コウチュウ目ハムシ科における?節味覚感覚子の普
遍性と寄主選択における役割に関する研究
著者
与謝野 舜
学位授与機関
Tohoku University
学位授与番号
11301
URL
http://hdl.handle.net/10097/00129323
コウチュウ目ハムシ科における
跗節味覚感覚子の普遍性と
寄主選択における役割に関する研究
コウチュウ目ハムシ科における
跗節味覚感覚子の普遍性と
寄主選択における役割に関する研究
専 攻 応用生命科学 専攻 指導教員 堀 雅敏 准教授 学籍番号 B7AD1210 氏 名 与謝野 舜目次
第 1 章 序論 1 第 2 章 コウチュウ目ハムシ科における跗節味覚感覚子の普遍性 5 第 1 節 外部形態観察によるハムシ科における跗節上の味覚感覚子の有無の調査 5 第 2 節 コウチュウ目ハムシ科の跗節味覚感覚子の内部形態 14 第 3 節 コウチュウ目における跗節味覚感覚子の普遍性の調査 26 第 4 節 考察 29 第 3 章 ハムシ科における跗節味覚感覚子を介した味受容 31 第 1 節 イチゴハムシの跗節味覚感覚子による塩受容 31 第 2 節 イチゴハムシの跗節味覚感覚子による糖受容 40 第 3 節 イチゴハムシの跗節味覚感覚子による苦味物質受容 43 第 4 節 イチゴハムシの跗節味覚感覚子による寄主植物葉表面ワックス受容 47 第 5 節 コガタルリハムシの跗節味覚感覚子による塩受容 51 第 6 節 考察 56 第 4 章 ハムシ科昆虫の跗節による味認識の役割 58 第 1 節 イチゴハムシのスクロース認識における跗節の重要性 58 第 2 節 イチゴハムシのブルシン認識における跗節の重要性 64 第 3 節 ヒメカメノコハムシのスクロース認識における跗節の重要性 68 第 4 節 オオニジュウヤホシテントウのスクロース認識における跗節の役割 71 第 5 節 イチゴハムシの寄主葉面ワックスの認識における跗節の重要性 75 第 6 節 イチゴハムシの寄主植物葉表面滲出物の認識における跗節の重要性 80 第 7 節 イチゴハムシの跗節による寄主の識別 84 第 8 節 考察 87 第 5 章 総合考察 89 摘要 93 謝辞 95 引用文献 961
第 1 章 序論
自然界に存在する化学物質は生物が周囲の環境を認識するために重要であり、化学物 質を受容する器官は、微生物から脊椎動物まで、全ての生物に備わっているといえる。 昆虫も例外ではなく、体外の化学物質を受容する器官を持ち、体表面に存在する感覚子 と呼ばれる器官がそれに相当する。受容された味物質は嗅覚または味覚として認識され、 昆虫の行動決定において重要な役割を果たす。昆虫ではこのような化学受容器は口器の 外側、触角、脚の先端節である跗節、交尾器、翅に存在していることが知られており、 特に味覚受容器は口器と触角、跗節、交尾器、翅に、嗅覚受容器は触角や口器に存在す る(Bruyne and Warr, 2005; Yarmolinsky et al., 2009)。昆虫の味覚受容器と嗅覚受容器 はそれぞれ味覚感覚子、嗅覚感覚子と呼ばれ、それぞれ異なる構造をもつ。味覚感覚子 は単孔性で先端に 1 つだけ孔(感覚子孔)をもつのに対し、嗅覚感覚子は多孔性の感覚 子で、先端および壁面に多数の感覚子孔をもつ(Mitchell et al., 1999)。これらの化学受 容感覚子は中空で、内腔は感覚子液と呼ばれる液体で満たされており、感覚子基部に存 在する複数の化学受容神経細胞の樹状突起が感覚子の開口部付近まで伸びている。感覚 子孔から取り込まれた化学物質は感覚子液に溶解し、樹状突起で受容されることで認識 される。嗅覚感覚子が受容するのは主に空気中に漂う物質であり、嗅覚感覚子自体が匂 い源の基質に直接触れる必要がないのに対し、味覚感覚子は主に物体と接触することに よって取り込まれた化学物質を受容する。 植食性昆虫の寄主選択において、嗅覚は寄主の探索に、味覚は寄主到達後の餌や産卵 場所としての適性の評価にそれぞれ用いられ、寄主選択のステップにおいて各感覚の重 要性が変化する(石川, 2007)。また、味覚は昆虫の摂食行動そのものにも大きく関わっ ており、摂食の可否の判定、口器(大顎や吻、口鬚など)を用いた摂食、摂食の開始に ともなう移動の抑制のすべてに味覚が関わっている(Thoma et al., 2017)。口器(口鬚、 口吻)、触角、跗節はすべて味覚器官だが、それぞれの味覚器官が果たす役割が異なる こともあるため、昆虫の行動決定における役割については味覚器官ごとに調べられてい る(Pontes et al., 2014; Thoma et al., 2017)。特に口器による味認識は摂食行動と密接 に関わっており、他の味覚器官と比較しても多くの知見が蓄積されている(Stocker, 1994; Murata et al., 2017)。 跗節による味認識についてはハエ目、チョウ目で特に知見が蓄積されており、形態学 的、電気生理学的、行動学的手法を用いた解析が多く行われている。ハエ目における跗 節による味認識に関する電気生理学的手法はクロバエ科の Phormia regina において確 立され(McCutchan, 1969)、この種やショウジョウバエにおいて跗節による味認識に つ い て 多 く の研 究 が 行わ れ て き た 。特 に 、 キイ ロ シ ョ ウ ジョ ウ バエ Drosophila melanogasterはモデルとしてよく用いられ、跗節で糖や苦味物質を認識すること、寄主2
(餌)の探索において跗節が重要な役割を果たしていることが明らかにされている (Stocker, 1994; Thoma et al., 2016)。また、キャベツハナバエDelia radicumでは、寄 主に含まれるグルコシノレート類を跗節上の化学感覚子で受容し、それを認識すること が産卵行動において重要であることが明らかにされている(Roessingh et al., 1992)。ま た、ショウジョウバエでは寄主探索に加えて配偶行動においても跗節による味認識が重 要であることが明らかにされており(Miyamoto and Amrein, 2008; Inoshita et al., 2011)、 ハエ目において跗節による味認識が生存上重要なさまざまな行動に大きく関与してい ることが分かっている。チョウ目では形態観察により跗節上の味覚感覚子の有無が複数 の科にわたって調査されており、シャクガ科(Geometridae)のウスジロエダシャク
Ectropis obliqua(Ma et al., 2016)、メイガ科(Pyralidae)の Homoeosoma nebulella
(Faucheux, 1991)、コナガ科(Plutellidae)のコナガPlutella xylostella(Qiu et al., 1998) タテハチョウ科(Nymphalidae)のアカタテハ Vanessa indica やツマグロヒョウモン
Argyreus hyperbius(Ômura et al., 2011)などで跗節味覚感覚子の存在が確認されてい ることから、跗節味覚感覚子はチョウ目に共通した特徴であると考えられる。また、ハ エ目と同様、跗節が寄主選択において味覚器官として用いられていることも明らかにさ れており、例えばオオモンシロチョウPieris Brassicae は跗節上に味覚感覚子をもち、 跗節による味認識が産卵行動において重要な役割を果た すことが報告されている (Chun and Schoonhoven, 1973; Renwick and Chew, 1994)。また、Helicoverpa assulta
では跗節で糖やアミノ酸を受容でき、それによって寄主の質を確認できることが示唆さ れている(Zhan, et al., 2011)。これらのことから、チョウ目においても跗節による味認 識が重要な役割を果たしていると考えられる。これらの目に加えて、ハチ目ではセイヨ ウ ミ ツ バ チ Apis mellifera が 跗 節 味 覚 感 覚 子 を も ち 、 跗 節 で 糖 を 認 識 す る こ と (Whitehead and Larsen, 1976; de Brito Sanchez et al., 2008 2014; de Brito Sanchez, 2011)、バッタ目ではトノサマバッタLocusta migratoriaの跗節上に味覚感覚子が存在 すること(Zhou et al., 2009)、サバクトビバッタSchistocerca gregariaが跗節上の味覚 感覚子で塩や糖、匂い物質を受容でき、跗節味覚感覚子による化学受容が忌避応答に関 わっていること(Newland, 1998)が明らかにされている。
