要約 グッピーは南米東北部原産の淡水魚で、観賞魚や蚊防除対策として世界各地で飼育あるいは放流されている。日 本でも観賞魚として一般的である一方、沖縄や温泉地などで野生化した集団が総合対策外来種に指定されている。 これらの日本におけるグッピーの導入起源や経路は不 明である。そこで本研究では日本のグッピーの起源を mtDNA における D-loop 領域約 320bp の塩基配列を用 いて調べた。沖縄などで野生化しているグッピー 16 集 団と観賞魚としてクローズドコロニーとして継代維持 連絡先:中嶋正道 東北大学大学院農学研究科 仙台市青葉区荒巻字青葉 468-1 (e-mail: [email protected]) 2020 年 1 月 7 日投稿 2020 年 6 月12日受理 〔原著論文〕
mtDNA の D-loop 領域の塩基配列より推定された日本におけるグッピー
Poecilia reticulata の起源
佐藤龍星
1、中嶋正道
1 1:東北大学大学院農学研究科、仙台市青葉区荒巻字青葉 468-1Origin of the guppy Poecilia reticulata in Japan by means of sequence of mtDNA
D-loop region
Ryusei SATOU
1and Masamichi NAKAJIMA
1Laboratory of Fish Genetics and Breeding Science, Graduate School of Agricultural Science, Tohoku University, Aoba, Sendai 980-0845, Japan
ABSTRACT
The guppy (Poecilia reticulata)is small freshwater fish native to South America from Venezuela to Brazil and has been introduced in Japan not only as ornamental fish but also as a biological control to mosquito. These introductions resulted some of populations colonized as an exotic species in Okinawa southern Island of Japan. The present study aimed to estimate introduction sources of the guppies in Japan by means of sequence analysis of 320 bp of mtDNA D-loop region. Sixteen wild populations and fourteen ornamental strains were examined. Six haplotypes (Hap1 ~ Hap6) were detected from the sequence analysis. All ornamental strains except Endrer's and populations collected in Singapore, Thailand and Venezuela had Hap1 which suggest the introduction sources of the haplotype from West Trinidad. Endrer's has Hap4 which suggest the introduction from East Venezuela and this haplotype was observed only in Endrer's in the present study. On the other hand, colonized populations collected from Okinawa Island have five types of haplotype, Hap1, Hap2, Hap3, Hap5 and Hap6. These haplotypes were originated from wide area of Northern South America. Hap2 was observed in North Guyana, Hap3 was observed in East Trinidad, Hap5 was observed in Suriname and East Guyana and Hap6 was observed in West Trinidad. Genetic variabilities in colonized populations of Okinawa Island were lower than that of original populations in South America. These results suggest that the origin of ornamental strain in Japan was originated from specific narrow area and small number of individuals, probably from West Trinidad. Meanwhile data of colonized populations in Okinawa Island suggested occurrence of multiple introduction or invasion from many origins from different area.
緒言 グッピー(Poecilia reticulata)はカダヤシ目カダヤ シ科グッピー属に分類される魚類で、南米北東部を原 産とする卵胎生の熱帯性淡水魚である。グッピーは、 2 ~ 4 ヵ月で成熟し、妊娠期間が約 1 ヵ月と世代交代 が早く、飼育が容易であり、観賞魚として高い人気が ある。また、雄は体色や模様、尾鰭の形が非常に多様 性に富み、主に雄の形質に関して選択された多くの系 統が作出されている(岩松 , 1993)。 観賞魚としてのグッピーの歴史は比較的浅い。1850 年に南米トリニダード島に生息する小型の魚として報 告されて以降、1920 年頃にはイギリスやアメリカ、 ドイツなど一部の国で観賞魚として飼育されるように なったのが始まりとされている。その後 1960 年代後 半までアメリカやドイツが中心となってグッピーの品 種改良が行われ、様々な系統が作出された。その後、 観賞用グッピーの市場の中心はシンガポールに移り、 シンガポールで養殖したグッピーが世界中に輸出され ている(岩崎 , 1997)。日本では 1930 年頃に観賞用と して輸入され、1960 年頃から一般的にも飼育される ようになった(岩崎 , 1997)。このように、日本にお ける観賞魚としてのグッピーの起源は南米トリニダー ド島と考えられるが遺伝的解析による起源の推定は行 われていない。 一方、グッピーは外来種としても有名である。グッ ピーの本来の生息域はベネズエラやガイアナ、スリナ ム、トリニダード・トバゴを含む南米北東部とされて いる(Rosen と Bailey, 1963)。しかし、人為的分散に より近年も生息域を拡大しており、自然生息域を除き 最低でも 69 ヵ国における野外での定着が確認されて いる(Daecon ら , 2011)。グッピーの他地域への導入 経路は主に 2 通りに分けられる。1 つは、蚊の対策と してグッピーを持ち込むという生物学的防除の意味を 持つ導入である(Houde, 1997)。インドやオーストラ リアでは水路や井戸へのグッピーの導入により、ボウ フラが駆除され、マラリア対策としての効果が挙げら れている(Ghosh ら , 2005;Borthwick, 1923)。もう 1 つは、観賞用飼育個体の野外遺棄である(McKay, 1984)。この 2 つの経路がグッピーへ分布拡大につな がる導入の機会を与えている。