor cells in pharyngeal mesoderm anterior to the early heart tube was discovered and named the “second heart field (SHF)”. Reciprocal signaling between cardiac NCC and cells originated from the SHF are essential for the development of the outflow tract and the aortic arch system. Discovery of the SHF has provided important implications for the interpretation of the cardiac outflow tract development and new insights into the morphogenesis of CHD.
要 旨 心臓神経堤細胞とは,神経管の耳胞から体節 3 までのセグメントに起源し,多分化能を有する細胞である. 間葉細胞として体の前後軸に対して分節的に移動し,第 III,IV,VI 咽頭弓に分布して,各咽頭弓動脈(胸部大 動脈)の平滑筋に分化する.流出路の発生には,臓側中胚葉の二次心臓領域細胞と心臓神経堤細胞の相互作用 が必要で,二次心臓領域細胞は流出路心筋・平滑筋層を形成し,流出路まで移動してきた心臓神経堤細胞は中 隔を形成する.これら心臓前駆細胞の異常により,心臓流出路異常のスペクトラムが生じる. Key words:
heart development, second heart field, outflow tract, pharyngeal arch, TBX1 はじめに 本セッションでは,最新の知見を紹介することに先 立って,小児循環器専門医に必要な知識を念頭に講演 した.小児循環器専門医修錬目標(平成 23 年版)では, 《総論》の「Ⅰ.心血管系の発生,発達,解剖,機能 B. 発生,胎生学」として,以下の項目があげられている. 本講演では,下線の項目に関連付けてまとめるように 心がけた. ・体静脈の胎生期の遺残や主な奇形(B) ・ 正常の肺静脈の発達過程と主な異常(例,肺静脈 閉塞)(B) ・ 原始心筒の主要な要素とループ形成のメカニズム(A) ・ 正常の主要構造の発達過程(例,半月弁,房室弁, 心中隔の分離)(A) ・ 心球および動脈幹の正常発達およびその異常によ る心奇形(A) ・ 大動脈弓の発達(A) ・ 心房・心室接合の発生学的基礎(B) ・ 右室および左室流出路の先天奇形を起こしうる発 達異常(B) ・ 心室−大血管不一致の発生学的基礎(B) ・ 冠動脈の発生と主な異常(B) ・ 刺激伝導系の発生(B) ・ 心奇形における神経堤組織の役割(B) * (A)必須項目:小児循環器の基本,(B)努力項目: Ⅱ 基礎
できるだけ習得すべき項目 また,本稿の執筆にあたっては,項目ごとにキーワー ド,キーセンテンスを箇条書きであげ,それに解説, 図を加えることによって,基本事項を修得し,理解を 深めやすいような構成とした. 1.心臓神経堤細胞 (1)神経管背側に起源(耳胞∼体節 3):外胚葉由来 (2) 多分化能を有する間葉細胞で,遊走・分化して各 組織を形成 ・胸部大動脈(第 III,IV,VI 弓)平滑筋 ・流出路中隔 ・ 胸腺,副甲状腺,感覚・交感神経節,色素細胞, 頭部骨格
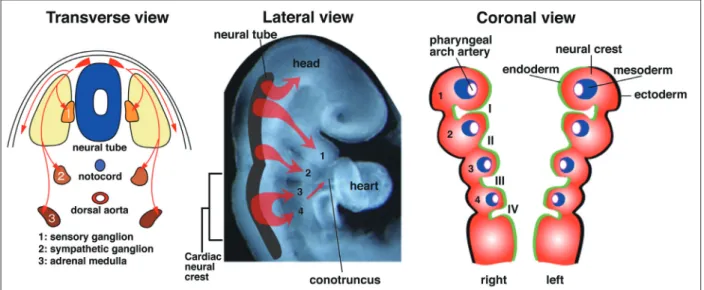
1)心臓神経堤細胞(cardiac neural crest cell)とは 神経堤細胞(neural crest cell)は,胎生期の神経原基 である神経管(neural tube:外胚葉)に由来し多分化能 を有する細胞集団である.神経管の背側に起源し,間 葉細胞として体の前後軸に対して分節的に遊走し, 様々な細胞(感覚神経細胞,グリア細胞,交感・副交 感神経細胞,副腎髄質,メラニン色素細胞など)に分 化する1)(Fig. 1).頭部から体節 3 までの神経管に由来
する頭部神経堤細胞(cranial neural crest cell)は,左右
6対の咽頭弓(pharyngeal arch)を形成し(Fig. 1),頭蓋 骨,軟骨,平滑筋,結合組織なども形成する.頭部神 経堤細胞の中で,耳胞(otocyst:耳の原基)から体節3(ま たはロンボメア:rhombomere 6~8)までの神経管に起 源する細胞群は,心臓神経堤細胞と呼ばれる1).心臓 神経堤細胞は,左右 6 対の咽頭弓のうち,第 III,IV, VI咽頭弓に分布し,間葉系細胞として各咽頭弓動脈, および胸腺,副甲状腺の形成に関与する.さらに,心 臓(円錐動脈幹:conotruncus)まで達する.心臓神経堤 細胞は胸部交感神経節,心臓神経節にも分化する. 2)心臓神経堤細胞と先天性心疾患 ニワトリ胚で神経堤細胞を摘除して発生させる実験 により,神経堤細胞が正常心臓大血管の発生に必要で あることが明らかにされた2).心臓神経堤細胞摘除ニ
