わが国で発生したA型および B 型乳児ボツリヌス
症起因菌の遺伝学的解析
著者
梅田 薫
内容記述
学位授与大学: Osaka Prefecture University(大阪
府立大学), 学位の種類: 博士(獣医学), 学位記番
号: 論獣第143号, 学位授与年月日: 2010-03-31,
指導教員: 小崎俊司.
大阪府立大学博士(獣医学)学位論文
わが国で発生したA型および B 型乳児ボツリヌス症
起因菌の遺伝学的解析
梅田
薫
目次 ページ 緒論・・・・・・・・・・・・・・・・・・・・・・・ 1 第 1 章 A 型乳児ボツリヌス症分離株の分子疫学的解析 緒言・・・・・・・・・・・・・・・・・・・・・・・ 6 材料と方法・・・・・・・・・・・・・・・・・・・・ 8 結果・・・・・・・・・・・・・・・・・・・・・・・ 11 考察・・・・・・・・・・・・・・・・・・・・・・・ 13 第 2 章 B 型乳児ボツリヌス症分離株の毒素遺伝子の解析 緒言・・・・・・・・・・・・・・・・・・・・・・・ 16 材料と方法・・・・・・・・・・・・・・・・・・・・ 18 結果・・・・・・・・・・・・・・・・・・・・・・・ 21 考察・・・・・・・・・・・・・・・・・・・・・・・ 26 第 3 章 Osaka05 株が産生する B 型毒素の抗原性の解析 緒言・・・・・・・・・・・・・・・・・・・・・・・ 30 材料と方法・・・・・・・・・・・・・・・・・・・・ 31 結果・・・・・・・・・・・・・・・・・・・・・・・ 33 34 考察・・・・・・・・・・・・・・・・・・・・・・・
第 4 章 連続継代培養によるボツリヌス B 型菌の毒素産生性への影響 緒言・・・・・・・・・・・・・・・・・・・・・・・ 37 材料と方法・・・・・・・・・・・・・・・・・・・・ 38 結果・・・・・・・・・・・・・・・・・・・・・・・ 39 考察・・・・・・・・・・・・・・・・・・・・・・・ 42 総合考察・・・・・・・・・・・・・・・・・・・・・ 44 結論・・・・・・・・・・・・・・・・・・・・・・・ 48 表・・・・・・・・・・・・・・・・・・・・・・・・ 49 図・・・・・・・・・・・・・・・・・・・・・・・・ 63 謝辞・・・・・・・・・・・・・・・・・・・・・・・ 79 参考文献・・・・・・・・・・・・・・・・・・・・・ 80
緒論 ボツリヌス菌(Clostridium botulinum)は、耐熱性芽胞を形成する偏性嫌 気性グラム陽性桿菌であり、強力な神経毒素(BoNT)を産生する (12)。本 菌は毒素の抗原性の違いにより A 型~G 型の 7 型に分類されている。ほと んどの菌株は 1 種類の型の毒素を産生するが、例外的に 2 種類の毒素を産生 する菌が分離されている (29)。また、本菌は生化学的性状および rRNA 遺 伝子の相同性から 4 つの群(第 I 群~第 IV 群)に分類することができるが 毒素型による分類とは一致しない (14, 47)。第 I 群にはすべての A 型菌とタ ンパク分解性 B、F 型菌が属し、第 II 群にはすべての E 型菌とタンパク非分 解性B、F 型菌が属している。第 III 群には C、D 型菌が属し、第 IV 群とし てはG 型菌のみが属している。また、一部の C. butyricum、C. baratii はそれ ぞれE 型、F 型毒素と極めて類似した毒素を産生する(Table 1)。ヒトにボ ツリヌス症を引き起こすのは A、B、E、F および G 型菌で、C および D 型 菌はトリや家畜などに中毒を起こす (27, 66)。 BoNT は分子量約 150 kDa の 1 本鎖ポリペプチドの形で産生され、培養液 中あるいは消化管内でトリプシンなどのタンパク分解酵素により分子内で
解離が起こり、ジスルフィド結合で結ばれた約50 kDa の軽鎖(Light chain; L)
と約100 kDa の重鎖(Heavy chain; H)の二本鎖構造に変化する(Figure 1)。
重鎖はN 末端領域(HN)およびC 末端領域(HC)の 2 つの機能的ドメイン に分けられ、重鎖 C 末端領域はさらに 2 つのサブドメイン(HCNおよび HCC) に分けられる (12, 77)。BoNT は生体内ではコリン作動性末梢神経に作用し、 アセチルコリンの遊離を阻害することにより麻痺を引き起こす。それぞれの ドメインが毒性発現に関連する一連の作用を担っている。HC ドメインは神 経細胞受容体への結合(特に HCC サブドメインが結合には重要と考えられ
ている)、HNドメインは神経細胞内移行に関与すると考えられており、L ド メインは亜鉛依存性プロテアーゼで、神経伝達物質の開口放出に関連するタ ンパク質を特異的に切断する (12, 47, 69)。 すべての型の BoNT は、菌体内で無毒成分との複合体を形成し、菌融解時 に放出される。複合体毒素は分子量の違いにより、LL 毒素(分子量約 90 万)、L 毒素(分子量約 50 万)および M 毒素(分子量約 30 万)に分けられ る。LL 毒素および L 毒素は血球凝集活性を持つが、M 毒素は持たない。A 型菌は 3 種類(LL、L、M)の毒素、B、C、D 型菌は 2 種類(L、M)の毒 素、E、F 型菌は M 毒素、G 型菌は L 毒素のそれぞれ 1 種類のみを産生する (35, 57)。 boNT 遺伝子は全長約 3.8 kbp であり、その上流には全長約 5 kbp~7 kbp
の無毒成分遺伝子群(ヘマグルチニン遺伝子[ha70, ha33, ha17]、転写調節因 子遺伝子[botR]、nontoxic-nonhemagglutinin 遺伝子[ntnh]、機能不明の遺伝子 [orf1, orf2, orf3, p47])がコードされている。boNT および無毒成分遺伝子群 は boNT 遺伝子クラスターと定義されている (23, 73)。
最近、boNT 遺伝子塩基配列の解析が進められ、A、B、C、D および E 型
毒素は、同一毒素型内におけるBoNT 分子の毒素アミノ酸残基が多様性を持
つことから、各型内で複数の subtype に分類されると報告された (29, 74)。
A 型毒素(BoNT/A)は 5 種類の subtype(A1、A2、A3, A4 および A5)に分 類されている (5, 18, 23)。BoNT/B は、第 I 群 B 型菌産生毒素が 3 種類(B1、 B2 および B3)、第 II 群 B 型菌産生毒素が 1 種類(Nonproteolytic)、B 型以
外の毒素遺伝子を保有する B 型菌(A[B]、Ab、AB、Bf など)産生毒素が 1
