Title 伊勢湾周辺域における淡水魚類・両生類を用いた比較系統地理学的研究( 本文(Fulltext) ) Author(s) 伊藤, 玄 Report No.(Doctoral Degree) 博士(農学) 甲第723号 Issue Date 2020-03-13 Type 博士論文 Version ETD URL http://hdl.handle.net/20.500.12099/79393 ※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。
伊勢湾周辺域における淡水魚類・両生類を用いた
比較系統地理学的研究
2019年
岐阜大学大学院連合農学研究科
生物生産科学
(岐阜大学)
伊藤 玄
伊勢湾周辺域における淡水魚類・両生類を用いた
比較系統地理学的研究
目次 第一章 緒言...............................4 第二章 伊勢湾周辺域におけるホトケドジョウの遺伝的集団構造 緒言...................................7 材料及び方法...............................8 結果..................................14 考察..................................19 第三章 伊勢湾周辺域におけるトウカイコガタスジシマドジョウの遺伝的集団構造 緒言..................................23 材料及び方法..............................23 結果..................................27 考察..................................32 第四章 伊勢湾周辺域におけるアカハライモリの遺伝的集団構造 緒言..................................36 材料及び方法..............................37 結果..................................45 考察..................................50 第五章 総合考察1 伊勢湾周辺域における遺伝的多様性の創出機構.....53 第六章 総合考察2 伊勢湾周辺域におけるホトケドジョウ・トウカイコガタスジシ マドジョウ・アカハライモリの保全単位................. 57
謝辞..................................62
第一章 緒言
比較系統地理学(comparative phylogeography)とは,分布域を共有し,類似した生 態もしくは生活史をもつ複数種間の遺伝的集団構造を比較する研究分野であり,地 域の生物相の形成プロセスの理解に有用である(Avise, 2000;Lomolino et al., 2017).淡水魚類や両生類などの水生生物は移動性が低いことから,これらの生物 の遺伝的集団構造は古環境や地史に影響を受けることが一般的である(Avice, 2000,2004).水生生物を対象とした比較系統地理学的研究は世界中で行われてお り,例えば北アメリカのフロリダ半島周辺やカリフォルニア州(Swift et al., 1985; Soltis et al., 2006;Schierenbeck, 2014),中央アメリカ(Bermingham and Martin, 1998;Hulsey et al., 2004),ヨーロッパ南部(Hewitt, 2000;Costedoat and Gilles, 2009),オーストラリア東部(Page and Hughes, 2014),台湾(Watanabe et al., 2007;Chiang et al., 2017)などの様々な地域において,生物の分布に影響を及ぼした 主要な地理的障壁や,気候変動による隔離と分散の歴史が明らかにされている.し かし,現在までに対象とされてきた地域は比較的広い範囲の地域であり,狭い範囲 の地域を対象とした研究は少ない.狭い地域では明瞭な遺伝的集団構造が相対的に 得られにくく,地域の生物相の形成プロセスを明らかにすることが困難であるため と考えられる. 狭い地域における集団構造の詳細な研究は,集団および地域生物相の進化の歴史 を理解するのに役立つと考えられる(Watanabe and Chiba, 2001; Watanabe and Mori, 2008; Koizumi et al., 2012).また,このような研究は,集団間の遺伝子流動と地理的 隔離の実際の状態を反映できるため,広範囲の生物地理学的研究よりも進化学的プ ロセスと種分化の研究により大きなインパクトを与えることができる(Schilthuizen et al., 2006; Kuchta et al., 2009; Chiba and Cowie, 2016).狭い地域での系統地理学的研 究では,地理的に容易に隔離される移動性の低い動物が対象として適していると考 えられ,実際にいくつかの地域では明瞭な遺伝的集団構造が得られている(例:ザ
リガニ,Koizumi et al., 2012;サンショウウオ,Kuchta et al., 2009;Sugawara et al., 2016;陸貝,Shimizu and Ueshima, 2000;Watanabe and Chiba, 2001).加えてこれら の研究は,詳細な保全単位[たとえば,Moritz(1994)で定義されている進化的重要 単位(ESU)など]の確立などに役立てることが可能であり,保全生態学にとって 特に重要である(Avise, 2000). 日本列島の本州中部に位置する愛知県・岐阜県・三重県にまたがる伊勢・三河湾 周辺の地域は,淡水魚などの水生生物において固有性の高い生物相が発達してお り,伊勢湾周辺域と呼ばれている(渡辺ほか,2006).伊勢湾周辺域では,更新世 に様々な山地の隆起や断層の活動が生じており(吉田・尾崎,2000;町田ほか, 2006),これらの地殻変動が淡水魚類や両生類の分布に大きな影響を与えたと考え られる.例えば鈴鹿山脈の隆起は,淡水魚ではカワバタモロコHemigrammocypris
rasborella,オイカワ Opsariichthys platypus,カワヒガイ Sarcocheilichthys variegatus variegatus,カマツカ Pseudogobio esocinus,およびナガレカマツカ P. agathonectris な
どの多くの淡水魚で,両生類ではヤマトサンショウウオHynobius vandenburghi にお いて伊勢湾周辺域と近畿地方の系統群に遺伝的に分断されたことがそれぞれの種内 のミトコンドリアDNA の系統地理によって示唆されている(Tominaga et al., 2009, 2016;Komiya et al., 2014;Watanabe et al., 2014;岐阜県立岐阜高等学校自然科学部生 物班,2014;藤谷ほか,2016;Kitanishi et al., 2016;Matsui et al., 2019).また,静 岡県西部と愛知県東部の間にまたがる弓張山地の隆起は,カワバタモロコを伊勢湾 周辺域と,浜名湖から藤枝市以西の地域(西静岡地域)の系統群に分断したと考え られている(Watanabe and Mori, 2008).これらの結果,伊勢湾周辺域は狭い地域で あるにも関わらず,他の地域とは遺伝的に異なる固有の集団の多い地域となったと 考えられる.
一方,最終氷期最盛期には海面が現在よりも100 m 以上低下していたため,水深 の浅い伊勢湾は陸化し,伊勢湾周辺域の水系は単一の古水系を形成したと考えられ ている(森山,2004).それによって,淡水魚についてはウシモツゴ Pseudorasbora
pugnax,ネコギギ Tachysurus ichikawai,カワバタモロコ,およびカワヒガイなど
が,両生類についてはヤマトサンショウウオが,この古水系を介して遺伝的に交流 したことが示唆されている(Watanabe and Nishida, 2003;Watanabe and Mori, 2008; 岐阜県立岐阜高等学校自然科学部生物班,2014;鈴木ほか,2016).ただしその 後,第四紀の間氷河期の海水面上昇時に水系が分割形成されたため(太田, 2009),氷期−間氷期サイクルが卓越していた時期では,分布の拡大と隔離が頻繁に 発生した可能性がある.これらの伊勢湾周辺域における歴史的な地形的変化は,こ の地域の動植物の個体群の細分化を引き起こした可能性がある.しかし,多くの淡 水魚における系統地理学的研究からは,過去の遺伝的集団構造は明確に示されてい ない(Kitagawa et al., 2001; Watanabe and Nishida, 2003; 堀川・向井,2007; Tominaga et al., 2009, 2016; Kakioka et al., 2013; Nakagawa et al., 2016; Kitanishi et al., 2016; 鈴木ほ か,2016).一方,水田周辺の池や小川湿地に生息するカワバタモロコとウシモツ ゴは,伊勢湾周辺域でわずかな遺伝的分化を示している(Watanabe and Mori, 2008).したがって,地史的イベントが地域の淡水魚に影響を与えた場合,移動性 の高い淡水魚よりは明瞭な遺伝的集団構造を保持している可能性がある. これらのことから本研究では,伊勢湾周辺域のような狭い地域においても移動性 の低い水生生物であれば,山地の隆起などの地史的影響による遺伝的集団構造を保 持している可能性を検証するために,ホトケドジョウ,アカハライモリおよびトウ カイコガタスジシマドジョウを対象に伊勢湾周辺域における遺伝的集団構造を推定 し比較した.総合考察では,上記3 種の系統地理パターンを比較し,生物地理学的 な考察を行うとともに,伊勢湾周辺域におけるどのような地史的イベントが遺伝的 集団構造の創出に重要な役割を担ってきたのかを考察した.また,これらの種は環 境省やさまざまな地域版のレッドリストに絶滅危惧もしくは準絶滅危惧としてリス トアップされている希少生物であるため,各種から得られた遺伝的集団構造を元に した保全単位についても検討した.