ワトリ胚(神経堤摘除胚:neural crest ablation embryo) では,円錐動脈幹・大動脈弓異常,および胸腺,副甲 状腺の低形成がおこる.円錐動脈幹異常は神経堤摘除 胚の約 90%に発生し,総動脈幹遺残(persistent truncus arteriosus:PTA)を特徴とする.大動脈弓異常は,神 経堤摘除胚の全例に認められ,大血管リモデリング (remodeling)の異常を特徴とする.
Fig. 1 Development of neural crest cells. Neural crest cells (shown in red) are multipotential cells that delaminate from the dorsal neural tube and migrate widely throughout the body as ectoderme-derived mesenchymal cells. Neural crest cells ultimately give rise to an enormous array of different cell types, tissues, and organs, including the peripheral and autonomic nervous systems, adrenal medulla, melanocytes, pharyngeal arches, and facial skeleton. A subregion of the cranial neural crest cells originating between the otocyst and somite 3 is the cardiac neural crest cells that migrate into the third, fourth, and sixth pharyngeal arches and the cardiac outflow tract (conotruncus). The transverse view of the embryonic trunk, lateral view of rostral region of the embryo, and a coronal view of the pharyngeal aches around embryonic days 9-10 in mice (equivalent to week 4-5 in humans) are shown.
2.大血管の発生 (1) 左右対称の 6 対の咽頭弓動脈のリモデリングによ り形成 ・左右第 I,II 弓:退縮して消失 ・ 左右第 III 弓:左右総頸動脈を形成 ・ 左第 IV 弓:(左)大動脈弓を形成 ・ 右第 IV 弓:右鎖骨下動脈近位部を形成 ・ 左右第 V 弓:哺乳類ではほとんど形成されない ・ 左右第 VI 弓:左右肺動脈起始部および(左のみ) 動脈管を形成 (2) 心臓神経堤細胞:第 III,IV,VI 弓・胸部大動脈 平滑筋に分化 (3) 大動脈弓異常:リモデリング(退縮・存続)の異常 により発生 1)大動脈弓の形態形成 3) 胎生 4 ∼ 5 週に,胎児の頸部器官である咽頭弓が, 頭側から尾側へ順に左右対称に 6 対形成され,各々神 経と動脈を含む(哺乳類では第 V および VI 咽頭弓は 目立たない)(Fig. 1).これらの動脈は咽頭弓動脈
(pharyngeal arch artery)と呼ばれ,動脈幹(truncus arteriosus) の頭側に連なる大動脈囊(aortic sac)から起始し,左右 の背側大動脈(dorsal aorta)につながる(Fig. 2).第 I,
II咽頭弓動脈は出現後急速に消失し,第 V 咽頭弓動 脈はまったく形成されないか,すぐに退縮する.第 III,IV,VI 咽頭弓動脈は存続し,リモデリングによっ て形態を変え,大動脈弓とその分枝,肺動脈,動脈管 を形成する(Fig. 2). ① 第 I 咽頭弓動脈:胎生 4 週までに大部分は消失する. 細い血管となって顎動脈(maxillary artery)を形成する. ② 第 II 咽頭弓動脈:発生後まもなく消失する.細い 血管となって舌骨動脈(hyoid artery),アブミ骨動脈 (stapedial artery)を形成する. ・ 第 III,IV,VI 咽頭弓動脈は,第 I,II 咽頭弓動 脈の消失と前後して発生し,背側大動脈に連結す る.大動脈弓は左側優位に発達し,心ループ形成 とともに左右非対称の形態となる. ③ 第 III 咽 頭 弓 動 脈: 近 位 部 は 総 頸 動 脈(common
Fig. 2 Development and anomalies of the aortic arch system. (A) Remodeling of the bilaterally symmetric third, fourth, and sixth pharyngeal arch arteries (PAA) into the aortic arch system is shown in a color coordinated fashion. Arrows indicate the migration of the cardiac neural crest cells.
