1994
性 腺 刺 激 ホル モ ン添 加 が ブ タ卵 細 胞 の成 熟 ・受精 と
3H ー メ チ オ ニ ンの取 り込 み に及 ぼ す影 響 につ い て
Effects of Gonadotrophins on the In vitro Maturation
and Fertilization of Pig Oocytes
辻 井 弘 忠 ・上 田 大 ・康 承 律
Hirotada TSUJII, Masaru UEDA, Seung-Yul KANG
信 州大学農学部 生物資源 開発学講座 〒399-45長 野県上 伊那郡
Faculty of Agriculture, Shinshu University, Nagano-Ken 399-45
Abstract: This study was conducted to determine the effect of PMSG and/or hCG (each 15 IU/ml) added to TCM -199 supplemented with 10% FCS and 3mg/m1
BSA on the maturation, fertilization and the incorporation of 3H - methionine of pig oocytes. The follicular oocytes were cultured for 48hr with or without supplementation of PMSG, hCG, given each alone or in combination to maturation medium
. The addition of PMSG with or without hCG to maturation medium was effective in inducting meiotic resumption in pig oocytes . Fertilization rate was significantly improved by addition of hCG alone or with PMSG
, compared to the control. From these results, it is suggested that the sperm penetration rate of oocyte was increased by hCG added to the maturation medium . The proportion of polyspermic oocytes
, however, was not different among the treatments
. Incorporation of methionine into oocytes measured after maturation culture was increased by addition of PMSG and/or hCG as compared to the control. These results indicate that PMSG and/or hCG added to medium during maturation is related to the development of cytoplasm of pig oocytes. Key Words: pig oocyte
, maturation, IVF, gonadotrophin (1994”N2ŒŽ18“úŽó•t1994”N5ŒŽ13“úŽó—•)
緒
言