種類(Bivalent)の 5 種類の subtype に分類されている (29)。BoNT/C および
BoNT/D では、それぞれ重鎖 C 末端領域(HC)が BoNT/D および BoNT/C と
る (29, 52)。BoNT/E は、E型菌産生 BoNT/E が 4 種類(E1、E2、E3 および E6)、C. butyricum 産生 BoNT/E が 2 種類(E4 および E5)の 6 種類の subtype
に分類されている (13)。 ヒトのボツリヌス症は発症機序により、食餌性ボツリヌス症、乳児ボツ リヌス症、創傷性ボツリヌス症および成人腸管定着性ボツリヌス症の4 種類 に分類される (27, 66, 75)。食餌性ボツリヌス症は、いわゆる古典的なボツ リヌス中毒であり、食品内に産生された毒素を摂取することにより発症する 典型的な食品内毒素型食中毒である (27, 66, 75)。摂取後 2~40 時間の潜伏 期のあと、複視、眼瞼下垂、口渇、嚥下困難、便秘、全身の筋弛緩などの症 状を呈し、重度の場合は呼吸筋の麻痺による呼吸不全で死に至る。致死率は 10~20%と他の細菌性食中毒と比較して高い。起因菌として、第 I 群および
第II 群の A、B、E、F 型菌、まれに BoNT/E 産生性 C. butyricum が分離され
る (47, 61, 85)。 乳児ボツリヌス症は、主として 1 歳未満の乳児がボツリヌス芽胞を経口 的に摂取後、腸管内で発芽、増殖し、産生された毒素を吸収することによっ て発症する感染型の疾患である (6, 16, 22)。本症は 1976 年に初めて米国で 報告された (51, 62)。乳児にのみ発症する原因として、腸内細菌叢が成人と 比較して未発達であることから、成人では消化管に定着しないボツリヌス菌 が定着しやすいためと考えられている。症状は便秘傾向に始まり、哺乳力低 下、無表情、眼瞼下垂、頸部筋肉の弛緩によって頭部を支えられなくなるこ となどが認められ、全身の筋力低下をきたす。呼吸障害が生じて重症化する と死に至ることもあるが、致死率は 2%程度とボツリヌス食中毒に比べて低 い。大部分の乳児ボツリヌス症例からは、第I 群菌の A 型、B 型菌が分離さ れることが知られている (47)。これまでに D 型、E 型、F 型菌による本症 の発生はなく、E 型、F 型毒素と極めて類似した毒素を産生する C. butyricum
あるいは C. baratii が分離された症例が数例報告されている (2, 26)。また、 わが国で C 型菌による乳児ボツリヌス症が 1 例報告されている (58)。創傷 性ボツリヌス症は、傷口から感染したボツリヌス菌の芽胞が創傷部位で発芽 し、局所で産生された毒素により中毒症状が引き起こされる疾患である (27, 66, 75)。野外での運動や作業中の感染が主であったが、近年は薬物常用者の 注射痕から感染がおきた症例が欧米でしばしば報告されている (60)。成人 腸管定着性ボツリヌス症は、1 歳以上の子供と成人において、乳児ボツリヌ ス症と同様腸管内でボツリヌス菌が定着、増殖して毒素を吸収し発症する疾 患である (27, 66, 75)。発症は外科手術や抗菌薬の投与による患者の腸内細 菌叢の破壊や菌交代現象に起因すると考えられている。 わが国では、ボツリヌス食中毒および乳児ボツリヌス症の発生が認めら れている (32)。乳児ボツリヌス症は、初めての国内症例が報告された 1986 年から2007 年までに 24 症例が報告されている(Table 2)。その起因菌は、 A 型菌が 16 症例、B 型菌が 3 症例、C 型菌が1症例、BoNT/E 産生性 C. butyricum が1症例、不明が 3 症例である。1986 年から翌 1987 年にかけて、 ハチミツを感染源とする 10 症例が連続して報告され、毒素型が確認できた 7 症例はすべて A 型であった。その後、当時の厚生省から 1 歳未満の乳児に ハチミツを与えないよう指導する通達が出されたこともあり、ハチミツを原 因とする同症の発生は報告されなくなった。しかし1990 年代以降、感染源 が不明な症例の発生が認められており、その毒素型は A 型だけでなく、B 型、C 型、BoNT/E 産生性 C. butyricum と多様である。 ボツリヌス食中毒は、1951 年から 2007 年までに、117 事例の発生が報告 されている(Table 3)。1990 年頃までは主に北海道、東北地方の伝統食品で ある魚の発酵食品(いずし、きりこみなど)を原因とする E 型食中毒事例 が毎年数件報告されていた。しかし 1990 年代以降、輸入食品や容器包装詰
低酸性食品を原因とする A 型および B 型食中毒事例や原因食品が不明な A 型事例が散発的に発生している。 本研究では、わが国で 1990 年以降発生動向が変化しつつある A 型および B 型乳児ボツリヌス症の起因菌を中心に遺伝学的解析を行った。第 1 章では、 簡便な分子型別法として boNT/A 遺伝子クラスター型別 Multiplex PCR 法を 確立し、A 型乳児ボツリヌス症分離株の boNT/A 遺伝子クラスター型別およ
びPulsed-Field Gel Electrophoresis(PFGE)型別を実施し、分子疫学的解析
行った。第2 章では、B 型乳児ボツリヌス症分離株について、毒素遺伝子塩
基配列の分子系統解析、PFGE 型別、boNT/B 遺伝子を検出する Southern blot hybridization を実施し、毒素遺伝子の解析および分子疫学的解析を行った。
また、BoNT/B subtype 型別 Multiplex PCR 法を確立した。第 3 章では、第 2
章で毒素遺伝子の分子系統解析から新たなBoNT/B subtype であると考えら れた乳児ボツリヌス症分離株 Osaka05 株産生毒素の抗原性の解析を行った。 第 4 章では、第 2 章で明らかにした B 型菌の毒素遺伝子の存在位置の多様 性が、毒素産生性および毒素プラスミドの安定性に及ぼす影響について検討 した。 以上の解析から、わが国で発生した乳児ボツリヌス症例の背景を明らか にし、新たな分子型別法を検討すると共に、毒素遺伝子、毒素抗原性や毒素 産生性などを調べることにより、ボツリヌス菌および産生毒素の遺伝学的多 様性の解明を試みた。