また、グッピーは温度 耐 性(Kanda ら , 1991)、 塩 分 耐 性( 鹿 野 と 藤 尾 , 1994)といった高い環境適応性や、1 匹の雌が 60 匹 を超える仔魚を一度に産むなど高い繁殖能力(Deacon ら , 2011)を有しておりこれらの性質が分布拡大に影 響していると考えられている。 日本の沖縄地方の河川で野生化したグッピーが見ら れるようになったのは 1970 年代の初め頃とされ(幸 地 , 1991)、主な導入経路は観賞用飼育個体の野外遺 棄と考えられている(石川ら , 2013)。1973 年には、 那覇市の天久や宜野湾市の伊佐など米軍基地周辺の溝 で見られる程度だったが、1979 年には中南部地域に 広くみられるようになった(幸地 , 1991)。そして、 嶋津(2011)による沖縄県での外来種の分布調査では、 グッピーは沖縄本島の 300 水系中 120 水系で確認され、 沖縄県の遊泳魚が分布する水系の 68% への侵入が明 らかとなった。日本において、グッピーは越冬可能な 水温である沖縄県の河川や温泉地しか定着できないも のの、定着可能な地域での広範な分布やその生態的影 響から、総合対策外来種に指定されている(環境省 我が国の生態系等に被害を及ぼすおそれのある外来種 リ ス ト よ り https://www.env.go.jp/press/files/jp/26594. pdf)。しかし、日本において定着した野生化グッピー の起源に関する研究は観賞魚の場合と同様行われてい ない。 一般的に、外来種の定着が生物多様性を脅かす脅威 であることは広く知られている(Vitousek ら , 1997)。 されている 14 系統を分析の対象とした。また比較のため南米ベネスエラの野生集団と東南アジアで野生化した 2 集 団(タイ、シンガポール)も分析した。その結果、6 ハプロタイプが得られ、このうち観賞魚系では Hap1 と Hap4 が観察され、Hap4 は Endrer's のみで観察された。Hap1 は西トリニダードで、Hap4 は東ベネズエラで報告されて いる。一方東南アジア野生化集団とベネズエラ集団でも Hap1 のみが観察されたのに対し、沖縄の野生化集団から は 5 種類のハプロタイプ(Hap1 ~ 3, 5, 6)が観察された。また、野生化集団における遺伝的変異性(ハプロタイプ数、 ハプロタイプ多様度、塩基多様度)は起源となった南米各地での変異性よりも低かった。これらの結果から、日本 における観賞魚系はトリニダード島のごく狭い範囲の少数個体、おそらくは西トリニダードを起源としている可能 性が示された。また、沖縄の野生化集団の起源は南米各地を起源とする個体の複数回の導入が起源となっている可 能性が示された。 キーワード:グッピー、ミトコンドリア DNA、D-loop、外来種、ハプロタイプ
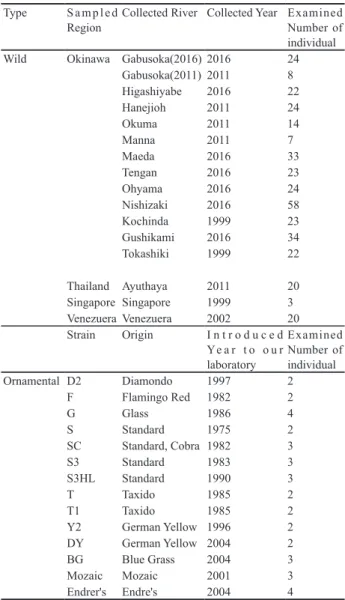
外来種がもたらす生物多様性への問題として捕食や競 合による他の在来生物の排除、近縁の在来種との交雑 による遺伝的浸食、生息環境の破壊、病害の伝播があ る( 鷲 谷 と 矢 原 , 1996)。 ブ ラ ッ ク バ ス( 青 木 ら , 2006)やブルーギル(河村ら , 2004)による食害など は日本でも見られ、非常に有名である。生態的な影響 が大きいとされるブラックバスは分布調査のほか、遺 伝学的な研究もおこなわれ、導入の起源や定着、分散 の過程について知見の蓄積が進んでいる(青木ら , 2006;北川ら , 2005;高村 , 2005;Yokogawa, 1998)。グッ ピーでは、分布調査(石川ら , 2013)による定着の現 状についての知見は蓄積されているものの、遺伝学的 な研究はほとんど行われていない。外来種の定着とそ の後の影響の予防のためにも、予想される侵入経路を 事前に予測し、対策を取る必要がある。さらに、すで に定着した生物についても、その導入の起源や定着過 程を調査し、把握しておくことが未定着地域への侵入 定着と定着地域への再導入防止のために重要である (高村 , 2005)。 グッピーは外来種として海外にも広く定着している ため、原産地である南米北東部や導入された地域の集 団に関する遺伝学的研究は多く報告されている。トリ ニダードのグッピー集団の遺伝的集団構造に関して、 アロザイム多型や mtDNA などの DNA 変異を用いた 研究が行われている。これらの研究によると、トリニ ダードのグッピー集団は西部と東部の水系間で大きな 遺伝的分化が見られており、約 60 ~ 120 万年前に分 岐したと推定されている(Carvalho ら , 1991;Fajen と Breden, 1992)。また、他地域の集団を含む系統解析に よると、ベネズエラ集団は西部トリニダード集団と系 統的に近縁であり、東部トリニダード集団とガイアナ 集団は単系統群をそれぞれ形成したという(Lindholm ら , 2005)。これらの結果は、グッピーは原産地であ る南米北東部において、いくつかの遺伝的に異なる集 団が存在することを示している。また、オーストラリ アでは 1910 年の生物学的防除を目的とした導入が最 初の導入とされ、以降複数回の導入報告がある(Vipan, 1910;Patrick 1987)。グッピーはオーストラリアにお いても観賞魚として有名なため、飼育個体の野外遺棄 による導入が行われた可能性があるという報告もある (McKay, 1984)。 こ う し た 報 告 を 受 け て 行 わ れ た mtDNA やマイクロサテライト DNA マーカーを用い た遺伝的集団構造解析の結果、生物学的防除による導 入と飼育個体の野外遺棄による導入のそれぞれの遺伝 的な証拠が示され、オーストラリアにおけるグッピー の異なる系統からの複数回の導入が明らかにされた (Lindholm ら , 2005)。 そこで本研究は、日本における観賞魚と野生化グッ ピーの mtDNA の D-loop 領域(320bp)を用いて、日 本に定着したグッピーの遺伝的起源の推定を目的とし た。 材料と方法 分析に用いたサンプル 分析に用いたサンプルの情報を図1、表1に示す。 本研究では、観賞魚系統 14 系統 37 個体、海外野生化 集団 3 集団 42 個体、日本野生化集団 14 集団 277 個体 のサンプルを用いて、mtDNA の Dloop 領域 320bp の 塩基配列を決定した。野生化集団は 2016 年に採集し たサンプル(沖縄県内 7 か所)とそれ以前に採集し、 本研究室でクローズドコロニーとして飼育していたサ ンプル(沖縄県内 6 か所)を「日本野生化集団」、そ れ以外を「海外野生化集団」とした。また、本研究室 で継代飼育されているクローズドコロニーから、観賞 魚 系 統 と し て D2、F、G、DY、BG、S、SC、S3、 S3HL、T、T1、Y2、Endrer's、Mozaic の 14 系 統 を 用 いた。これら 14 系統は全てペットショップから購入 した個体を起源とし、当研究室でクローズドコロニー として維持されている。S 系統は 1975 年に当研究室 に導入されたスタンダードと呼ばれる野生型に近いと される系統、SC は 1983 年に S 系統の雌とキングコブ ラの雄との交配個体を起源とする系統、S3 は 1983 年 に S 系統から分離された系統、S3HL は 1990 年に S3 系統から高温に強い個体を選抜し作成した系統であ る。D2 系統は 1992 年にダイアモンド(D)として導 入された個体の中から黄色素胞を有さない個体を選抜 Okuma R. Hanejioh R. Gabusoka R. Manna R. Higashiyabe R. Maeda R. Tengan R. Ohyama Kochinda Gushikami Nishizaki Tokashiki Fig. 1