体 外 で の 卵 母 細 胞 の成 熟 培 養(IVM)で は,第2減 数 分 裂 中期 に 達 す る と と もに,受 精 能 お よ び発 生 能 を もつ卵 子 が得 られ るか 否 か が課 題 と な る.そ の た め に はinvivoに お い て 減 数 分裂 再 開 に関 わ って い るFSHとLH,あ る い は,こ れ らFSHとLH様 作 用 を有 す るPMSG, hCGの 培 養 液 へ の 添 加 が そ の 成熟 に必 要 で あ る と考 え られ る.牛 の 卵 胞 卵 子 の 成 熟 は,TCM-199培 養 液 に10%前 後 の 血 清 を 加 え た 培 養 液 で 成 熟 卵 子 を得 て い る報 告1)も あ る が,性 腺 刺 激 ホル モ ンを添 加 して い る例 も見 られ る2,3).さ らに,成 熟 用 培養 液 に性 腺 刺 激 ホル モ ン を添 加 す る こ と に よ り,第2減 数 分 裂 中期 に達 す る こ とに加 えて,受 精 能,発 生 能 を 含 ん だ総 合 的 な 卵 子 の能 力 が 高 め られ る こ と が,マ ウ ス4,5),ラ ッ ト6,7,8),山羊9,10),羊11,12,13)およ び 牛14,15,16)に お いて 報 告 され て い る. 一 方,豚 卵 母 細 胞 のIVMに お い て も一 般 的 に成 熟 用 培養 液 に性 腺 刺 激 ホ ル モ ン が添 加 され て お り17,18,P),FSHとLHの 添 加 が卵 子 の成 熟 を 高 め る こ とをMattioliら20)が 報 告 して い る.し か し,FSH,LH様 作 用 を も つPMSG,hCGに 関 して は,近 年一 般 的 に成 熟 用 培 養 液 に添 加 され て は い る もの の,そ れ ら 自体 が体 外 成 熟 ・受 精 ・培 養 の過 程 に どの よ うに影 響 し て い るの か詳 細 に調 べ た報 告 は少 な い.そ こで 本 実 験 で は,成 熟 用 培 養 液 に 添 加 したPMSG とhCGが,卵 母 細 胞 の 第2減 数 分 裂 中 期 へ の成 熟 と受 精,多 精 子 侵 入 お よ び ア ミノ酸 の 取 り 込 み に与 え る影 響 に つ い て 調 べ る こ と を 目的 と した. 材 料 お よび 方 法 卵 母 細 胞 の 採 取:食 肉 セ ン ター に て,性 成 熟 豚 か ら黄 体 の な い卵 巣 の み を採 取 し,39℃ に保 温 した0.85%生 理 食 塩 水(100mg/1の 硫 酸 カ ナマ イ シ ンを 含 む)に 浸 した 状 態 で 研 究 室 に持 ち帰 り, 38℃ の 生 理 食 塩 水 で3回 洗 浄 した 後,直 径2-5mmの 卵 胞 の 内 容 物 を18G針 付 デ ィ ス ポ ー ザ ブ ル シ リン ジで 吸 引採 取 した.緊 密 な数 層 の 卵 丘 細 胞 に 包 まれ,黒 く均 一 な細 胞 質 を もつ卵 母 細 胞 の み を実 験 に供 した.培 養 液 は,TCM-199に10%FCSと3mg/mlBSA(ナ カ ラ イ テ ス ク株 式 会 社)を 添 加 した もの を 基 礎 培 養 液(BM)と し,こ れ に15IU/mlPMSG(ピ ー メ ッ ク ス,三 共 ゾ ー キ)と15IU/mlhCG(プ ベ ロー ゲ ン,三 共 ゾー キ)を 添 加 す る組 み 合 わせ に よ って以 下 の5区 の 実 験 区 を 設 け た.[1]BMの み(Control区),[2]BM+PMSG(+ PMSG区),[3]BM+hCG(+hCG区),[4]BM+PMSG+hCG(+PMSG,+hCG区), [5] PMSG添 加 培 養 液 で24時 間 培 養 後,hCG添 加 培 養 液 で24時 間 培 養(PMSG→hCG区) . 流 動 パ ラ フ ィ ン下 に 各 実 験 区 の培 養 液 ドロ ッ プ(0.4ml)を 作 り,卵 母 細 胞 を20-30個 ず つ 導 入 し,37℃,5%CO2,95%空 気 の 条 件 下 で48時 間 成 熟 培 養 を 行 っ た. 凍 結 精 子 の 処 理 と前 培 養:体 外 受 精 に供 した 精 子 は 長 野 県 畜 産 試 験 場 の ラ ン ドレー ス雄 の 射 出 凍 結 精 液 を使 用 した.精 子 の 洗 浄,前 培 養 お よび 受 精 用 の 培 養 液 に はBO液(5mMカ フ ェ イ ン と3mg/mlBSAを 含 む)を 用 い た.