人工生命による生態進化モデルとその応用に関する 研究
著者 武藤 敦子
学位名 博士(工学)
学位授与番号 13903乙第265号 学位授与年月日 2010‑03‑16
URL http://id.nii.ac.jp/1476/00002916/
人工生命による生態進化モデルと その応用に関する研究
2 0 1 0
年武 藤 敦 子
i
目 次
第
1
章 序論1
1.1
研究の背景. . . . 1
1.1.1
人工生命の先天的情報. . . . 3
1.1.2
人工生命の後天的情報. . . . 4
1.1.3
人工生命の生物学的応用. . . . 6
1.1.4
人工生命の工学的応用. . . . 8
1.2
研究の目的. . . . 12
1.3
研究の内容. . . . 13
第
2
章 多出力二分決定グラフのAPPLY
交叉を用いた食物連鎖モデル15 2.1
はじめに. . . . 15
2.2 BDD
とAPPLY
演算. . . . 16
2.2.1 BDD . . . . 16
2.2.2 APPLY
演算. . . . 17
2.3 n-BDD
を用いた遺伝子表現. . . . 19
2.3.1
多出力二分決定グラフ(n-BDD) . . . . 19
2.3.2 n-BDD
の遺伝的操作. . . . 19
2.4 APPLY
交叉の提案. . . . 20
2.4.1 APPLY
交叉の効果. . . . 21
2.5
生態系モデル. . . . 24
2.5.1
生態系モデルの概要. . . . 24
2.5.2 APPLY
交叉を用いた交配の導入. . . . 25
2.6
実験. . . . 26
2.6.1
実験環境. . . . 26
2.6.2
結果と評価. . . . 27
2.7
おわりに. . . . 30
第
3
章 多出力二分決定グラフのFlexible APPLY
交叉を用いた生態分化モデル33 3.1
はじめに. . . . 34
3.2
生態分化モデル. . . . 35
3.2.1
エージェントの定義. . . . 35
3.2.2
エージェントの生成. . . . 35
3.2.3
生殖隔離. . . . 36
3.2.4
エージェントの知覚と行動. . . . 36
3.2.5
エージェントの内部状態. . . . 36
3.2.6
エージェントの死滅. . . . 37
3.2.7
生態進化メカニズム. . . . 37
3.2.8
生態分化メカニズム. . . . 37
3.3 Flexible APPLY
交叉の提案. . . . 38
3.3.1
占有率. . . . 39
3.3.2 Flexible APPLY
交叉. . . . 39
3.3.3 Flexible APPLY
交叉の効果. . . . 40
3.3.4
実験結果. . . . 41
3.4
実験. . . . 42
3.4.1
エージェントの適温帯. . . . 43
3.4.2
エージェントの食糧. . . . 43
3.4.3
行動戦略. . . . 44
3.4.4
エネルギー変化量. . . . 44
3.4.5
実験結果. . . . 45
3.4.6
環境順応値と行動戦略. . . . 47
3.4.7
生殖隔離の影響. . . . 48
3.4.8 Flexible APPLY
交叉の効果. . . . 50
3.5
おわりに. . . . 51
第
4
章 同調遺伝子とミームを用いた性選択モデルによる循環型流行の発現53 4.1
はじめに. . . . 53
4.2
エージェントモデル. . . . 55
iii
4.2.1
エージェントの定義. . . . 55
4.2.2
同調遺伝子. . . . 55
4.2.3
形質の派手さの定義. . . . 57
4.2.4
エージェントの行動. . . . 57
4.3
実験. . . . 60
4.3.1
実験環境. . . . 60
4.3.2
実験結果. . . . 61
4.3.3
考察. . . . 62
4.4
おわりに. . . . 68
第
5
章 同調・差別化欲求を持つエージェントモデルによる多種循環型流行の発現69 5.1
はじめに. . . . 69
5.2
エージェントモデル. . . . 70
5.2.1
エージェントの定義. . . . 70
5.2.2
同調化欲求遺伝子. . . . 71
5.2.3
形質の派手さの定義. . . . 72
5.2.4
エージェントの行動. . . . 72
5.3
実験. . . . 76
5.3.1
環境. . . . 76
5.3.2
結果. . . . 77
5.3.3
考察. . . . 78
5.3.4
カタジロクロシトドの流行現象. . . . 80
5.3.5
従来モデルとの比較. . . . 80
5.3.6
参照集団数を変化させた実験. . . . 81
5.4
おわりに. . . . 82
第
6
章 動的多段交叉を用いた実数値遺伝的アルゴリズムの効率化85 6.1
はじめに. . . . 85
6.2
多段交叉による局所探索. . . . 86
6.2.1
探索履歴の利用. . . . 86
6.2.2
多段交叉. . . . 86
6.2.3
多段交叉を用いた世代交代モデル. . . . 88
6.2.4
多段交叉を用いたMGG
の挙動. . . . 89
6.3
動的多段交叉. . . . 92
6.4
実験. . . . 93
6.4.1
テスト関数を用いた実験. . . . 93
6.4.2
実験結果と考察. . . . 93
6.4.3