して作成した系統、F 系統は 1982 年に導入されたフ ラミンゴレッドの系統、G 系統は 1987 年に導入され たキングコブラ(当時はグラスと呼ばれていた)を起 源とする系統、T は 1985 年に導入されたタキシードで、 T 1は同年にタキシードから体色がゴールデンの個体 を 分 離 し て 作 成 し た 系 統。Y2 は 1996 年 に、DY は 2004 年に導入されたそれぞれ異なるドイツイエロー を起源としている。BG は 2004 年に導入されたブルー グラスを起源としている。Mozaic は 2001 年にシンガ ポールから導入した系統。Endrer's は 2004 年に導入さ れた系統である。海外野生化集団としてシンガポール、 アユタヤ、ベネズエラの 3 集団を用いた。海外野生化 集団は現地の小川で繁殖していた個体を採集した。シ ンガポールとアユタヤは現地の研究者に、ベネズエラ は観賞魚の輸入業者(神畑養魚 ( 株 ))に採集を依頼 した。それぞれの個体は採集後、DNA 抽出用に尾鰭 の一部を切り取り、99.5% エタノールに入れて保存し た。 DNA は各個体の尾鰭のサンプルより通常のフェ ノール・クロロホルム法を用いて抽出した。抽出した DNA は 50µl の TE buffer(10mM Tris-HCl, 1mM EDTA) に溶解し、- 20℃で保存した。
mtDNA (D-loop 領域)の増幅と塩基配列の決定
D-loop 領域の増幅用プライマーとして、L15926 (5'- TCAAAGCTTACACCAGTCTTGTAAACC -3'; Kocher ら , 1989)および H16498 (5'- CCTGAACTAGGAACCA GATG -3'; Shields と Kocher, 1991)を用いて PCR によ り mtDNA の D-loop 領域を増幅した。PCR 反応液は、 1 サンプルにつき DW を 2.6µl、2 × Mighty Amp Buffer Ver.3(TaKaRa)を 5.0µl、各プライマー(10pmol)を 0.6µl ずつ、Mighty Amp® DNA Polymerase Ver.3(1.25U/µl TaKaRa)を 0.2µl を加え、各鋳型 DNA サンプルを 1.0µl 加えて総量 10µl の反応液とした。PCR 反応は 98℃ -10 秒、49℃ -20 秒、68℃ -30 秒を 1 サイクルとして 25 サイクル行った。
PCR によって増幅した遺伝子配列の塩基配列決定 は Cycle Sequence 法で行った。Cycle Sequence 反応液 の組成は、1 サンプルあたり DW 6.25µl、Big Dye 5 × Sequencing Buffer v3.1 を 1.75µl、プライマー(10pmol/ µl) 0.5µl、Big Dye v3.1 Sequencing RT-100 0.5µl、精製 後の PCR 産物 1.0µl を加えて総量 10µl の反応液とした。 また、プライマーは L15926 を用いた。Cycle Sequence は、96℃ -10 秒、50℃ -5 秒、60℃ -2 分を 1 サイクル として 25 サイクル行った。 塩基配列の決定には 3500xl Genetic Analyzer(Applied Biosystems)を使用した。 系統解析 塩基配列データを MEGA ver.6.06(Tamura ら , 2013) に入力し、実装されている MUSCLE(Edgar, 2004)を 用いて複合アライメントを行った。各集団で重複して 見られたハプロタイプは 1 つのハプロタイプとして絞 り、データセットに用いた。系統解析として本研究で 確認された 6 つのハプロタイプ、Genbank に登録され ている 67 のハプロタイプ(表 2:Lindholm ら , 2005; Taylor と Breden, 2000; Alexander と Breden, 2004, unpublished)、外群として Genbank に登録されている
Poecilia parae (Lindholm ら , 2005)の配列を加えた 74
の塩基配列(322bp)を用いて、近隣結合法に基づく 系統樹(Saitou と Nei, 1987)を作成した。モデルは MEGA に実装されている Kimura 2- parameter model を Type Sampled
Region Collected River Collected Year Examined Number of individual Wild Okinawa Gabusoka(2016) 2016 24
Gabusoka(2011) 2011 8 Higashiyabe 2016 22 Hanejioh 2011 24 Okuma 2011 14 Manna 2011 7 Maeda 2016 33 Tengan 2016 23 Ohyama 2016 24 Nishizaki 2016 58 Kochinda 1999 23 Gushikami 2016 34 Tokashiki 1999 22 Thailand Ayuthaya 2011 20 Singapore Singapore 1999 3 Venezuera Venezuera 2002 20 Strain Origin I n t r o d u c e d Ye a r t o o u r laboratory Examined Number of individual Ornamental D2 Diamondo 1997 2 F Flamingo Red 1982 2 G Glass 1986 4 S Standard 1975 2 SC Standard, Cobra 1982 3 S3 Standard 1983 3 S3HL Standard 1990 3 T Taxido 1985 2 T1 Taxido 1985 2 Y2 German Yellow 1996 2 DY German Yellow 2004 2 BG Blue Grass 2004 3 Mozaic Mozaic 2001 3 Endrer's Endre's 2004 4
選択した。各ノードについて、1000 回試行のブート ストラップ値により評価した。作成した系統樹は Figtree ver1.4.3(http://tree.bio.ed.ac.uk/software/figtree/) を用いて描写した。 ハプロタイプネットワーク 本研究で用いたサンプルで確認されたハプロタイプ 間の類縁関係を調べるため、TCS1.21(Clement ら , 2000)に実装されている統計的最節約アルゴリズム (Templeton ら , 1992)に基づいて、ハプロタイプネッ トワークを作成したネットワークを構築する際は、コ ネクションリミットを 50Step に固定し、塩基挿入 / 欠 失を 5th state として扱った。 遺伝的多様性と遺伝的分化の評価 集団の遺伝的多様性は、Arlequin ver.3.5.1.2(Excoffier と Lischer, 2010)を用いて、ハプロタイプ数(H)、ハ プロタイプ多様度 Hd(Nei, 1973)、塩基多様度π(%) (Nei と Tajima, 1981)をそれぞれ算出し、評価した。 ハプロタイプ多様度はハプロタイプ頻度を反映し、塩 基多様度はハプロタイプ頻度に遺伝的距離が反映され る。mtDNA D-loop 部分領域(320bp)を用いて、各集 団間における遺伝的分化の指標である pairwise Φ st (Excoffier ら , 1992)を Arlequin ver.3.5.1.2 で算出し、
遺伝的分化の程度を評価した。 結 果 1.検出されたハプロタイプ 決定した塩基配列データで複合アライメントを行っ た結果、全サンプル 356 個体からハプロタイプは 6 種 類検出され、それぞれ Hap1~6 とした。ハプロタイプ 間の塩基置換サイトは 19 サイトだった(表 3)。また、
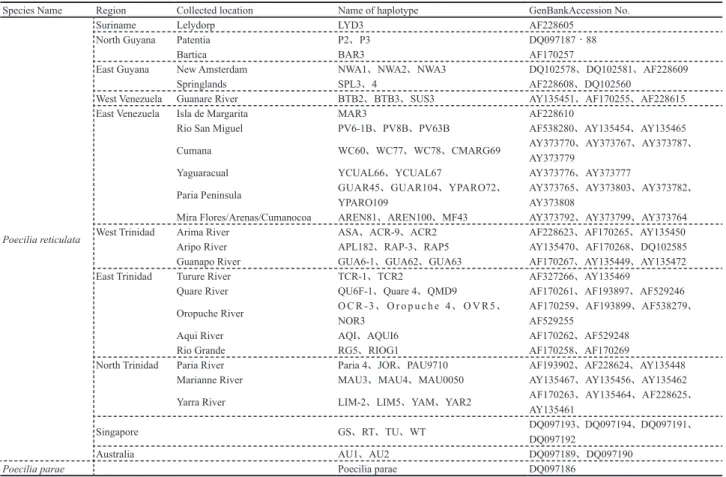
Species Name Region Collected location Name of haplotype GenBankAccession No.