液 体 窒 素 中 か ら取 り出 した 凍 結 精 液 を ス トロ ー(精 液0.5 ml/本)ご と40℃ の湯 に50-60秒 間 漬 け て 融 解 した.さ ら に 遠 沈 管 に移 し,あ ら か じめ38℃ に温 め て お い たBO液1.0mlを 加 え 十 分 撹拌 後,500gで5分 間 遠 心 分 離 した.上 清 液 を捨 て, 同様 の操 作 で2度 洗 浄 し,2×108/mlの 濃 度 に調 整 し,37℃,5%CO2,95%空 気 の 条 件 下 で 3時 間 前 培 養 を行 った 。 体 外 受 精 とそ の後 の 培 養:流 動 パ ラ フ ィ ン下 に 用 意 したBO液 の ドロ ップ(0.4ml)に 成 熟 培 養 した 卵 母 細 胞20-30個 を 移 し,前 培 養 を終 了 した精 子 を2×106/mlの 濃 度 に な る よ うに 導 入 し
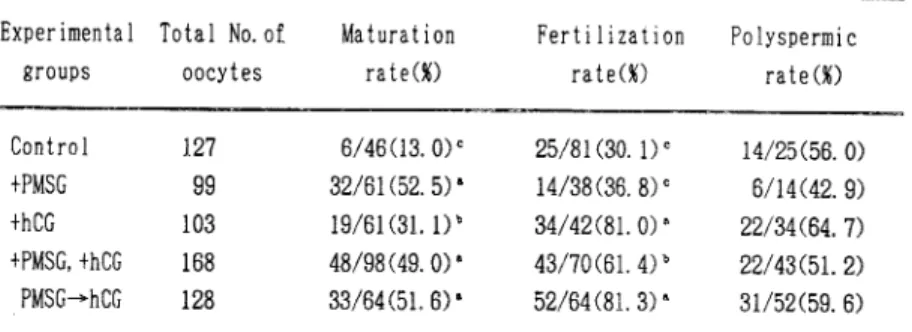
て媒 精 を行 った。 媒 精 後7-8時 間 目に卵 子 を取 り出 し,ピ ペ ッテ ィン グ に よ り周 囲 に付 着 して い る精 子 を取 り除 い た後 に10%FCSを 加 え たTCM-199の ドロ ップ へ移 し ,媒 精 後18-20時 間 目ま で培 養 した. 成 熟 率 と受 精 率,多 精 子 侵 入 率 の算 定:成 熟 培 養 終 了後 の卵 子 の 一 部 は カル ノ ア液(メ タ ノー ル:酢 酸=3:1)で 脱 脂 ・固 定 し,成 熟 率 の判 定 の た め の 観 察 に 供 した.ま た,媒 精後18か ら 20時 間 目の 卵 子 は 同様 に 固 定 し,受 精 率 お よび 多 精 子 侵 入 率 の 算 定 の た め の 観 察 に供 した 。固 定 は いず れ も3-5日 間 行 い,1%ア セ トオ ル セ イ ソで 染 色 した 後 に ノマ ル ス キ ー 微 分 干 渉 装 置 付 きの 倒 立 顕 微 鏡 を用 い観 察 を行 っ た . ア ミ ノ酸 の取 り込み:卵 胞 か らの採 取 直 後 のGV期 お よ び,成 熟 培 養 直 後,顕 微 鏡 下 で極 体 の 放 出 の観 察 され たMII期 の卵 母 細 胞 を選 び ,ア ミノ酸取 り込 みの実験 に供す るため ピペ ッテ ィ ン グ に よ り卵 丘 細 胞 を と り除 い た .5μCi/mlの3H-メ チ オ ニ ン を100μl入 れ た0 .5ml容 マ イ ク ロチ ュ ー ブ に,1サ ン プ ル 当 た り10個 の卵 子 を 入 れ,さ らにBO液 を加 え合 計200μl に な る よ う に定 容 した.マ イ ク ロ チ ュ ー ブを37℃ の イ ソ キ ュ ベ ー タ ー 内 に 入 れ1時 間培 養 した 後 に,10%TCAを 等 量(200μl)加 え反 応 を停 止 させ た.各 サ ン プ ル は-20℃ の ス トッカ ー で 保 存 後,ミ リポ ア フ ィル ター 上 で吸 引 濾 過 し,5%TCAと70%エ タ ノー ル で 洗 浄 しタ ンパ ク分 画 と した.フ ィル ター を乾 燥 させ た後,液 体 シ ソ チ レー シ ョ ソカ ウ ン ター で カ ウ ン トした.単 位 はcpm/oocyteで 表 した. 統 計 分 析:成 熟 率,受 精 率,多 精 子 侵 入 率 はx2検 定 で,ア ミノ酸 取 り込 み量 の データ はt検 定 に よ り比 較 し,不 等 分 散 で あ る もの はCochran-Cox検 定 を用 い て 比 較 した . 結 果 各 実験 区 で48時 間 の 成 熟 培 養 を行 っ た卵 母 細 胞 の成 熟 率 な ら び に 受 精 率 を表1に 示 した.成 熟 率 は,Control区 に比 べ他 の 実 験 区 で は 有 意 に 高 か っ た(P<0.05).ま た,+hCG区 に比 べ , +PMSG区,+PMSG,+hCG区,PMSG→hCG区 は有 意 に高 い成 熟 率 を示 した(P<0 .05).