これらの形態学的、電気生理学的、行動学的知見に加え、近年では味受容神経 ( Gustatory receptor neurons: GRNs ) 上 で 発 現 す る 味 物 質 の 受 容 体 ( Gustatory receptors: GRs)について分子生物学的な知見も蓄積されており、各受容体の発現部位 や役割が調査されてきている(Thorne et al., 2004)。例えば、チョウ目のナミアゲハで は跗節で発現している Sf9 という受容体が産卵刺激物質であるシネフリンの認識に必 要であることが明らかにされているほか(Ozaki et al., 2011)、ヒメアカタテハVanessa carduiにおいて跗節上で発現している GRs の多様性が調査されている(Suzuki, et al., 2018)。特にショウジョウバエでは Gal4-UAS システムを用いた GRs のノックアウト
3
個体が利用可能なため(Ishimoto and Tanimura, 2004)、跗節で発現する GRs の役割に 関する知見が蓄積されてきている(Miyamoto and Amrein, 2008; Thoma et al., 2016)。
一方、コウチュウ目における跗節による味認識に関する知見は非常に少ない。コロラ ドハムシChrysolina brunsvicensisにおいて KCl や NaCl といった塩や寄主植物に含ま れる Hypericine が跗節上の味覚感覚子によって受容されることが明らかにされている ほか(Rees, 1969)、ヨツモンマメゾウムシCallosobruchus maculatusにおいて跗節の有 無が産卵行動に影響することが明らかにされている(Messina et al., 1987)ものの、コ ウチュウ目において跗節による味認識がどのような役割を果たしているかはほとんど 明らかになっていない。また、跗節上の味覚感覚子の存在を形態学的に調査した例も少 なく、ハムシ科のマメゾウムシ亜科の 2 種(Mbata et al., 1997)やハネカクシ亜科(Betz, 2003)で跗節上の味覚感覚子の有無が調査されているものの、コウチュウ目において跗 節上の味覚感覚子が共通した特徴であるかも不明である。また、コウチュウ目ではコク ヌストモドキTribolium castaneumの跗節の GRNs 上での GRs の発現を調査した例が あるものの(Adbel-latief, 2007)、脚での GRs の発現について調査した報告は他にはほ とんどなく、それを利用した味覚器官としての跗節の役割の調査はいまだ行われていな いのが現状である。 コウチュウ目には昆虫の約 4 割が属するとされ、37 万種以上が記載される非常に大 きな分類群である(Gullan and Cranston, 2014)。このことから、昆虫一般の味認識メ カニズムの解明を行う上でコウチュウ目の味認識に関する知見を蓄積することは必須 であるといえる。また、コウチュウ目は多くの植食性昆虫を含み、植食生昆虫の 50% 以上がこの目に含まれる(Farrel, 1998)。そのため、コウチュウ目の味認識について理 解することは植食性昆虫の寄主選択メカニズムの解明にもつながると考えられる。跗節 は特に飛翔を行う昆虫が目標となる物体(植物など)に着地した際に最初に物体に触れ る器官であり、跗節による味認識の有無や、その役割を明らかにすることは昆虫の寄主 選択メカニズムの解明において必須であると考える。 コウチュウ目の中で、ハムシ上科はゾウムシ上科とともに最も植食性に特化した分類 群であり、この 2 上科で 135000 種以上が記載され(Lawrence, 1982; 森本,1986)、コ ウチュウ目の植食性種の 8 割以上がこの 2 上科に分類される(Farrel, 1998)。ハムシ上 科はカミキリムシ科とハムシ科からなるが、ハムシ科は植食性に特化していることから 多くの農業害虫を含む。例えば、コロラドハムシLeptinotarsa decemlineataはアメリカ 大陸を中心に北半球に分布し、ジャガイモの主要な害虫種としてあげられる(Palli, 2014 )。 キ ス ジ ノ ミ ハ ム シ Phyllotreta striolata は ア ブ ラ ナ 科 の 重 要 害 虫 で あ り (Andersen et al., 2006)、幼虫が根部を食害するためダイコンでは特に重大な問題とな っている。また、ハムシ科害虫でも化学殺虫剤に対する抵抗性が発達してきていること から(Nauen and Denholm, 2005; Gassman et al., 2011)、殺虫剤とは異なるアプローチ
4 を用いた害虫防除技術が必要とされてきており、ハムシ科の寄主選択メカニズムを解明 することはハムシ科害虫に対する新たな防除法を検討するうえでも重要だと考えられ る。ハムシ科の寄主選択において跗節による味認識が果たす役割はこれまで不明であっ たが、跗節による味認識を利用した害虫防除法が確立されれば、殺虫剤使用の削減にも つながり、より環境負荷の少ない害虫防除が可能になることも考えられる。以上のこと から、本研究ではハムシ科に注目し、形態学的手法を用いて跗節上の化学受容器の有無 を網羅的に調査した。加えて、電気生理学的なアプローチから跗節による味受容を確認 するとともに、行動学的調査を行い、ハムシ科において跗節による味認識が果たす役割 について研究を行った。
5 第 2 章
コウチュウ目ハムシ科における跗節味覚感覚子の普遍性
跗節上の化学感覚子の有無を明らかにするには、形態観察が有効な手法である。そこ で、コウチュウ目ハムシ科に属する種を中心として跗節の外部形態および内部形態を観 察し、化学感覚子の有無を調査した。 第 1 節 外部形態観察によるハムシ科における跗節上の味覚感覚子の有無の調査 昆虫の化学感覚子の分布と形態を調査するには、電子顕微鏡を用いた観察が有効であ る。昆虫の感覚子は機械感覚子と化学感覚子に大別され、孔の有無でそれらを識別でき る。化学感覚子には嗅覚感覚子と味覚感覚子があるが、嗅覚感覚子は表面に多数の孔を もち、味覚感覚子は先端に 1 つだけ孔をもつ(Zacharuk, 1980)ことから、外部形態か ら化学覚感覚子の種類も推定できる。そこで、跗節の外部形態を走査型電子顕微鏡 (Scanning electron microscope: SEM)を用いて観察し、全 17 亜科の各亜科に属する 種の跗節上に味覚感覚子と推定される感覚子が存在するか調査した。 1.材料および方法 供試虫 観察に供試した種を表 1 に示した。ハムシ科の各亜科から、少なくとも 1 種を観察に 供試し、17 亜科 49 種の跗節を観察した。 SEM 観察試料の作製 供試虫は冷凍殺虫し、跗節の汚れを取り除くため超音波洗浄機(SONO CLEANER、 株式会社カイジョー、東京)を用いて洗浄した。その後、跗節を傷つけないように脛節 と跗節を切り離した。作業は顕微鏡下で行い、跗節の切断には鋭利なピンセットを用い た。切断した跗節はアルミニウム基質の SEM 用カーボン両面テープ(日新 EM 株式会 社、東京)を用いてアルミニウム製の SEM 試料台(日新 EM 株式会社、東京)に固定 し、乾燥させた。その後、イオンスパッタ(MSP-1S、株式会社真空デバイス、茨城) を用いて白金パラジウムでコーティングし、観察試料とした。 SEM による跗節の観察 観察には走査型電子顕微鏡(SU8000Type II、株式会社日立製作所、東京)を用いた。 供試した種の跗節を観察し、化学感覚子と推定される感覚子(感覚子孔をもつ感覚子) の有無を調査した。6 2.結果 跗節の観察を行った全種で跗節上に先端孔をもつ感覚子の存在が確認され(図 1、 表 1)、それらは形態から味覚感覚子と推定された。各亜科で観察された跗節上の味覚 感覚子の例を図 2 および図 3 に示した。観察された跗節味覚感覚子には、先端孔の周囲 に明瞭な隆起または溝をもつものと先端孔の周囲に溝がみられないものがみられたた め、本研究ではそれぞれを Type I、Type II の感覚子とした。Type I の感覚子の例とし てイチゴハムシ、TypeII の感覚子の例としてカメノコハムシの跗節と跗節上で観察さ れた味覚感覚子を図 4、図 5 に示した。Type I の感覚子はカタビロハムシ亜科、マメゾ ウムシ亜科、コガネハムシ亜科、ネクイハムシ亜科、クビホソハムシ亜科、ナガハムシ 亜科、サルハムシ亜科、ヒゲナガハムシ亜科、ノミハムシ亜科、ハムシ亜科、ツヤハム シ亜科、ツツハムシ亜科、ナガツツハムシ亜科の 13 亜科だけで観察され(図 2、図 3)、 Type II の感覚子はホソハムシ亜科、カメノコハムシ亜科、トゲハムシ亜科、コブハム シ亜科の 4 亜科だけで観察された(図 2、図 3)。いずれの種においても、味覚感覚子の 数は機械感覚子の数よりも少なかった。また、味覚感覚子は跗節の左右に対称に位置し ている傾向がみられた。