(B) Interrupted aortic arch type B results from abnormal regression of the left fourth PAA. (C) Double aortic arch results from abnormal development of the right fourth PAA and persistence of the right dorsal aorta. (D) Aberrant right subclavian artery results from abnormal regression of the right fourth PAA.
Ao: aorta, BCA: brachiocephalic artery, DA: ductus arteriosus, LCC: left common carotid artery, LSC: left subclavian artery, PA: pulmonary arteries, RCC: right common carotid artery, RSC: right subclavian artery.
carotid artery),遠位部は背側大動脈と結合し内頸動 脈(internal carotid artery)を形成する.
④ 第 IV 咽頭弓動脈:左側は左総頸動脈と左の鎖骨下 動脈(subclavian artery)の間の大動脈弓(aortic arch) の一部を形成し,右側は右鎖骨下動脈の最も近位部 を形成する.
⑤ 第 VI 咽頭弓動脈:肺動脈末梢は肺原基から発生し, 原始肺動脈(primitive pulmonary artery)の主要分枝は 第 VI 咽頭弓動脈の形成以前にすでに存在する.胎 生 29 日頃,第 VI 咽頭弓動脈が発生し肺動脈幹と接 続する.右側は,近位部は右肺動脈起始部となり,遠 位部は背側大動脈との連絡を絶つが,左側は,近位 部は左肺動脈起始部を形成し,遠位部は動脈管 (ductus arteriosus)として存続する.左右肺動脈起始 部は左右末梢肺動脈と接続する. ⑥ 大動脈囊:両側に引き伸ばされ,右側は腕頭動脈を, 左側は総頸動脈の起始部を形成する. ⑦ 背側大動脈:左右対称に発生し,分枝として分節間 動 脈 が 認 め ら れ る. 第 7 分 節 間 動 脈(segmental artery)は左右とも鎖骨下動脈を形成する.咽頭弓動 脈系のリモデリングと胎児頸部・胸部の形態形成に 伴い,左背側大動脈の第 3 ∼ 7 分節は短縮する.こ の短縮により左第 7 分節間動脈からリモデリングし た左鎖骨下動脈は頭側へ移動し,左総頸動脈の直後 に起始することになる.右背側大動脈の第 3 ∼ 7 分 節も短縮し,右第 7 分節間動脈は右第 IV 咽頭弓動 脈と接続して右鎖骨下動脈遠位部を形成する.第 10以下の分節間動脈は肋間動脈となる.右背側大 動脈第 8 分節,および第 III ∼ IV 咽頭弓動脈間の 背側大動脈(頸動脈管)はリモデリングにより閉塞・ 消失する. 2)大動脈弓発生異常の考え方 正常の大動脈弓発生過程では,前述のように大血管 リモデリングにより咽頭弓動脈および背側大動脈の一 部は存続・形態改変し,一部は退縮・消失する.大血 管リモデリング,すなわち存続と退縮のパターンの異 常により,大動脈弓異常が発生する(Fig. 2).特に左 右の第 IV 咽頭弓動脈,第 VI 咽頭弓動脈末梢側(動脈 管),背側大動脈の存続・退縮がどのように生じるか を考えることが,正常な大動脈弓の発生および大動脈 弓の異常を理解するうえで重要である. 