Table 1:Effect of PMSG and/or hCG added to maturation medium on in vitro maturation and fertilization of pig oocytes
°_cValues with different superscripts are significantly different(P<0 .05). +PMSG,+hCG : Addition of PMSG and/or hCG in maturation medium.
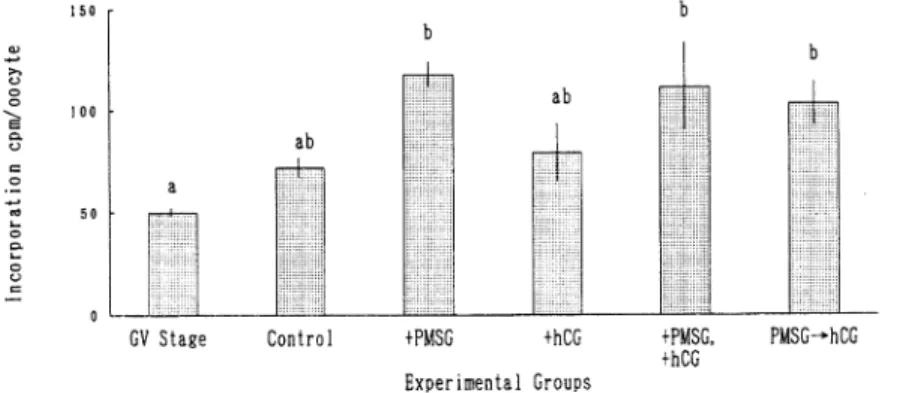
受 精 率 は,Control区 に比 べ て+PMSG区 で は 有 意 な 差 は見 られ な か っ た が,+hCG区, +PMSG,+hCG区,PMSG→hCG区 で は 有 意 に 高 い受 精 率 が得 られ た(P<0.05).ま た,+ PMSG,+hCG区 に比 べ て+hCG区 とPMSG→hCG区 の 受 精 率 は有 意 に高 か っ た(P<0.05). また,受 精 卵 に お け る 多精 子 侵 入 率 につ い て は,各 実 験 区 の 間 に有 意 な差 は 見 られ な か った. 各 実 験 区 の 成 熟 培 養 後 の ア ミノ酸 の 取 り込 み 量(平 均 値 ± 標 準 誤 差)を 図1に 示 した.そ の 結 果,GV期 の卵 子 に比 べ,48時 間 成 熟 培 養 後 のMII期 卵 子 に お い て,全 て の実 験 区 で ア ミ ノ 酸 の取 り込 み 量 は増 加 す る傾 向 が 見 られ,+PMSG区 で は 有 意 に 高 い値 で あ っ た(P<0.05). また,Control区 に比 べ+PMSG区 で は 取 り込 み 量 は 有 意 に 高 か っ た(P<0.05).
Figure 1: Incorporation of 3H-methionine into oocytes matured in addition of PMSG and/or hCG.
( M•}S.D,a,b;p<0.01)
+PMSG,+hCG : Addition of PMSG and/or hCG in maturation medium. PMSG•¨hCG : Addition of hCG followed by PMSG in maturation medium.