第1章 A 型乳児ボツリヌス症分離株の分子疫学的解析 緒言 乳児ボツリヌス症は、1976 年に世界で初めての症例がアメリカで報告さ れてから 30 年以上が経過し、現在日本および欧米においては、ボツリヌス 症の中で最も発生件数が多い疾患となっている (22, 41)。わが国ではこれま でに 24 症例の発生が報告されているが、中でも A 型菌による症例は 16 症 例と最も多い(Table 2)。わが国で初めての症例が報告された 1986 年から翌 1987 年にかけて、全国各地で連続して 10 症例が報告されたが、毒素型が確 認できた 7 症例はすべて A 型であった。この期間の症例の患者にはすべて ハチミツ摂取歴があり、またハチミツからもボツリヌス菌が検出されたため、 当時の厚生症はハチミツが乳児ボツリヌス症の感染源であることを認め、1 歳未満の乳児にハチミツを与えないよう指導する通達を公布した(昭和 62 年 10 月;乳児ボツリヌス症の予防対策について)。その結果、1989 年を最 後にハチミツが関連すると考えられる同症の発生は報告されていない。一方、 1990 年以降ハチミツ摂取歴のない症例が報告されているが、2004 年までは 数年に1 症例程度の散発的な発生のみであった。しかし、2005 年以降 2007 年までは年に 2 症例ずつの発生が報告されており、現在増加傾向にあると言 える。1990 年以降のハチミツ摂取歴の無い症例の毒素型は、A 型 7 症例、B 型3 症例、C 型 1 症例、BoNT/E 産生性 C. butyricum 1 症例と、ハチミツ摂 取歴がある症例と比較して多様性に富んでいる。その内、感染源が推定され ているのは、1996 年東京で発生した A 型症例(自家製野菜スープ)および 2006 年宮城県で発生した A 型症例(粉ミルクを調製する井戸水)の 2 症例 のみであり、A 型 5 症例を含む 10 症例の感染源は不明である (2, 3, 32, 33, 37,
58, 81)。 一方、わが国で発生した A 型ボツリヌス食中毒において最も大規模な事 例は、1984 年の熊本県産辛子レンコンによる事例であるが、1990 年以降、 A 型事例の散発的な発生が報告されている(Table 3)。1999 年には、発生場 所(東京都、千葉県および大阪市)や感染源(それぞれ感染源不明、容器包 装詰ハヤシライスの具、感染源不明)が異なる 3 事例が連続して発生してい る (32, 33, 40)。 BoNT/A は現在 5 種類の subtype(A1~A5)に分類される。ほとんどの株 がA1 もしくは A2 に属し、A3, A4 および A5 は各 1 株のみが報告されてい る (5, 18, 29)。boNT 遺伝子およびその上流に位置する無毒成分遺伝子群は boNT 遺伝子クラスターと定義されており、それらの遺伝子構成は毒素型お
よび毒素subtype によって、HA クラスターと Orfx クラスターの 2 つに分類
される (19, 34)。HA クラスターは、boNT 遺伝子の上流に、ヘマグルチニン
遺 伝 子 (ha70, ha33, ha17) 、 転 写 調 節 因 子 遺 伝 子 (botR) 、
nontoxic-nonhemagglutinin 遺伝子(ntnh)が存在し、LL 毒素、L 毒素、M 毒素 を形成するタンパク群をコードしている。一方、Orfx クラスターは、ntnh および botR は保有しているが、ヘマグルチニン遺伝子を持たず、代わりに 機能不明の orfx 遺伝子(orfx1, orfx2, orfx3)および p47 遺伝子が存在し、そ
の産物は M 毒素のみを形成する。BoNT/A1、BoNT/B、BoNT/C、BoNT/D お
よび BoNT/G 産生菌は HA クラスターを保有し、BoNT/A2、BoNT/A3、
BoNT/A4、BoNT/E および BoNT/F 産生菌は Orfx クラスターを保有している (34, 57)。
1986 年から 1987 年にかけてわが国で発生した A 型乳児ボツリヌス症分離
株と、保存 A 型菌株や A 型ボツリヌス食中毒分離株とは、産生毒素の抗原
毒分離株は LL 毒素、L 毒素および M 毒素を産生するが、乳児ボツリヌス症
分離株は M 毒素のみを産生することも明らかされた (64, 79)。その後、乳
児ボツリヌス症分離株産生毒素は BoNT/A2、保存菌株や食中毒分離株産生
毒素はBoNT/A1 と定義された (15, 74)。
Franciosa ら (23) は、BoNT/A1 および HA クラスターを保有する A 型菌 を boNT/A 遺伝子クラスター1、BoNT/A2 および Orfx クラスターを保有す
るA 型菌を boNT/A 遺伝子クラスター2、BoNT/A1 および毒素産生を伴わな
い(不活性な)B 型毒素遺伝子、HA クラスターおよび Orfx クラスターを保
有する A 型菌を boNT/A 遺伝子クラスター3 と定義し、PCR および PCR
restriction fragment length polymorphism (PCR-RFLP) 法を用いた型別法を報
告した(Figure 2)。boNT/A 遺伝子クラスター型別は A 型菌の分子型別法と しても応用されている (23)。 本章では、Multiplex PCR 法による簡便な boNT/A 遺伝子クラスター型別 法を新たに構築するとともに、わが国で発生した乳児ボツリヌス症の背景を 明らかにするため、boNT/A 遺伝子クラスター型別および PFGE を用いて様々 な由来の A 型菌株の遺伝子型別を実施し、分子疫学的解析を行った。 材料と方法 供試菌株と培養条件: わが国で発生した A 型乳児ボツリヌス症分離株 10 株、A 型食中毒分離株 4 株、米国カリフォルニア州で発生した乳児ボツリヌス症分離株 6 株、食品 分離株2 株、および保存株 5 株の、計 27 株の A 型菌を供試した (Table 4)。 他の Clostridium 属菌 12 株(Figure 3)を PCR のコントロールとして供試し た。各菌株は0.3% glucose および 0.2% soluble starch (Difco, Becton Dickinson
and Co., Franklin Lakes, NJ) 添加 cooked meat medium (Difco, Becton Dickinson and Co.)で 30°C、18 時間嫌気条件下で静置培養した。
boNT/A 遺伝子クラスタータイプを決定する Muliplex PCR 法:
菌株の培養液 1 ml から、DNeasy Blood and Tissue Kit (Qiagen Inc., Valencia, CA)を使用してゲノム DNA を抽出した。DNA は 100 μl の elution buffer で溶