実問題を用いた実験. . . . 94
6.4.4
実験結果と考察. . . . 99
6.5
おわりに. . . . 99
第
7
章 結論103
付 録A
シンプレクス交叉(SPX)107
付 録B SRM
制御パラメータ最適化問題108 B.1
適応度計算. . . . 108
付 録C
ロトカボルテラ(Lotka-Volterra)系110
付 録
D
関数最適化問題111
謝辞
116
参 考 文 献
119
研究業績
133
v
図 目 次
1.1
人工生命研究.. . . . 2
1.2
自然科学と人工生命の研究アプローチ.. . . . 2
1.3
群れ行動シミュレーションの様子(赤:捕食者,黒:被捕食者).. . . 3
1.4 n-BDD(n=4)
の例.. . . . 4
1.5
文化伝播シミュレーションの様子.. . . . 5
1.6
獲得した歌オートマトンの例.. . . . 5
1.7
種分化.. . . . 7
1.8
世代交代モデルの概念図.. . . . 9
2.1 BDD
のグラフ表現(a)
とデータ構造(b). . . . . 17
2.2 h(= f ◦ g)
のアルゴリズム.. . . . 18
2.3 n-BDD
の例(n= 4). . . . . 19
2.4 n-BDD
の遺伝的操作.. . . . 20
2.5 APPLY
交叉の例.. . . . 21
2.6
肉食エージェントの固定された戦略.. . . . 22
2.7
適応度平均の推移.. . . . 23
2.8
平均節点数の推移.. . . . 23
2.9
草食エージェントのBDD
の推移(例). . . . . 25
2.10
エージェントを扱う1
ステップの流れ.. . . . 27
2.11
シミュレーションの様子(59
×59). . . . . 28
2.12
エージェント別個体数推移.. . . . 29
2.13
エージェント別個体数推移(安定期間).. . . . 30
2.14 100
回のシミュレーション中の最長安定期間のステップ数分布.. . . . 31
2.15
絶滅までのステップ数の分布.. . . . 31
3.1
環境順応値と行動戦略によるエネルギー蓄積量の概念図.. . . . 38
3.2 Flexible APPLY
交叉の例.. . . . 40
3.3
目標とするn-BDD (n=22). . . . . 41
3.4
世代毎の平均適応度の推移.. . . . 42
3.5
時間毎の平均適応度の推移.. . . . 43
3.6
気温の知覚.. . . . 44
3.7
実験のスナップショット(左)
と発現した行動パターン(右). . . . . 46
3.8
安定期におけるエリア毎の個体数推移および気温変化.. . . . 47
3.9
行動戦略st
stayを採るエージェントのn-BDD
の一例.. . . . 48
3.10
行動戦略st
migrateを採るエージェントのn-BDD
の一例.. . . . 49
3.11
安定期における行動戦略st
stay,stmigrateを採るエージェントの環境順 応値b
iの分布.. . . . 49
3.12
安定期における行動戦略st
stay,stmigrateを採るエージェントの環境順 応値b
iの分布(生殖隔離なし). . . . . 50
4.1
提案エージェントモデル.. . . . 56
4.2
エージェントの行動モデル.. . . . 58
4.3
模倣対象となるオスa
kの選定(a)
とオスa
jおよびメスa
iの模倣行動(b). 60 4.4
実験(1)
における嗜好別メスエージェント平均生存割合.. . . . 62
4.5
実験(3)
における嗜好別メスエージェント平均生存割合.. . . . 63
4.6
実験(4)
における嗜好別メスエージェント平均生存割合.. . . . 63
4.7
実験(1)
における嗜好別メスエージェント生存割合*.. . . . 64
4.8
実験(1)
におけるメスエージェント嗜好遺伝子生存割合*.. . . . 65
4.9
実験(1)
におけるメスエージェント嗜好ミーム生存割合*.. . . . 65
4.10
実験(1)
における非同調エージェント生存割合*.. . . . 66
4.11
実験(1)
におけるメスの嗜好およびオスの形質の平均値.. . . . 67
4.12
実験(2)
におけるメスの嗜好およびオスの形質の平均値.. . . . 67
5.1
エージェントの1
ステップの行動モデル.. . . . 73
5.2
嗜好別メスエージェント生存割合.. . . . 77
5.3
同調化欲求遺伝子の平均値.. . . . 78
5.4
「クレイズ」および「ブーム」発生回数の平均値.. . . . 81
vii
6.1 SPX
を用いた多段交叉の例.. . . . 88
6.2
世代交代の流れ.. . . . 89
6.3 100
世代毎の平均成功回数および平均失敗回数(Schwefel 1.2 関数).. 90 6.4 100
世代毎の平均成功回数および平均失敗回数(Rastrigin関数).. . . 91
6.5
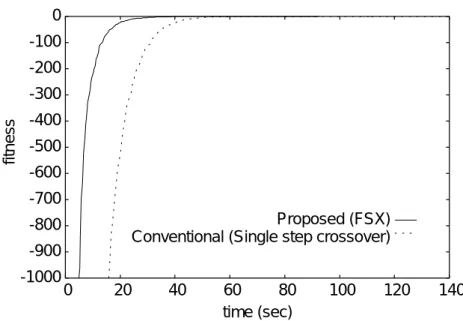
平均適応度の推移(Schwefel 1.2関数).. . . . 95
6.6 100
世代毎の段数選択回数の推移(Schwefel 1.2関数).. . . . 95
6.7
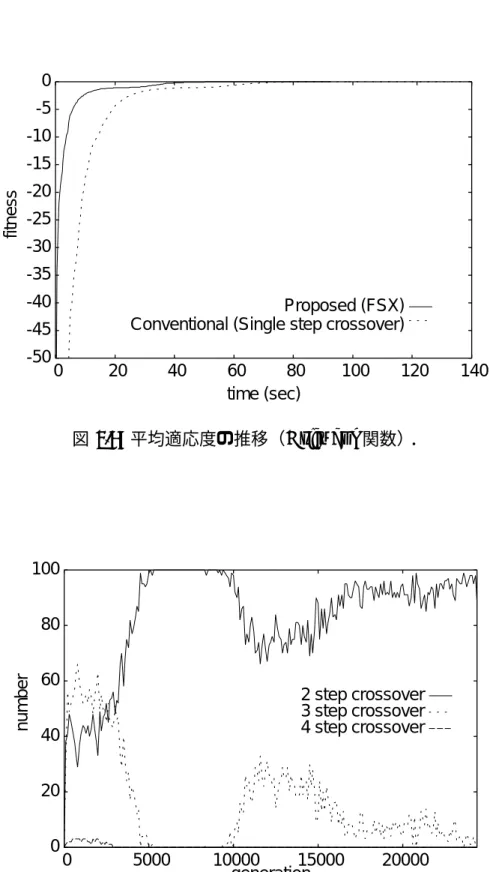
平均適応度の推移(Rosenbrock関数).. . . . 96