考 察 Mattioliら20)は 性 腺 刺 激 ホ ル モ ン を含 まな い 培 養 液 で豚 の 卵 母 細 胞 を 成 熟 させ た 時 の 成 熟 率 (35%)に 比 べ,FSH及 びLHを 添 加 した培 養 液 で は成 熟 率 が86%,76%と 向 上 した と報 告 して い る.本 実 験 に お け る成 熟 率 はControl区 に比 べ,PMSG,hCGを 添 加 した 他 の どの 実 験 区 に お い て も有 意 に 高 ま り,PMSG及 びhCGが 豚 卵 母 細 胞 の減 数 分 裂 再 開 始 誘 起 に 有 効 で あ る こ と が示 され た.ま た,+PMSG,+hCG区,+PMSG区,PMSG→hCG区 に比 べ, +hCGで 有 意 に成 熟 率 が低 か った.こ れ は,in vivoに お い てLHサ ージ に よ って 減 数 分 裂 の 再 開 が 起 こ る こ と を 考 え る と,や や 矛 盾 して い る よ う に 思 わ れ る.こ の こ と に 関 して は, Galliら11)が,PMSGを 投 与 した 羊 の 卵 巣 卵 母 細 胞 を採 取 し,hMG(human menopausal gonadotropin)を 添 加 した培 養 液 で成 熟 培 養 す る こ と に よ って 第2減 数 分 裂 中期 卵 子83.8% を得 て い る こ とや,ま た,in vivoに お い て 卵 母 細 胞 はLHサ ージ の作 用 を受 け る前 にFSH の作 用 を受 け て お り,さ らに,排 卵 の 前 後 に はFSHサ ージが 発 現 し,血 中FSHレ ベ ル も上 昇 す る こ と か ら,卵 母 細 胞 の 成 熟 に はLHま た はhCGに よ る減 数 分 裂 再 開 の 誘 起 作 用 を 受 け る前 にFSHやPMSGの 作 用 を 受 け る必 要 が あ る.こ れ は卵 丘 細 胞 に お け る減 数 分 裂 分 裂 再 開 の誘 起 因 子21)にFSHやPMSGが 作 用 す る必 要 が あ る の で は な い か と思 わ れ た.
一 方,PMSGとhCGの 両 方 の作 用 を受 け た+PMSG ,+hCG区 お よ びPMSG→hCG区 で は成 熟 率 は高 く,PMSG ,hCGの 相 互 作 用 が 核 成 熟 に 必 要 で あ る と示 唆 され た.ま た+P MSG区 に お い て も+PMSG,+hCG区,PMSG→hCG区 と同様 に高 い 成 熟 率 が 得 られ た.P MSGはFSHと 同様 の生 物 学 的 作 用 に 加 え て弱 い な が ら もLH様 作 用 も有 す る こ とか ら ,結 局+PMSG,+hCG区 と同 様 の効 果 が得 られ た の で は な い か と推 察 され た. Eppigら4)はFSHを 添 加 した成 熟 用 培 養 液 で 成 熟 させ た マ ウ スの 卵 母 細 胞 は ,体 外 受 精 後 の2細 胞 あ るい は 胚 盤 胞 ま で の 発 生 率 が 高 ま る と報 告 して い る .ま たZhangら6)はPMSGを 投 与 した ラ ッ トか ら得 た 卵 母 細 胞 お よび,FSHを 添 加 した 成 熟 用 培 養 液 で 成 熟 させ た ラ ッ ト 卵 母 細 胞 で 高 い受 精 率 を 得 た と報 告 して い る.こ れ ら は卵 母 細 胞 の 受 精 能 に 関 してFSHが 有 用 で あ る こ と を示 して い る.一 方,豚 に 関 して はMattioli20)お よ びNaitoら22)の 報 告 の 中 で,FSH添 加 培 養 液 で 成 熟 させ た卵 母 細 胞 の 受 精 率 の 効 果 が あ る が ,性 腺 刺 激 ホ ル モ ン無 添 加 区 を含 む 他 の ど の実 験 区 と も有 意 な差 は な く,高 い受精 率 を得 てい る.