出し、使用するまで 4°C で保管した。
boNT/A 遺伝子クラスタータイピングに用いたプライマーの塩基配列を
Table 5 に示す。2 つのプライマーセット、BoNT/A1 と BoNT/A2 を型別する 3 つのプライマーを含む “boNT/A1, A2” および、ha33 および p47 遺伝子を
検出する4 つのプライマーを含む “ha33, p47”を用いて型別を行った(Figure
2)。これらのプライマーは GenBank に登録されている boNT/A, ha33 および
p47 遺伝子の塩基配列を基に設計した。“boNT/A1, A2”プライマーセットは、
forward プライマーが BoNT/A1 あるいは BoNT/A2 遺伝子特異的に、reverse プライマーが両方の遺伝子にアニーリングするように設計した。
PCR は、鋳型 DNA 0.1~1 ng、各プライマー 0.25 μM、TaKaRa Ex Taq (TaKaRa Shuzo, Kyoto, Japan) 1.25 U, TaKaRa Ex Taq buffer 2.5 μl および各 dNTP 200 μl を含む溶液を滅菌水で全容量を 25 μl に調製した。DNA の増幅 反応は、PCR system 9700 (Applied Biosystems Inc., Foster City, CA)を用いて、 熱変性(94°C 1 分)、30 サイクルの増幅反応(熱変性 94°C 1 分、アニーリ
ング55°C 1 分、伸長反応 72°C 1 分)、最終伸長反応(72°C 5 分)で実施し
た。PCR 産物は 3% Nu-Sieve agarose gel (Camblex Bio Science, Rockland ME, U.S.A.) を使用して電気泳動し、ethidium bromide 染色後、写真撮影を行っ た。不活性な boNT/B 遺伝子は、boNT/A~boNT/G 遺伝子を検出する PCR 法 (83) を用いて検出した。
PFGE 法:
PFGE ゲルプラグの作成は Hielm らの方法に準じて行った (28)。各菌株を Brain Heart Infusion 培地 (Difco, Becton Dickinson and Co.)で 18 時間嫌気
培養後、培養液を 8,000 rpm, 5 分間、4°C で遠心分離し、沈渣を 4% (vol/vol)
ホルムアルデヒド加 Pett IV (PIV) buffer (10 mM Tris-HCl [pH 8.0] , 1 M NaCl)に懸濁し、氷上に 1 時間放置した。遠心洗浄後、同 buffer に懸濁し、 OD600(nm) = 1.3 に調製した。等量の 1.2% SeaKem Gold agarose (Camblex Bio Science) を混合し、0.75 mm サンプルプラグキャスターに流し込み固化させ
た。固化したゲルプラグを Lysis buffer (10 mM Tris-HCl [pH 8.0], 100 mM
EDTA [pH 8.0], 50 mM NaCl, 0.2% sodium deoxycholate, 0.5% sodium
laurylsarcosine, 0.5% Brij 58, 1 mg/ml lysozyme, 20 U/ml mutanolysin) に浸漬 し37°C、18 時間処理した。続いて、ProK buffer (50 mM Tris-HCl [pH 8.0], 100 mM EDTA [pH 8.0], 0.2% sodium deoxycholate, 0.5% sodium laurylsarcosine, 2 mg/ml proteinase K) 2 ml に入れ 50°C、48 時間処理した。ゲルプラグを泳動
時の大きさにカットし、4 mM Pefabloc SC 加 Tris-EDTA (TE) buffer 中に移し、
50°C、30 分処理を 2 回行い、proteinase K の不活化を行った。TE buffer で
平衡化した後、制限酵素用 buffer 200 μl 中に移し、20 U の制限酵素を用い
て各酵素の至適温度にて 18 時間処理した。制限酵素は、Sma I(GE Healthcare
UK Ltd., Little Chalfont, Buckinghamshire, UK)、Xho I (Roche Diagnostics, Mannheim, Germany)、Sac II (GE Healthcare UK Ltd.)および、Nru I(TaKaRa Shuzo)を使用した。処理後のゲルプラグは 1%アガロースゲル(BioRad Laboratories)、0.5×Tris Borate EDTA buffer を用いて、CHEF-DR®III system (BioRad Laboratories)にて電圧 6 V/cm, パルスタイム 0.5-40 秒、14°C で 20
差異が生じた場合に別パターンとした。FPQuest software ver. 4.5 (BioRad Laboratories)を用いて、類似性係数 Dice、デンドログラムタイプ UPGMA 法
にて、バンドパターンの解析および系統樹の作成を行った。
結果
boNT/A 遺伝子クラスター型別:
boNT/A 遺伝子クラスター型別を行うための Multiplex PCR 法を検討した。
A 型菌 27 株およびコントロール株 12 株の、プライマーセット”boNT/A1, A2”
および “ha33, p47”を用いた PCR の結果を Figure 3 に示し、A 型菌のクラス
ター型別結果を Table 4 にまとめた。11 株(CB111 株、89E00033-1 株、
89E00035-1 株、83E00080 株、2137-1-77 株、802-1 株、Denken 株、97A 株、 62A 株、33A 株および 36A 株)から boNT/A1 遺伝子(665 bp)および ha33
遺伝子(543 bp)が検出され、クラスタータイプ1に型別された。8 株(Chiba
H 株、Kyoto F 株、KZ1828 株、7I03 H 株、7I05 F 株、7I05 H 株、Y8036 株
および804-1H 株)から boNT/A2 遺伝子(440 bp)および p47 遺伝子(344 bp)
が検出され、クラスタータイプ 2 に型別された。8 株(Hiroshima 1 株、