6.8 100
世代毎の段数選択回数の推移(Rosenbrock関数).. . . . 96
6.9
平均適応度の推移(Griewank関数).. . . . 97
6.10 100
世代毎の段数選択回数の推移(Griewank関数).. . . . 97
6.11
平均適応度の推移(Rastrigin関数).. . . . 98
6.12 100
世代毎の段数選択回数の推移(Rastrigin関数).. . . . 98
6.13
平均適応度の推移.. . . . 100
6.14 10
世代毎の段数選択回数の推移.. . . . 100
C.1
ロトカボルテラ系の個体数変動の例.. . . . 110
D.1 Schwefel 1.2
関数.. . . . 112
D.2 Rosenbrock
関数.. . . . 113
D.3 Griewank
関数.. . . . 114
D.4 Schwefel
関数.. . . . 114
D.5 Rastrigin
関数.. . . . 115
ix
表 目 次
1.1
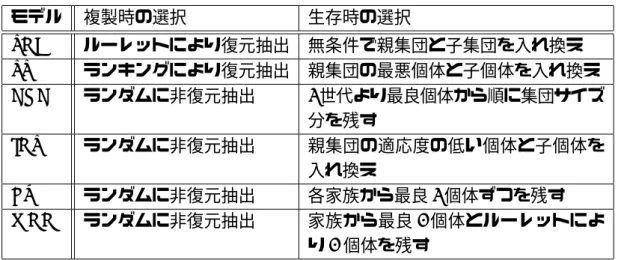
代表的な世代交代モデルの構成.. . . . 10
1.2
代表的な世代交代モデルの特徴.. . . . 10
1.3
交叉オペレータの比較.. . . . 11
2.1
入力ビット列の割り当てと意味.. . . . 26
2.2
エージェントが選択できる行動.. . . . 28
2.3
肉食動物又は草食動物絶滅までの平均ステップ数.. . . . 29
3.1
エージェントaiの知覚情報.. . . . 45
3.2
エージェントaiの行動.. . . . 45
4.1
実験の組合せと結果.. . . . 61
4.2
多数派に安定した嗜好の内訳.. . . . 68
5.1
「クレイズ」および「ブーム」回数の20
試行の平均値と標準偏差.. . 80
6.1
実験パラメータ.. . . . 94
6.2
最適解獲得平均計算時間(単位:sec). . . . . 94
6.3
実験パラメータ.. . . . 99
6.4
最適解獲得平均計算時間(単位:sec). . . . . 101
B.1
制御パラメータの探索範囲と間隔.. . . . 108
D.1
テスト関数の特徴.. . . . 115
1
第 1 章 序論
1.1
研究の背景生命に関わる諸現象を工学的手法で再現し解明しようとする生物学的研究,あるい は生命的な振る舞いにヒントを得た工学的研究が様々な分野で展開されている. こ のような研究アプローチは人工生命を用いて行なわれる.
人工生命研究は,一見して生命のように見えるものを工学的手法を用いて創造する 試みであり,1987 年に第
1
回人工生命国際会議においてChristopher Langton
が提唱 した概念である[1][2][3][4].人工生命は,もともと生物の進化のメカニズムのモデルと
して,集団遺伝学や進化生物学など,生物・進化に関連する学問分野(これらを総称し
て,以下,生物学分野) での利用を目的としていた.一方で,Holland[5]によって遺伝 的アルゴリズムの手法が定式化されてからは,工学的応用を目的とした組合せ最適化 問題や各種の最大値探索問題などへの人工生命手法の適用が盛んになっている.それ らの手法は総称して進化的計算と呼ばれ,代表的な手法として,遺伝的アルゴリズム[5],進化戦略 [6],進化的プログラミング [7],遺伝的プログラミング [8]
があり,最適化,学習,解析などの応用をされている.
以上をまとめると,人工生命研究は,図
1.1
に示すように生物システムを模倣する 進化シミュレータとしての生物学的応用と,進化的計算を用いた最大値探索問題に代 表される生物モデルの工学的応用に大別することができる[9].
人工生命の生物学的応用研究の位置付けは難しく,確立された研究方法というもの が存在しない.従来の典型的な自然科学では,具体的な現象やデータを観察し,そこか ら一般性を持つ仮説,法則,理論を推論する方法や,反対に,仮説,法則,理論を立て,
実験や観察を行なうことにより妥当性を裏付けていく方法が用いられる(図
1.2(a)).
↢‛ቇ
↢‛ࡕ࠺࡞ߩᎿቇ⊛
ታࠪࠬ࠹ࡓ߳ߩᔕ↪
↢‛ࠪࠬ࠹ࡓߩᮨ୮ߣ ߘߩࡔࠞ࠾࠭ࡓߩ⸃
↢‛␠ળ
ੱᎿ↢
Ꮏቇ
図
1.1:
人工生命研究.一方で,典型的な人工生命モデルは,具体的な現象やデータとのつながりは必ずしも 強く維持せずに,何らかの仮説や法則を制約として概念レベルにおいて人工生命モデ ルを構成する
[10].そして,計算機の中で時間発展させ,興味深い創発現象を起こし
得た時に,その挙動を従来の仮説や法則の中に位置付けながら解釈する.その解釈の 中で,あるいは元の人工生命モデルのレベルで,実際の現象やデータとの比較が可能 な場合もある.本研究では,人工生命の生物学的応用を図
1.2(b)
に示すような研究アプローチで行 う.特に本論文で行う部分は,人工生命モデルを計算機上で構成するための人工生命 手法の提案と,実際にそれらの手法を用いた人工生命モデルの構成である.人工生命 モデルは仮説・法則や具体的な現象・データ等を参照して作成し,計算機上での進化C⥄ὼ⑼ቇ
⺑ᴺೣ
⽎
࠺࠲
ᬌ⸽ ផ⺰
⺑ᴺೣ ⽎
࠺࠲
ޓޓޓ᭴ᚑ 㧔ੱᎿ↢ᚻᴺ㧕
ੱᎿ↢ࡕ࠺࡞
⸃㉼
ᬌ⸽DੱᎿ↢
図
1.2:
自然科学と人工生命の研究アプローチ.1.1.
研究の背景3
により創発した人工生命体の挙動を元の仮説・法則や現象・データと比較しながら解 釈または検証する.