7 表 1:跗節上の化学受容感覚子の有無を調査したハムシ科昆虫 亜科 種 学名 カタビロハムシ亜科 カタビロハムシ Colobaspis japonicus ワモンナガハムシ Zeugophora annulata マメゾウムシ亜科 アズキゾウムシ Callosobruchus chinensis コガネハムシ亜科 フェモラータオオモモブトハムシ Sagra femorata
ネクイハムシ亜科 エゾオオネクイハムシ Plateumaris constricticollis babai
ヒラタネクイハムシ Donacia hiurai クビホソハムシ亜科 ヤマイモハムシ Lema honorata アカクビナガハムシ Lilioceris subpolita ナガハムシ亜科 クロナガハムシ Orsodacne arakii ホソハムシ亜科 カバノキハムシ Syneta adamsi サルハムシ亜科 トビサルハムシ Trichochrysea japana マダラアラゲサルハムシ Demotina fasciculata アカガネサルハムシ Acrothinium gaschkevitchii チャイロサルハムシ Basilepta balyi ヒゲナガハムシ亜科 イタドリハムシ Gallerucida bifasciata キクビアオハムシ Agelasa nigriceps クロウリハムシ Aulacophora nigripennis クワハムシ Fleutiauxia armata サンゴジュハムシ Pyrrhalta humeralis イチゴハムシ Galerucella grisescens ネギオオアラメハムシ Galeruca extensa ウリハムシモドキ Atrachya menetriest ノミハムシ亜科 テントウノミハムシ Argopistes biplagiata タマアシトビハムシ Philopona vibex フタホシオオノミハムシ Pseudodera xanthospila スジカミナリハムシ Altica latericosta ハムシ亜科 クルミハムシ Gastrolina depressa コガタルリハムシ Gastrophysa altrocyanea ハッカハムシ Chrysolina exanthematica フジハムシ Gonioctena rubripennis ルリハムシ Linaeidea aenea カメノコハムシ亜科 ジンガサハムシ Aspidomorpha indica ヒメカメノコハムシ Cassida piperata アオカメノコハムシ Cassida rubiginosa ミドリカメノコハムシ Cassida erudite イチモンジカメノコハムシ Thlaspida cribrosa ルイスジンガサハムシ Thlaspida lewisii
8 カメノコハムシ Cassida nebulosi セモンジンガサハムシ Cassida versicolor トゲハムシ亜科 カタビロトゲハムシ Dactylispa subquadrata ヒメキベリトゲハムシ Dactylispa angulosa キムネクロナガハムシ Brontispa longissima ツヤハムシ亜科 ドウガネツヤハムシ Oomorphoides cupreatus コブハムシ亜科 ツツジコブハムシ Chlamisus Laticollis ツツハムシ亜科 キアシルリツツハムシ Cryptocephalus fortunatus バラルリツツハムシ Cryptocephalus approximatus クロボシツツハムシ Cryptocephalus signaticeps ナガツツハムシ亜科 キボシルリハムシ Smaragdina aurita ムナキルリハムシ Smaragdina semiaurantiaca
9
図 1:跗節上の味覚感覚子の有無を調査したハムシ科の各亜科の種数 観察した全ての種で跗節上に味覚感覚子の存在が確認された。 図は森本ら(1996)より。
10
図 2. ハムシ科の各亜科で観察された跗節味覚感覚子 1
A. カタビロトゲハムシDactylispa subquadrate(トゲハムシ亜科) B. キボシルリハムシSmaragdina aurita(ナガツツハムシ亜科) C. クロナガハムシOrsodacne arakii(ナガハムシ亜科)
D. エゾオオネクイハムシPlateumaris constricticollis babai(ネクイハムシ亜科) E. フタホシオオノミハムシPsedodera xanthospila(ノミハムシ亜科)
F. クルミハムシGastrolina depressa(ハムシ亜科)
G. イチゴハムシGalerucella grisescens(ヒゲナガハムシ亜科) H. カバノキハムシSyneta adamsi(ホソハムシ亜科)
11 図 3. ハムシ科の各亜科で観察された跗節味覚感覚子 2 A. カタビロハムシColobaspis japonica(カタビロハムシ亜科) B. カメノコハムシCassida nebulosa(カメノコハムシ亜科) C. アカクビナガハムシLilioceris subpolita(クビホソハムシ亜科) D. フェモラータオオモモブトハムシSagra femorata(コガネハムシ亜科) E. ツツジコブハムシChlamisus laticollis(コブハムシ亜科) F. マダラアラゲサルハムシDemotina fasciculata(サルハムシ亜科) G. バラルリツツハムシCryptocephalus approximatus(ツツハムシ亜科) H. ドウガネツヤハムシOomorphoides cupreatus(ツヤハムシ亜科)
12
図 4. イチゴハムシの跗節味覚感覚子の外部形態
A:イチゴハムシの跗節先端節、*:先端孔を備えた感覚子 B:イチゴハムシの跗節上の先端孔を備えた感覚子
13
図 5:カメノコハムシの跗節味覚感覚子の外部形態 A:カメノコハムシの跗節
B:カメノコハムシの跗節先端節、*:先端孔を備えた感覚子 C:カメノコハムシの跗節上の先端孔を備えた感覚子
14 第 2 節 コウチュウ目ハムシ科の跗節味覚感覚子の内部形態 外部形態観察でみられる感覚子孔の有無だけでは、その感覚子が化学感覚子であるか を判定するには不十分である。これは、観察で感覚子孔だと推定されたものが実際に跗 節内部まで続いているか SEM を用いた観察だけでは判断できないためである。一般に、 化学感覚子は中空で、その内部は跗節内部とつながっており、感覚子内腔には化学物質 を受容する神経が感覚子基部から先端孔付近まで伸びている。外部形態から化学感覚子 と推定されたものがこれらの内部形態的特徴を備えていれば、外部形態観察の結果を裏 付けできる。そこで、ハムシ科で外部形態観察から跗節味覚感覚子であると推定された 感覚子について透過型電子顕微鏡(Transmission electron microscope: TEM)を用いて 内部形態観察を行い、味覚感覚子としての特徴を内部にも備えているか調査し、外部形 態観察による跗節味覚感覚子の有無の判定が妥当であるか確認した。 1.材料および方法 供試虫 外部形態観察から跗節味覚感覚子の存在が確認された 3 種(イチゴハムシ、カメノコ ハムシ、ヒメカメノコハムシ)を観察に供試した。 イチゴハムシ(図 6)は旧東北大学農学部構内(宮城県仙台市青葉区堤通雨宮町)で 採集したものを初代とし、24±1 ℃、明暗条件 16L: 8D の昆虫飼育槽内で累代飼育し たものを用いた。イチゴハムシは成育ステージごとにシャーレに分け、食草と水分を十 分に与えて飼育した。食草のエゾノギシギシRumex obtusifoliusは東北大学青葉山新キ ャンパス構内(宮城県仙台市青葉区荒巻字青葉)、旧東北大学農学部構内(宮城県仙台 市青葉区雨宮)および宮城県仙台市青葉区内に自生するもの、または東北大学青葉山新 キャンパス内の温室で栽培したものを使用した。試料の作成には羽化後 3~8 日のイチ ゴハムシの成虫を雌雄の区別なく供試した。 カメノコハムシ(図 7A)は旧東北大学農学部構内で採集したものを初代とし、24 ± 1 ℃、明暗条件 16L: 8D の昆虫飼育槽内で飼育したものを用いた。飼育はシャーレ内で 行い、食草と水分を十分に与えて飼育した。食草にはアカザC. album var. centrorubrum