本 実験 では,+PM SG区 の 受 精 率 は低 く,む しろ,hCGの 添 加 が受 精 率 の 向上 に必 要 で あ る と 推 察 され た. 多 精 子 侵 入 率 は各 実 験 区 の 間 で 有 意 な 差 は な か った .豚 は多精 子侵入 が起 こ り易 い種 で,人 工 授 精 で 卵 管 内 に 多数 の 精 子 を注 入 す る と多 精 子 侵 入 率 が極 端 に 高 くな る と言 わ れ て い る23). そ の た め一 度 に多 数 の精 子 が 周 りに存 在 す る体 外 受 精 で は 多 精 子 侵 入 が 頻 繁 に起 こ り ,そ の 後 の 発 生 率 が 極 端 に低 くな る と 言 わ れ て い る24).in vivoで は,多 精 子 侵 入 を 拒 否 す る た め に , 卵 子 は一 精 子 の侵 入 刺 激 に よ っ て透 明 帯 反 応 を起 こす が,こ れは卵子細胞 質の作用 が強 く関係 して い る.本 実 験 で は 全 実 験 区 の卵 子 と も同 様 な率 で 多 精 子 侵 入 が 起 こ っ て お り ,細 胞 質 の 成 熟 度 合 い は 各 実 験 区 間 で 同 様 で あ っ た と 考 え られ る.ま た,Yoshidaら25)の 体 内成 熟 卵 子 を 用 い た体 外 受 精 率 の結 果 に よ る と,卵 子 を提 供 す る雌 個 体 に よ って も多 精 子 侵 入 率 に差 が あ る こ とか ら,本 実 験 で の 多 精 子 侵 入 率 の 高 低 につ い て考 察 す る こ とは 難 し く,従 って 細 胞 質 の 成 熟 が充 分 で あ る か ど うか は 他 の 方 法 に よ って 検 討 しな け れ ば な らな い.一 方,多 精 子 侵 入 に関 す る精 子側 の 問 題 と して,Nagaiら26)は 精 子 前 培 養 濃 度 に違 い よ っ て 受 精 率 が 変 わ り,受 精 率 が 向上 す る と と もに 多精 子 侵 入 率 が増 加 す る と報 告 し,Rathら24)は 媒 精 時 に お け る一 卵 子 あ た りの 精 子 数 が 増 加 す る と多 精 子 侵 入 率 も増 加 す る と報 告 して い る.従 って,多 精 子 侵 入 を 防 ぐには 精 子 側 の 処 理 に つ い て も検 討 す べ きで あ る と考 え られ た. 3H-メ チ オ ニ ン の取 り込 み は ,物 質 代 謝,特 に タ ンパ ク合 成 の 指 標 と な り得 る もの で あ り, 卵 母 細 胞 や 胚 の 生存 力 ひ い て は核 や 細 胞 質 の成 熟 と非 常 に 関 係 が深 い と考 え られ る.加 え て雄 胚 の グル コース代 謝 は 高 い27)と され て い る.ま た,マ ウス 胚 で は潜 在 的 活 性 が 高 い胚 は,代 謝 率 を 高 め27),さ ら に異 常 発 生 や 生 存 率 低 下 を防 ぐ29)と い う報 告 もあ る.こ れ らの こ と か ら, ア ミ ノ酸 等 の代 謝 活 性 を調 べ る こ とで 卵 子 の生 存 力 の高 低 を あ る程 度 推 察 し得 る と考 え られ る. 本 実 験 の 結 果,核 の成 熟 が 進 ん だ卵 子 ほ ど,生 存 活 性 が高 く,ま た 細 胞 質 の 成熟 も進 んで い る と考 え られ た.今 後,受 精 前 後 の透 明 帯 の変 化 な ど を的 確 に測 定 で き る方 法 等 を 検 討 す る こ に よ っ て,多 精 子 侵 入 防 御 の 機 構 が解 明 され る と思 わ れ る.