3.心臓流出路の発生 (1) 胎生期の流出路(円錐動脈幹)は右室から起始 ・円錐口の左方移動(looping) ・円錐動脈幹の回旋(隆起=中隔がねじれて形成) 大動脈−左心室 肺動脈−右心室 (2) 二次心臓領域細胞と心臓神経堤細胞の相互作用が 必要 ・二次心臓領域細胞:流出路心筋・平滑筋を形成 ・心臓神経堤細胞:隆起=中隔を形成 (3) 流出路中隔の形成異常:PTA,TOF,TOF 型 DORV 1)心臓流出路の形成1) 心臓流出路(outflow tract)は,胎生期の円錐動脈幹
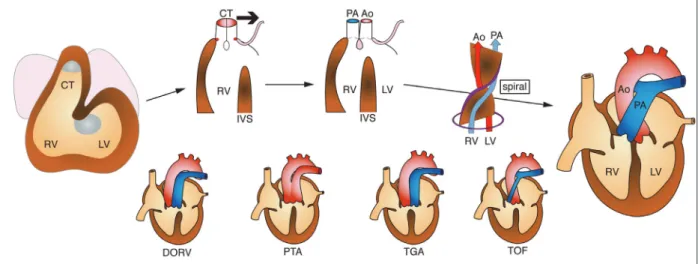
Fig. 3 Development and defects of the cardiac outflow tract. See detail in the text. Bold arrow represents leftward movement of conotruncus (CT).
RV: right ventricle, LV; left ventricle, IVS: interventricular septum, Ao: aorta, PA: pulmonary artery, DORV: double outlet right ventricle, PTA: persistent truncus arteriosus, TGA: transposition of the great arteries, TOF: tetralogy of Fallot.
から形成される.胎生 4 週頃,原始心筒は looping し て左右心室の形態が明らかになり,一本の導管の形態 をした円錐動脈幹が,右心室原基(心球部:bulbus cordisとも呼ばれる)と大動脈囊の間で伸長する(Fig. 3).円錐動脈幹には,左側および右側から円錐動脈幹 隆起(conotruncal cushion/swelling)が形成され,円錐動 脈幹(特に円錐部)の短縮に伴ってねじれ(spiral)なが ら癒合し,円錐動脈幹中隔を形成していく.同時に, 円錐動脈幹の右心室への開口部(円錐口)は左方へ移動 していく.その結果,肺動脈と大動脈が分離し,それ ぞれ右心室と左心室に整列(alignment)する.円錐部(漏 斗部)中隔と膜様部心室中隔が接合し,肺動脈円錐が 存続,大動脈円錐が吸収されることにより,大動脈弁 僧帽弁線維性結合(aortico-mitral fibrous continuity)が形 成される. まとめると,流出路の発生は,(1)円錐動脈幹が形成 されて伸長し,looping に伴って,(2)円錐動脈幹隆起 から中隔の形成,(3)円錐動脈幹の短縮,(4)円錐口の左 方移動,が同時進行し,(5)円錐部(漏斗部)中隔と膜様 部中隔の接合,(6)肺動脈円錐の存続と大動脈円錐の吸 収,により完成する(Fig. 3). 2)心臓流出路の異常1) 円錐動脈幹の発生異常に起因する心臓流出路の異常 は,先天性心疾患の約 30%を占める.円錐動脈幹異 常は,多くの場合,新生児期から症状を有し,内科的・ 外科的治療を必要とする.また,症例によって病態に バリエーションが多く,いまだに予後不良な場合もあ るが,適切な診断・治療・管理によって予後を改善す ることができる可能性も高いため,小児循環器科の診 療において特に重要である.主要な心臓流出路異常に ついては,すべて証明されているわけではないが,以 下のような機序が推定されている(Fig. 3).