およびシロザC. albumを用いた。食草は旧東北大学農学部構内に自生するもの、また は東北大学農学部青葉山新キャンパス内の温室で栽培したものを用いた。試料の作成に は成虫を雌雄の区別なく供試した。 ヒメカメノコハムシ(図 7B)は旧東北大学農学部の圃場で採集したものを初代とし、 24 ± 1 ℃、明暗条件 16L: 8D の昆虫飼育槽内で飼育し、累代したものを用いた。飼育 はシャーレ内で行い、食草と水分を十分に与えて飼育した。食草にはアカザおよびシロ ザを用いた。食草は旧東北大学農学部構内に自生していたもの、または東北大学青葉山
15 新キャンパス内の温室で栽培したものを用いた。試料の作成には成虫を雌雄の区別なく 供試した。 TEM 観察試料の作製 供試虫の脚部を胸部から切り離し、エタノールで脱脂したカミソリを用いて 2.5%グ ルタルアルデヒド溶液中で爪の先端を切除したのち、跗節を脛節から切り離した。切り 離した各跗節は 2.5%グルタルアルデヒド溶液中に沈め、冷蔵庫で一晩固定した(タン パク質固定)。跗節が固定液中に沈まない場合は真空デシケーターを用いて跗節表面お よび内部にトラップされた気泡を除去し、跗節を固定液中に沈めた。その後リン酸緩衝 液で洗浄し、1%四酸化オスミウム溶液を用いて 2 時間固定した(脂質固定)。四酸化オ スミウムによる固定後、再びリン酸緩衝液で洗浄し、エタノール脱水系列による脱水を 行った。脱水後、溶媒を酸化プロピレンに置換した。酸化プロピレンとエポキシ樹脂 (Epon)の混合比率を変えながら溶媒を樹脂に置換した。樹脂置換後の各跗節は樹脂 に包埋し、定温乾燥機(DRN320DB、アドバンテック東洋株式会社、東京)を用いて樹 脂を重合させ、樹脂ブロック試料とした。観察に際しては、樹脂ブロック試料をウルト ラミクロトーム(ウルトラカット S、Leica Microsystems GmbH、Wetzlar、Germany) を用いて超薄切し、超薄切片としたものをシートメッシュ上に乗せ、観察した。 詳細な資料作成手順は表 2 に示した。
16
17
図 7:カメノコハムシとヒメカメノコハムシ A:カメノコハムシ
18 表 2:TEM 観察用の樹脂包埋試料の作成方法 試薬 作業手順 1 緩 衝 液 の 作製 ・ リン酸二 水素ナト リウム・2 水和物 ・ リン酸水 素二ナト リウム・ 12 水 和 物 1.1 リン酸二水素ナトリウム・2 水和物とリン酸水 素二ナトリウム・12 水和物をそれぞれ純水に溶 かし、0.2 M リン酸二水素ナトリウム水溶液お よび 0.2 M リン酸水素二ナトリウム水溶液を作 製する。 1.2 攪拌しながら 0.2 M リン酸二水素ナトリウム水 溶液と 0.2 M リン酸水素二ナトリウム水溶液を 混合し、リン酸緩衝液を作製する。混合する際 は pH メーターを使用し、pH が 7.0±0.2 にな るように調製する。 2 前 固 定 ( タ ン パ ク 質 固 定) ・ 25%グル タルアル デヒド溶 液 ・ 0.2 M リ ン酸緩衝 液(pH 7) 2.1 25%グルタルアルデヒド溶液をリン酸緩衝液 で希釈し、2.5%グルタルアルデヒド溶液(前固 定液、浸透速度 0.5 mm/h)とする。 2.2 イチゴハムシを生きたまま超音波洗浄機で洗 浄する(数秒程度)。 2.3 実体顕微鏡下でイチゴハムシの脚を基部から 抜き取り、前固定液中で爪の先端を切断する。 爪の切断にはエタノールで脱脂したカミソリ を用いる。 2.4 前固定液中で跗節を脚から切り離し、前固定液 の入った固定ビンに入れる。 2.5 固定ビンを真空デシケーターに入れ、数回脱気 を繰り返す。それでも沈まない場合は超音波洗 浄機で振動を与え、跗節が前固定液に沈むよう にする。跗節が沈まない場合はこれらの作業を 沈むまで繰り返す。 2.6 冷蔵庫で 1 晩固定する。 3 洗浄 ・ 0.2 M リ ン酸緩衝 液(pH 7) 3.1 跗節を洗浄液(リン酸緩衝液)に移し、10 分間 洗浄する。よく攪拌し、洗浄は 3 回行う。 4 後 固 定(脂 ・ 2% 四 酸 化オスミ ウム溶液 4.1 2%四酸化オスミウム溶液をリン酸緩衝液で希 釈し、1%四酸化オスミウム溶液(後固定液、浸 透速度 0.8 mm/h)とする。
19 質 固 定) ・ 0.2 M リ ン酸緩衝 液(pH 7) 4.2 洗浄後の跗節を後固定液に移し、室温で 2 時間 固定する。 5 洗浄 ・ 0.2 M リ ン酸緩衝 液(pH 7) 5.1 跗節を洗浄液(リン酸緩衝液)に移し、1 分間 洗浄する。 6 脱水 ・ 50%エタ ノール ・ 70%エタ ノール ・ 90%エタ ノール ・ 無水エタ ノール 6.1 エタノールの濃度を徐々に変えていき、試料の 脱水を行う。脱水は第 S1 表に示した順序で行 う。 第 S1 表 脱水系列 試薬 時間(分) 1 50%エタノール 10 2 70%エタノール 10 3 90%エタノール 15 4 99.9%エタノール 20×3 回 7 溶 媒 置換 ・ 酸化プロ ピレン 7.1 跗節を酸化プロピレンに移し、15 分間の溶媒置 換を 2 回行う。 ※固定ビンの蓋を占めると内圧が上がって勢 いよく開いてしまうため、蓋は上に乗せる程度 にする。 8 樹 脂 調合 ・ Quetol 812 ・ Araldite ・ DDSA ・ DMP-30 8.1 樹脂浸透用の樹脂(Epon)を調合する。第 S2 表にって調合を行う。調合を行う際にはマグネ ットスターラーを用いて攪拌する。 ※樹脂の攪拌が不十分だと試料の硬さが均一 にならず、きれいな超薄切片を作成できないた め、よく攪拌する。 第 S2 表 Quetol812+Araldite 混合樹脂(Epon) 配合割合 重量(g) Quetol 812 5.5 Araldite 4.5 DDSA 10.0 DMP-30 0.3 9 樹 脂 置換
・ Epon 9.1 Epon と酸化プロピレン(PO)の混合割合を変 えながら、徐々に樹脂へ置換する。Epon と PO
20 の混合にはマグネットスターラーや超音波洗 浄機を用い、よく混合する。樹脂置換は第 S3 表 の手順に従って行う。 第 S3 表 樹脂置換の手順 試薬 時間(時間) Epon : PO = 3 : 7 6 Epon : PO = 5 : 5 12 Epon : PO = 8 : 2 6 樹脂調合 Epon : PO = 10 : 0 12 Epon : PO = 10 : 0 6 樹脂調合 Epon : PO = 8 : 2 および 2 回目の Epon : PO = 10 : 0 の置換作業の直前に新しく Epon を調合 する。 10 包埋・ 重合 ・ Epon 10.1 包埋作業を行う直前に新しく Epon を調合す る。 10.2 シリコン型に樹脂(Epon)を流し入れ、脱気し て気泡を除去する。試料を樹脂内に設置する。 この際、樹脂置換用の古い樹脂が包埋用の樹脂 に混ざらないようにする。 10.3 第 S4 表にしたがって定温乾燥機(DRN320DB、 アドバンテック東洋株式会社、大阪)を用いて 温度を段階的に上げながら樹脂を重合させる。 ※定温乾燥機には同時にシリカゲルも入れて 除湿を行う。(湿度が高いと樹脂がうまく重合 しない。) 第 S4 表 重合時の温度管理 温度(℃) 時間(時間) 30 6 45 6 60 36 11 ト リ ミ ン グ・厚 切り 11.1 作製した樹脂ブロックの試料を含む部分を 0.5 mm 四方にトリミングする。(超音波カッター を用いると樹脂のゆがみが少ない。)
21 11.2 XAC ナイフを用いて準超薄切片(厚さ 2.0 µm) を作製。スライドグラスに切片を乗せ、伸展機 を用いて 60 ℃に加熱し切片を伸展させる。ト ルイジンブルー溶液で切片を染色し、試料が切 断されているか確認する。 12 超 薄 切 片 作成 12.1 ダイヤモンドナイフを用いて試料の超薄切を 行い、超薄切片(厚さ 80 nm)を作成する。 12.2 作製した超薄切片はメッシュ数 100 のグリッド に乗せる。