Poecilia reticulata
Suriname Lelydorp LYD3 AF228605 North Guyana Patentia P2、P3 DQ097187‐88
Bartica BAR3 AF170257
East Guyana New Amsterdam NWA1、NWA2、NWA3 DQ102578、DQ102581、AF228609 Springlands SPL3、4 AF228608、DQ102560
West Venezuela Guanare River BTB2、BTB3、SUS3 AY135451、AF170255、AF228615 East Venezuela Isla de Margarita MAR3 AF228610
Rio San Miguel PV6-1B、PV8B、PV63B AF538280、AY135454、AY135465 Cumana WC60、WC77、WC78、CMARG69 AY373770、AY373767、AY373787、
AY373779
Yaguaracual YCUAL66、YCUAL67 AY373776、AY373777
Paria Peninsula GUAR45、GUAR104、YPARO72、YPARO109 AY373765、AY373803、AY373782、AY373808 Mira Flores/Arenas/Cumanocoa AREN81、AREN100、MF43 AY373792、AY373799、AY373764 West Trinidad Arima River ASA、ACR-9、ACR2 AF228623、AF170265、AY135450 Aripo River APL182、RAP-3、RAP5 AY135470、AF170268、DQ102585 Guanapo River GUA6-1、GUA62、GUA63 AF170267、AY135449、AY135472 East Trinidad Turure River TCR-1、TCR2 AF327266、AY135469
Quare River QU6F-1、Quare 4、QMD9 AF170261、AF193897、AF529246 Oropuche River O C R -3、O r o p u c h e 4、O V R5、NOR3 AF170259、AF193899、AF538279、AF529255 Aqui River AQI、AQUI6 AF170262、AF529248
Rio Grande RG5、RIOG1 AF170258、AF170269
North Trinidad Paria River Paria 4、JOR、PAU9710 AF193902、AF228624、AY135448 Marianne River MAU3、MAU4、MAU0050 AY135467、AY135456、AY135462 Yarra River LIM-2、LIM5、YAM、YAR2 AF170263、AY135464、AF228625、AY135461 Singapore GS、RT、TU、WT DQ097193、DQ097194、DQ097191、
DQ097192
Australia AU1、AU2 DQ097189、DQ097190
Poecilia parae Poecilia parae DQ097186
Table 2 Sequence data of mtDNA D-loop region used for the comparison with the datas obtained in this study.
Haplotype 33 36 37 44 52 109 133 157 158 205 209 210 278 283 286 287 301 302 308 Hap1 A A C G C G G G C G A G A A C T C A C Hap2 T ・ A ・ ・ A A ・ ・ A C T ・ ・ ・ G T ・ T Hap3 ・ T A A T A ・ A T A ・ ・ T ・ T G ・ ・ T Hap4 ・ ・ A ・ ・ A ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ Hap5 ・ ・ A ・ ・ A ・ ・ ・ A ・ ・ ・ ・ ・ G ・ G ・ Hap6 ・ ・ A ・ ・ ・ ・ ・ ・ ・ ・ ・ ・ G ・ ・ ・ ・ ・
Hap1、2、4、6 が 既 報 の ハ プ ロ タ イ プ と 一 致 し、 Hap3、5 は既報のハプロタイプと一致せず、新規のハ プロタイプだった。 2.各系統・集団内で検出されたハプロタイプ 各系統・集団内で検出されたハプロタイプとそのハ プロタイプを持つ個体数を表 4 に示した。また、観賞 魚系統 14 系統について見てみると、全サンプル中エ ンドラーズの系統のみ Hap4 であったが、それ以外の 系統は全て Hap1 であった。海外野生化集団では、3 集団全てが Hap1 であった。日本野生化集団は 5 つの ハ プ ロ タ イ プ(Hap1、2、3、5、6) が 確 認 さ れ た。 Hap1 は Maeda、Nishizaki、Tokashiki で単型として見 ら れ、Gushikami、Higashiyabe、Ohyama、Hanejioh、 Kochinda でも確認された。Hap2 は Gabusoka(2016)、 Gabusoka(2011)、Okuma で 単 型 と し て 見 ら れ、 Gushikami、Higashiyabe、Hanejioh、Kochinda でも確認 された。Hap3 は Tengan、Manna で単型として見られ、 Gushikami、Higashiyabe でも確認された。Hap5、6 は Ohyama でのみ確認された。 3. mtDNA に基づくハプロタイプ間の系統類縁関係推 定 本研究で検出されたハプロタイプの遺伝的起源を推 定するために、GenBank の配列情報を加えて近隣結合 法に基づく系統樹を構築した(図 2)。各枝の上にブー トストラップ確率(2 桁の整数)を示す。ハプロタイ プ名の右側にそれぞれの遺伝的起源とされる国名を示 した。本研究で検出されたハプロタイプに注目すると、 0.02 GUA6-1 GUA62 PV8B CMARG69 AQUI6 LIM-2 NWA1 TCR-1 TCR2 WC78 RT MAU3 AU2 APL182 SPL3 WC60 ASA QMD9 ACR2 Quare4 MF43 ACR-9 MAR3 WC77 AQI NWA2 Y A M YCUAL67 RAP-3 LIM5 SPL4 YAR2 NOR3 RIOG1 RG5 GS AU1 AREN81 YCUAL66 OCR-3 NWA3 MAU0050 W T P3 AREN100 Oropuche4 GUAR45 P2 MAU4 QU6F-1 RAP5 OVR5 Hap3 TU PAU9710 Poecilia_parae GUA63 JOR Paria4 BAR3 East Trinidad North Guyana West Trinidad East Venezuela North Trinidad Suriname East Guyana East Venezuela East, West Venezuela 94 59 94 51 65 67 74 58 68 57 55 BTB3 SUS3 65 GUAR104 YPAR0109 BTB2 YPARO72 PV6-1B 65 PLVY6D33B Hap5 73 67 83 61 63 Hap2 Hap1 Hap6 Hap4 Fig.2 Fig.2 Genetic relationship among haplotypes observed in this study and registered sequences in data base.