① 両大血管右室起始(double outlet right ventricle:DORV): 円錐口の左方移動の障害,円錐の存続・吸収の異常 ② 完全大血管転位(transposition of the great arteries:TGA):
円錐動脈幹の回旋不全,円錐の存続・吸収の異常 ③ 総動脈幹遺残(persistent truncus arteriosus:PTA):円
錐・動脈幹中隔の形成不全
④ Fallot 四徴症(tetralogy of Fallot:TOF):肺動脈円錐 の低形成,大動脈の malalignment
3)心臓流出路を形成する細胞4)
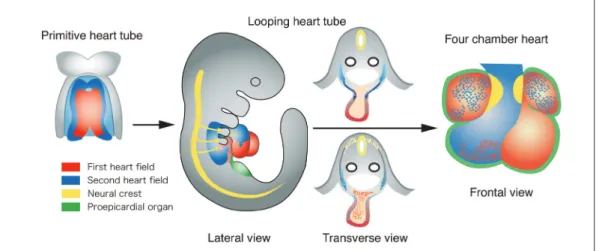
円錐動脈幹の発生には,原始心筒を形成する側板中 胚葉(lateral plate mesoderm: LPM)の細胞(一次心臓領 域:first heart field:FHF と呼ばれる)と,前述のよう に心臓神経堤細胞が重要な役割を果たすことが以前か ら知られていた.2001 年,これらの細胞の他に,臓 側中胚葉(splanchnic mesoderm)領域に由来し,原始心 筒が looping し始める頃に,咽頭弓に連なる心筒の流 出路側から流入し,心筋・心内膜層および平滑筋層を 形成する心臓前駆細胞が発見された.この新たな心臓 前駆細胞の発生領域は,二次心臓領域(second heart field:SHF)と呼ばれるようになった.さらに近年, 二次心臓領域は心臓の流出路側だけでなく,流入路側 からも流入し,心房の一部を形成する心筋細胞を供給 することが示された. Fig. 4 に心臓流出路の形成に関与する,由来の異な
Fig. 4 Developmental origins of the outflow tract and other regions of the heart. The first heart field (red) gives rise to the primitive heart tube and eventually the left ventricle, the atria, and a part of the right ventricle. The second heart field (blue) gives rise to the outflow tract, the right ventricle, and a part of the atria. The cardiac neural crest (yellow) gives rise to the outflow tract cushion that eventually forms the outflow tract septum. The proepicardial organ (green) gives rise to the epicardium. See more details in the text.
る細胞の動きを示す.側板中胚葉由来の細胞は,胚の 正中に原始心筒(Fig. 4:赤)を形成する.咽頭弓臓側 中胚葉の二次心臓領域に由来する細胞(Fig. 4:青)は, 原始心筒が形成された頃に動脈幹部に移動し(Fig. 4:青矢印),流出路心筋層(一部,心内膜層)に分布 する.神経管背側(Fig. 4:黄)に起源する心臓神経堤 細胞(Fig. 4:黄)は,原始心筒が looping する頃,動脈 幹部に移動し(Fig. 4:黄矢印)円錐動脈幹隆起に沿っ て分布し,その成長に関与する.心臓流出路の形成に は,二次心臓領域由来の細胞(青色),心臓神経堤細胞 (黄色)の相互作用が必要である.最終的に,一次心臓 領域由来細胞は主に左心室と心房(Fig. 4:赤),二次 心臓領域由来細胞は主に右心室と流出路を形成する (Fig. 4:青). 