22 TEM による跗節内部形態の観察 観察には透過型電子顕微鏡(H-7650(ZeroA)、株式会社日立製作所、東京)を用いた。 四酸化オスミウムによる電子染色だけでは明瞭な画像が得られない場合には、クエン酸 鉛と Ti ブルーを用いた 2 重染色を行い、観察した。 2.結果 イチゴハムシの跗節上の感覚子の横断面を図 8 に示す。イチゴハムシでみられた Type I の感覚子では、感覚子内腔に神経鞘に包まれた 4 本の樹状突起外節が観察され た(図 8A)。感覚子内腔の樹状突起は先端部、基部の両方で観察された(図 8A―8C)。 また、感覚子基部では感覚子先端でみられた 4 本の樹状突起に加え、新たに 1 つの樹状 突起が観察された(図 8C)。跗節内部でも鞘細胞に囲まれた 5 本の神経細胞が観察され た(図 8D)。一方、外部形態から感覚子孔をもたないとされた感覚子では感覚子内腔に 神経細胞は観察されなかった(図 8E)。Type I の跗節内部で見られた神経細胞では、円 状の構造が 2 つ対になっているものが 9 対観察された(図 9)。 カメノコハムシ、ヒメカメノコハムシの跗節上の感覚子(Type II)の横断面を図 10 に示す。イチゴハムシと同様、カメノコハムシ亜科の 2 種で観察された味覚感覚子の内 腔にも神経鞘に包まれた 4 本の樹状突起外節が観察された。また、これらの 2 種におい ても、先端孔をもたない感覚子は内腔に神経を備えていなかった。
23 図 8:イチゴハムシの跗節上の感覚子の内部形態 A:跗節味覚感覚子先端部の横断面、ds:神経鞘、ods:樹状突起外節、cw:表皮 B:跗節味覚感覚子基部の横断面、ds:神経鞘、cw:表皮 C:跗節味覚感覚子のソケット部の断面、ds:神経鞘、cw:表皮 D:跗節内部にみられた神経群、矢印:神経細胞 E:跗節上の機械感覚子先端部の横断面
24
図 9:イチゴハムシの跗節内部
25 図 10:カメノコハムシ、ヒメカメノコハムシの跗節味覚感覚子の内部形態 A:カメノコハムシの跗節味覚感覚子先端部の横断面、ds:神経鞘、ods:樹状突起外 節、cw:表皮 B:ヒメカメノコハムシの跗節味覚感覚子先端部の横断面、ds:神経鞘、ods:樹状突 起外節、cw:表皮
26 第 3 節 コウチュウ目における跗節味覚感覚子の普遍性の調査 外部形態観察、内部形態観察からハムシ科において跗節上の味覚感覚子は共通した特 徴であることが示唆された。この特徴がハムシ科だけでなくコウチュウ目全体を通して 共通しているか明らかにするため、SEM および TEM を用いてコウチュウ目の他科に 属する種の跗節を観察し、味覚感覚子の有無を調査した。 1.材料および方法 供試虫 カブトムシ(コガネムシ科)、ナガヒラタムシ(ナガヒラタムシ科)、ヨツボシテント ウダマシ(テントウダマシ科)、オオニジュウヤホシテントウ(テントウムシ科)の跗 節を観察した。カブトムシ、ヨツボシテントウダマシは野外で採集した個体を用いた。 ナガヒラタムシ、オオニジュウヤホシテントウは飼育個体を用いた。 SEM 観察試料の作製 供試虫は冷凍殺虫し、跗節の汚れを取り除くため超音波洗浄機(SONO CLEANER) を用いて洗浄した。その後跗節を傷つけないように脛節と跗節を切り離した。作業は顕 微鏡下で行い、跗節の切断には鋭利なピンセットを用いた。切断した跗節はカーボンテ ープを用いて SEM 試料台に固定し、乾燥させた。その後、イオンスパッタ(MSP-1S) を用いて跗節を白金パラジウムでコーティングし、観察試料とした。 SEM による跗節の観察 ハムシ科の跗節の観察に用いたものと同じ SEM を用いて観察を行った。供試した種の 跗節を観察し、味覚感覚子と推定される感覚子(先端孔をもつ感覚子)の有無を調査し た。 TEM 観察試料の作製 供試虫 オオニジュウヤホシテントウHenosepilachna vigintioctomaculata の成虫を観察に用 いた。宮城県名取市にある雷神山古墳のジャガイモ畑から採集したものを初代とし、24 ± 1 ℃、明暗条件 16L: 8D の昆虫飼育槽内で飼育した個体を用いた。成育ステージご とにプラスチックシャーレに分け、食草および水分を適宜補充して飼育した。食草には 旧東北大学農学部の圃場または温室、東北大学青葉山新キャンパス内の温室で栽培した トマトSolanum lycopersicum の葉を与えた。
27 試料の作製 TEM 観察試料の作成方法は第 1 章第 2 節に準じた。ただし、跗節のサイズがイチゴ ハムシなどよりも大きいことから、オスミウム酸を用いた後固定の固定時間は 2.5 h と した。 TEM による跗節の観察 ハムシ科の跗節の観察に用いたものと同じ TEM を用いて観察を行った。 2.結果 オオニジュウヤホシテントウの跗節上で観察された感覚子の外部形態および内部形 態を図 11 に示した。外部形態観察では先端孔をもつ感覚子は観察されず、内部形態観 察でも味覚神経を備える感覚子は確認されなかった。また、コガネムシ科のカブトムシ、 ナガヒラタムシ科のナガヒラタムシ、テントウダマシ科のヨツボシテントウダマシでは、 オオニジュウヤホシテントウの跗節上で観察されたものと類似した感覚子が確認され たものの、跗節味覚感覚子は観察されなかった。外部形態観察でみられた感覚子はハム シ科において観察された跗節上の機械感覚子と類似していた。
28
図 11:オオニジュウヤホシテントウの跗節上の感覚子の外部形態および内部形態 A:跗節上の感覚子先端
29 第 4 節 考察 外部形態観察から、観察を行ったすべての種(全亜科)の跗節上に先端孔をもつ感覚 子が存在することが明らかになった。さらに、内部形態観察から、外部形態観察で先端 孔が確認された感覚子はその形状に関わらず、感覚子内腔に神経細胞の樹状突起を備え ていることが明らかになった。これらのことから、外部形態観察の結果が内部形態観察 からも裏付けられ、ハムシ科において跗節味覚感覚子の存在は共通した特徴であると考 えられた。 今回観察を行った 3 種では、感覚子内腔の樹状突起の数はいずれも 4 本であった。味 覚感覚子が備える味覚神経の本数は種や個体上の分布箇所によって異なる可能性は考 えられるが、4 本、または 2 本の神経細胞を備えるものが多く報告されており、ハムシ 科においてもそれが共通していると考えられた。ショウジョウバエ(Nayak and Singh, 1983)やオオユスリカChironomus anthracinus(Wülker et al., 1982)では跗節上の味 覚感覚子内が大室と小室に分かれていることが明らかにされているが、本研究ではその ような味覚感覚子は観察されなかった。観察された味覚感覚子横断面はセイヨウミツバ チで報告されているものに類似しており、分類群によって味覚感覚子の構造が異なる可 能性が示唆された。味覚感覚子が備える味覚細胞から伸長する樹状突起は、感覚子先端 付近、または感覚子の途中まで伸びることが明らかにされている(Zacharuk, 1980)。こ のことから感覚子内腔でみられた樹状突起は味覚細胞から伸びていると考えられ、外部 形態から味覚感覚子であると推定されたものは内部形態からも味覚感覚子であると裏 付けられた。また、味覚感覚子内の味覚神経はそれぞれ別の味を受容し、単独の味覚感 覚子が複数の味を受容することが明らかにされている(Ishimoto and Tanimura, 2004; Bruyne and Warr, 2005)。本研究の形態学的調査でも複数の神経が味覚感覚子内腔にみ られたことから、ハムシ科の跗節味覚感覚子内にみられた 4 本の樹状突起はそれぞれ異 なる味を受容すると考えられた。 単孔性の感覚子は機械感覚神経を備えることも知られており、機械感覚神経から伸び る樹状突起は感覚子基部またはその下部に樹状突起の終末が存在する( Zacharuk, 1980)。本研究の観察でもイチゴハムシの跗節味覚感覚子の基部にまで伸びる樹状突起 が観察された。また、機械感覚細胞は細管体(tubular body)と呼ばれる構造をもち、 これは感覚子の基部に位置するとされる。イチゴハムシの跗節味覚感覚子基部にもショ ウジョウバエやセイヨウミツバチで観察された 細管体と類似した構造がみられた (Whitehead and Larsen, 1976; Nayak and Singh, 1983)。また、イチゴハムシの跗節上 の機械感覚子の基部にも同様の構造がみられた。これらのことから、イチゴハムシの跗 節味覚感覚子基部まで伸びる神経細胞は機械感覚細胞であり、ハムシ科の味覚感覚子も 機械感覚神経を備えると考えられた。