Population, Strain N Hap1 Hap2 Hap3 Hap4 Hap5 Hap6
Gabusoka(2016) 24 24 Gabusoka(2011) 7 7 Higashiyabu 22 20 1 1 Hanejioh 24 15 9 Okuma 14 14 Mannna 7 7 Maeda 30 30 Tengan 23 23 Ohyama 24 16 4 4 Nishizaki 58 58 Kochinda 3 2 1 Gushikami 34 5 28 1 Tokashiki 4 4 Ayuthaya 19 19 Singapore 20 20 Venezuera 3 3 D2 2 2 F 2 2 G 4 4 DY 2 2 BG 3 3 S 2 2 SC 3 3 S3 3 3 S3HL 3 3 T 2 2 T1 2 2 Y2 2 2 Mozaic 3 3 Endrer's 4 4
Table 4 Detected haplotype and its observed number of individual in eacu collected location and strain.
観賞魚系統、海外野生化集団が含まれた Hap1 は West Trinidad で見つかったハプロタイプに近い関係を示し た。次に、日本野生化集団で多く見られた Hap2 は、 North Guyana で見つかったハプロタイプのクレードに 含まれた。Hap3 は East Trinidad で見つかったハプロ タイプに近い関係を示した。観賞魚系統のエンドラー ズだけで見つかった Hap4 は East Venezuela で見つかっ たハプロタイプに近い関係を示した。日本野生化集団 の Ohyama だ け で 見 つ か っ た Hap5、6 は そ れ ぞ れ Suriname・East Guyana と West Trinidad で見つかるハプ ロタイプに近い関係を示した。Hap1、Hap3、Hap4 で はブートストラップ値による支持は得られなかった。 これは今回調べた D-loop 部分領域が 320bp と短かっ たことに起因すると考えられる。正確性を高めるため にはより長い領域を用いた分析を行う必要がある。 4.mtDNA に基づくハプロタイプ間の類縁関係推定 mtDNA の Dloop 領域(320bp)に基づき、本研究で 検出されたハプロタイプについて統計的最節約法によ る ハ プ ロ タ イ プ ネ ッ ト ワ ー ク を 構 築 し た( 図 3)。 Hap1 が最も多くの系統・集団で共有されている。そ の Hap1 から 2 塩基置換で Hap4、6 が派生している。 さらに Hap4 から 3 塩基置換で Hap5 が派生し、8 塩基 置換で Hap2 が、9 塩基置換で Hap3 が派生しているこ とがわかった。Hap2、3、5、6 は観賞魚系統で確認さ れず、日本野生化集団でのみ確認された。 5.mtDNA に基づく各集団の遺伝的多様性 mtDNA の Dloop 領域(320bp)に基づく、各集団の 遺伝的多様性の指標となるハプロタイプ多様度 Hd、 塩基多様度π(%)を日本野生化集団の集団毎に算出 した(表 5)。集団中で 3 つのハプロタイプが検出さ れ た Gushikami 集 団 で は Hd=0.308、 π =1.000、 Higashiyabe 集団では Hd=0.178、π =0.612、Ohyama 集 団では Hd=0.522、π =0.598 だった。集団中で 2 つの ハ プ ロ タ イ プ が 検 出 さ れ た Hanejioh 集 団 で は Hd=0.489、 π =1.529、Kochinda 集 団 で は Hd=0.667、 π =2.083 だった。それ以外の集団は単型だった。 6.mtDNA に基づく集団間の遺伝的分化 D-loop 領域(320bp)の塩基配列に基づき、日本野 生化集団間の遺伝的分化の指標 pairwise φ ST を算出 した(表 6)。φ ST 値は 0.086 ~ 1.000 の範囲をとった。 Hap4
Hap5 Hap1 Hap6
Hap2
Hap3 8
Fig. 3
Fig.3 Relationship among 6 haplotypes observed in this study.
Sampled
location N Observed No. of Haplotype Haplotype diversity diversity π (% )Nucleotide
Gabusoka (2016) 24 1 0.000 0.000 Gabusoka (2011) 7 1 0.000 0.000 Higashiyabe 22 3 0.178 0.612 Hanejioh 24 2 0.489 1.529 Okuma 14 1 0.000 0.000 Manna 7 1 0.000 0.000 Maeda 30 1 0.000 0.000 Tengan 23 1 0.000 0.000 Ohyama 24 3 0.522 0.598 Nishizaki 58 1 0.000 0.000 Kochinda 3 2 0.667 2.083 Gushikami 34 3 0.308 1.000 Tokashiki 4 1 0.000 0.000
Table 5 Estimated genetic diversity obtained from the variations observed in mtDNA D-loop region.
Gabusoka
(2016) Gabusoka (2011) Higashiyabe Hanejioh Okuma Manna Maeda Tengan Ohyama Nishizaki Kochinda Gushikami Tokashiki
Gabusoka (2016) ― *** *** *** *** *** *** *** *** * *** Gabusoka (2011) 0.000 ― *** ** *** *** *** *** *** ** Higashiyabe 0.903 0.849 ― *** *** *** *** * *** Hanejioh 0.609 0.478 0.188 ― *** *** *** *** *** *** *** Okuma 0.000 0.000 0.877 0.544 ― *** *** *** *** *** * *** Manna 1.000 1.000 0.872 0.706 1.000 ― *** *** *** *** *** ** Maeda 1.000 1.000 0.039 0.380 1.000 1.000 ― *** *** *** Tengan 1.000 1.000 0.916 0.793 1.000 0.000 1.000 ― *** *** *** Ohyama 0.899 0.848 0.040 0.207 0.874 0.873 0.205 0.915 ― *** *** Nishizaki 1.000 1.000 0.086 0.493 1.000 1.000 0.000 1.000 0.298 ― Kochinda 0.891 0.708 0.104 -0.225 0.828 0.849 0.706 0.946 0.141 0.826 ― *** Gushikami 0.097 0.013 0.682 0.341 0.063 0.784 0.804 0.839 0.683 0.854 0.363 ― *** Tokashiki 1.000 1.000 -0.122 0.152 1.000 1.000 0.000 1.000 1.000 0.000 0.111 0.693 ―
Table 6 Estimated genetic differences ( φ ST) among sampled locations.