Miyagi2006 株、Iwate2007 株、Renkon 株、CB121 株、Osaka99 株、89E00064-3
株および89E00086-1 株)から、boNT/A1 遺伝子、ha33 遺伝子および p47 遺
伝子が検出された。同時に不活性な boNT/B 遺伝子も検出されたため、クラ
スタータイプ 3 に型別された。”boNT/A1, A2” を用いた PCR では、コントロ
ール株12 株;B 型菌 3 株(Okra 株、111 株および Osaka05 株)、C 型菌 1 株
(CB-19 株)、D 型菌 1 株(1873 株)、E 型菌 1 株(Iwanai 株)、F 型菌 1 株 (Langeland 株)、E 型毒素産生性 C. butyricum 1 株(5262 株)、C. sporogenes 1 株(ATCC19404 株)、C. bifementas1株(ATCC638 株)、C. perfringens 1 株
(ATCC13124 株)および C. difficile 1 株(ATCC43593 株)から PCR 産物は 検出されなかった(Figure 3A)。“ha33, p47”を用いた PCR では、B 型菌 3 株 から ha33 遺伝子が検出され、F 型菌 1 株から p47 遺伝子が検出された(Figure 3B)。
PFGE 型別:
供試A 型菌 27 株の Sma I 切断 PFGE 型を Table 4 に示し、PFGE パターン
およびパターン間の類似性を基に作成したデンドログラムを Figure 4 に示
した。27 株は 22 の PFGE 型(S1~S22)に分類され、パターン間の類似性
は 29.6%~100%であった。1986 年から 1987 年に発生した乳児ボツリヌス
症分離株 7 株は 5 つの PFGE 型に分類され、パターン間の類似性は 78.6%~
100%であった。Chiba-H 株と KZ1828 株、および Kyoto-F 株と 7I03-H 株は
それぞれ同一の PFGE 型を示し、また両パターンの類似性は 96.3%であった。 7I05-F 株と 7I05-H 株は同一症例において、患者便および感染源と考えられ たハチミツから分離された株である。2 株は異なる PFGE 型を示したが、パ ターンの類似性は 96.6%であった。Y8036 株は、ブラジル産ハチミツ分離株 804-1H 株と高い類似性(92.9%)を示した。また、他の乳児ボツリヌス症 分離株6 株も 804-1H 株とは 81.5%~89.7%の類似性を示した。 1999 年から 2007 年に発生した乳児ボツリヌス症分離株 3 株は 2 つの PFGE 型に分類され、両パターンの類似性は 62.1%であった。Hiroshima1 株 および Miyagi2006 株は、わが国で 1999 年に発生した食中毒分離株 CB121
株と同一の PFGE 型を示した。これら 3 株は、Xho I、Sac II および Nru I 切
断PFGE パターンも同一であった(Figure 5)。Iwate2007 株は、米国で 1989
年に発生した乳児ボツリヌス症分離株89E00064-3 株と 92.9%の類似性を示
わが国で発生した乳児ボツリヌス症分離株において、1986 年から 1987 年 発生症例分離株 7 株と、1999 年から 2007 年発生症例分離株 3 株の PFGE パ ターン間の類似性は 42.9%~66.7%であった。また、わが国で発生した食中 毒事例分離株 4 株はそれぞれ異なる PFGE 型を示した。 考察 本章において、boNT/A 遺伝子クラスター型別を行う新たな Multiplex PCR 法を確立した。本法はリファレンス株(62A 株および Kyoto F 株)を、それ ぞれ既知のクラスタータイプ(クラスタータイプ 1 および 2) (23) に型別 することができた。検出限界は、”boNT/A1, A2” PCR が 5.5×10~2.8×102
cells/ml で、“ha33, p47” PCR が 2.1×102~2.1×103 cells/ml であった。PCR-RFLP
法による BoNT/A1, A2 型別と個別の PCR による ha33 遺伝子および p47 遺 伝 子 検 出 を 組 み 合 わ せ た boNT/A 遺 伝 子 ク ラ ス タ ー タ イ プ 型 別 法 が 、 Franciosa ら (23) から報告されているが、本研究で構築した Multiplex PCR 法は、より簡便、迅速なだけでなく、特異的および高感度であることから、 汎用性が高く、分子型別だけでなくボツリヌス症検査にも応用できる可能性 がある。 わが国で 1986 年から 1987 年に発生し、ハチミツ摂取歴のある A 型乳児 ボツリヌス症分離株 7 株はすべてクラスタータイプ 2 に分類され、南米産ハ チミツと関連する可能性が示唆された。ハチミツは主に中国、ヨーロッパ、 北アメリカおよび南アメリカで生産され、世界各国で輸出入される国際貿易 品である。南米では土壌中から A 型ボツリヌス菌が高率に分離されること (48)、および 1982 年から 2005 年までに南米で発生した乳児ボツリヌス症 370 症例中 369 症例が A 型菌の症例であると報告されている (41)。しかし、こ
れらの分離 A 型菌の毒素 subtype や遺伝子型などは不明である。 一方、1999 年から 2007 年に発生し、ハチミツ摂取歴のない A 型乳児ボツ リヌス症分離株はすべて boNT/A 遺伝子クラスタータイプ 3 に分類され、 1986 年から 1987 年に発生した本症分離株とは遺伝子型が著しく異なってい た。Hiroshima1 株および Miyagi2006 株は、食中毒分離株 CB121 株と同一の 遺伝子型を示した。わが国において乳児ボツリヌス症とボツリヌス食中毒と の関連が示唆された初めての事例である。しかし、これらの症例および事例 の発生場所、発生時期および感染源はそれぞれ異なっており、疫学的な関連 性は見出せない。また、わが国における環境中(土壌、河川、湖沼など)の ボツリヌス菌汚染調査の結果、E 型菌や C 型菌はそれぞれ北海道・東北地 方、中部地方を中心に分離されるが、A 型菌および B 型菌はほとんど分離 されていない (53, 87, 89)。これより、分離株の遺伝子型が同一であった乳 児ボツリヌス症例と食中毒事例には共通して何らかの輸入品を含む原材料 などが関与していた可能性が示唆される。Iwate2007 株は米国での乳児ボツ リヌス症分離株と相同性の高い PFGE パターンを示した。この 2 株間にも疫 学的な関連性は見出せないが、わが国と米国の乳児ボツリヌス症分離株間の 関連性が示されたことは興味深い。近年、海外ではハチミツの他に、粉ミル ク (11, 36)、ベビーフード (4, 59)、ハウスダスト (54) がそれぞれ乳児ボツ リヌス症の感染源として報告されている。わが国で発生した乳児ボツリヌス 症例が、食中毒事例や海外の乳児ボツリヌス症例と関連する可能性が示され たことは、ボツリヌス症発生の背景を考える上で有益なな情報になると考え られる。今後は、乳児が喫食する食品や環境物だけでなく様々な食品、原材 料や環境物について、ボツリヌス症のリスク検討が必要である。 A 型菌において、boNT/A 遺伝子クラスタータイプと分布地域との関連が 報告されている。米国で分離される A 型菌は、主にクラスタータイプ 1 ま