第二章 伊勢湾周辺域におけるホトケドジョウの遺伝的集団構造 緒 言 ホトケドジョウLefua echigonia(コイ目フクドジョウ科)は,日本列島に固有の 純淡水魚である(Kottelat, 2012;中島・内山,2017).ホトケドジョウの生息地は, 基本的に谷戸の湧水河川に限定されているため,大河川を通じた移動分散が少ない と考えられ,移動性は低いと考えられる(柿野ほか,2006a,2006b;Mitsuo et al. 2013).このようなホトケドジョウの生態的特徴は,伊勢湾周辺域における地史と 集団分化のプロセスの関連性を明らかにするために適していると考えられる.ホト ケドジョウの分布域を網羅した系統地理学的研究では,mtDNA に基づいて東北,北 陸,山形,北関東,南関東,東海,近畿の7 つの主要な系統に細分されることが示 された.(Saka et al., 2003;Aiki et al., 2009;Miyazaki et al., 2011).7 つの主要な系 統のうち,東海系統には少なくとも遺伝的に2 つの地域系統が含まれている
(Miyazaki et al., 2017).しかし,Miyazaki et al.(2017)の主なサンプリング地点 は,静岡県西部から伊勢湾周辺域の東端であった.したがって,伊勢湾周辺域にお ける東海系統の細分化の主な要因は明らかにされていない.その要因を明らかにす るためには,伊勢湾周辺域における分布範囲の全域からホトケドジョウを収集する 必要がある. そこで本研究では,系統地理を調査するために,mtDNA の cyt b 領域を使用し て,伊勢湾周辺域におけるホトケドジョウの遺伝的集団構造を調べた.比較のため に,伊勢湾周辺域と西静岡地域の分布範囲全体から標本をサンプリングした.調査 結果に基づいて,地理的イベントの形成年代と遺伝的系統間の分岐時間との関係に ついて議論した.
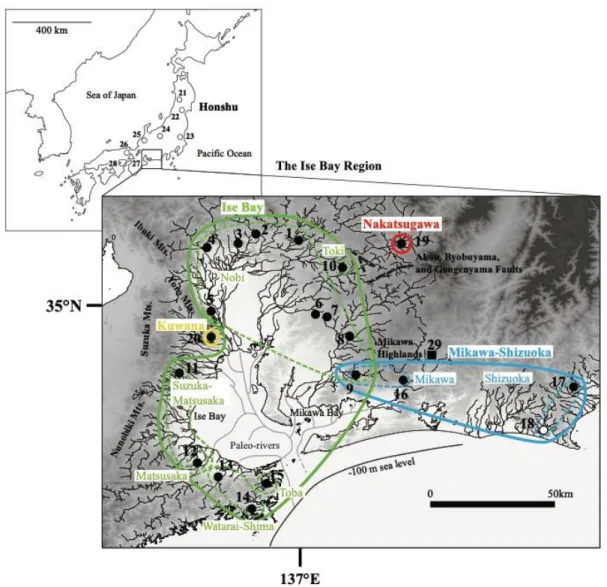
材料と方法 2014 年から 2015 年にかけて,伊勢湾周辺域と西静岡地域の 19 地点から合計 145 個体のホトケドジョウを採集した(Fig. Ⅱ-1; Table Ⅱ-1).各個体はタモ網を用いて 採集した.採集した個体から尾鰭の一部を切除し,99 %エタノールで保管し,– 20 °C のフリーザ内で保管した.すべての個体は,採集した地点に放流した.加え て,トウカイナガレホトケドジョウを愛知県新城市(Loc. 29)から採集し,同様の 方法で処理した.
DNA 抽出は,Quick Gene DNA tissue kit S(Kurabo 社)を用いて行った.mtDNA のcyt b 領域を対象とした PCR 増幅と塩基配列の決定には,L14736-Glu(5′-AAC CAC CGT TGT TAT TCA ACT A-3′)(Mabuchi et al., 2006)と H15913-Thr-AH (5′-CCG ATC TTC GGA TTA CAA GAC CG-3′)(向井ほか,2011)を,mtDNA の調節 領域(CR)の増幅塩基配列の決定には,L15998-Pro(5’-TAC CCC AAA CTC CCA AAG CTA-3’)(Alvarado Bremer and Zhang 1998)と H690(5’-GTT GAG GCT TGC ATG TAT A-3’)(Kumazawa et al. 1996)のプライマー対を用いた.CR の増幅とシ ークエンスは,cyt b 領域の異なるハプロタイプが得られた個体からのみ行った. PCR には,Ex Taq バッファーと Ex Taq DNA ポリメラーゼ(タカラバイオ社)を使 用し,常法に従ってサーマルサイクラ―で 95 °C 1 分,55 °C 1 分,72 °C 2 分の温度 サイクルを30 回繰り返して増幅を行った.PCR 産物については ExoSAP-IT キット (GE ヘルスケアジャパン社)を用いて処理した後,増幅した時と同じプライマーを それぞれ用いて,BigDye Terminator Cycle Sequencing Kit ver. 3.1(アプライドバイオ システムズ社)によるシークエンス反応をL 鎖と H 鎖について行い,ABI 3130 Genetic Analyzer で塩基配列を決定した.得られた両鎖の塩基配列については個体ご とに結合させて,両鎖の不一致が無いことを確認した上でDDBJ,EMBL,GenBank のデータベースに登録した(cyt b:LC079000–LC079010,LC079012–LC079031, CR:LC404130–LC404154).また,集団遺伝データは遺伝的多様性データベース GEDIMAP(Watanabe et al., 2010a)に登録した(P1980–1999).
Fig. II-1 Sampling localities of Lefua echigonia (circles), L. torrentis (triangles), and L. sp.
2. (square). Localities with closed symbols indicate collection points in the present study, and localities with open symbols indicate collection sites used by Saka et al. (2003). Distribution patterns for each sub- and local lineage of L. echigonia in the Ise Bay and West-Shizuoka regions are shown. Solid lines indicate four sub-lineages, and thin broken lines indicate eight local lineages. The paleo-river system follows Moriyama (2004).
Table II-1 List of sampling localities, number of specimens (n), and haplotypes in Lefua
species group
得られた塩基配列は,CLUSTAL X, Version 1.83(Thompson et al., 1997)を用いて 多重整列を行った.なお,CR の多重配列では,挿入と欠損の箇所は除いた.Cyt b のデータセットを用いた系統樹の推定には,西静岡地域の塩基配列(Loc. 18)は, DDBJ から入手した(Table Ⅱ-1).加えて,Saka et al.(2003)で調べられた他のホ トケドジョウの地域系統[北陸:(AB080141,Loc. 21),東北(AB080155,Loc. 22),北関東(AB080151,Loc. 23),南関東(AB080163,Loc. 24),近畿 (AB080176,Loc. 25 および AB080181,Loc. 26]や,ナガレホトケドジョウの地域 系統(AB080182,山陽,Loc. 27 および AB080183,紀伊・四国,Loc. 28)の塩基配 列,本研究で収集したトウカイナガレホトケドジョウ(LC079031,Loc. 29)を外群 として使用した(Fig. Ⅱ-1;Table Ⅱ-1). 系統解析は,最尤法とベイズ法を用いた.最尤法には,raxmlGUI, Version 1.3 (Silvestro and Michalak, 2012)を用いた.Cyt b,CR の各データセット共に,塩基配 列におけるコドンの1 番目,2 番目,3 番目の進化速度の違いを考慮し,それぞれに パーティションを設定し,GTR+G モデルを適用した.各枝の信頼性については 1000 回のブートストラップ確率を求めた.ベイズ法には,MrBayes, Version 3.2.6 (Ronquist et al., 2012)を用いた.Cyt b 領域のデータセットでは,Kakusan4 (Tanabe, 2011)を用いて BIC 基準に基づいたモデルテストを行い,1 番目,2 番 目,3 番目のコドンに,GTR+G,K80+G,HKY+G の各モデルを適用した.CR のデ ータセットでは,パーティションの設定はせず,HKY+G モデルを適用した.解析 は,マルコフ連鎖モンテカルロ(MCMC)法によって 10000000 世代繰り返し,100 世代ごとにサンプリングを行い,burnin を 25000 世代に設定した.その際に, Average Standard Deviation of Split Frequencies(ASDSF)が 0.01 未満になったことで MCMC が定常状態に達したと判断した.
Cyt b データセットにおいて,MEGA6(Tamura et al., 2013)を用いて,p-distance による各亜系統と地域系統間の遺伝距離を求め,標準誤差を1000 回のブートストラ
ップ確率によって評価した(Table Ⅱ-2).加えて,ハプロタイプネットワークを TCS 1.2.1(Clement et al., 2000)を用いて作製した.