30 結合繊毛部では神経細胞内の微小管がみられ、9 対の微小管が円形に並んでいる様子 が観察された。これは昆虫の化学受容感覚子へと伸長する神経細胞の結合繊毛部で一般 にみられるパターンであり(Zacharuk, 1980)、これらの神経細胞が味覚感覚子へと伸 長していることの裏付けとなった。また、観察された細胞の中には中央部にも微小管ら しき構造を備えるものもあった。カメムシ目の一種である Pyrops candelaria では繊毛 部で中心にも微小管を備える神経細胞の存在が報告されており(Lewis and Marshall, 1970)、本研究で見られたものも微小管であると考えられた。
先端の形状からハムシ科の跗節味覚感覚子は少なくとも 2 種類に分類できることが 明らかになった。これらの感覚子の先端で見られた構造は Zacharuk(1980)が a simple pit pore または a sculptured porous point として言及した構造に酷似しており、その文 献で著者らは a sculptured porous point をもつ味覚感覚子が開閉する可能性についても 言及している。このことから、本研究の観察でハムシ科に見られた 2 種類の跗節味覚感 覚子はそれぞれ異なる役割を果たす可能性が考えられた。2 種類の跗節味覚感覚子は同 一亜科に属する種ではどちらか一方で共通しており、亜科内で異なる種類の跗節味覚感 覚子が観察されることはなかった。Type II の跗節味覚感覚子が観察されたのはホソハ ムシ亜科、カメノコハムシ亜科、トゲハムシ亜科、コブハムシ亜科の 4 亜科だけである が 、 これ らの 亜科 のうち 、 特に カメ ノコ ハムシ 亜 科と トゲ ハム シ亜科 は とも に Cryptophalinae に属し、核 18S rRNA、28S rRNA、ミトコンドリア 16S rRNA のホモ ロジー解析から系統的に非常に近いことが報告されている(Gomez-Zurita et al., 2007)。 また、ともに単子葉植物食の系統に分類され(Wilf et al., 2000; Gomez-Zurita et al., 2007)、この 2 亜科を 1 つの亜科(Cassidinae)として扱う研究もある。カメノコハム シ亜科とトゲハムシ亜科の跗節は 4 節からなるが、この点で偽 4 節を呈する他の亜科 と異なっていることや、ミトコンドリアゲノムを解析した研究からやや独立した系統だ と考えられる(Nie et al., 2019)。ホソハムシ亜科は種数が少なく、系統的な位置づけが 不明瞭な部分もあるが、ミトコンドリアゲノムの解析からは、この亜科がハムシ科の分 化の初期の段階で他の主要なクレードから分化したことが示唆されている(Nie et al., 2019)。これらを合わせて考えると、Type II の感覚子は各亜科で独立して新しく獲得さ れた形質ではなく、分化の過程でどちらかの種類の感覚子のみが失われていったと推測 される。 コガネムシ科、ナガヒラタムシ科、テントウダマシ科、テントウムシ科のそれぞれに 属する種では跗節上に味覚感覚子の存在が確認されなかった。先行研究から、テントウ ムシ科ではインゲンテントウでも形態観察で跗節上の味覚感覚子の存在を確認できな かったと報告されていることから(Fischer and Kogan, 1986)、テントウムシ科は跗節 味覚感覚子をもたない可能性が考えられた。これらのことは、コウチュウ目において跗 節上の味覚感覚子は必ずしも共通した特徴ではないことを示唆すると考えられた。
31
第 3 章 ハムシ科における跗節味覚感覚子を介した味受容
第 2 章での形態学的調査から、ハムシ科において跗節味覚感覚子が共通した特徴であ ることが示唆された。また、イチゴハムシとカメノコハムシの跗節味覚感覚子の内部形 態観察から、ハムシ科の跗節味覚感覚子は内腔に 4 本の樹状突起を備えることが明らか になった。しかし、これらの神経が実際に味物質を受容するかは不明である。また、ハ エ目では一般的に味覚感覚子は 4 本の樹状突起を備え、例えば糖、水、低濃度の塩、高 濃度の塩を受容する神経を備える味覚感覚子の存在が報告されている(Yarmolinsky, 2009)。このことから、ハムシ科の跗節味覚感覚子が備える味覚神経もそれぞれ異なる 味を受容することが考えられた。これらの味覚ニューロンは味物質を受容すると活動電 位を生じさせるため(山元, 1986)、それを計測することで実際に味物質を受容するか確 認できる。そこで、飼育が容易で、これまでに摂食や寄主の探索に関わる化学物質が明 らかにされている(Matsuda and Matsumoto, 1974; Matsuda, 1983; Adati and Matsuda, 1993; Hori et al., 2006)イチゴハムシをモデルとして電気生理学的試験を行い、ハムシ 科の跗節味覚感覚子による味受容の有無や、各感覚子が受容する味物質の種類を調査し た。 第 1 節 イチゴハムシの跗節味覚感覚子による塩受容 昆虫の味覚感覚子による塩受容に関しては複数の目で報告されている(McCutchan, 1969; Rees, 1969)。また、NaCl や KCl は味受容細胞の神経応答の記録を行う際に電解 質として一般的に用いられることから、他の物質の受容を調査するうえでも塩受容の有 無を明らかにすることは必須である。そこで、形態観察から味覚感覚子とされたイチゴ ハムシの跗節上の感覚子の塩受容の有無を調査した。 1.材料および方法 供試虫 第 2 章第 2 節と同様の方法で累代飼育したイチゴハムシの成虫を用いた。試験には 羽化後 3~8 日後の個体を用いた。 供試物質 KCl(和光一級、和光純薬工業株式会社、大阪)、および NaCl(和光一級、和光純薬 工業株式会社、大阪)を用いた。 電気生理応答の記録32
試験にはチップレコーディング法を用いた(Hodgson et al., 1955: Marion-Poll & Van Der Pers, 1995; Delventhal and Carlson, 2014)。供試虫はピンセットを用いて鞘翅、中 脚、後脚を切除して用いた。切除処理後のイチゴハムシを練りゴムを用いてスライドグ ラス上に固定した。この際、電気生理応答の記録を行う感覚子が存在する部位が観察で きる状態にし、前胸と中胸の間の体節が露出するように固定した(図 12)。不関電極、 記録電極にはガラス電極を使用した。マイクロピペットプラー(P-97、Sutter Instrument、 California、US)を用いて作成したガラスマイクロピペットの先端をセラミックタイル カッターで適切な長さにカットし、ガラス電極に使用した。不関電極は、100 mM KCl 水溶液で満たしたガラスマイクロピペットに銀線を挿し込んだものとし、供試虫の前胸 と中胸の間の体節から、前胸に向けて挿入した(図 12)。記録電極には不関電極よりも 先端が長いガラスマイクロピペットを使用し、試験溶液を満たしたマイクロピペットに 銀線を差し込んだものを記録電極とした。記録の際には、マイクロマニピュレーター (MHW-103、株式会社成茂科学機械研究所、東京)を用い、光学顕微鏡(SteREO Discovery V20、Carl Zeiss、Oberkochen、Germany)下で、供試する跗節味覚感覚子の 先端が記録電極の先端口に入るように近づけた。得られた神経電位応答は微小電極用増 幅機(Taste Probe DTP-1、SYNTECH、Hilversum、Netherlands)を経由した後、これ と接続されたデータ収集・解析システム(IDAC4、SYNTECH)を用いてデータを収集 し、データ解析ソフト(Autospike ver. 3.9、SYNTECH)を用いて解析した。電気生理 応答の記録は試験溶液が感覚子の先端に触れた瞬間に開始されるように設定し、3 秒間 記録した。同一の感覚子から連続して電気生理応答を記録する際には、低い濃度の試験 溶液から順に作用させた。また、同一の試験溶液は1つの感覚子に対し 1 度だけ作用さ せた。直前に使用した試験溶液の影響を最小限にするため、連続した記録と記録の間に は 3 分以上の間隔を設け(Loy et al., 2016)、純水で味覚感覚子先端を洗浄した。前脚 跗節による味受容または味認識が複数の昆虫種で報告されていることから(Chun and Schoonhoven, 1973; Ozaki et al., 2011)、前脚跗節先端節上に存在する味覚感覚子の電 気生理応答を調査した。本研究では前脚跗節先端節上の異なる位置に存在する 3 つの感 覚子(C1–C3、図 13)の電気生理応答を記録し、解析した。味物質を作用させる際に生 じ得る物理的な刺激が記録に影響を及ぼす可能性があること、断続的な刺激に対して味 覚神経の応答(神経電位の発火頻度)が小さくなっていくことから、記録開始後 200~ 700 ms に生じたスパイク数をカウントすることで味覚神経の応答を定量化した (Dahanukar et al., 2001; Weiss et al., 2011)。