1.1.1
人工生命の先天的情報地球上に存在する生命体が先天的に保持する情報として遺伝情報がある.人工生命 体の遺伝子表現としては,ビット列によるもの
[11][12][13][14][15],オートマトンを用
いたもの[16][17],木構造で表現するもの [18][19][20][21][22][23]
など様々な表現方法が 存在する.例えば,人工生命体の遺伝子を群れ行動決定のためのパラメータとしてビット列で 表現した研究がある
[24][25][26][27][28].これは,ある限られた範囲のフィールド上に
おいて群れ行動をするエージェント集団を存在させたシミュレーションモデルである(図
1.3).群れエージェント集団は遺伝子として群れ行動決定のためのパラメータを持
ち,適応度評価値を群れエージェントが全滅するまでの期間とすることで,群れエー ジェント集団が捕食エージェントから逃れて長く生存し続けるためのパラメータを自 然淘汰により獲得する.
図
1.3:
群れ行動シミュレーションの様子(赤:捕食者,黒:被捕食者).また,人工生命体が先天的に持つ行動戦略を遺伝的プログラミングを応用した多出 力二分決定グラフ
(n-output Binary Decision Diagram ;n-BDD)
で遺伝子表現した研究がある
[18][19][20][21][22][23].1978
年にAkers[29]
によって考案されたBDD
は論理 関数の表現方法の一つであり,その記憶効率や処理速度に優れることからLSI
やCAD
の分野を中心に,様々な分野に応用されている.BDDの出力値を0
か1
の2
通りからn
通りに拡張したものがn-BDD
(図1.4)である [20].n-BDD
の変数を用いてエージェ ントの知覚情報を表現し,n-BDDの出力,即ち定数節点をエージェントのn
種類の行 動に対応させることで,自律エージェントの行動戦略を表現できる.これを遺伝的プ ログラミング技法によって進化させることでエージェントの行動戦略の最適化を行う ことができる.詳細については本研究の第2
章において述べる.X3
0 X11
X2
A B C D
0
0
1
1
terminal node decision node
図
1.4: n-BDD(n=4)
の例.1.1.2
人工生命の後天的情報地球上には,先天的情報である遺伝子に加え生まれた後に学習や模倣により獲得 する後天的情報を持つ生命体が存在する
[30].近年,後天的に獲得した情報を文化
伝達子(ミーム)として表現する研究が盛んに行なわれている[31][32].ミームとは,
R.Dawkins[33]
により提唱された,模倣・教示・学習等によって後天的に獲得する文化伝達子の概念である.人工生命体のミーム表現としては,1.1.1節において述べた遺伝 子表現と同じように様々な表現方法が考えられる.
例えば,文化嗜好を経験によって後天的に獲得するミームとして定義することで,文 化嗜好を人工生命体に蓄積し伝播させるモデルがある
[34][35][36].図 1.5
は,文化伝 播シミュレーションのスナップショットであり,赤で示すA
というオブジェクトを嗜1.1.
研究の背景5
好するエージェントと青で示す
B
というオブジェクトを嗜好するエージェントが地域 毎に密集して存在し文化を伝播している様子が分かる.objectA objectB
agent(preferA) agent(preferB)
図
1.5:
文化伝播シミュレーションの様子.また,性淘汰の環境下における鳥の求愛の歌表現を後天的情報としてオートマトン を用いて表現した研究がある
[30].図 1.6
は,後天的に獲得した歌オートマトンの一例 であり,後天的情報である歌表現が学習により複雑に変化していることが分かる.0
c b e d
1
a e
d a
2
b
3
c
c b a e
4d c a d e
b b
e a c
d
5a d e
図
1.6:
獲得した歌オートマトンの例.1.1.3
人工生命の生物学的応用人工生命の生物学的応用として,本研究では未だ明らかとなっていない生命体の諸 現象について新たな進化モデルを提案し,実際に計算機上で構成した人工生命体の挙 動を確認することでそのメカニズムを探る.人工生命の構成には,先に述べた先天的 情報である遺伝子および後天的情報であるミーム等の人工生命表現を,構成するモデ ルによって組み合わせて用い,さらに必要に応じて人工生命手法(進化演算子)を定 義する.
本研究では,生命体の生態進化に関わる現象として主に二つを取り上げる.一つは 生物多様性を引き起こす要因となっている「種分化」,一つは異性をめぐる競争を通 じて起きる進化である「性選択(性淘汰)」である.どちらも,進化生物学における重 要な理論であり多くの生物学者が興味を抱いている研究分野である.
種分化
自然界に存在する生命体は進化の過程で様々な生態を発現し,その種類も多様化し ている.種の多様性を引き起こす生態の分化がどのようなメカニズムで生じるのかを 解明することは重要である
[37].種分化とは新しい生物学的種が誕生する進化プロセ
スの一つであり,種分化しつつある集団がどの程度母集団から地理的に隔離されてい るかで,以下のように3
分類できる.図1.7
にそれぞれの特徴を示す.( 1 )
異所的種分化( 2 )
側所的種分化( 3 )
同所的種分化一般的に種分化の多くが地理的隔離のある「異所的種分化」であると考えられている
が
[38],部分的に地理的隔離のない「側所的種分化」の確認例もいくつか存在する.近
年では地理的隔離の全くない「同所的種分化」を示す事例研究が報告され注目を集め
ている
[39][40].人工生命分野において同所的種分化を扱ったものには,性淘汰のみで
起こりうることを示したもの
[41][42]
や,相互作用による表現型の分化が遺伝型の進化 を促す様子を表現した研究[43]
などが報告されているがその数は事例研究と同様に少 ないため,さらなる研究報告が求められている.1.1.
研究の背景7
図
1.7:
種分化.性選択(性淘汰)
生態には,クジャクのオスの羽のように生物が生存する上で過度に派手で不利であ ると考えられる現象が見られる.
Darwin
はこのような現象を性選択(性淘汰)と定義 することで説明した[44][45].性選択には,異性による選り好み(配偶者選択)が存在
する.配偶者選択にどのようなメカニズムが働いているかを示す理論モデルは大きく 二種類に分けられる.一つはランナウェイ説のように,メスの選好の基準が生存上の 有利さとは無関係な場合,もう一つはハンディキャップ説のように生存上の有利さに繋 がる形質を選好の基準にしている場合である.ある形質や信号がランナウェイによっ て発達したのか,ハンディキャップによるものなのかは判断が難しい場合が多く,個々 の事例に関してさらなる研究報告が求められている[46].