上記の系統推定によって得られた系統間の分岐年代を,BEAST ver. 1.8.2
(Drummond et al., 2012)を用いて推定した.Cyt b データセットを使用し,コドンそ れぞれにパーティションを設定し,BIC によって選択された TN93+I モデルを適用 し,Yule (speciation) tree prior を選択した.本プログラムでは,uncorrelated lognormal relaxed clock モデルを仮定し,共通祖先年代(tMRCA)を推定するために,2 点のノ ードに較正点を設定した.1 つ目は,ホトケドジョウ東海系統および近畿系統が,約 100−150 万年前の伊吹・鈴鹿・布引山脈の隆起(川辺,1994)によって分断された と仮定し,較正点1(CA1)として設定した.この較正は,事前分布を対数正規分 布(lognormal prior distribution)と仮定し,95%信頼区間を約 1.1−1.5 Mya,平均 1.3 Mya,標準偏差 0.1,offset を 0 として設定した.2 つ目に,中津川系統が,更新世中 期の恵那・中津川盆地を形成した赤河・屏風山・権現山断層の活動によって分断さ れたと仮定し(約20−80 万年前:森山 1985),較正点 2(CA2)として設定した. この較正は,事前分布を対数正規分布(lognormal prior distribution)と仮定し,95% 信頼区間を約0.35−0.68 Mya,平均 0.5 Mya,標準偏差 0.2,offset を 0 として設定し た.他のすべてのモデルパラメータは,初期値で設定した.MCMC 分析は,分析ご とに5000 万世代の 2 つの独立したランを実行した.1000 世代ごとにサンプリングを 行い,burnin として初期サンプルの 10%を削除した.TRACER v1.6(Rambaut et al., 2014)を使用して,チェーンの定常分布と有効サンプルサイズ(ES > 200)への収束 を確認した.コンセンサスツリーは,BEAST package の TreeAnnotator v1.8.2 を使用 して作成し,FigTree v1.4.2(Rambaut, 2014)を用いて系統樹を視覚化した.ベイズ 系統樹の各枝の評価は事後確率(PP)によって行った.
Table II-2 Genetic distances among sub-lineages and local lineages in Lefua echigonia
Sub-lineages Ise Bay Mikawa-Shizuoka Nakatsugawa Kuwana Ise Bay
Mikawa-Shizuoka 0.026 ± 0.004
Nakatsugawa 0.034 ± 0.005 0.038 ± 0.005
Kuwana 0.049 ± 0.006 0.050 ± 0.006 0.046 ± 0.006
Other Kinki 0.055 ± 0.006 0.056 ± 0.006 0.052 ± 0.006 0.044 ± 0.006
Local lineages Nobi Toki Matsusaka Suzuka-Matsusaka Ise-Shima Mikawa Nobi Toki 0.015 ± 0.004 Matsusaka 0.011 ± 0.003 0.012 ± 0.003 Suzuka-Matsusaka 0.015 ± 0.003 0.014 ± 0.004 0.009 ± 0.003 Ise-Shima 0.016 ± 0.003 0.016 ± 0.003 0.011 ± 0.002 0.013 ± 0.003 Mikawa 0.027 ± 0.004 0.031 ± 0.005 0.026 ± 0.005 0.030 ± 0.005 0.031 ± 0.005 Shizuoka 0.024 ± 0.004 0.026 ± 0.005 0.022 ± 0.004 0.025 ± 0.005 0.026 ± 0.004 0.023 ± 0.004 Genetic distances were calculated based on uncorrected p-distance. Data represent the mean ± standard error.
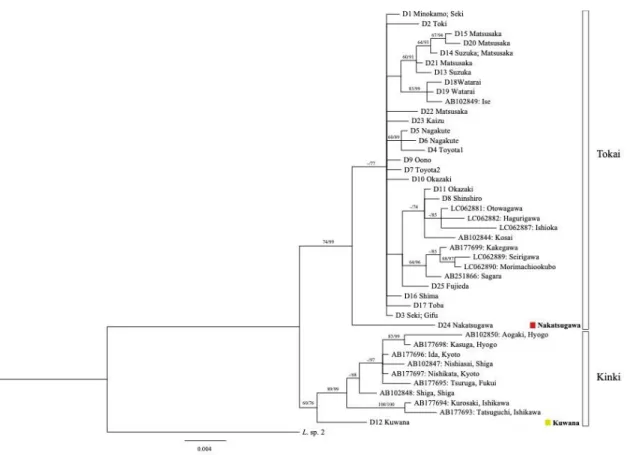
結 果 各系統の地理的分布 伊勢湾周辺域と西静岡地域から採集した合計145 個体から mtDNA の cyt b 領域 1023 bp を決定したところ,合計 30 ハプロタイプ(L1-30)が確 認された(Table Ⅱ-1).Cyt b データセットでは,最尤法とベイズ法で推定された樹 形はほとんど同じトポロジーを示し(Fig. II-2),伊勢湾周辺域に東海系統および近 畿系統が分布することを確認した.この系統樹の東海および近畿系統は,遺伝距離 (uncorrected p-distance)では 0.053 ± 0.005 SE(標準誤差)と,明確な単系統群を 形成した(近畿系統,BP = 78%,PP = 95%,東海系統,BP = 85%,PP = 100%) (Fig. II-2;Table II-2).伊勢湾周辺域から得られた近畿系統は,近畿地方の姉妹群 からの初期の分岐を示し,遺伝距離(uncorrected p-distance)では 0.044 ± 0.006 SE と,明確な分岐が見られたため,桑名系統と名付けた.桑名系統は,員弁川水系で のみ得られた(Loc.20;Table II-1).ただし,最尤法では,桑名系統の位置のみ異 なっていた.最尤法では桑名系統は,東海・近畿系統と姉妹群を形成したが,ブ−ト ストラップ値(BP)は 60%と低かった(Fig. Ⅱ−3).一方で,ベイズ法では桑名系 統は近系統に内包されることが89%と高い事後確率(PP)で示された.CR データ セットでは,最尤法,ベイズ法共に,cyt b データセットにおけるベイズ法と同様の 結果が示された(Fig. Ⅱ−4).すなわち,桑名系統は近畿系統に内包される系統であ ると判断した(BP = 60%,PP = 76%). 先行研究で明らかにされていた東海系統は,本研究で3 つの系統(伊勢湾,三川 静岡,中津川)に分けられ,これらの系統の独立性は強く支持された(BP ≥ 85%, PP ≥ 96%).中津川系統は,恵那・中津川流域の木曽中流域で発見され(Loc. 19),伊勢湾および三河・静岡系統を含む姉妹クレードから最初に分岐し,遺伝距 離(uncorrected p-distance)では 0.035 ± 0.005 SE と,明瞭な分岐が見られた(Fig. II-2).
Fig. II-2 Time-calibrated Bayesian inference tree of the 1023-bp cytochrome b gene
sequences of Lefua echigonia individuals from populations from the Ise Bay and West-Shizuoka regions and several major lineages. The tree is dated using the uncorrelated
lognormal relaxed clock model with node-age constants (calibration point (CA) 1 and CA2). Numbers at nodes indicate maximum likelihood (ML) bootstrap probabilities (left) and Bayesian posterior probabilities (right) >70% (both parameters). Bars show the credibility intervals as 95% highest probability density. Numbers in parentheses indicate the number of specimens. The asterisk indicates another position pattern of the Kuwana sub-lineage in the case of the ML tree (see the text). The statistical parsimony network of six local lineages within the Ise Bay sub-lineage derived from the cyt b sequences is shown.
Fig. II-3 Maximum likelihood (ML) tree of the 1023-bp cytochrome b gene sequences of
Lefua echigonia individuals from populations from the Ise Bay and West-Shizuoka regions
and several major lineages. Numbers at nodes indicate ML bootstrap probabilities (left) and Bayesian posterior probabilities (right) > 70% (both parameters). The asterisk indicates the node of the Kuwana sub-lineage obtained by ML method which was different from that obtained by Bayesian inference method (see the text).
Fig II-4 Bayesian phylogenetic tree of the 872-bp control region sequences of Lefua
echigonia individuals from populations of Ise Bay, West-Shizuoka regions, and Lefua sp. 2. Numbers at nodes indicate maximum likelihood bootstrap probabilities (left) and Bayesian
伊勢湾系統は,三河・静岡系統から分岐しており,遺伝距離(uncorrected p-distance)は 0.026 ± 0.004 SE だった.本系統は,伊勢湾周辺域の東側と西側に広く 分布しており,6 つの地域系統(濃尾,土岐,鈴鹿・松阪,松阪,志摩・度会,鳥 羽)に分けられた.これらの地域系統については,遺伝距離(uncorrected p-distance)は 0.011 ± 0.002 SE であり,各地域系統の単系統性は高い値で支持された (BP ≥ 77%,PP = 100%).これらの中で,濃尾地域系統は,木曽川,庄内川,矢 作川水系から得られ(Loc.1−9;Table II-1),土岐地域系統は庄内川中流域で発見 された(Loc.10).これら 2 つの地域系統の分布は,伊勢湾の東側に限定されてい た.鈴鹿・松阪地域系統は,鈴鹿川水系と金剛川水系(Loc.11 および 12)からの み,松阪地域系統は金剛川水系(Loc.12)からのみ,志摩・度会地域系統は宮川, 西川水系(Loc. 13,14),鳥羽系統は鴨川水系(Loc. 15)からのみ得られ,これら 4 つの地域系統の分布は,伊勢湾の西側に限定されていた.ほぼすべての地域から は,1 つの地域系統しか得られなかったが,金剛川(Loc.12)からは,2 つの地域 系統(鈴鹿松阪と松阪)が得られた. 最後に,三河・静岡系統は,伊勢湾地域の東端(三河高原周辺)と西静岡地域に 分布していた.三河・静岡系統は,2 つの地域系統(三河,静岡)に分けられ,遺伝 距離(uncorrected p-distance)は 0.017 ± 0.003 SE であり,これらの地域系統の単系 統性は強く支持された(BP = 98%,PP = 100%).これらの地域系統のうち,三河 地域系統は,矢作川水系と豊川水系で発見され(Loc.9 と 16),静岡系統は瀬戸川 水系と菊川水系で発見された(Loc.17 と 18).矢作川水系の東支流である Loc. 9 において,三河・静岡系統の三河地域系統と伊勢湾系統の濃尾地域系統の分布が重 なっていた(Table II-1,Fig. II-1).