たは3 であり、ヨーロッパでは主にクラスタータイプ2であることが報告さ れている (23, 29)。わが国で分離された A 型菌は、クラスタータイプ1、2 または3 に分類され、米国やヨーロッパとは異なる傾向を示すことが明らか になった。また、クラスタータイプは菌株の由来、分離された年代やハチミ ツ摂取歴の有無などによって異なっていたため、本法は分離株の疫学的な差 異を反映していると考えられた。クラスタータイプ型別はPFGE 法と比較す ると型別能は低いが、得られた結果の相互比較が容易であるため、研究室間 や国際間における分子型比較法として優れている。よって、本章で構築した Multiplex PCR 法は、日本国内だけでなく国外における分離菌の疫学解析に 有用な手段になり得ると考えられる。
第2章 B 型乳児ボツリヌス症分離株の毒素遺伝子の解析
緒言
ボツリヌス B 型菌は、タンパク分解性や発育至適温度などの生化学的性
状やrRNA の塩基配列が著しく異なる第 I 群菌および第 II 群菌に分けられる
(Table 1)。現在、boNT/B 遺伝子塩基配列の分子系統解析から、BoNT/B は
5 つの subtype に分類されることが報告されている (29)。第 I 群 B 型菌産生
毒 素 が 3 種類(B1, B2 および B3)、第 II 群 B 型菌産生毒素が 1 種類
(Nonproteolytic)、B 型以外の毒素遺伝子を同時に保有する B 型(A[B], Ab,
AB, Bf など)産生毒素が1種類(Bivalent)に分類される (29)。第 I 群 B 型 菌産生毒素は、ほとんどの株が B1 もしくは B2 subtype に属し、現在 B3 subtype としては 1 株(CDC795 株)のみが報告されている。 B 型ボツリヌス食中毒からは第 I 群および第 II 群菌が分離されるが、B 型 乳児ボツリヌス症からは第 I 群菌のみが分離される (47)。わが国の B 型ボ ツリヌス食中毒は、1951 年から 2007 年までの間に、1969 年に宮崎県で発生 した西ドイツ産キャビアによる事例、1984 年に栃木県で発生した感染源不 明事例および、1998 年に東京都で発生したイタリア産瓶詰めグリーンオリ ーブによる事例の 3 事例が報告されている(Table 3)。乳児ボツリヌス症は、 1986 年から 2007 年までに報告されている 24 症例中、3 症例の B 型症例が 報告されている(Table 2)。1995 年 3 月に石川県においてわが国ではじめて の B 型乳児ボツリヌス症の発生が報告されて以来、10 年間発生の報告はな かったが、2005 年 10 月に大阪市内、2006 年に 9 月に大阪府下(高槻市内) で相次いで 2 症例が発生した。3 症例とも患者は典型的な症状を示し、予後 は良好であったが、ハチミツ摂取歴はなく、感染源は特定されていない (1,
32, 33, 37, 55, 88)。2005 年、2006 年の症例では、患者宅のベビーフード、飲 料、漢方薬、ハウスダストなどの検査が行われたが、どの検体からもボツリ ヌス菌およびボツリヌス毒素は検出されなかった。これまでに、1995 年の 症例から分離された B 型菌 111 株の産生毒素は、BoNT/B 産生標準菌株とし て広く用いられてきた Okra 株の産生毒素と、毒力、抗原性および生物活性 が異なることが示されている (31, 42, 44)。その後、それぞれの boNT/B 遺伝 子の解析が行われた結果、Okra 株産生毒素が BoNT/B1、111 株産生毒素が BoNT/B2 と定義され、両者の塩基配列およびアミノ酸配列の相同性は、そ れぞれ97.6%および 95.7%であることが明らかにされた (31, 74)。 また、従来 boNT/B 遺伝子は染色体上に存在すると考えられてきた (30, 57, 86) が、2007 年に Okra 株の全ゲノム塩基配列が決定され、本菌株の boNT/B 遺伝子が約 147 kbp のプラスミドに存在していることが明らかになった (73)。しかし、他の B 型菌の毒素遺伝子の存在位置については明らかにされ ておらず、毒素遺伝子の存在位置と BoNT/B subtype や菌株の由来との関連 については検討されていない。 本章では、わが国で発生した B 型乳児ボツリヌス症分離株の遺伝子解析 および分子疫学的解析を行った。2005 年および 2006 年に発生した乳児ボツ リヌス症患者便から分離された第I 群 B 型菌 Osaka05 株および Osaka06 株の boNT/B 遺伝子塩基配列全長を決定し、既知の boNT/B 遺伝子とともに分子系 統解析を実施した。わが国で発生した乳児ボツリヌス症分離株に様々な由来 の B 型菌を加えて PFGE による分子型別を行った。また、制限酵素処理を
行わない PFGE および boNT/B 遺伝子を検出する Southern blot hybridization
法により供試菌株の boNT/B 遺伝子存在位置を明らかにした。新たな分子型
材料と方法 供試菌株および DNA 抽出: わが国で発生したB型乳児ボツリヌス症分離株3 株、米国で発生した乳児 ボツリヌス症分離株 5 株、食中毒分離株 1 株、食品分離株 4 株および保存株 2 株の、計 15 株の I 群 B 型菌を供試した(Table 6)。他の Clostridium 属菌 11 株を PCR のコントロールとして供試した (Figure 10)。タンパク分解性を 含む生化学的性状は、API 20A kit(Biomerieux, France)を用いて解析した。