人工生命分野において性選択を扱ったものには,オスを形質遺伝子,メスを嗜好遺 伝子で構成し計算機上で進化実験を行った研究
[47]
がある.また,性選択の過程には アズマヤドリのアズマヤのようにオスがメスを引きつけるために後天的に装飾すると いう現象がみられる[48].このような生物は,配偶者選択において先天的な身体的特
徴に加え後天的な嗜好対象オブジェクト(ミーム)[49][50][51][52]
を参照していると考 えられる.1.1.4
人工生命の工学的応用人工生命の工学的応用として進化的計算の中で最も代表的な手法に遺伝的アルゴリ ズムがある.遺伝的アルゴリズムの工学的応用としては,超
LSI
のパターン配置,ス ケジューリング,ニューラルネットワークの構成,広域ネットワークのルーティング,画像やデザインなど広範な問題がある.必ずしも常に最良の解が得られるとは限らな いが,コンピュータのパワーがあれば,他の方法より効率良くかなり良好な解を得る ことができる
[3].遺伝的アルゴリズムを適用するとよいと考えられる代表的な工学的
問題として,巡回セールスマン問題やナップザック問題などに代表される組合せ最適 化問題がある.巡回セールスマン問題とは,与えられた複数の都市の全てを,各都市 をそれぞれ1
回だけ訪問するという条件のもとで巡回する際の経路長を最小にする問 題であり,ナップザック問題とは,大きさが異なる複数の荷物を容量の決まった袋に ちょうどよく詰め込む問題である[53].
しかし,これらの問題は計算理論の検証のための問題という意味合いが濃く,実際 の工学的問題を考えた時,実問題の多くはシミュレータを用いた評価を伴うため計算 負荷が高く,試行錯誤的な探索手法である遺伝的アルゴリズムを用いることでさらに 計算時間がかかるという問題点がある.
遺伝的アルゴリズムの計算負荷を解消することを目的とし,探索履歴を用いた交叉 を分散遺伝的アルゴリズムに適用する研究がある
[54][55][56].ここでは,生成された
個体のシミュレーションを伴う評価に探索履歴との類似度を用いた適応度予測を用い ることで評価計算コストを削減し,対象問題による交叉回数の制限のない複数回交叉 手法を実現した.しかしながら,実問題を遺伝的アルゴリズムに適用する際には遺伝 子表現に実数値を用いる方が効率的であり,実数値遺伝的アルゴリズムが近年では主 流となっている.よって,本研究では,実数値遺伝的アルゴリズムにおける探索の効 率化を目的とした新たな探索手法を考える[57].
世代交代モデルの設計
進化的計算において,よりよい個体を次世代に生み出すための多くの拡張手法が提 案されている
[58][59][60][61][62][63][64][65].よりよい個体を次世代に生み出すために
は,適切な世代交代モデルおよび交叉の設計が不可欠である.遺伝的アルゴリズムにおける世代交代モデルの代表例として,Simple Genetic Al-
1.1.
研究の背景9
gorithm(SGA)
がある.今までに,多様性の観点からSGA
に対して改善を行ったIter-
ated Genetic Search(IGS), Steady State(SS), CHC, Elite Recombination(ER), Minimal Generation Gap(MGG)
などが提案されている[66].これらのモデルには,
「世代交代 の連続化」を目的として子を生成した親個体にも生存の機会を与える戦略が採用されている.
IGS,SS
では親は無条件で残るのに対し,CHC,ER,MGGでは,子との競争に勝った親のみが生存を許される戦略である.世代交代の一般的な枠組みとして,
図
1.8
に示すような2
種類の選択,すなわち複製選択(Reproduction selection)と生 存選択(Survival selection)があり,CHC,ER,MGGのいずれも複製選択はランダ ムに非復元抽出であるが,生存選択がそれぞれ異なる.CHCの生存選択は,親子2
世 代の中から適応度(fitness)の高い順に集団サイズ分の個体を次世代に残すという方 法,ERは各家族(親2
個体,子2
個体)の中から最良2
個体を次世代に残すという方 法,MGGはER
を発展させたもので,各家族の中から最良1
個体とルーレット選択に より選ばれた1
個体を次世代に残すという方法である.それぞれの方法は問題により 向き不向きがあるとされている.それぞれの世代交代モデルの構成を表1.1
に,特徴 を表1.2
に示す.Childbirth Present
generation
Reproduction selection
Next generation Survival
selection Fitness
evaluation
Alternation-generation model
図
1.8:
世代交代モデルの概念図.これらの世代交代モデルでは,SGAがその構造がシンプルであることを理由に多用 されていたが,近年では多峰性の形状を持つ問題に対して性能の良い
MGG
が一般的 に多く用いられている.本研究では,第6
章において以下に示すMGG
を多親用に拡 張したモデル[67]
を用いることとした.「MGGを多親用に拡張したモデル」
( 1 )
個体集団から一定数の親個体をランダムに選ぶ.( 2 )
交叉によって一定数の子個体を生成する.表
1.1:
代表的な世代交代モデルの構成.モデル 複製時の選択 生存時の選択
SGA
ルーレットにより復元抽出 無条件で親集団と子集団を入れ換えSS
ランキングにより復元抽出 親集団の最悪個体と子個体を入れ換えCHC
ランダムに非復元抽出2
世代より最良個体から順に集団サイズ分を残す
IGS
ランダムに非復元抽出 親集団の適応度の低い個体と子個体を 入れ換えER
ランダムに非復元抽出 各家族から最良2
個体ずつを残すMGG
ランダムに非復元抽出 家族から最良1
個体とルーレットにより
1
個体を残す表
1.2:
代表的な世代交代モデルの特徴.モデル 適応度の使い方 世 代 交 代 の 連続化
世 代 交 代 の 限定化
生 存 選 択 の 局所化
SGA
基数的 離散的 全体的 なしSS
序数的 連続的 部分的 なしCHC
序数的 連続的 全体的 なしIGS
序数的 連続的 部分的 なしER
序数的 連続的 全体的 ありMGG
主に序数的 連続的 部分的 あり( 3 ) (1)
の親個体からランダムに2
個体を非復元抽出する.( 4 ) (3)
の2
個体と(2)
の子個体の中から最良個体および適応度のランキングによるルーレット選択で選んだ
1
個体を選び,個体集団に戻す.交叉オペレータの設計
喜多らは遺伝的アルゴリズムを構成する選択・世代交代モデル,交叉などの遺伝演算 子の設計に関して,「機能分担仮説」[68]「統計量の遺伝」[69]と呼ばれる考え方を提案 している.これは,遺伝演算子それぞれの担うべき機能を明確にし,その機能を考慮し た設計指針に基づいて遺伝演算子を設計するべきであるという考え方である.喜多ら が提案する設計指針をよく満たしている実数値遺伝的アルゴリズムの交叉手法の代表
1.1.