分岐年代推定 分岐年代推定の結果,東海と近畿系統の分岐年代は約1.17 Mya [95%信頼区間(HPD),0.96–1.39 Mya],中津川系統と姉妹クレードの分岐年代 は約0.64 Mya(95%HPD,0.45–0.84 Mya),伊勢湾と三河・静岡系統の分岐年代は 0.51 Mya(95%HPD,0.34−0.69 Mya)と推定された.伊勢湾系統内の地域系統は,6
つの地域系統に分けられた.これらのローカル系統の分岐は,約0.34 Mya(95% HPD,0.21–0.48 Mya)で発生すると推定された.三河・静岡系統の地域系統の分岐 年代は,約0.38 Mya(95%HPD,0.21–0.57 Mya)と推定された.近畿系統内では, 桑名系統は伊勢湾周辺域側に,他の近畿系統が伊吹山,鈴鹿山,布引山の反対側に 分布していた.近畿系統におけるこれらの系統の分岐年代は,約0.91 Mya(95% HPD,0.60–1.25 Mya)と推定された.平均進化速度(ucld. mean)は,cyt b 領域で 0.032 と推定された. 考 察 本研究では,伊勢湾周辺域のホトケドジョウにおいて,4 つの mtDNA 系統の存在 が明らかになった.そのうちの3系統(伊勢湾,三河・静岡,中津川系統)は東海 系統に含まれ,残りの1系統(桑名系統)は近畿系統に含まれた.Cyt b および CR で推定した系統解析の結果から,桑名系統はおそらく近畿系統に内包され,他の近 畿系統のハプロタイプと深く分岐した系統であると考えられる.ホトケドジョウの 東系統と近畿系統の分岐は,他のいくつかの淡水魚で見られるように,伊吹山,鈴 鹿山,布引山の形成によって生じたと考えられる(例えば,イチモンジタナゴ
Acheilognathus cyanostigma,Kitazima et al., 2015;シロヒレタビラ Acheilognathus tabira tabira,梅村ほか,2012;ゼゼラ Biwia zezera,Watanabe et al., 2010b;オイカワ Opsariichthys platypus,Kitanishi et al., 2016;カマツカ Pseudogobio esocinus,
Tominaga et al., 2009,2016;カワヒガイ Sarcocheilichthys variegatus variegatus, Komiya et al., 2014;鈴木ほか,2016).これらの他の淡水魚では,伊勢湾地域の在 来系統ハプロタイプに加えて,近畿地方の系統(近畿系統)に属するハプロタイプ も検出されている.これらの淡水魚場合,伊勢湾地域で検出された近畿系統のハプ ロタイプは近畿地方の在来系統のハプロタイプと同一または類似していたため,近 畿地方からの人為的な移入の結果である可能性が示された(堀川・向井,2007; Tominaga et al.2009;梅村ほか,2012;Kitazima et al., 2015;Kitanishi et al., 2016;鈴
木ほか,2016).一方,ホトケドジョウの場合,桑名亜系と近畿系統の他の系統と の間の遺伝的距離は比較的大きく,長期的な隔離を示唆している.しかし,先行研 究では,mtDNA の cyt b 領域で調べられた近畿は,2 地点のみだった(Saka et al., 2003).CR を用いた Miyazaki et al.(2011)で,多くの CR を用いた解析を行ったこ とから,本研究でもCR を用いた桑名系統の系統学的位置を再分析した.その結 果,CR でも,桑名亜系統のハプロタイプは近畿系統の他のハプロタイプに対応して いなかった(Fig. II-4).したがって,本研究の結果に基づいて,桑名系統は伊勢湾 周辺域に在来の系統であると考えられた.近畿系統に属する桑名亜系は,伊勢湾地 域の西端の鈴鹿・養老山地間にある1 地点のみに局在していた(Fig. 1,Table II-1).この地理的分布から,桑名系統の存在は,東海と近畿系統が鈴鹿山脈の隆起の みで分断されたわけではなく,養老山地の存在が大きいことを示唆する.多くの日 本産淡水魚類の系統地理学的研究では,系統間の分岐年代の較正点として鈴鹿山脈 の隆起年代を用いている(例えば,Komiya et al. 2014;Watanabe et al. 2014;
Kitanishi et al. 2016;Tominaga et al. 2016 など).桑名系統の存在は,鈴鹿山脈周辺 において異なる年代で分断が生じた可能性を示唆するものであり,鈴鹿・養老山地 間(員弁・牧田川水系)に分布する個体群のさらなる調査が必要である.
中津川系統は,伊勢湾周辺域北部の東濃地方の恵那・中津川地域に局在していた (Fig. II-1,3,Table II-1).東濃地方は,第四紀における断層運動に起因する丘陵 地で構成されている.恵那・中津川地域は,更新世中期に隆起した赤河,屏風山, 権現山断層によって伊勢湾地域の他の地域から隔離された(森山,1985;町田ほ か,2006).東海丘陵地要素とよばれる湿地性植物に関する先行研究(例えばハナ ノキAcer pycnanthum;シデコブシ Magnolia stellate)は,これらの植物の分布が東濃 の断層運動によって制限されていることを示唆している(糸魚川,2011,2015). したがって,中津川系統はおそらく,この地域の断層運動によって隔離されたと考 えられ,伊勢湾周辺域の他の系統から中期更新世の約0.46−0.86 Mya に分岐したと推 定される東濃地方固有の系統であると考えられる.しかし,同様の系統地理学的パ
ターンは本地域における他の淡水魚の先行研究からは報告されていない(例えば, Kitagawa et al., 2001;Watanabe and Mori, 2008;Kitanishi et al., 2016 など).おそら く,ほかの淡水魚類は,木曽川本流を介して分散することができる遊泳力を持ち合 わせているためと考えられる. Miyazaki et al.(2017)は,mtDNA の CR 領域の配列に基づいて東海系統の種内変 異を調査し,伊勢湾周辺域とその周辺に2 つの遺伝的グループが存在することを報 告した(西静岡地域に分布する「グループA」および伊勢湾地域の東端に分布する 「グループB」).分布パターンに基づき,本研究の三河・静岡系統は「グループ A」,伊勢湾系統は「グループ B」にそれぞれ対応すると判断した.三河・静岡系統 と伊勢湾系統の同所的な分布は,ホトケドジョウのCR の塩基配列を解析した Miyazaki et al.(2017)でも報告されている.これらの結果は,河川争奪か古水系の 接続によって分散したものと考えられる. 伊勢湾系統(Miyazaki et al., 2017 における「グループ B」に対応)は,桑名系統, 中津川系統が見つかった地域を除き,伊勢湾の西側(三重地域)と東側(愛知岐阜 地域)に広く分布していた.伊勢湾と三河湾のすべての河川系は氷河期に接続し, 単一の大きな古河川系を形成したと考えられている(Fig. II-3,森山,2004).この 古水系は,伊勢湾の西側と東側の間の淡水魚や湿地性植物の遺伝子流動を促進した ことが示唆されている(Watanabe and Nishida, 2003;Watanabe and Mori, 2008;鈴木 ほか,2016;Saeki et al., 2015).ホトケドジョウでも,同様に古水系によって交流 した可能性が高い. 本研究では,伊勢湾内に6 つ,三河・静岡系統内に 2 つの地域系統が分けられ た.各地域系統は,分布が局所的であるものが多いが,分布の境に明瞭な地理的障 壁が見られない系統もある.松阪地域系統と鈴鹿・松阪系統は1 地点から同所的に 確認されていることから,明瞭な地理的障壁によって隔離され,成立したものとは 考えにくい.ホトケドジョウの移動性は,非常に低いと考えられており(Mitsuo et al., 2013),主に谷戸の細流に生息している(柿野,2016a,2016b).そして,河川
本流にはほとんど出現しないと言われている(細江・古屋,2008).この生態的特 徴は,おそらくこれらの地域系統の分断を容易にしたものと考えられる.