4.TBX1 と二次心臓領域の研究の基礎・臨床医学的インパクト (1) 分子遺伝学的 ・ TBX1 は 22q11.2 欠失症候群の先天性心疾患の主 要責任遺伝子 (2)心臓発生学的(メカニズム) ・ 二次心臓領域細胞は,流出路と右心室(そして心 房の一部)を形成 ・ Tbx1 は二次心臓領域に発現し,心臓神経堤細胞 には発現しない ⇨先天性心疾患の発症に関する新たなコンセプト ・ 二次心臓領域と心臓神経堤細胞の異常による 流出路異常のスペクトラム 1)TBX1 と 22q11.2 欠失症候群 T-box 型転写因子をコードする TBX1 は,心臓流出 路の異常を高率に合併する 22q11.2 欠失症候群の主要 な責任遺伝子である5).Tbx1 ヘテロ変異マウスでは 10∼ 50%に大動脈弓異常が,Tbx1 ホモ変異マウスで は総動脈幹遺残,胸腺・副甲状腺低形成,口蓋裂,顔 面頭蓋骨異常など,22q11.2 欠失症候群の主要症状す べてが認められる6-8). 私たちは,心臓流出路の発生における Tbx1 の発現 を解析し,Tbx1 が二次心臓領域由来の心臓前駆細胞 に発現し,心臓神経堤細胞には発現しないことを明ら かにした9, 10).この結果は,私たち(そしておそらく, すべての 22q11.2 欠失症候群の研究者)にとって衝撃 的であった.何故なら従来 22q11.2 欠失症候群のプラ イマリーな病因は,心臓神経堤細胞の異常と考えられ ており,責任遺伝子は心臓神経堤細胞に発現すると推 測されていたからである.さらに私たちは細胞系譜解 析により,Tbx1 を発現した二次心臓領域由来の細胞 は,胎生初期には流出路全体と右心室原基を形成し,胎 生後期∼出生直前にはおもに右心室流出路から肺動脈 主幹部および肺動脈弁を形成することを明らかにした11).
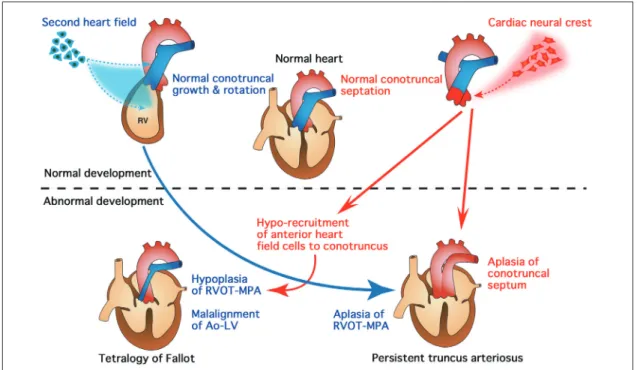
2)心臓流出路異常の発症機構に関する考察 以上の臨床心臓発生学の知見と従来の形態形成学か ら,心臓流出路の発生異常による先天性心疾患(PTA, TOF)について,Fig. 5 のような発症機序が考察される4, 11). PTAは心臓神経堤細胞を除去したニワトリ胚で高率 に認められることから,心臓神経堤細胞の発生異常に より円錐動脈幹中隔が完全欠損して発症すると考えら れる2).TOF の発症機序は完全に解明されているわけ ではないが,円錐動脈幹の不完全な回旋および中隔形 成に起因すると考えられている.すなわち,円錐動脈 幹 の 回 旋 異 常 に よ り 流 出 路 中 隔 と 筋 性 部 中 隔 の alignmentが障害され,結果として大動脈が心室中隔 欠損の上に騎乗する.そして円錐動脈幹隆起により異 常に前方偏位した中隔が形成され,肺動脈弁・漏斗部 狭窄が発生するという説である12).これに対し,Van Praaghは肺動脈弁下漏斗部(または円錐)の低形成が, 漏斗部狭窄と流出路中隔の malalignment の原因である とする説を唱えている12).二次心臓領域が発見される 以前には,TOF の発症機序は心臓神経堤細胞の発生 異常によって説明されることが多かった.しかし,最 近の研究成果から,TOF の発症にはむしろ二次心臓 領域細胞の発生異常が大きく関与すると考えられる. 二次心臓領域細胞が形成する肺動脈弁下漏斗部(円錐) の低形成の結果として TOF が発症すると考える,Van Praaghの説に近い考え方である.一方,心臓神経堤細 胞の発生異常も二次的に二次心臓領域細胞の流出路へ の供給を障害することから,同様に TOF 発症の原因 となり得る.二次心臓領域細胞の発生異常が非常に高 度で,主肺動脈および漏斗部がまったく形成されなけ れば,心臓からの流出路は大動脈だけになり,肺動脈 閉鎖兼心室中隔欠損ないし PTA の形態になる場合も あると考えられる4, 11). おわりに 臨床心臓発生学は,自然・生物の神秘を探求するも のであり,先天性心疾患の成因解明と予防・再生医療 への基礎として発展すべきものである.今世紀,心臓 流出路の発生における心臓神経堤細胞と二次心臓領域 細胞の相互作用の重要性が明らかとなり,多彩な心臓 流出路異常のスペクトラムが形成されるメカニズムに 新たな考察が生まれた.しかし,この考察も,臨床心 臓発生学のさらなる発展により書きかえられ,着実に “自然の本質”に近づいていくことが期待される. 臨床心臓発生学の最大の課題として,複雑な心臓大 血管形成を制御する多くの分子の相互作用からなる ネットワークが,どのように形態形成に機能している のか,すなわち“From gene to morphology”を解明する ことがあげられる.革新的な研究・発想が求められて いる.本稿が,多くの忙しい若手臨床医にとって,心 臓大血管の発生と先天性心疾患の基礎について,理解 を深める一助となれば,誠に幸いである. 謝 辞 講演の機会を与えてくださいました,日本小児循環器学 会の皆様に深謝します.また,図を作成していただきまし た山岸千尋先生に感謝します. 【参 考 文 献】 ̶̶̶̶̶̶̶̶̶̶̶̶̶̶̶̶̶̶̶̶ 1) 山岸敬幸 : 流出路の発生とその異常 . 山岸敬幸 , 白石 公 , 編 : 先天性心疾患を理解するための臨床心臓発生学 . メ ジカルビュー社 , 東京 , 2007, pp131-139