研究の背景11
的なものに,Unimodal Normal Distribution Crossover(UNDX)[70][71], UNDX-m[68],
Simplex Crossover(SPX)[72]
がある.表
1.3:
交叉オペレータの比較.形質遺伝 変 数 間 依存
座標軸方 向へ依存
スケール への依存
統 計 量 遺伝
子個体分布
バイナリ × × ? ? × ?
UNDX
○ ○ ○ × ○ 正規分布UNDX-m
○ ○ ○ △ ○ 正規分布SPX
○ ○ ○ ○ ○ 一様これらの実数値遺伝的アルゴリズムにおける代表的な
3
種類の交叉演算子およびバ イナリー遺伝的アルゴリズムの交叉演算子を比較をしたものが表1.3
である[72].機能
分担仮説および統計量の遺伝は,遺伝的アルゴリズムの設計の自由度をコントロール するための一つの仮説に過ぎないが,本研究ではこれらを有効な設計指針であると考 え,ここで述べた必要条件を全て満たすSPX
を第6
章での実数値遺伝的アルゴリズム において用いることとした.なお,SPXの詳細は付録A
に示す.また,本研究ではいくつか存在する遺伝的アルゴリズムの工学的応用例
[68][73]
の中 でも特に,シミュレーションを伴うために評価値計算に時間のかかる実問題に焦点を 当てる.具体的には,次に示すメカトロニクスおよびパワーエレクトロニクスの分野 における実問題を取り上げる.工学的応用問題
メカトロニクスやパワーエレクトロニクスの分野では,各種設計パラメータの試行 錯誤的で煩雑な決定作業を軽減するために,遺伝的アルゴリズムの最適化能力に着目 した自律パラメータ設計に関する研究が報告されている
[74][75][76].その中で,構造
設計段階で形状寸法が定められたSwitched Reluctance Motor(SRM)に対し,その
最大出力性能と対応する最適制御パラメータを遺伝的アルゴリズムを用いて探索する 研究が行なわれている[77][78].探索に遺伝的アルゴリズムを適用することで他手法と
比較して大幅な計算時間削減を実現しているが,出力性能評価のための反復計算を伴 う特性計算によって1
回の適応度計算に時間がかかるため,依然として計算時間削減 の余地が残っている.遺伝子表現,適応度関数等,詳細については付録B
にて述べる.以上に述べた世代交代モデルおよび交叉オペレータの設計指針に基づき,本研究で は,実数値遺伝的アルゴリズムの効率化のための新たな進化オペレータを提案し,
SRM
制御パラメータ最適化問題に適用し評価する.1.2
研究の目的今日,人工生命研究の目的は二つに大別することができる
[9][79].一つは,生物を
分析,模倣することによって,生命の本質を理解する,あるいは生物システムの挙動 や進化のメカニズムを解明することである.もう一つは,生物システムをモデル化し,そのモデルに基づいた工学的な実際のシステムを構築することである
[9][80].本論文
の第2
章から第5
章までの人工生命は前者を目的とし,本論文の第6
章は後者を目的 としている.人工生命の生物学的応用として,本研究では実在する各種生命体の生態現象,具体 的には,食物連鎖,種分化,性選択についてモデル化を行い,計算機上で人工生命体が 実在する生命体の諸現象を進化により創発する様子を確認する.そうすることで,生 命の誕生,生態分化,生態進化システムなどのメカニズムを明らかにすることが最大 の目的である.ここで,1.1節において述べたように実在する生命体の挙動を正確に再 現することが人工生命の目的ではないことに注意したい.さらに,各種人工生命体の 構成のための計算手法を提案することで人工生命研究・生物学研究への工学的一助と なることを目指す.
人工生命の工学的応用として代表的なものが進化的計算である.進化的計算は,そ の膨大な計算負荷などの問題のために実用化は難しいとされていた.しかし,近年の 計算機の処理能力の向上やコストの軽減などの理由で計算量の問題は解消されつつあ るため,アルゴリズム上の探索効率を上げることで工学的応用に耐えうる進化的計算 手法の提案を目指す.
第
2
章では,より現実に近い安定した人工生命モデルの生成が目的である.不安定な 環境の中で自己適応していくシステムの研究として,微妙なバランスの上に成り立っ ている生態系の計算機シミュレーションがある.生態系のシミュレーションではより 現実に近くかつ単純でシミュレートが容易なモデルを構築し,生物の諸現象を計算機 上で調べることが課題となる.ここでは,生命体が世代交代をしていく上で欠かすこ との出来ない生殖活動に着目し,交配を導入することでより現実に近いモデルを生成1.3.
研究の内容13
する.これにより長く安定したシミュレーションを確保することが目的である.
第
3
章では,種の多様性を引き起こす生態の分化がどのようなメカニズムで生じる のかを解明することが目的である.自然界に存在する生命体は進化の過程で様々な生 態を発現し,その種類も多様化している.ここでは,同所的種分化がどのようなプロ セスで発生したかを明らかにする.第
4
章,第5
章では,性選択が存在する環境下での遺伝子とミームが及ぼす相互作 用について解明することが目的である.それぞれの章において,人工生命体に同調性・非同調性の特徴を付け,その特徴によって異なる行動を行なうことで,配偶者選択に おける嗜好の遷移に与える影響を明らかにする.