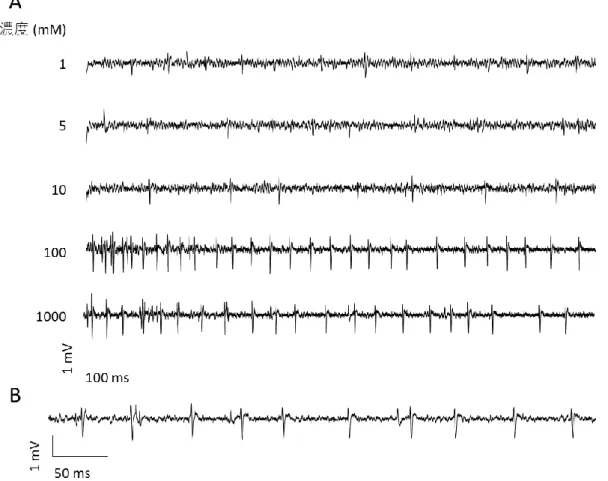
1、5、10、100、1000 の KCl、または NaCl 水溶液を跗節味覚感覚子に作用させ、電 気生理応答を記録した。
33 イチゴハムシ雌雄成虫の跗節味覚感覚子の KCl に対する応答を図 14 に示した。供試 した感覚子は全て KCl に対して濃度依存的な電気生理応答を示した。感覚子 C3 が KCl を受容して生じたスパイクの頻度はいずれの濃度でも感覚子 C1、C2 と比べて低かっ た。KCl 受容によって生じたスパイクの形状は図 15 示した。KCl 受容によって生じた スパイクは振幅が 1.2~1.5 mV で、マイナス側に突出した形状であった。 イチゴハムシ雌の跗節味覚感覚子の NaCl に対する応答を図 16 に示した。供試した感 覚子は全て、NaCl に対して濃度依存的な電気生理応答を示した。NaCl 受容によって生 じたスパイクの形状は図 17 に示した。NaCl 受容で生じたスパイクは KCl 受容で生じ たスパイクと類似しており、振幅が 1.2~1.5 mV で、マイナス側に突出した形状であっ た。
34
35
図 13:イチゴハムシの前脚跗節先端節
36 図 14:イチゴハムシの前脚跗節先端節上に存在する味覚感覚子の KCl に対する応答 A:雌の跗節味覚感覚子の KCl に対する応答 感覚子 C1–C3 の応答をスパイク頻度で示した(C1: N = 7, C2: N = 12, C3: N = 12) B:雄の跗節味覚感覚子の KCl に対する応答 感覚子 C2 の応答をスパイク頻度で示した(N = 5)
37
図 15:イチゴハムシ(雌)の跗節味覚感覚子の KCl に対する応答(波形) 感覚子 C2 から得られた波形を示した。
A:各濃度の KCl に対して生じた波形
38
図 16:イチゴハムシの前脚跗節先端節上に存在する味覚感覚子の NaCl に対する応答 感覚子 C1、C2 の応答をスパイク頻度で示した(C1: N = 4, C2: N = 12)
39
図 17:イチゴハムシ(雌)の跗節味覚感覚子の NaCl に対する応答(波形) 感覚子 C2 から得られた波形を示した。
A:各濃度の NaCl に対して生じた波形
40
第 2 節 イチゴハムシの跗節味覚感覚子による糖受容
糖は昆虫のエネルギー源であり(松香, 1984)、摂食刺激物質として働くことから餌の 摂取の可否を決定するうえで重要な物質である (Matsuda and Matsumoto, 1974: Nishida, 2014)。昆虫の味覚感覚子は一般に糖を受容する神経を備えるとされることか ら(Bruyne and Warr, 2005)、イチゴハムシの味覚感覚子が糖を受容するか調査した。 1.材料と方法 供試虫 試験に用いたイチゴハムシは第 3 章第 1 節に準じた。 供試物質 糖にはスクロース(和光特級、和光純薬工業株式会社、大阪)を用いた。 電気生理応答の記録 第 3 章第 1 節と同様の方法を用いた。 試験溶液として、0.1、1、3.16、10 mM のスクロース水溶液を用いた。各溶液には、 電解質として最終的な濃度が 5 mM となるように KCl を加えた。 2.結果 イチゴハムシ雌雄成虫の跗節味覚感覚子のスクロースに対する応答を図 18 に示した。 供試した感覚子のうち、C2 と C3 の感覚子はスクロースに対して濃度依存的な電気生 理応答を示したが、感覚子 C1 はいずれの濃度のスクロース水溶液を作用させても電気 生理応答を示さなかった(図 18)。スクロース受容によって生じたスパイクを図 19 示 した。スクロース受容で生じたスパイクは、振幅が 1~1.2 mV で、プラスとマイナスの 振幅がほぼ等しい形状であった。
41 図 18:イチゴハムシの前脚跗節先端節上に存在する味覚感覚子のスクロースに対する 応答 A:雌の跗節味覚感覚子のスクロースに対する応答 感覚子 C1–C3 の応答をスパイク頻度で示した(N = 12) B:雄の跗節味覚感覚子のスクロースに対する応答 感覚子 C2 の応答をスパイク頻度で示した(N = 5)
42
図 19:イチゴハムシ(雌)の跗節味覚感覚子のスクロースに対する応答(波形) 感覚子 C2 から得られた波形を示した。
A:各濃度のスクロース水溶液を作用させた際に得られた波形 B:10 mM スクロース水溶液を作用させた際に生じた波形の拡大
43
第 3 節 イチゴハムシの跗節味覚感覚子による苦味物質受容
毒物質や摂食阻害物質は一般に苦味として認識され、さまざまな種で苦味受容神経を 備える味覚感覚子の存在が報告されている(Weiss et al, 2011; Sparks and Dickens, 2016; Seada et al., 2018)。苦味の認識は適切な寄主の選択や毒の回避において重要であるこ とから、ハムシ科も跗節で苦味を受容できる可能性が高い。そこで本節では、ハムシの 跗節による苦味受容について電気生理応答試験により調査した。 1.材料と方法 供試虫 試験に用いたイチゴハムシは第 3 章第 1 節に準じた。 供試物質 苦味物質にはブルシン(和光特級、和光純薬工業株式会社、大阪)を用いた。 電気生理応答の記録 第 3 章第 1 節と同様の方法を用いた。 試験溶液として、0.001、0.01、0.1、1 mM のブルシン水溶液を用いた。各溶液には、 電解質として最終的な濃度が 5 mM となるように KCl を加えた。 2.結果 イチゴハムシ雌雄成虫の跗節味覚感覚子のブルシンに対する応答を図 20 に示した。 試験に供試したすべての感覚子がブルシンに対して濃度依存的な応答を示した(図 20)。 ブルシン受容によって生じたスパイクの形状を図 21 に示した。ブルシン受容で生じた スパイクの振幅は 1~1.2 mV であり、プラス側よりもマイナス側に大きく突出する形 状であった。スパイクの形状は塩受容によって生じたスパイクとやや類似していたもの の、振幅が異なっていた(図 22)。
44 図 20:イチゴハムシの前脚跗節先端節上に存在する味覚感覚子のブルシンに対する応 答 A:雌の跗節味覚感覚子のブルシンに対する応答 感覚子 C1–C3 の応答をスパイク頻度で示した(C1: N = 7, C2: N = 12, C3: N = 12)。 B:雄の跗節味覚感覚子のブルシンに対する応答 感覚子 C2 の応答をスパイク頻度で示した(N = 5)。
45
図 21:イチゴハムシ(雌)の跗節味覚感覚子のブルシンに対する応答(波形) 感覚子 C2 から得られた波形を示した。
A:各濃度のブルシン水溶液を作用させた際に得られた波形 B:0.1 mM ブルシン水溶液を作用させた際に生じた波形の拡大
46
47
第 4 節 イチゴハムシの跗節味覚感覚子による寄主植物葉表面ワックス受容