すべての菌株が B 型以外の不活性な boNT 遺伝子を保有しないことを
boNT/A~boNT/G 遺伝子を検出する PCR 法 (83) で確認した。各菌株は、0.3%
glucose および 0.2% soluble starch (Difco, Becton Dickinson and Co.) 添加 cooked meat medium (Difco, Becton Dickinson and Co.)で 30°C、18 時間嫌気条
件下で静置培養した。ゲノム DNA は菌株の培養液 1 ml から、DNeasy Blood
and Tissue Kit (Qiagen Inc.)を使用して抽出した。DNA は 100 μl の elution buffer で溶出し、使用するまで 4°C で保管した。
boNT/B 遺伝子および無毒成分遺伝子の塩基配列の決定:
BoNT/B 遺伝子および無毒成分遺伝子(ha70, ha17, ha33, botR および ntnh) は、Table 7 に示した増幅用プライマーを用いた PCR 法によって増幅した。 PCR の反応液は、鋳型 DNA 10 ng、各プライマー 0.5 μl、TaKaRa LA Taq (TaKaRa Shuzo) 2.5 U、LA Taq buffer 5 μl、MgCl2 2.5 mM および各 dNTP 400 μM を含む溶液を滅菌水で全容量を 50 μl に調製した。DNA の増幅反応は、 PCR system 9700 (Applied Biosystems Inc.)を用いて、熱変性(94°C 1 分)、25
サイクルの増幅反応(熱変性94°C 1 分、アニーリング 50°C 1 分、伸長反応
Table 7 に示したシーケンス用プライマーを使用し、ABI PRISM BigDye cycle sequencing kit (Applied BioSystems Inc.)を用いたダイレクトシーケンス法に よって決定した。
BoNT/B C 末端領域遺伝子の塩基配列の決定:
BoNT/B の C 末端領域遺伝子 400 bp は、プライマー;Forward
[5’-GAAAGTCAAATTCTCAATC-3’; position in the coding region, 3445 to 3463 base]および、reverse [5’-CAAAATTTAGCTACATCCT-3’; position, 3961 to 3943 base] を用いて増幅し、PCR 産物からダイレクトシーケンス法によって塩基
配列を決定した。PCR の反応液は、鋳型 DNA 1 ng、各プライマー 0.5 μl、
TaKaRa LA Taq (TaKaRa Shuzo) 2.5 U、LA Taq buffer 5 μl、MgCl2 2.5 mM およ
び各dNTP 200 μM を含む溶液を滅菌水で全容量を 50 μl に調製した。DNA
の増幅反応は、PCR system 9700 (Applied Biosystems Inc.)を用いて、熱変性
(94°C 1 分)、25 サイクルの増幅反応(熱変性 94°C 1 分、アニーリング 50°C
1 分、伸長反応 65°C 2 分)、最終伸長反応(65°C 5 分)で実施した。
塩基配列の accession numbers:
本研究で決定した塩基配列は、DDBJ/EMBL/GenBank database に登録し、
accession number は AB302852-AB302865 である(Table 6)。
分子系統解析:
塩基配列は、Clustal X version 1.83(パラメーターClustal W 1.6)でアライ ンメントした。系統樹は、1,000 回のブートストラップ反復を行い、遺伝的
距離はKimura two-parameter 法にて計算し、近接結合法を用いて形成した
PFGE:
PFGE ゲルプラグの作成は Hielm らの方法に準じ (28)、前章と同様に行っ た。20 U の制限酵素 Sma I(GE Healthcare UK Ltd.)を用いて、200 μl の制
限酵素用buffer 中で 25°C で 18 時間処理したゲルプラグおよび、制限酵素
処理を行わないゲルプラグを、1%アガロースゲル(BioRad Laboratories)、
0.5×TBE buffer を用いて、CHEF-DR®III system (BioRad Laboratories)にて電
圧6V/cm, パルスタイム 0.5-40 秒、14°C で 20 時間泳動した。Ethidium bromide
染色後、写真撮影した。FingerprintingTM II ソフトウエア (BioRad Laboratories)
を用いて、類似性係数 Dice、デンドログラムタイプ UPGMA 法にて、バン
ドパターンの解析および系統樹の作成を行った。DNA フラグメントの分子
量(kbp)はサイズマーカー λ Ladder (Bio Rad Laboratories)の分子量から推 定した。
Southern blot hybridization:
PFGE 後のアガロースゲルから DNA フラグメントを Hybond N+ nylon
membrane (GE healthcare UK. Ltd.)に、20×Saline-sodium citrate (SSC) buffer
を用いた18 時間のキャピラリートランスファー法によって転写した。DNA
は1200×10μJ/cm2、3 回の UV 照射(CL Ultra violet Cross Linker, UVP)に てメンブレンに定着させた。boNT/B 遺伝子および 16S rRNA 遺伝子を検出す
るDIG ラベル DNA プローブは、Okra 株から抽出した DNA を鋳型とし、
boNT/B 遺伝子は Szabo ら (78)、16S rRNA 遺伝子は Marshall ら (49) のプラ
イマーを用いて、PCR DIG Probe Synthesis Kit (Roche Diagnostics) を使用し
て作製した。メンブレンは、DIG-Easy Hyb (Roche Diagnostics)中で 37°C で 2
中42°C で 18 時間処理し、ハイブリダイゼーションを行った。続いて、0.1 % SDS を含む 1×SSC buffer で室温、15 分間、3 回洗浄し、さらに 0.1 % SDS
を含む0.1×SSC buffer で 60°C(boNT/B 遺伝子プローブ)もしくは 65°C(16S