個体数が少ない(N ≦ 4)集団のφ ST 値の多くは統 計的に有意ではない(p > 0.05)場合が多かった。また、 各集団中で検出されるハプロタイプ数が少ない(H ≦ 3)ため、集団内で確認されるハプロタイプが異なる と高い遺伝的分化を示す傾向にあった。 考 察 観賞魚系統のグッピーの遺伝的起源 観賞魚系統 14 系統からは Hap1 と 4 が確認され、 エンドラーズのみで Hap4 が観察された。系統解析で は、Hap1 は West Trinidad と、Hap4 は East Venezuela と遺伝的に近縁であることが明らかになった。 観賞魚系統は全て単一のハプロタイプを示し、異な るハプロタイプの混成は見られなかった。理由として、 起源がごく限られた地域であったこと、おそらくは グッピーの発見時に West Trinidad から持ち帰られた個 体が観賞魚としてのグッピーの起源となったためと考 えられる。グッピーの本来の生息域はベネズエラやト リニダード・トバゴを含む南米北東部である(Rosen と Bailey, 1963)。観賞魚系統のグッピーは 1960 年代 後半までアメリカやドイツを中心に生産されていた が、1970 年代以降はシンガポールに生産の中心が移っ た。アメリカやドイツで作られた系統が元となりシン ガポールにおいて生産されるようになった。そして、 シンガポールが観賞魚系統のグッピーの世界的な産地 となり、日本を含む世界中にシンガポールで養殖した グッピーを輸出している(Cheong, 1996)ことから単 一起源のグッピーが観賞魚として世界中に広まったも のと考えられる。観賞魚系統のグッピーには様々な色 彩・体型・体模様の系統が存在する。このような特定 の形質を有する系統の作出には、選抜による遺伝的改 良が必要となる。特定の形質を有する少数の親魚を繰 り返し行うことにより特定の形質をそろえていくこと になる。グッピーの系統作成の過程でも同様の過程を 経ているものと考えられ、そのために、系統内が単一 のハプロタイプとなったと考えられる。 Endrer’s は元々 Venezuela の一部地域にのみ生息して おり、他地域の野生型グッピーと比較して、体の模様 や体型など形態的特徴のほか、部分的な生殖的隔離が 認められていることから、Poecilia wingei と呼ばれ、 グッピーの亜種とされた(Poester ら , 2005)。mtDNA の cytochrome b 領域(953bp)を用いた North-eastern Venezuela 集団の解析では、P. wingei を含む集団は同 じ水系内の集団との遺伝的差異が認められたという (Herdegen ら , 2014)。系統解析に用いた East Venezuela
に由来するハプロタイプ CMARG69 と WC78 は先行 研究(Alexander と Breden, 2004)においてP. wingei と され別種とされている。本研究の結果では系統解析に おいて他の East Venezuela 起源のハプロタイプと近い 関係を示した。P. wingei が別種となるかどうかに関し てはさらに長い領域の比較や核 DNA を用いた解析が 必要と考えられる。 2.海外野生化集団の遺伝的起源 海外野生化集団は 3 集団全個体が Hap1 を示し、系 統解析では West Trinidad と遺伝的に近縁であることが 明らかになった。タイ、シンガポールはグッピーの本 来の生息地ではなく、蚊の生物学的防除などの理由で グッピーの導入報告がある地域である(Deacon ら , 2011; FAO, 2017; Herre, 1940)。アユタヤ、WS は集団 内で単一のハプロタイプ(Hap1)しか検出されず、 遺伝的起源は観賞魚と同様に West Trinidad であると考 えられる。導入の経緯などを考慮すると、東南アジア に導入されたグッピーはグッピー飼育が一般的になっ た初期の観賞魚起源の個体が用いられたと考えられ る。一方、Venezuela の野生化個体は他の地域の野生 化個体と同様に Hap1 であった。Venezuela をはじめと する原産地域では観賞魚由来の野良グッピーが問題と なっている。分析に用いた Venezuela が野良グッピー であり、Venezuela においても観賞魚由来の野良グッ ピーが蔓延していることを示している。 3.日本野生化集団の遺伝的起源 日本野生化集団は 14 集団で Hap1、2、3、5、6 の 5 つ の ハ プ ロ タ イ プ が 確 認 さ れ た。 系 統 解 析 で は、 Hap1、6 は West Trinidad、Hap2 は North Guyana、Hap3 は East Trinidad、Hap5 は Suriname・East Guyana と遺 伝的に近縁であることが明らかになった。 日本における野生化グッピーの導入歴は 1960 年代 に観賞用魚の野外遺棄とされており(石川ら , 2013)、 その後、沖縄地方の河川や日本各地の温泉地などで野 生化したグッピーがみられるようになった(山岸ら , 1962)。しかし、日本野生化集団では複数のハプロタ イプが確認され、14 集団中 5 集団でハプロタイプの 混成が見られた。今回確認されたハプロタイプのうち、 Hap1、2、4、6 は既報のハプロタイプと一致したが、 Hap3、5 は未報告のハプロタイプであった。一般的に、 魚 類 の mtDNA の 塩 基 置 換 率 は お よ そ 100 万 年 に 1~2% とされている(Near ら , 2003)ことから、導入 以前から元々遺伝的に異なっていた可能性が考えられ
る。また、いくつかの集団で見られたハプロタイプの 混成は、集団への複数回の導入が行われたためと考え られる。 系統解析により、各ハプロタイプの遺伝的起源を調 べてみると、観賞魚系統でも見られた Hap1 は West Trinidad を遺伝的起源としたグループに含まれ、Hap2 は North Guyana のグループに含まれた。Hap2 と同ク レード内の AU1 は、オーストラリアで確認されたハ プロタイプである。オーストラリア導入集団を対象に mtDNA を用いて遺伝的起源を調査した先行研究で、 AU1 は North Guyana を遺伝的起源とすることが明ら かにされ、これは、蚊の生物的防除を目的とした North Guyana からオーストラリアへの導入の報告と一 致していた(Lindholm ら , 2005)。ほかにも、日本野 生 化 集 団 で の み 確 認 さ れ た Hap3 は East Trinidad、 Hap5 は Suriname・East Guyana、Hap6 は West Trinidad のグループと近い関係を示した。 公式の記録では沖縄県で蚊の生物的防除を目的とし たグッピーの導入は行われていない。そのため、グッ ピーの導入経路は観賞用飼育個体の野外遺棄が主と考 えられていたが(石川ら , 2013)、本研究により、観 賞魚以外の系統が、経緯は不明であるが、複数回の導 入が行われたことが明らかとなった。しかし、今回の 分析ではブートストラップ値から支持されない分岐が いくつか見られている。これは今回分析に用いた領域 が 320bp と短かったことによると考えられる。正確性 を高めるためには今後より長い領域や他の領域を含め た解析を行う必要がある。 4.mtDNA の Dloop 領域に基づく遺伝的多様性 mtDNA の Dloop 領域の各集団における遺伝的多様 性 の 指 標 は、 ハ プ ロ タ イ プ が 3 つ 確 認 で き た Gushikami、Higashiyabe、Ooyama は Hd(ハプロタイ プ多様度)が 0.178 ~ 0.522、π(塩基多様度)が 0.598 ~ 1.000% となった。ハプロタイプが 2 つ確認できた Hanejioh、Kochinda は Hd が 0.489 ~ 0.667、πは 1.529 ~ 2.083% となった。