2) Kirby ML, Waldo KL: Role of the neural crest in congenital heart disease. Circulation 1990; 82: 332-340
3) 内田敬子 , 山岸敬幸 : 大動脈弓の発生とその異常 . 山岸 敬幸 . 白石 公 , 編 : 先天性心疾患を理解するための臨床 心臓発生学 . メジカルビュー社 , 東京 , 2007, pp143-150 4) 山岸敬幸 , 山岸千尋 : トピックス 二次心臓形成領域 . 山 岸敬幸 , 白石 公 , 編 : 先天性心疾患を理解するための臨 床心臓発生学 . メジカルビュー社 , 東京 , 2007, pp140-142
5) Yamagishi H, Srivastava D: Unraveling the genetic and developmental mysteries of 22q11 deletion syndrome. Trends Mol Med 2003; 9: 383-389
6) Merscher S, Funke B, Epstein JA, et al: TBX1 is responsible for cardiovascular defects in velo-cardio-facial/DiGeorge syndrome. Cell 2001; 104: 619-629
7) Jerome LA, Papaioannou VE: DiGeorge syndrome phenotype in mice mutant for the T-box gene, Tbx1. Nat Genet 2001; 27: 286-291
8) Lindsay EA, Vitelli F, Su H, et al: Tbx1 haploinsufficieny in the DiGeorge syndrome region causes aortic arch defects in mice. Nature 2001; 410: 97-101
9) Garg V, Yamagishi C, Hu T, et al: Tbx1, a DiGeorge syndrome candidate gene, is regulated by sonic hedgehog during pharyngeal arch development. Dev Biol 2001; 235: 62-73 10) Yamagishi H, Maeda J, Hu T, et al: Tbx1 is regulated by
tissue-specific forkhead proteins through a common Sonic hedgehog-responsive enhancer. Genes Dev 2003; 17: 269-281 11) Maeda J, Yamagishi H, McAnally J, et al: Tbx1 is regulated
by forkhead proteins in the secondary heart field. Dev Dyn 2006: 235; 701-710
12) Siwik ES, Patel CR, Zahka KG: Tetralogy of Fallot. In Allen HD, et al. eds. Moss and Adams’ Heart Disease in Infants, Children, and Adolescnets including The Fetus and Young Adult. Lippincott Williams & Wilkins, Philadelphia, 2001