第三章 伊勢湾周辺域におけるトウカイコガタスジシマドジョウの遺伝的集団構造
緒 言
トウカイコガタスジシマドジョウCobitis minamorii tokaiensis は,西日本に広く分 布するコガタスジシマドジョウの1 亜種であり,東海地方の伊勢・三河湾周辺と遠 州灘西部の流入河川(愛知県・岐阜県・三重県・静岡県)にのみ分布する(中島・ 内山,2017).本亜種は,河川下流域や,平野部の農業用水路・水田・河川敷湿地 帯などの氾濫原に主に生息し,産卵の際に浅い水路や水田などに移動すると考えら れている(中島・内山,2017).これまで東海地方における詳細な遺伝的集団構造 が調べられているドジョウ類として,アジメドジョウNiwaella delicata(Kitagawa et al., 2001)や,トウカイナガレホトケドジョウ Lefua tokaiensis(Miyazaki et al., 2017) があるが,本亜種の生息環境や生活史は,これらのドジョウ類とは大きく異なって おり,生態の違いと遺伝的集団構造の関係を明らかにするには好材料であると考え られる.
材料と方法
2014 年から 2019 年にかけて,合計 16 地点 165 個体[瑞穂(Mizuho)6 個体;関 (Seki)16 個体;山県(Yamagata)24 個体;垂井(Tarui)19 個体;一宮
(Ichinomiya)1 個体;大口(Oguchi)12 個体;名古屋(Nagoya)12 個体;西尾 1 (Nishio1)2 個体;西尾 2(Nishio2)4 個体;松阪 1(Matsusaka1)7 個体;松阪 2 (Matsusaka2)13 個体;伊勢(Ise)16 個体;湖西(Kosai)4 個体;浜松 1
(Hamamatsu1)10 個体;浜松 2(Hamamatsu2)9 個体;森(Mori)10 個体]のトウ カイコガタスジシマドジョウを,タモ網を用いて採集した(Fig. 1;Table III-1).加えて,比較用に岐阜県から 1 個体,滋賀県から 1 個体のニシシマドジョウ
Fig. III-1 Sampling localities of Cobitis minamorii tokaiensis (closed circles). The paleo-river
Table III-1 List of sampling localities for Cobitis minamorii tokaiensis
*number of haplotypes; **gene diversity; ***nucleotide diversity, **** Grounds of regional division described in Results
しくは腹鰭の一部を99.5%エタノール中に浸漬し,DNA の抽出を行うまで–80°C の 超低温フリーザ内で保管した.
DNA 抽出には,DNeasy Bood & Tissue Kit(キアゲン社)を用いた.mtDNA の cyt
b 領域を対象とした PCR 増幅と塩基配列の決定には,Glu-L(5’-GAA GAA CCA
CCG TTG TTA TTC AA-3’)と Thr-H(5’-ACC TCC RAT CTY CGG ATT ACA-3’)の プライマー対(Šlechtová et al., 2006)を用いた.PCR には,Ex Taq バッファーと Ex Taq DNA ポリメラーゼ(タカラバイオ社)を使用し,常法に従ってサーマルサイク ラ―で95°C・1 分,55°C・1 分,72°C・2 分の温度サイクルを 30 回繰り返して増幅 を行った.PCR 産物については ExoSAP-IT キット(GE ヘルスケアジャパン社)を 用いて処理した後,増幅した時と同じプライマーをそれぞれ用いて,BigDye Terminator Cycle Sequencing Kit ver. 3.1(アプライドバイオシステムズ社)によるシ ークエンス反応をL 鎖と H 鎖について行い,ABI 3130 Genetic Analyzer で塩基配列 を決定した.得られた両鎖の塩基配列については個体ごとに結合させて,両鎖の不 一致が無いことを確認した上でDDBJ,EMBL,GenBank のデータベースに登録した (登録番号 LC464984–LC465017,LC487327).また,集団遺伝データについては 遺伝的多様性データベースGEDIMAP(Watanabe et al., 2010)に登録した(登録番号 P2307–2320,2352,2353).
比較のための多重整列には,CLUSTAL X ver. 1.83(Thompson et al., 1997)を用 い,系統樹の推定には,最尤法とベイズ法を用いた.最尤法には,raxmlGUI ver. 1.3 (Silvestro and Michalak, 2012)を用いた.塩基配列におけるコドンの 1 番目,2 番 目,3 番目の進化速度の違いを考慮し,それぞれにパーティションを設定し, GTR+G モデルを適用した.各枝の信頼性については 1000 回のブートストラップ確 率を求めた.ベイズ法には,MrBayes ver. 3.2.6(Ronquist et al., 2012)を用いた. Kakusan4(Tanabe, 2011)を用いてベイズ情報量規準(BIC)に基づいたモデルテス トを行い,1 番目,2 番目,3 番目のコドンに,K80,F81,GTR の各モデルを適用 した.解析にあたっては,マルコフ連鎖モンテカルロ(MCMC)法によってマルコ
フ連鎖を10000000 世代繰り返し,100 世代ごとにサンプリングを行い,burnin を 25000 世代に設定した.その際に,Average Standard Deviation of Split Frequencies (ASDSF)が 0.01 未満になったことで MCMC が定常状態に達したと判断した.系 統樹の外群には,ビワコガタスジシマドジョウC. m. oumiensis(AP010782, Saitoh et al., 2010),岐阜県産と滋賀県産のニシシマドジョウ(LC465018–LC465019, 本研 究),ドジョウMisgurnus anguillicaudatus(AP011291, Miya et al., 2015)の相同配列 を用いた.加えて,ハプロタイプネットワークをTCS 1.2.1(Clement et al., 2000)を 用いて作成した.MEGA6(Tamura et al., 2013)を用いて,p-distance による遺伝距離 を求め,1000 回のブートストラップ検定により標準誤差を求めた.採集地点間の遺 伝的分化について調べるため,Arlequin ver. 3.5(Excoffier and Lischer, 2010)を用い てΦst(Excoffier et al., 1992)を算出し,sequential Bonferroni 補正(Rice, 1989)を行 った.ハプロタイプ多様度(h)および塩基多様度(π)の算出には Arlequin ver. 3.5 を用いた.採集を行った9 つの水系ごとについて,Arlequin ver. 3.5 を用いて分子分 散分析(AMOVA)を行った.ただし,Φst,AMOVA,h,およびπの各分析では, 個体数が2 以下の地点(一宮,西尾 1)については解析に含めなかった. 結 果 165 個体のトウカイコガタスジシマドジョウの mtDNA の cyt b 領域 1045 bp の塩基 配列を決定した結果,35 種類のハプロタイプ(T1–T35)が得られた(Table Ⅲ-2).各地点における h とπは,それぞれ 0.385–0.871 と 0.0009–0.0071 であった (Table Ⅲ-1).得られたハプロタイプの塩基配列を用いて最尤法とベイズ法により 系統樹を推定したところ,ほとんど同じ樹形を示し,系統的に明瞭に区別できるク レードはみられなかった(Fig. Ⅲ−2).しかし,ハプロタイプの分布には地域的な 偏りがみられ,28 ハプロタイプ(T1–T28)が伊勢湾周辺域(長良川,揖斐川,木曽 川,庄内川,矢作川,櫛田川,江川水系)からのみ見出され,7 ハプロタイプ (T29–T35)が西静岡地域(都田川,馬込川,太田川水系)からのみ見出された.伊
Table III-2 Distribution of each haplotype for 16 sampling sites *G rounds for re gi ona l di vi sion d es cri be d in R es ul ts
Fig. III-2 Bayesian tree of the 1045 bp cytochrome b gene sequences of Cobitis minamorii
tokaiensis individuals from the Aichi-Gifu (gray circles), Mie (black circles), and
West-Shizuoka (white circles) regions. Numbers at nodes indicate maximum likelihood bootstrap probabilities (left) and Bayesian posterior probabilities (right) > 70 % (both parameters). The statistical parsimony network of C. m. tokaiensis is shown to the left of the tree. Pie charts indicate relative frequencies of haplotypes found in the three regions.