第
6
章では,進化的計算手法の工学的応用,特に計算負荷の高いシミュレーション を伴う実問題への適用が目的である.遺伝的アルゴリズムを用いた探索において,探 索局面は常に動的に変化をする.よって,探索過程を常に評価してそれにあった戦略 を用いて探索をすすめることは重要だと考える.ここでは,探索過程を考慮に入れた 実数値遺伝的アルゴリズムを提案しアルゴリズム上の探索効率を上げることで工学的 応用に耐えうる計算手法の実現を目指す.1.3
研究の内容本研究は,全体の構成として大きく
2
つに分けられる.第2
章から第5
章は,人工生 命の生物学的応用についての研究である.特に,第2
章および第3
章の前半では,人 工生命を計算機上に実装するために必要となる計算手法を提案し,それらの章の後半 および第4
章,第5
章において,人工生命を用いた各種生態進化モデルを計算機上で 構成した結果について考察を行う.第6
章は,人工生命の工学的応用についての研究 である.ここでは,実数値遺伝的アルゴリズムの効率化を行う交叉手法を提案し,計 算機上に実装した結果について述べる.始めに,第
2
章では,人工生命を表現するための手段として多出力二分決定グラフ(n-output Binary Decision Diagram;n-BDD)
の交叉手法を提案する.被食,捕食関係 のある多種類の人工生命体をn-BDD
を用いて計算機上に存在させ,不安定な環境の 中で自然淘汰による進化を繰り返し自己適応させる人工生命モデルを提案する.ここ では,エージェントが環境の中で他のエージェントと相互に干渉しながら生態系全体 で安定した食物連鎖関係を創発することを示す.第
3
章では,変数順序の違うn-BDD
同士の交叉を可能とした交叉手法を提案する.この交叉手法を用いて,身体的特徴差を持つ各々の人工生命体が,環境変化に順応し て異なる生命体に分化していく様子を計算機上で表現する.本システムにより,似た 遺伝子を持った人工生命体がわずかな身体的特徴差(環境順応値)により,異なった 方向へ互いに進化(分化)していくことを明らかにする.
第
4
章では,実在する一部の動物に見られる配偶者選択におけるメスの嗜好の流行 現象がメスの嗜好ミームおよび同調性・非同調性に関与していると考え,遺伝子とミー ムをあわせもつ人工生命体の進化モデルに同調遺伝子を加えたモデルを提案する.同 調遺伝子により模倣行動または独創行動を行い,世代交代により進化することで,配 偶者選択における嗜好の循環型流行の発現を確認する.第
5
章では,第4
章の同調遺伝子に代わり同調・差別化欲求の強さを表す遺伝子を 新たに定義し,さらに同調化行動および差別化行動を行うエージェントモデルを提案 し,計算機シミュレーションを行う.実験において,オスを好むメスの嗜好に2
種類 の循環型流行の発現を確認する.また,同調・差別化欲求の存在がこれらの流行現象 に与える影響について考察する.第
6
章では,実数値遺伝的アルゴリズムにおいて,限られた生成子個体数での効率 的な探索を行うモデルを目指し動的多段交叉を提案する.動的多段交叉は,評価値の 良い個体を用いて交叉を段階的に行うことで進化を促し,その段数を探索過程に応じ て動的に変化させる交叉法である.動的多段交叉を代表的な世代交代モデルに適用す ることで,最良個体の進化を促しつつ集団の多様性を維持できるモデルを実現したこ とを示す.最後に,第
7
章において,本論文を総括し,今後の研究の展望について述べる.15
第 2 章
多出力二分決定グラフの APPLY 交叉を 用いた食物連鎖モデル
本章では,多出力二分決定グラフ
(n-output Binary Decision Diagram ;n-BDD)
を用い た生態系モデルにおいて利用可能な交配方法を提案する.n-BDD
は,人工生命の行動 戦略の表現に適しているが,遺伝子操作に交叉の機能が存在しない.そこで,n-BDD
の 交叉方法としてBDD
の二項論理演算を拡張したAPPLY
交叉を定義し用いる.APPLY
交叉は両親の行動戦略を確率的に均等に継承できることに特徴があり,エージェント の交配に用いることに適していると考えられる.簡単な競合問題を用いてAPPLY
交 叉の有効性を確認し,さらに,被食,捕食関係のある多種類の人工生命体を存在させ 交配による世代交代を繰り返すことで,他の個体と相互に干渉しながら生態系全体で 安定した食物連鎖関係が創発した.2.1
はじめに生命的な振る舞いにヒントを得た手法の研究あるいは生命に関わる諸現象を工学的 手法で再現し解明しようとする研究が様々な分野で展開されている.不安定な環境の 中で自己適応していくシステムの研究として,微妙なバランスの上に成り立っている 生態系のシミュレーションはその中の一つである.
Holland
によって最初に考案された生態系モデルEcho
は,全ての生態系に共通の一般的な性質の獲得のために,生態系を可能な限り単純化している
[5][81][82].高階ら
は,個々の生命体を独立のプログラムで記述した生態系のシミュレーションを行った[83][84].我々は,自然界に存在する生命体を 3
種類に単純化した生態系において,被食者,捕食者の行動戦略を
n-BDD
を用いて表現し,シミュレートする研究を行ってきた
[19][20].n-BDD
は文献[19][20]
が示すように入力情報に対して行動を一つに決定す る問題に適しており,生態系シミュレーションの実現のために有用である.生態系の計算機シミュレーションではより現実に近くかつ単純でシミュレートが容 易なモデルを構築し,生物の諸現象を計算機上で調べることが課題となる.本章は,
生命体が世代交代をしていく上で欠かすことの出来ない生殖活動に着目し,交配を導 入することでより現実に近いモデルを生成する.これにより長く安定したシミュレー ションを確保することが目的である.