残りの集団は単型で、遺伝的多 様性は Hd、πともに 0 だった。 ベネズエラやトリニダードを含む南米野生集団での mtDNA の Dloop 領域の遺伝的多様性は Hd が 0.783 ~ 0.986、 π が 0.003 ~ 0.015 で あ る(Alexander ら , 2006)。南米野生集団と日本野生化集団の遺伝的多様 性を比較すると、日本野生化集団は Hd が低い値を示 したのに対し、πは非常に高い値を示した。Hd が低 い値を示した理由として、日本へグッピーが導入され る際に生じたボトルネックと創始者効果によって、定 着の際にハプロタイプが減少したことが挙げられる。 導入によるハプロタイプの減少は、オーストラリアの グッピー導入集団でも確認されている(Lindholm ら , 2005)。 ま た、 ヨ ー ロ ッ パ の カ ダ ヤ シ(Gambusia holbrokki)導入集団(Vidal ら , 2010)やアメリカのコ イ 科 魚 類 Sacramento pikeminnow(Ptychocheilus grandis)導入集団(Kinziger ら , 2014)でも同様の現 象が確認されるなど、多くの外来種で導入時のボトル ネックと創始者効果によるハプロタイプの減少が知ら れている。πが高い値を示した理由として、ハプロタ イプの遺伝的起源の違いが関係している。πはハプロ タイプ頻度に遺伝的距離が反映されて算出されるが、 本研究の日本野生化集団で見られた各ハプロタイプは 遺伝的起源が異なる地域に由来することが示された。 同一地域内で確認されたハプロタイプで算出した先行 研究(Alexander ら , 2006)に対し、異なる地域に由来 するハプロタイプを含む日本野生化集団のほうが、遺 伝的距離は高くなるため、日本野生化集団のほうが高 い値を示したと考えられる。 中城湾における流れ藻の魚類相調査でグッピーが採 集されており、台風や大雨で一時的に海に流された後、 周辺河川への遡上による分布域を拡大している可能性 が指摘されている(立原ら , 2002)。今後、遺伝子流 動が人為的なものか、自然によるものなのかを明らか にするため、水系内集団や河口の近い河川集団のサン プルを増やすことやマイクロサテライトなどの両性遺 伝マーカーを用いることが、より詳細な遺伝的構造の 把握が必要になると考えられる。 謝 辞 実験の期間中、グッピーの維持管理に尽力していた だいた中嶋登代子技官に厚く感謝の意を表します。ま た、沖縄県でのサンプル採集に関して助言いただいた 琉球大学理学部海洋自然科学科の立原一憲准教授、ア ユタヤ ( タイ ) のサンプルを提供いただいたラジャマ ンガラ工科大学アユタヤ校の T. Ngamsiri 博士、シン ガポールのサンプルを提供いただいたシンガポールポ リテクニックの A. F. Yuen 博士に厚く御礼申し上げま す。 引用文献
Alexander HJ, Breden F. 2004. Sexual isolation and extreme morphological divergence in the Cumana guppy: a possible case of incipient speciation. Journal of
evolutionary biology, 17: 1238-1254.
Alexander HJ, Taylor JS, Wu SST, Breden F. 2006. Parallel evolution and vicariance in the guppy (Poecilia
reticulata) over multiple spatial and temporal scales.
Evolution, 60: 2352-2369.
青木大輔 , 中山祐一郎 , 林正人 , 岩崎魚成 . 2006. 琵琶 湖 に お け る オ オ ク チ バ ス フ ロ リ ダ 半 島 産 亜 種 (Micropterus salmoides floridanus) のミトコンドリア DNA 調節領域の多様性と導入起源 . 保全生態学研 究 , 11, 53-60.
Borthwick T. 1923. An Anti-mosquito campaign in Adelaide. Health, 1: 259-265.
Carvalho GR, Shaw PW, Magurran AE, Seghers BH. 1991. Marked genetic divergence revealed by allozymes among populations of the guppy Poecilia reticulata (Poeciliidae), in Trinidad. Biological Journal of the Linnean ociety, 42: 389-405.
Cheong L. 1996. Overview of the current international trade in ornamental fish, with special reference to Singapore. Revue scientifique et technique (International Office of Epizootics), 15: 445-481.
Clement M, Posada DCKA, Crandall KA. 2000. TCS: a computer program to estimate gene genealogies. Molecular ecology, 9: 1657-1659.
Deacon AE, Ramnarine IW, Magurran AE. 2011. How reproductive ecology contributes to the spread of a globally invasive fish. PLoS One, 6, e24416.
Edgar RC. 2004. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic acids research, 32: 1792-1797.
Excoffier L, Lischer HE. 2010. Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Molecular ecology resources, 10: 564-567.
Excoffier, L., Smouse, P. E., & Quattro, J. M. (1992). Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics, 131(2), 479-491.
Fajen A, Breden F. 1992. Mitochondrial DNA sequence variation among natural populations of the Trinidad guppy, Poecilia reticulata. Evolution, 46: 1457-1465. FAO(2017) DIAS - Database on Introductions of Aquatic
Species
http://www.fao.org/fishery/dias/en
Ghosh SK, Tiwari SN, SatHigashiyabeanarayan TS, Sampath TRR, Sharma VP, Nanda N, Subbarao SK. 2005. Larvivorous fish in wells target the malaria vector sibling species of the Anopheles culicifacies complex in villages in Karnataka, India.