勢湾周辺域と西静岡地域間のハプロタイプの遺伝距離(p-distance)の平均±標準偏 差は,0.006 ± 0.001 であった.伊勢湾周辺域から得られた 28 ハプロタイプの多く は各水系に固有のものであり,8 ハプロタイプが長良川水系から,3 ハプロタイプが 揖斐川水系から,5 ハプロタイプが庄内川水系から,2 ハプロタイプが矢作川水系か ら,1 ハプロタイプが櫛田川水系から,1 ハプロタイプが江川水系からのみ見出され た(Table Ⅲ-2).しかし,ハプロタイプ T1 と T3 は伊勢湾周辺域に広く見られ,特 にハプロタイプT1 は 12 地点中 10 地点から見出された(Table Ⅲ-2).西静岡地域 からのみ得られた7 ハプロタイプのうち,2 ハプロタイプが太田川水系からのみ見出 された(Table Ⅲ-2).ハプロタイプネットワークを作成したところ,西静岡地域か ら得られたハプロタイプT29–T35 は,T31 を中心とする近縁なグループとしてまと まった(Fig. Ⅲ-2).伊勢湾周辺域から得られたハプロタイプ T1–T28 は,西静岡地 域から得られたハプロタイプT31 を挟み,2 つのグループ(T1 を中心とした 10 ハプ ロタイプのグループと,T3 を中心とした 18 ハプロタイプから成るグループ)に分 けられたが,それら2 つのハプロタイプグループの地理的分布に明瞭な違いはみら れなかった. 地点間のペアワイズΦstを計算した結果,西静岡地域の4 地点のうち 3 地点(浜松 1,浜松 2,森)は,ほとんどの伊勢湾周辺域の地点との組み合わせにおいて有意に 分化していた(Φst = 0.146–0.578:P < 0.01)(Table Ⅲ-3).ただし伊勢湾周辺域に 地理的に最も近い湖西(地点No.13)は,伊勢湾周辺域のどの地点とも有意な遺伝的 分化を示さなかった.また,三重県の3 地点のうち 2 地点(松阪 2,伊勢)も,多く の組み合わせにおいて有意に分化していた(Φst = 0.219–0.578:P < 0.01)(Table Ⅲ-3).ただし松阪 1 は,伊勢湾周辺域のどの地点とも有意な遺伝的分化を示さなかっ た. 水系間での遺伝的分化については,個体群を9 つの水系(Table Ⅲ-1)に分けて AMOVA 分析を行った.その結果,水系間での遺伝的変異がトウカイコガタスジシ マドジョウの遺伝的変異の21.35%(P < 0.01),水系内の地点間の遺伝的変異が
Table III-3 Pairwise Φst among local populations of Cobitis minamorii tokaiensis * G rounds for re gi ona l di vi sion d es cri be d in Re sul ts, **P < 0.01
12.67%(P < 0.01),地点内の遺伝的変異が 65.98%(P < 0.00)であり,いずれも有 意にゼロから離れていた(Table Ⅲ-4). 考 察 トウカイコガタスジシマドジョウのmtDNA ハプロタイプの系統樹を推定した結 果,系統的に明瞭に区別できるクレードは得られなかった.しかし,ペアワイズΦst の比較をした結果,西静岡地域と伊勢湾周辺域間,および三重県と伊勢湾周辺の他 地域(愛知県,岐阜県)の間では,多くの地点の組み合わせで有意な分化が見出さ れたことから,遺伝的に3 つの地域(西静岡地域,三重地域,愛知岐阜地域)に分 化していると考えられた(Table Ⅲ-2). 特に西静岡地域の4 地点は,固有のハプロタイプのみが分布することとペアワイ ズΦstに基づく集団分化の検定結果から,伊勢湾周辺域(三重地域と愛知岐阜地域) のトウカイコガタスジシマドジョウとは明確に分化していると考えられた.ただ し,これらのハプロタイプは系統的に伊勢湾周辺域から得られたハプロタイプグル ープに内包され,Φstでは湖西がどの地点とも有意な遺伝的分化を示さなかった.湖 西ではサンプル数が4 個体と少ないことから,Φstに有意差がみられなかったことも 考えられるが,いずれにせよ本亜種は地質年代的なスケールで比較的最近まで伊勢 湾周辺域と西静岡地域間で遺伝的に交流しており,遺伝的分化が生じたのは比較的 新しい時代であると考えられる.本亜種と同様に東海地方に分布する淡水魚類の中 では,河川源流域に生息する山地性のトウカイナガレホトケドジョウの系統地理的 パターン(Miyazaki et al., 2017)が,トウカイコガタスジシマドジョウの系統地理パ ターンとやや類似している.Miyazaki et al.(2017)によると,トウカイナガレホト ケドジョウのmtDNA の CR 領域の解析では伊勢湾周辺域と西静岡地域間で遺伝的差 異はほとんどなく,特に豊川水系(伊勢湾周辺域)と天竜川水系(西静岡地域)の 個体群からは同一のハプロタイプが得られており,上流域で生じた河川争奪によ り,両水系の個体群が遺伝的に交流したことが示唆されている.しかし,トウカイ
Table III-4 Hierarchical analysis of molecular variance (AMOVA) among regions of
コガタスジシマドジョウは主に河川下流域に生息する平地性の淡水魚類であること から,山地性のトウカイナガレホトケドジョウで推測された上流域の河川争奪が遺 伝的交流の要因とは考えにくい.また,同じ平地性の淡水魚類であるカワバタモロ コは,伊勢湾周辺域と西静岡地域に対応する明瞭な2 つの系統に分かれることが cyt
b 領域の遺伝的解析により明らかにされている(Watanabe and Mori, 2008;Watanabe
et al., 2014).カワバタモロコにみられた 2 系統は,弓張山地の隆起により分断され たと推測されている(Watanabe and Mori, 2008).弓張山地は更新世前期(約 180–80 万年前)に隆起したと考えられており(森山,1990),カワバタモロコの 2 系統間 で推定された分岐年代と矛盾しない(Watanabe et al., 2009,2014).しかし,トウカ イコガタスジシマドジョウについては,伊勢湾周辺域と西静岡地域に対応する明瞭 なクレードに分かれなかったことから,弓張山地の隆起以降に両地域間で遺伝的な 交流があったと考えられる.約60 万年前以降の時代は,約 10 万年のサイクルで氷 期と間氷期が交互に到来したとされている(北村,2010).この過去 60 万年間にお ける氷期の海水準は最終氷期と同程度であり,どの氷期でも海面が約100 m 低下し た(北村,2010).そのため,現在の遠州灘から渥美半島沿岸の河川が下流部で接 続することで遺伝的な交流が行われた可能性が考えられる.ただし,伊勢湾周辺域 と西静岡地域の河川が,海退期に下流域で接続していたという地形学的証拠は得ら れておらず,河川水系の接続は洪水など大規模な出水時の一時的なものであった可 能性もある.なお,カワバタモロコでは,西静岡地域の複数地点から伊勢湾周辺域 の系統に属するハプロタイプが得られている(Watanabe et al., 2009).もしこれらの ハプロタイプが自然分布であれば,カワバタモロコもトウカイコガタスジシマドジ ョウと同様に,弓張山地の隆起以降の河川下流域の接続によって伊勢湾周辺域と西 静岡地域間で遺伝的に交流した可能性が考えられる. 三重県の3 地点(三重地域)も,伊勢湾周辺域の他の地点とは遺伝的に分化して いることが,ハプロタイプの分布とペアワイズΦstの検定結果から考えられた.ただ し,三重県の3 地点から得られた 5 ハプロタイプ中 2 ハプロタイプ(T1 と T3)は,
長良川・揖斐川・庄内川・矢作川水系からも確認されている.伊勢湾周辺域では, 氷期に海水面が低下したことで,湾内に流入する水系が合流して単一の古水系とな ったと考えられている(森山,2004).ウシモツゴやカワバタモロコ,カワヒガイ などでは,この単一の古水系を介した伊勢湾流入河川間での遺伝的交流が示唆され ているため(Watanabe and Mori, 2008;鈴木ほか,2016),トウカイコガタスジシマ ドジョウの場合も,古水系を介した遺伝的交流によって共通のハプロタイプが分布 していると考えられる.また,ペアワイズΦstの検定結果では,松阪1 のみ伊勢湾周 辺域のどの地点とも有意な遺伝的分化を示さなかったが,松阪1 のサンプル数は 7 個体と少ないために有意差がみられなかった可能性がある.松阪1の個体群と他地 域との間の遺伝的分化の有無については,今後個体数を増やして解析する必要があ る. 三重県以外の伊勢湾周辺域(愛知岐阜地域)では,各水系でのみみられる固有の ハプロタイプが多数得られた.しかし,地点間のΦstの比較においては,山県と関 (長良川水系)の間でのみ遺伝的分化が示されたにすぎず,明確な集団構造は見出 せなかった.ただし,AMOVA 分析によって,水系間の遺伝的変異が 21.35%存在す ることが示された.本種の遺伝的変異の多く(65.98%)は各地点の個体群内にある ことから,相対的に水系間の分化は小さいものの,今後各地点の解析個体数を増や すことや,種内多型が多いとされるマイクロサテライトマーカーを利用するなどの 解析を行うことで,愛知岐阜地域内での集団構造が明確にできる可能性がある.