食葉性昆虫は寄主の葉上に到達すると、葉表面ワックスの化学的、物理的特性に影響 を受けて試咬や産卵といった行動を決定することが知られている(石川, 2007; Powell et al., 1999; Braccini et al., 2015)。チョウ目では幼虫の寄主となる植物の葉上に到達す ると、跗節で葉を叩くドラミングと呼ばれる行動をとることが知られており、跗節で葉 表面または葉内の味物質を認識することでその植物が産卵場所として適切であるか判 断できる(Renwick and Chew, 1994)。また、イチゴハムシをはじめとするハムシ科昆 虫においても寄主植物の葉表面ワックスに含まれる物質に対する選好性が報告されて いる(Adati and Matsuda, 1993)。そこで、本節ではイチゴハムシの跗節味覚感覚子が 寄主であるエゾノギシギシRumex obtusifolius(タデ科)の葉表面ワックスを受容する か調査した。 1.材料と方法 供試虫 試験に用いたイチゴハムシは第 3 章第 1 節に準じた。 供試植物 試験には東北大学青葉山新キャンパス構内に自生していたエゾノギシギシを採集し て用いた。 葉表面ワックスの抽出 抽出には無傷の完全展開葉を用いた。採集した葉はスキャナー(GTX-9000:セイコ ーエプソン株式会社、長野)を用いて撮影し、画像解析用フリーソフト ImageJ を用い て面積を測定した。抽出溶媒にはヘキサン(和光一級、富士フィルム和光純薬株式会社、 大阪)を用い、1 回の抽出に 800 mL の溶媒を使用した。ワックスの抽出は葉をヘキサ ンに 30 秒ディッピングすることで行った。抽出液はろ紙(ADVANTEC®︎ No.1、アド バンテック東洋株式会社、東京)を用いてろ過し、その後ロータリーバキュームエバポ レーター(N1200A-S 型、東京理科機械株式会社、東京)を用いて 39 ℃で減圧乾固し、 粗抽出物を得た。 試験溶液の作製 エゾノギシギシ葉表面ワックスは疎水性の物質を含み、純水に溶解させるのは難しい ことから、界面活性剤であるポリオキシエチレンソルビタンモノウラレート(生化学用、
48 フジフィルム和光純薬株式会社)を 1%の濃度で含む水溶液に溶かし、試験溶液とした。 試験溶液には電解質として最終的な濃度が 5 mM となるように KCl を加えた。 電気生理応答の記録 第 3 章第 1 節と同様の方法を用いた。 2.結果 イチゴハムシ雌雄成虫の跗節味覚感覚子のエゾノギシギシ葉表面ワックスに対する 応答を図 23 示した。供試した感覚子のうち、感覚子 C2 のみがエゾノギシギシ葉表面 ワックス溶液に対し濃度依存的な応答を示した。葉表面ワックス受容で生じたスパイク を図 24 に示した。葉表面ワックス溶液を作用させた際には、振幅の異なる 2 種類のス パイクが生じた。振幅が小さいものをスパイク a、大きいものをスパイク b とした。ス パイク a の振幅は 0.8~1 mV であり、多くがややマイナス側に突出した形状を示した。 スパイク b の振幅は 1~1.2 mV で、プラス側とマイナス側の振幅がほぼ同等であった。
49 図 23:イチゴハムシの前脚跗節先端節上に存在する味覚感覚子のエゾノギシギシ葉表 面ワックスに対する応答 A:雌の跗節味覚感覚子の葉表面ワックスに対する応答 感覚子 C1–C3 の応答をスパイク頻度で示した(N = 12)。 B:雄の跗節味覚感覚子の葉表面ワックスに対する応答 感覚子 C2 の応答をスパイク頻度で示した(N = 5)。
50 図 24:イチゴハムシ(雌)の跗節味覚感覚子のエゾノギシギシ葉表面ワックス溶液に 対する応答(波形) 感覚子 C2 から得られた波形を示した。 A:各濃度のワックス溶液に対する波形 B:1 µg/µL のワックス溶液を作用させた際にみられたスパイク。白矢印:スパイク a、 黒矢印:スパイク b
51 第 5 節 コガタルリハムシの跗節味覚感覚子による塩受容 第 4 節までの結果から、イチゴハムシの跗節味覚感覚子が実際に味を受容する能力を 備えることが明らかになった。ハムシ科で観察された跗節味覚感覚子に味を受容する能 力があるかさらに裏付けるため、イチゴハムシ(ヒゲナガハムシ亜科)とは異なる亜科 に属するコガタルリハムシGastrophysa atrosyanea(ハムシ亜科)を用いて電気生理応 答試験を行った。 1.材料および方法 供試虫 コガタルリハムシは宮城県仙台市青葉区荒巻字青葉の東北大学青葉山新キャンパス の農学研究科の圃場内で採集し、24 ± 1 ℃、明暗条件 16L: 8D の昆虫飼育槽内で飼 育したものを用いた。食草のエゾノギシギシは東北大学青葉山新キャンパス構内および 宮城県仙台市青葉区内に自生していたもの、または東北大学青葉山新キャンパス内の温 室で栽培したものを使用した。試験には雄成虫を用いた。 供試物質 味物質には KCl(和光一級)を用いた。 電気生理応答の記録 第 4 章第 1 節に準じた。コガタルリハムシ(図 25)はイチゴハムシよりも 大きく、より硬い表皮を持つため、セロハンテープを用いてスライドグラス上 に固定して試験に用いた(図 26)。 1、5、10、100、1000 mM の KCl 水溶液を跗節味覚感覚子に作用させ、電気生理応 答を記録した。 2.結果 コガタルリハムシの前脚跗節上の味覚感覚子の KCl に対する応答を図 27 に示した。 試験に供試した感覚子は KCl に対して濃度依存的な応答を示した。KCl 受容で生じた 波形とスパイクの形状を図 28 に示した。KCl 受容で生じたスパイクの形状はイチゴハ ムシでみられたものと類似していた。
52
53
54
図 27:コガタルリハムシの前脚跗節先端節上の味覚感覚子の KCl に対する応答 単一の感覚子の応答をスパイク頻度で示した(N = 5)。
55
図 28:コガタルリハムシ前脚跗節味覚感覚子の KCl 水溶液に対する神経応答(波形) 各濃度の KCl に対する応答を示した。
56 第 6 節 考察 供試した感覚子は KCl と NaCl のそれぞれの水溶液に対して電気生理応答を示した ことから、イチゴハムシの跗節味覚感覚子の内腔にみられた 4 本の神経細胞樹状突起外 節のうちの 1 つは塩の受容体をもつと考えられた。昆虫の塩受容体としては、キイロシ ョウジョウバエでイオンチャネル内蔵型レセプターである Ir76b という受容体が報告 されており(Freeman and Dahanukar, 2015)、ハムシ科も同様にイオンチャネル内蔵型 レセプターを味覚神経細胞上にもつと考えられた。KCl と NaCl の受容によって生じた 神経細胞の活動電位はスパイクの振幅、形状が類似していた。通常、味覚の種類(甘味、 苦味等)はある 1 つの種類の感覚細胞によって決定されるのが一般的であるため、これ らの物質は同一の神経細胞によって受容されると考えられた。しかし、ショウジョウバ エでは複数の異なる種類の神経細胞、例えば「甘味受容細胞」や「苦味受容細胞」も塩 を受容することが明らかになっている(Jaeger et al., 2018)。このことから、KCl と NaCl が複数の感覚細胞で受容される可能性もある。キイロショウジョウバエの唇弁上の味覚 感覚子は低濃度の塩に応答するニューロンと高濃度の塩に応答するニューロンを備え るが(Jaeger et al., 2018)、本研究でも KCl の受容によって生じた跗節味覚感覚子から のスパイクは低濃度と高濃度で異なっていた。これが異なる感覚細胞の応答によるもの であれば、ショウジョウバエと同様に塩受容細胞が複数存在するとも考えられる。しか し、スパイクの形状は神経細胞の順化などによっても変化しうるため、それを明らかに するためには塩受容体の発現部位を詳細に明らかにする必要がある。 スクロースに対しては感覚子 C2 と C3 が電気生理応答を示し、感覚子 C1 では電気 生理応答は見られなかった。一方、苦味物質であるブルシンに対しては供試した 3 つの 感覚子の全てが電気生理応答を示した。このことから、感覚子によって受容する味物質 が異なると考えられた。スクロースのような糖は一般に甘味として認識され、ブルシン のような物質は苦味として認識される。先行研究から、苦味受容ニューロンは甘味受容 ニューロンに比べて広範な種類の味覚感覚子内に存在することが示唆されている (Weiss et al, 2011)。本研究でも甘味受容を示した感覚子は苦味受容を示したものより も少なかったことから、イチゴハムシでも苦味受容ニューロンがより多くの味覚感覚子 内に存在していると考えられた。 塩(KCl、NaCl)とスクロース、ブルシンの受容によって生じたスパイクはその振幅 や形状がそれぞれで異なっていた。これは、塩とスクロース、ブルシンがそれぞれ別々 の感覚細胞で受容されるためと考えられた。もしそうであれば、それぞれの味物質は異 なる種類の味として認識されると考えられた。 イチゴハムシの跗節上の味覚感覚子 C2 はその寄主であるエゾノギシギシの葉表面ワ ックスに対して電気生理応答を示した。葉表面ワックスは疎水性の物質を多く含むため、