有性生殖における交配には,交配相手の選択とその交配方法の
2
点が重要となり,こ れらに関しては今までに以下に示すような様々なモデルが提案されている.交配相手 の選択に関しては,よく似た個体のみを交配相手とするモデル[85]
や,逆に似た個体 同士の交配(近親婚)
を禁じたモデル[86]
や,交配タグの一致した個体だけが相手とな るモデル[5]
などがある.また,Hillisは初期集団を2
次元格子状に配置させ,空間に おいて近傍の個体同士だけが交配を行うモデルを発案した[87].生態系モデル Echo
に おける交配は,各エージェントが持っている染色体を2
点交叉で組み合わせて2
匹の 子を形成しており,Hillisは二倍体染色体を用いたより実世界に近い交配方法を提案し ている.本研究では,この中から
n-BDD
を用いた生態系モデル[19]
において利用可能な交 配方法を利用し拡張する.交配相手の選択には,[5][85]のモデルで提案された異種間 の交配を禁止する手法を採用し,また[87]
において提案された空間的に制限された相 手との交配をさらに拡張し,その中から評価値の高いエージェントと交配することと した.交配方法はBDD
の二項論理演算[88]
を拡張したAPPLY
交叉を定義し用いる.APPLY
交叉はn-BDD
のグラフ構造を利用しながら両親の行動戦略を確率的に均等に継承できることに特徴があり,n-BDDを用いた本モデルにおけるエージェントの交配 に用いることに適していると考えられる.
2.2 BDD
とAPPLY
演算2.2.1 BDD
1978
年にAkers[29]
によって考案されたBDD
は論理関数の表現方法の一つであり,その記憶効率や処理速度の面での優秀さから
LSI
やCAD
の分野を中心に,様々な分 野に応用されている.2.2. BDD
とAPPLY
演算17
false or true
1 0
x1
x2 x3
(a)
0 0 - -
1 1 - -
(a) x1 c b (b) x2 1 0 (c) x3 0 1
(b)
図2.1: BDD
のグラフ表現(a)
とデータ構造(b).
図
2.1(a)
にBDD
の一例を示す.図2.1(a)
の丸で表される節点は変数節点である.変数節点は
0
枝と1
枝の2
種類の枝を持つ.BDDの変数節点が持つ変数の値を1
つに定 めることで,1つの定数節点を対応させることができる.すなわち,根の節点から始 めて変数節点に書かれた変数の値が0
の時は0
枝をたどり,1のときは1
枝をたどる.こうして最終的にたどり着く四角で表される節点に書かれた値が出力値である.四角 で表される定数節点は
0
か1
の2
通りの値を持ち,これによってBDD
は論理関数を表 現する.BDD
の節点は変数記号(または定数値),0枝につながる節点ラベル,1枝につな がる節点ラベルの3
つの属性を組にして持ち,この組の集合を保持し,その内1
つを 根節点(BDDf
に対し,f.topと表記)として指定することで1
つのBDD
を表すこと ができる.本論文では,BDDf
に対し,f.topの節点の0
枝,1枝に続くグラフをf
0,f
1と書くことにする.つまり,f.top = (x
i, f
0, f
1) (2.1)
と定義される.f0,f1は
0
枝,1枝につづくサブグラフだが,そのサブグラフが表現 する論理式と同一視する.図2.1(b)
にBDD
をこの表現で表した例を示す.根節点から葉への順に現われる変数の順序(変数順位)を一定にすることで,
BDD
に 関する計算効率が良くなる.本研究においても与えられた変数順位に従うこととする.2.2.2 APPLY
演算APPLY
演算はBryant[88]
が考案したBDD
のための二項論理演算の計算手続きであり,二つの論理関数
f
,gのBDD
に対し二項論理演算(AND,ORなど)の結果を表すBDD
を生成する.二項論理演算(◦)は次のように定義される.f ◦ g(x
1, x
2, . . . , x
k) = f (x
1, x
2, . . . , x
k) ◦ g(x
1, x
2, . . . , x
k) (2.2)
この二項論理演算はShannon
の展開式により次のように表される.f ◦ g(x
1, . . . , x
i, . . . , x
k) =
f (x
1, . . . , x
i−1, 0, x
i+1, . . . , x
k)
◦ g(x
1, . . . , x
i−1, 0, x
i+1, . . . , x
k); if x
i= 0 f (x
1, . . . , x
i−1, 1, x
i+1, . . . , x
k)
◦ g(x
1, . . . , x
i−1, 1, x
i+1, . . . , x
k); if x
i= 1
(2.3)
ここで図
2.1
で与えたデータ構造によってBDD
が表現されるとする.式(2.3)
にお いて,f.top= (x
i, f
0, f
1)
ならばf (x
0, . . . , x
i−1, 0, x
i+1, . . . , x
k) = f
0,f (x
0, . . . , x
i−1, 1, x
i+1, . . . , x
k) = f
1である.したがって,BDDの上位の変数から順に展開して,それぞれの部分グラフ同士の演 算を再帰的に実行し,定数値に関する自明な演算になったところで,再帰を打ち切る ことで結果を得ることができる.この方法で
h(= f ◦ g)
を求めるアルゴリズムを図2.2
に示す.( 1 ) f, g
のいずれかが定数のとき,演算子◦
に応じ,f◦ g
を求める.( 2 ) f.top
とg.top
の順位が同じとき,h
0← f
0◦ g
0, h
1← f
1◦ g
1if h
0= h
1then h ← h
0else h ← (f.top, h
0, h
1) ( 3 ) f.top
がg.top
より上位のとき,h
0← f
0◦ g, h
1← f
1◦ g
( 4 ) f.top
がg.top
より下位のとき,h
0← f ◦ g
0, h
1← f ◦ g
0図
2.2: h(= f ◦ g)
のアルゴリズム.このアルゴリズムでは,生成される各
![表 2.2: エージェントが選択できる行動. walk (W) ランダムに 8 近傍に移動する runaway (R) 肉食エージェントから遠ざかる 方向へ移動する eat (E) 食物の方向へ移動して食べる do-nothing (N) 動かない approach (A) 仲間に接近する 各行動の詳細は [20] と同じ 返されているのが分かる.また,捕食者-被食者の個体群モデルを表す Lotka-Volterra 方程式 [89] を仮定すると食物連鎖関係が成立した生態系で横軸に捕食者数,縦軸に被 食者](https://thumb-ap.123doks.com/thumbv2/123deta/6463149.2149771/41.892.264.579.659.975/エージェント選択できるランダムエージェント食べる分かるモデル.webp)