要 約 本 実 験 はTCM-199に10%FCSと3mg/mlBSAを 添 加 した 基 礎 培 地 にPMSGとhCG 添 加 が 豚 卵 母 細 胞 の 成 熟,受 精 お よ び3Hー メ チ オ ニ ン の取 り込 み に及 ぼ す 影 響 に つ い て調 べ た.実 験 区 は,(1)基 礎 培 地,(2)PMSG(15IU/ml)添 加,(3)hCG(15IU/ml)添 加,(4)PMSG+h CG(+P,h)添 加 お よ び(5)PMSG24時 間 後,hCG24時 間(P→h)の5区 を 設 け,48時 間 培 養 を行 った.こ の 結 果,PMSGGお よ びhCGを 基 礎 培 養 液 に添 加 す る こ とは,豚 卵 母 細 胞 の 減 数 分 裂 再 開 誘 起 に有 効 で あ った.受 精 率 は 対 照 に 比 べ,+hCG区,+P,h区,P→h 区 で 有 意 に 高 く,hCGの 添 加 が 受 精 率 の 向 上 を 促 進 す る こ と が示 され た.し か し,多 精 子 侵 入 率 に つ い て,実 験 区 間 で 差 は 見 られ な か っ た.上 記5区 の 成 熟 培 養 後 の3Hー メ チ オ ニ ン の 取 り込 み を 測 定 した 結 果,対 照 区 よ りPMSG,hCG添 加 した 区 で 高 くな る傾 向 が 見 られ, PMSGお よびhCG添 加 が,卵 母 細 胞 の 細 胞 質 の 成 熟 に 関与 して い る こ とが 明 ら か に な った. 謝 辞 豚凍 結精液 を提供下 さい ま した長野 県畜 産試験場養 豚部 保科 和 夫 研究 員 に厚 くお礼 申 し上げ ます。 引 用 文 献
1) Goto, K., Iwai, M,. Takuma, Y., and Nakanishi, Y.(1992): Co-culture of in
vitro fertilized bovine embryos with different cell monolayers. J. Anim. Sci.
70:1449-1453.
2) Tanu, P and Bavister, B. D.(1991): In vitro-matured/in vitro-fertilized bovine
oocytes can develop into morulae/blastcystsin chemically defined, protein free
culture media. Biol.Reprod. 45:736-742.
3) Younis, A. I., and Brackett, B. G.(1992):Thyroid stimulating hormone enhancement
of bovine oocyte maturation in vitro. Mol .Reprod. Dev. 31:144-151.
4) Eppig. J.J., Schroeder, A. C., and O' Brien, M. J.(1992):Developmental capacity
of mouse oocytes matured in vitro: effects of gonadotrophic stimulation, follicular origin and oocytes size. J. Reprod. Fert. 95:119-127.
5) Jinno, M., SandowB. A., and Hodgen, G. C.(1989):Enhancement of the
developmental potential of mouse oocytes matured in vitro by gonadotropins and
ethelendiamenetetraacetic acid(EDTA). J. In Vitro Fertil Embryo Transfer
6:36-40.
6) Zhang, X. and Armstrong,D. T.(1989):Effects of follicle stimulating hormone
and ovarian steroids during in vitro meiotic maturation on fertilization of rat
7) Shalgi, R., Dekel.N., and Kraicer , P. F. (1979):The effect ofLH on the fertilizability
and developmental capacity of rat oocytes matured in vitro . J. Reprod . Fert .
55:429-435.
8) Vanderhyden ,B .C. and Armstrong , D. T. (1990):Effect of gonadotropins and
granulosa cell secretions on maturation and fertilization of rat oocytes in vitro . Mol. Dev. 26:337-346.
9) Kumar, J., Osborn. J. C., and Cameron , A. W. N. (1991):Luteinizing hormones
and follicle stimulating hormone induce premature condensation of chromatin in
goat(capra hircus) oocytes. Reprod. Ferti l. Dev. 3:585-591.
10) Younis, A. I. ,Zuelke, K. A., Harper, K. M ., Oliveira, M. A. L., and Brackett, B. G.(1991): In vitro fertilization of goat oocytes . Biol. Reprod. 44:1177-1182.
11) Galli, C. and Moor, R. M.(1991):Gonadotrophin requirements for the in vitro
maturation of sheep oocytes and their subsequent embryonic development .