第四章 伊勢湾周辺域におけるアカハライモリの遺伝的集団構造
緒 言
アカハライモリCynops pyrrhogaster は,有尾目イモリ科に属し,本州,四国,九 州に分布する日本固有種である(関,2016).本種の地理的変異は,形態的特徴と 繁殖行動の違いから6 種族に(Kawamura and Sawada, 1959;Sawada, 1963a,b),ア ロザイム分析によって5 集団に(Hayashi and Matsui, 1988,1990),ミトコンドリア DNA(mtDNA)の種内系統から 4 系統群(クレード)に(Tominaga et al., 2013; Tominaga et al., 2015)分けられる.これらの研究の中では,mtDNA の 4 つのクレー ド内に,さらに複数の地理的なサブクレードも見られるため(Tominaga et al., 2013; Tominaga et al., 2015),アカハライモリの地理的分化を検出する上で mtDNA は優れ たマーカーであると言える.本州中部のアカハライモリは,mtDNA の 4 クレードの うち主に中部日本(CENTRAL)クレードが分布する.中部日本クレードは,中部地 方から中国地方までの広域に分布し(Tominaga et al., 2013),中部地方全域から近畿 地方東部に分布する中部(Chubu)サブクレードと, 近畿地方西部から中国地方に かけて分布する近畿(Kinki)サブクレードに大別される(Tominaga et al., 2013). また,中部サブクレード内では,伊豆半島中央部に生息する集団のハプロタイプ (中部伊豆系統Central Izu lineage)が他の中部サブクレードのハプロタイプとは区 別できることが知られている(Tominaga et al., 2015).このように,アカハライモリ のmtDNA 系統は地域ごとに細かく分かれており,伊勢湾周辺域には上記の内,中部 サブクレードが広く分布するとされている.しかし,これまでの研究では伊勢湾周 辺域における調査地点が少なく,この地域における詳細な遺伝的集団構造は明らか ではない.アカハライモリは,低地から山地の水田,池,湿地,川岸の水たまりな どの止水域で見られ(関,2016),水域間の移動はあまりないという報告もあるた め(竹内ほか,2008),伊勢湾周辺域における水生生物の遺伝的集団構造の形成要 因を明らかにするには好材料であると考えられる.
材料と方法
2014 年から 2015 年にかけて,伊勢湾周辺域を中心とした合計 20 地点 99 個体[美 濃加茂(Minokama)11 個体;多治見(Tajimi)5 個体;関(Seki)6 個体;岐阜 1 (Gifu1)2 個体;中津川(Nakatsugawa)9 個体;八百津(Yaotsu)9 個体;瀬戸 (Seto)4 個体;矢並(Yanami)3 個体;根尾(Neo)9 個体;新城(Shinshiro)4 個 体;恵那(Ena)1 個体;高山(Takayama)1 個体;鳳来寺(Horaiji)3 個体;志摩 (Shima)9 個体;鳥羽(Toba)4 個体;度会(Watarai)5 個体;上石津
(Kamiishizu)1 個体;谷汲(Tanigumi)5 個体;日野(Hino)2 個体;高島
(Takashima)6 個体]のアカハライモリを採集した(Fig IV-1, Table IV-1).採集し た個体は,尾の先端の一部を99.5 %エタノール中に浸漬し,DNA の抽出を行うま で–80 °C の超低温フリーザ内で保管した.
DNA 抽出には,Quick Gene DNA Tissue Kit S(クラボウ社)を用いた.mtDNA の ND6,tRNA-glu,および cyt b 領域を対象とした PCR 増幅と塩基配列の決定には, sala_cytb_F_N1(5’-AAY TAC CCC AYG CCT CMG GAT AWG GYT C-3’)と
cytb_R4_cynops(5’-GGT RGT GCC CGG GYG CAC CTA GAG AAG G-3’)のプライ マー対(Tominaga et al., 2010)を用いた.PCR には,Crimson Taq PCR sampler のバ ッファー(ニューイングランドバイオラボ社)とEx Taq DNA ポリメラーゼ(タカ ラバイオ社)を使用し,常法に従ってサーマルサイクラ―で95 °C 1 分,55 °C 1 分,72 °C 2 分の温度サイクルを 30 回繰り返して増幅を行った.PCR 産物について はExoSAP-IT キット(GE ヘルスケアジャパン社)を用いて処理した後,増幅した 時と同じプライマーに加え,SLD_cytb_H2(5’-TAG GCR AAT ARR AAR TAY CAY TCT GG-3’)(Tominaga et al., 2010)も用いて,BigDye Terminator Cycle Sequencing Kit ver. 3.1(アプライドバイオシステムズ社)によるシークエンス反応を行い,ABI 3130 Genetic Analyzer で塩基配列を決定した.得られた塩基配列については個体ごと
Fig. IV-1 Sampling localities of Cynops pyrrhogaster. Localities with black circles indicate
collection sites of the Widespread lineage, white circles indicate collection sites of the East Kinki-West Ise Bay lineage, and gray circles indicate collection sites of Central Izu lineage (this lineage follows Tominaga et al., 2013). The asterisks indicate collection sites used by Tominaga et al. (2013).
Site
no. Collection site
Lineage*
n Haplotype (n)
1 Minokamo, Gifu Others 11 C1 (3), C10 (1), C11 (1), H38 (6) 2 Tajimi, Gifu Others 5 C2 (2), C3 (1), C4 (1), C5 (1)
3 Seki, Gifu Others 6 C6 (1), C7 (1), C8 (1), C9 (2), C10 (1) 4 Gifu1, Gifu Others 2 C20 (1), H38 (1)
5 Nakatsugawa, Gifu
Others
9 C2 (4), C12 (3), C20 (1), H38 (1)
6 Yaotsu, Gifu Others 9 C13 (1), H38 (8) 7 Seto, Aichi Others 4 C2 (1), C4 (2), C19 (1) 8 Yanami, Aichi Others 3 C2 (2), C4 (1)
9 Neo, Gifu Others 9 C9 (1), C10 (1), C20 (1), C21 (1), C22 (1), H38 (2), H43 (2)
10 Shinshiro, Aichi Others 4 C27 (4) 11 Ena, Gifu Others 1 C4 (1)
12 Takayama1, Gifu
Others
1 H38 (1)
13 Houraiji, Aichi Others 2 C2 (1), C27 (1)
Mikawa-Shizuoka 1 C28 (1)
14 Shima, Mie
Shima-Nagoya 9 C23 (6), C24 (1), C25 (2)
Table IV-1 List of sampling localities, number of specimens (n), and haplotypes in Cynops
15 Toba, Mie
Shima-Nagoya 4 C23 (3), C26 (1)
16 Watarai, Mie East Kinki-
West Ise Bay 1 C30 (1)
Kii Mts. 4 C29 (1), C31 (1), C32 (1), H53 (1)
17 Kamiishizu, Gifu
East Kinki-
West Ise Bay 1 C33 (1)
18 Tanigumi, Gifu Others 4 C15 (1), C16 (1), C17 (1), C18 (1)
East Kinki-
West Ise Bay 1 C14 (1) 19 Hino, Shiga Others 1 C38 (1)
East Kinki-
West Ise Bay 1 H65 (1)
20 Takashima, Shiga
Others
3 C4 (1), C34 (1), C36 (1)
East Kinki-
West Ise Bay 1 C35 (1)
Kinki 2 C37 (1), H70 (1) 21 Suzu, Ishikawa Others - AB754652: H59 22 Himi, Toyama Others - AB754652: H59 23 Nomi, Ishikawa Others - AB754678: H55
24 Taroyama, Toyama
Others
- AB754631: H38
25 Arimine,Toyama Others - AB754631: H38 26 Sakaki, Nagano Others - AB754634: H41
27 Azumino, Nagano
Others
- AB754656: H63
28 Hida, Gifu Others - AB754631: H38, AB754635: H42
29 Takayama2, Gifu
Others
- AB754631: H38
30 Shirakawa, Gifu Others - AB754644: H51 31 Ono, Fukui Others - AB754649: H56 32 Takefu, Fukui Others - AB754654: H61
33 Minamiechizen, Fukui
Others
- AB754637: H44, AB754636: H43
34 Ibigawa, Gifu Others - AB754637: H44, AB754636: H43, AB754655: H62
35 Nagahama, Shiga
Others
- AB754655: H62
36 Ibuki, Shiga Others - AB754636: H43
East Kinki-
West Ise Bay - AB754658: H65
37 Hikone, Shiga East Kinki-
West Ise Bay - AB754658: H65 38 Imazu, Shiga Others - AB754653: H60
39 Yokkaichi, Shiga
East Kinki-
West Ise Bay - AB754658: H65
40 Eigenji, Shiga East Kinki-
West Ise Bay - AB754658: H65
41 Koga, Shiga East Kinki-
42 Gifu2, Gifu Others - AB754645: H52 43 Hiraya, Nagano Others - AB754633: H40
44 Nagoya, Aichi
Shima-Nagoya - AB754639: H46 45 Shitara, Aichi Others - AB754632: H39
46 Misakubo, Shizuoka Mikawa-Shizuoka - AB754643: H50 47 Shizuoka, Shizuoka
Mikawa-Shizuoka - AB754641: H48, AB754642: H49
48 Kakegawa, Shizuoka
Others
- AB754638: H45
49 Izu, Shizuoka Central Izu** - AB754662: H69 50 Miyama, Kyoto Others - AB754651: H58 51 Iwakura, Kyoto Others - AB754650: H57
52 Yase, Kyoto East Kinki-
West Ise Bay - AB754661: H68
53 Nara, Nara East Kinki-
West Ise Bay - AB754659: H66
54 Ibaragi, Osaka East Kinki-
West Ise Bay - AB754660: H67
55 Tondabayashi, Osaka
Others
- AB754630: H37
56 Taiki, Mie Kii Mts. - AB754646: H53
57 Kihoku, Mie Kii Mts. - AB754646: H53, AB754647: H54
58 Minabe, Wakayama
59 Wakayama, Wakayama
Others
- AB754657: H64
”C” indicates haplotypes found in this study
“H” indicates same haplotypes with Tominaga et al. (2013) Site number 21–59 were cited from Tominaga et al. (2013) Gifu2 is equivalent to “Gifu” of Tominaga et al. (2013)
Takayama2 is equivalent to “Takayama” of Tominaga et al. (2013) * The grounds of lineages were described in Results
に結合させて,不一致が無いことを確認した上でDDBJ,EMBL,GenBank のデータ ベースに登録した(登録番号 LC520004–LC520041).