Transactions of the ROhyamaal Society of Tropical Medicine and Higashiyabegiene, 99: 101-105.
Herdegen M, Alexander HJ, Babik W, Mavárez J, Breden F, Radwan J. 2014. Population structure of guppies in north-eastern Venezuela, the area of putative incipient speciation. BMC evolutionary biology, 14: 28.
Herre AWCT. 1940. Additions to the fish fauna of Malaya and notes on rare or little known Malayan and Bornean fishes. Bulletin of the Raffles Museum, 16: 27-61. Houde AE. 1997. Sex, color, and mate choice in guppies.
Princeton University Press. Princeton
石川哲郎 , 高田未来美 , 徳永圭史 , 立原一憲 . 2013. 沖 縄島に導入された外来純淡水魚類の定着状況およ び分布パターン . 保全生態学研究 , 8, 5-18. 岩崎登 . 1997. ブリーディング・テクニック グッピー 専科 , マリン企画 , 東京 . 岩松鷹司 . 1993. メダカ学 , サイエンティスト社 , 東京 . Kanda N, Nakajima M, FujiOhyama. 1991. Strain
differences at thermal resistance in the guppy, Poecilia
reticulata. Tohoku journal of agricultural research, 42:
25-31.
河村功一 , 米倉竜次 , & 石川正樹 . 2004. ミトコンドリ ア DNA の制限酵素切断多型から見た日本・韓国 産ブルーギルの遺伝的特徴 . 水産育種 , 33: 93-100. Kinziger, A. P, Nakamoto RJ, Harvey BC. 2014. Local-scale
invasion pathways and small founder numbers in introduced Sacramento pikeminnow (Ptychocheilus
grandis). Conservation genetics, 15: 1-9.
北川えみ , 北川忠生 , 能宗斉正 , 吉谷圭介 , 細谷和海 . 2005. オオクチバスフロリダ半島産亜種由来遺伝 子 の 池 原 貯 水 池 に お け る 増 加 と 他 湖 沼 へ の 拡 散 . 日本水産学会誌 , 71: 146-150.
Kocher TD, Thomas WK, Meyer A, Edwards SV, Pääbo S, Villablanca FX, Wilson AC. 1989. Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proceedings of the National Academy of Sciences, 86: 6196-6200. 幸地良仁 . 1991. トーイユからリュウキュウアユまで
とっておきの話 . 沖縄の川魚 . 沖縄出版 . 那覇 Lindholm AK, Breden F, Alexander HJ, Chan WK, Thakurta
SG, Brooks R. 2005. Invasion success and genetic diversity of introduced populations of guppies Poecilia
reticulata in Australia. Molecular Ecology, 14:
3671-3682.
McKay RJ. 1984. Introductions of exotic fishes in Australia. Distribution, biology, and management of exotic fishes. Johns Hopkins University Press, Baltimore, Maryland, pp. 177-199.
Near TJ, Kassler TW, Koppelman JB, Dillman CB, Philipp DP. 2003. Speciation in North American black basses, Micropterus (Actinopterygii: Centrarchidae). Evolution, 57: 1610-1621.
Nei M. 1973. Analysis of gene diversity in subdivided populations. Proceedings of the National Academy of Sciences, 70: 3321-3323.
Nei M, Tajima F. 1981. DNA polymorphism detectable by restriction endonucleases. Genetics, 97: 145-163. Patrick R. 1987. A history of health & medicine in
Queensland, 1824-1960. University of Queensland Press. Brisbane. Queensland.
Poeser FN, Kempkes M, Isbrücker IJ. 2005. Description of
Poecilia (Acanthophacelus) wingei n. sp. from the Paría
P e n i n s u l a , Ve n e z u e l a , i n c l u d i n g n o t e s o n
Acanthophacelus Eigenmann, 1907 and other subgenera
of Poecilia Bloch and Schneider, 1801 (Teleostei, Cyprinodontiformes, Poeciliidae). Contributions to Zoology, 74: 97-115.
Rosen DE, Bailey RM. 1963. The poeciliid fishes ( C y p r i n i d o n t i f o r m e s ) , t h e i r s t r u c t u r e , zoogeograpHigashiyabe and systematics. Bull. Am. Mus. Nat. Hist. N.Y. 126: 1-176.
Saitou N, Nei M. 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular biology and evolution, 4: 406-425.
Shields GF, Kocher TD. 1991. Phylogenetic relationships of North American ursids based on analysis of mitochondrial DNA. Evolution, 45: 218-221.
鹿野隆人 , 藤尾芳久 . 1994. グッピーにおける塩分耐性 の系統差について . 水産育種 . 20: 47-53. 嶋津信彦 . 2011. 2010 年夏沖縄島 300 水系における外 来水生生物と在来魚の分布記録 . 保全生態学研究 , 16: 99-110. 立原一憲 , 徳永桂史 , 地村佳純 . 2002 沖縄県の外来魚 類 .( 日 本 生 態 学 会 編 ) 外 来 種 ハ ン ド ブ ッ ク . pp248 - 249. 地人書館 , 東京 . 高村健二 . 2005. 日本産ブラックバスにおけるミトコ ンドリア DNA ハプロタイプの分布 . 魚類学雑誌 , 52: 107-114.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular biology and evolution, 30: 2725-2729.
Taylor JS, Breden F. 2000. Slipped-strand mispairing at noncontiguous repeats in Poecilia reticulata: a model for minisatellite birth. Genetics, 155: 1313-1320. Templeton AR, Crandall KA, Sing CF. 1992. A cladistic
analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics, 132: 619-633.
Vidal O, García-Berthou E, Tedesco PA, García-Marín JL. 2010. Origin and genetic diversity of mosquitofish (Gambusia holbrooki) introduced to Europe. Biological Invasions, 12: 841-851.
Vipan JAM. 1910. Malaria and the ‘millions’ fish (Girardinus poeciloides). In Proceedings of the Zoological Society of London, 146-147.
Vitousek PMAEDA'antonio CM, Loope LL, Rejmanek M, Westbrooks R. 1997. Introduced species: a significant component of human-caused global change. New Zealand Journal of Ecology, 21: 1-16.
山岸宏 , 中村譲 , 和田芳武 , 沖野外輝夫 , 中本信忠 , 落 合照雄 . 1962. 上山田・戸倉温泉地域に棲息する グッピーについて . 衛生動物 , 17: 48-58.
Yokogawa K. 1998. Morphological variabilities and genetic structures of largemouth bass Micropterus salmoides in Japanese fresh waters. Aquaculture Science, 46: 321-332.
鷲谷いずみ , 矢原徹一 . 1996. 保全生態学入門-遺伝子 から景観まで- , 文一総合出版 . 東京