Therio-genology 6:1083 -1093.
12) Moor, R. M. and Trounson, A. Q.(1977):Hormonal and follicular factors affecting
maturation of sheep oocytes in vitro and their subsequent developmental
capacity. J. Reprod. Fertil. 49:101-109.
13) Staigmiller, R. B. and Moor, R. M.(1984):Effect of follicle cells on the maturation and developmental competence of ovine oocytes matured outside the follicle. Gamete Res. 9:221-229.
14) Brackett, B. G., Younis, A. I., and Fayrer-Hosken, R. A.(1989):Enhanced
viability after in vitro fertilization of bovine oocytes matured in vivo with high concentration of luteinizing hormone. Fertil. Steril. 52:319-324.
15) Younis, A. I., Brackett. B. G., and Fayrer-Hosken ,R. A.(1989):Influence of
serum and hormones on maturation and fertilization of bovine oocytes in vitro.
Gamete Res. 23:189-201.
16) Zuelke ,K. A. and Brackett, B. G.(1990):Luteinizing hormone enhanced in vitro
maturation of bovine oocytes with and without protein supplementation. Biol .Rep-rod. 43:784-787.
17) Mattioli, M., Galeati, G., and Seren, E.(1988):Effect of follicle somatic cells
during pig oocytes maturation on egg penetrability and male pronucleus
formation. Gamete Res. 20:177-183.
18) Yoshida, M., Ishizaki ,Y., and Kawagishi ,H.(1990):Blastcyst formation by pig
embryos resulting from in vitro fertilization of oocytes matured in vitro. J. Reprod. Fertil. 88:1-8.
Iwasaki, S. , and Hanada,A.(1988):In-vitro fertilization of pig oocytes by frozen boar spermatozoa. J. Reprod. Fertil. 84:585-591.
20) Mattioli, M., Bacci, M. L., Galeati, G., and Seren, E.(1991):Effects of LH and
FSH on the maturation of pig oocytes in vitro. Theriogenology 36:95-105.
21) Sato, E., Matsuo, M., and Miyamoto, H. (1990):Meiotic maturation of bovine
oocytes in vitro:Improvement of meiotic competence by dibutyryl cyclic adenosine
3',5'-monophosphate. J. Anim.Sci. 68: 1182-1187.
22) Naito, K., Fukuda, Y. and Toyoda, Y.(1988):Effects of follicular fluid on male
pronucleus formation in porcine oocytes matured in vitro. Gamete Res. 21:
289-295.
23) R. H. F. Hunter and Leglise, P. C.(1992):Polyspermic fertilization following tubal surgery in pig with particular reference to the role of the isthmus. J. Reprod. Fertil. 24:233. 1971.
24) Rath, D.(1992):Experiments to improve in vitro fertilization techniques for in
vivo matured porcine oocytes. Theriogenology 37:885-896.
25) Yoshida, M.(1987):In vitro fertilization of pig oocytes matured in vitro.
Jpn. J. Vet. Sci. 49:711-718.
26) Nagai, T., Niwa, K., and Iritani, A.(1984):Effect of sperm concentration during
preincubation in a defined medium on fertilization in vitro of pig follicular
oocytes. J. Reprod. Fertil. 70:271-275.
27) Tiffin, G. J., Rieger,D., Betteridge, K. J., Yadov, B. R., and King, W. D.(1991):
Glucose and glutamine metaborism in pre-attachment cattle embryos in relation
to sex and stage of development. J. Reprod. Fert. 93:125-132.
28) Epstein, C. J., Tucker, G., Travis, B., and Gropp , A.(1977):Gene dosage for
isocitrate dehydrogenase in mouse embryos trisomic for chromosome I. Nature
267:615-616.
29) Magnuson, T.(1990):Maturations and chromosomal abnormalities:how they are
useful for studying genetic control of early mammalian development. In experimental
approaches to mammalian embryonic development. pp.437-474.Eds J. Rossant