比較のための多重整列には,CLUSTAL X Version 1.83(Thompson et al., 1997)を 用い,系統樹の推定には,最尤法とベイズ法を用いた.最尤法には,raxmlGUI, Version 1.3(Silvestro and Michalak, 2012)を用いた.塩基配列における各領域とコド ンの進化速度の違いを考慮し,ND6 の 1 番目,2 番目,3 番目のコドン,tRNA-glu, cyt b の 1 番目,2 番目,3 番目のコドンにパーティションを設定して GTR+G モデル を適用した.各枝の信頼性については1000 回のブートストラップ確率を求めた.ベ イズ法には,MrBayes, Version 3.2.6 (Ronquist et al., 2012)を用いた.Kakusan4 (Tanabe,2011)を用いてベイズ情報量規準(BIC)に基づいたモデルテストを行 い,ND6 の 1 番目,2 番目,3 番目のコドン,tRNA-glu,cyt b の 1 番目,2 番目,3 番目のコドンにパーティションを設定して,HKY85+G,F81+G,HKY+G,K80+G, K80+G,HKY,GTR+G の各モデルを適用した.解析にあたっては,マルコフ連鎖モ ンテカルロ(MCMC)法によってマルコフ連鎖を 10000000 世代繰り返し,100 世代 ごとにサンプリングを行い,burnin を 25000 世代に設定した.その際に,Average Standard Deviation of Split Frequencies(ASDSF)が 0.01 未満になったことで MCMC が定常状態に達したと判断した.系統樹には,Tominaga et al.(2013)で明らかにさ れた中部日本クレードの中部サブクレードに含まれるハプロタイプの相同配列も用 いた(Table Ⅳ-1).系統樹の外群には,Tominaga et al.(2013)で明らかにされた 中部日本クレードの近畿サブクレード(AB754663:H70)および西日本
(WESTERN)クレード(AB754679)の相同配列を用いた.地理的に遺伝的分化が 示唆された地域間について,MEGA6(Tamura et al., 2013)を用いて,p-distance によ る遺伝距離を求め,1000 回のブートストラップ検定により標準誤差を求めた.
結 果
99 個体のアカハライモリの mtDNA の ND6,tRNA-glu,および cyt b 領域 1402 bp の塩基配列を決定した結果,本研究で新たに得られたハプロタイプが38 種類(C1– C38),Tominaga et al.(2013)と共通のハプロタイプが 5 種類(H38,H43,H53, H65,H70)得られた(Table Ⅳ-1,Ⅳ-2).得られたハプロタイプの塩基配列を用い て最尤法とベイズ法により系統樹を推定したところ,ほとんど同じ樹形を示した (Fig. Ⅳ−2).得られた樹形は,Tominaga et al.(2013)と同様に中部日本クレード 内に2 つのサブクレード(中部,近畿)が認められた.近畿サブクレードについて は,ブートストラップ確率,事後確率共に高い値を示した(100 %)ものの,中部サ ブクレードに相当するクレードのブートストラップ確率,事後確率は共に低い値を 示した(< 60 %).ただし,Tominaga et al.(2013)では共に高い値が得られている ことから,本研究でも中部サブクレードを1 つの単系統群として扱った.本研究で 得られたハプロタイプのうち,41 種類は中部サブクレードに,2 種類は近畿サブク レードに含まれた.中部サブクレードについては,Tominaga et al.(2013)と同様に 中部伊豆系統が最初に分岐した.次にブートストラップ確率,ベイズ法の事後確率 共に83 %以上で支持される系統(東近畿−西伊勢湾系統)と,ブートストラップ確 率は低いものの(53 %),事後確率は 98 %で支持される系統(広域系統)に分岐し た.広域系統内では,ブートストラップ確率,事後確率共に高い値(> 85 %)で支 持された地域系統が3 つ(志摩−名古屋,三河−静岡,紀伊山地地域系統)認められ た(Fig. Ⅳ−2).その他の 27 種類のハプロタイプは,Tominaga et al.(2013)から得 られた19 種類のハプロタイプも含めて系統的に明瞭に区別できなかった(以下,そ の他のハプロタイプグループとする).各系統の分布範囲については,東近畿−西伊 勢湾系統は度会,上石津,谷汲,日野,高島に加え,Tominaga et al.(2013)におけ る伊吹(Ibuki),彦根(Hikone),八日市(Yokaichi),永源寺(Eigenji),甲賀 (Koga),八瀬(Yase),奈良(Nara),茨木(Ibaragi)から得られたハプロタイ プも含まれた.志摩−名古屋地域系統は,志摩および鳥羽に加え,Tominaga et al.
Table IV-2 Distribution of each haplotype for 20 sampling sites * T he grounds of re gi ona l di vi sion w ere d es cri be d in Re sul ts
Fig. IV-2 Bayesian tree of the 1402 bp NADH6, tRNA-glu, and cytochrome b genes
sequences of Cynops pyrrhogaster individuals from mainly the Ise Bay region. Numbers at nodes indicate maximum likelihood bootstrap probabilities (left) and Bayesian posterior probabilities (right) > 70 % (both parameters). ”C” indicates haplotypes found in this study. “H” indicates same haplotypes with Tominaga et al. (2013). Accession numbers (AB…) indicates cited from Tominaga et al. (2013). *Cited from Tominaga et al. (2013). **Cited from Tominaga et al. (2015).
(2013)における名古屋(Nagoya)から得られたハプロタイプも含まれた.三河−静 岡地域系統は鳳来寺に加え,Tominaga et al.(2013)における静岡(Shizuoka),水 窪(Misakubo),みなべ(Minabe)から得られたハプロタイプも含まれた.紀伊山 地地域系統は度会に加え,Tominaga et al.(2013)における大紀(Taiki),紀北 (Kihoku)から得られたハプロタイプも含まれた.その他のハプロタイプグループ は,系統的には明瞭に区別できなかったものの,本研究で得られた27 種類のうち 17 種類のハプロタイプは各地点に固有であり,C1,C11 が美濃加茂のみから,C5 が多 治見のみから,C6−8 が関のみから,C12 が中津川のみから,C13 が八百津のみか ら,C19 が瀬戸のみから,C3 が矢並のみから,C21−C22 が根尾のみから,C15−C18 が谷汲のみから,C38 が日野のみから見出された(Table Ⅳ-2).しかし,広域に分 布するハプロタイプも見られ,C4 は伊勢湾周辺域内の 4 地点,C2 は伊勢湾周辺域 外の高島を含む5 地点,H38 は Tominaga et al.(2013)で確認された太郎山 (Taroyama),有峰(Arimine),飛騨(Hida),高山 2(Takayama2)を含めて伊勢 湾周辺域内外の合計10 地点から見出された.近畿サブクレードについては,本研究 では高島からのみ得られた. 複数の系統/地域系統が同所的に見られた地点として,三河−静岡地域系統は鳳来 寺においてその他のハプロタイプグループと,東近畿−西伊勢湾系統は度会において 紀伊山地地域系統と,谷汲・日野・高島においてその他のハプロタイプグループと 同所的に確認された(Table Ⅳ-2).高島からは,近畿サブクレードのハプロタイプ も同所的に確認された(Table Ⅳ-2).ハプロタイプ間の遺伝距離(p-distance)は, 中部サブクレード内に確認された3 系統(広域,東近畿−西伊勢湾,中部伊豆系統) の間では0.009−0.0019 の間であり,広域系統内に確認された 3 地域系統(志摩−名古 屋,三河−静岡,紀伊山地地域系統)間では0.007−0.009 の間であった(Table Ⅳ-3).
Widespread East Kinki-West Ise Bay East Kinki-West Ise Bay 0.009±0.002
Central Izu 0.019±0.003 0.016±0.003
Shima-Nagoya Mikawa-Shizuoka Kii Mts. Mikawa-Shizuoka 0.007±0.002
Kii Mts. 0.008±0.002 0.009±0.002
Others 0.006±0.001 0.006±0.001 0.008±0.002
Table IV-3 Genetic distance among lineages within Chubu subclade and lineages