rRNA 遺伝子プローブ)、15 分間、2 回洗浄した。ハイブリダイゼーション
シグナルは、DIG Luminescent Detection Kit (Roche Diagnostics)を用いて検出 した。
BoNT/B subtype を識別する Multiplex PCR 法:
boNT/B1、boNT/B2 および boNT/Osaka05 遺伝子を検出する Multiplex PCR
は、Table 8 に示した 4 種のプライマーを混合して行った。これらのプライ
マーはGenBank に登録されている boNT 遺伝子塩基配列および本研究で決定
した boNT/B 遺伝子塩基配列に基づいて、Forward プライマーは boNT/B1、
boNT/B2 および boNT/Osaka05 遺伝子に共通で、reverse プライマーは BoNT/B
C 末端領域の塩基配列の多様性を利用し、それぞれ特異的な遺伝子にアニー リングするように設計した。PCR の反応液は、鋳型 DNA 0.3~1 ng、各プラ イマー 0.25 μM、TaKaRa Ex Taq (TaKaRa Shuzo) 1.25 U, TaKaRa Ex Taq buffer 2.5 μl および各 dNTP 200 μl を含む溶液を滅菌水で全容量を 25 μl に調製した。 DNA の増幅反応は、PCR system 9700 (Applied Biosystems Inc.)を用いて、熱 変性(94°C 1 分)、25~35 サイクルの増幅反応(熱変性 94°C 1 分、アニー
リング55°C 1 分、伸長反応 72°C 1 分)、最終伸長反応(72°C 5 分)で実施
した。PCR 産物は 3% Nu-Sieve agarose gels (Camblex Bio Science) を使用し て電気泳動し、ethidium bromide 染色後、写真撮影を行った。
Osaka05 株および Osaka06 株 boNT/B 遺伝子塩基配列の分子系統解析:
Osaka05 株および Osaka06 株の boNT/B 遺伝子塩基配列全長(3,876 bp)を
決定した。両塩基配列は 98.8%の相同性を有していた。さらに現在 GenBank に登録されている 24 株の boNT/B 遺伝子塩基配列(Table 6)との比較を行 った。Osaka06 株は B2 subtype に分類されている CDC6291 株の配列と 100% 一致していた。Osaka05 株はどの配列とも一致しなかった。Osaka05 株、 Osaka06 株および前述の 24 株の boNT/B 遺伝子塩基配列全長を基に分子系統 解析を行い、構築した系統樹を Figure 6A に示した。26 株は、5 つの subtype クラスター(B1, B2, B3, Bivalent および Nonproteolytic)に分類されたが、 Osaka05 株はどのクラスターにも属さなかった。Osaka06 株は B2 subtype に
属し、他のB2 subtype 株と 99.3%~100%の相同性を示した。新たなクラス
ターに分類された Osaka05 株は、B2 subtype 株と 98.8%~99.1%、B3 subtype
株 と 98.9 % 、 B1 subtype 株 と 97.7 % ~ 98.0 % 、 Bivalent 株 と 97.4 % 、 Nonproteolytic 株と 95.7%~96.2%の相同性を示した。
Osaka05 株、Okra 株(B1 subtype)および 111 株(B2 subtype)の boNT/B
および無毒成分遺伝子塩基配列およびアミノ酸配列の比較:
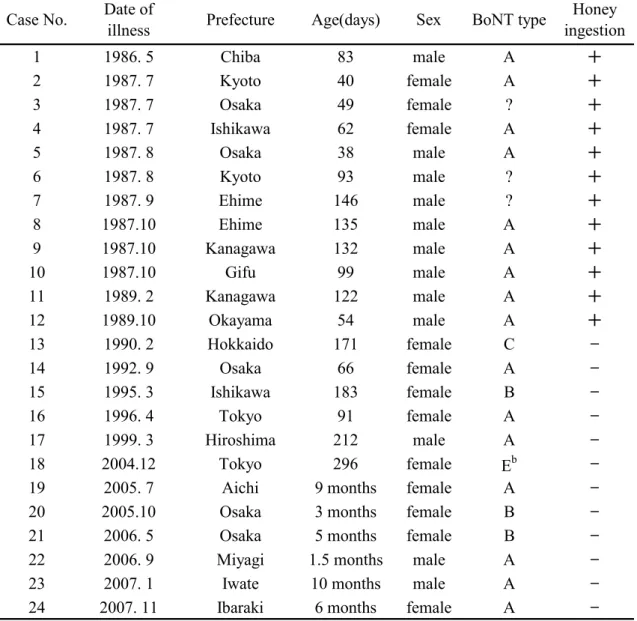
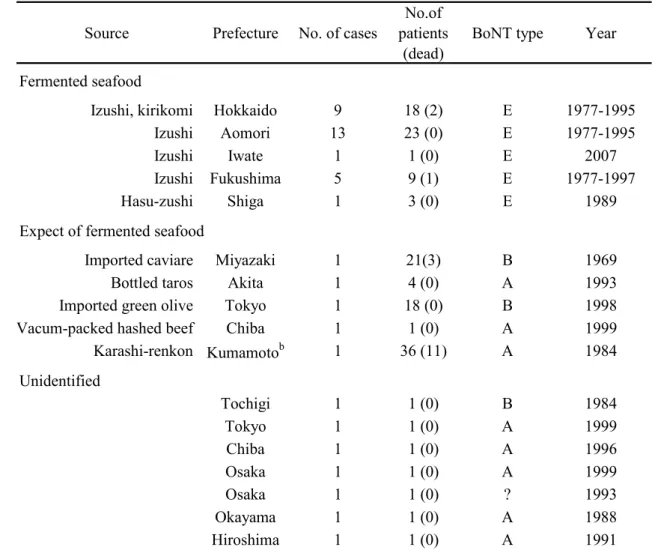
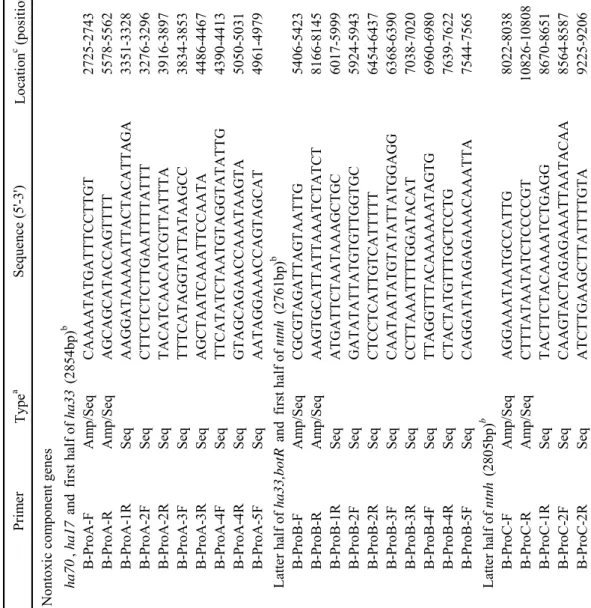
Osaka05 株、Okra 株および 111 株の boNT/B および無毒成分遺伝子(ha70; 1,881 bp、ha17; 441 bp、ha33; 879 bp、botR; 537 bp および ntnh; 3,594 bp)の
塩基配列およびアミノ酸配列の相同性をTable 9 に示した。Osaka05 株およ び111 株の無毒成分遺伝子塩基配列を決定し、Okra 株は GenBank に登録さ れている配列(Accession No.AB232927)を用いた。boNT/B 遺伝子塩基配列 およびアミノ酸配列の相同性は、Osaka05 株と Okra 株ではそれぞれ 97.9% および 96.2%、Osaka05 株と 111 株では 99.0%および 98.5%、また Okra 株 と 111 株では 97.6%および 95.7%であった。boNT/B 遺伝子クラスターの構
成(boNT/B 遺伝子の上流に ha70, ha17, ha33, botR および ntnh)は 3 株間で 共通していた。ha33 以外の無毒成分の塩基配列およびアミノ酸配列の相同 性はOsaka05 株と Okra 株ではそれぞれ 97.9%~99.6%および 96.7%~99.0%、 Osaka05 株と 111 株では 98.6%~99.6%および 98.4%から 100%、また Okra 株と 111 株では 97.8%~98.9%および 96.4%~98.1%であった。一方 ha33 で は 、3 株間の遺伝子塩基配列およびアミノ配列の相同性が、それぞれ 92.3%~95.2%および 83.6%~90.1%であり、他の無毒成分遺伝子および boNT/B 遺伝子と比較して著しく低かった。
Okra 株、111 株および Osaka05 株の BoNT/B アミノ酸配列を比較した結果、 61 残基の変異が検出された(Figure 7)。変異アミノ酸は、軽鎖(L: 441 aa) に2 残基、重鎖(H: 850 aa)に 59 残基が存在し、重鎖の変異をドメインご とに分けると、重鎖 N 末端領域(HN: 420 aa)に 22 残基、重鎖 C 末端領域 (HC: 430 aa)に 37 残基が存在し、HCの変異をサブドメインごとに分ける と、 HCNサブドメイン(167 aa)に 12 残基、HCCサブドメイン(263 aa)に 25 残基が存在していた。
Osaka05 株と Okra 株の BoNT/B 各ドメインおよびサブドメインのアミノ
酸配列の相同性は、99.5%(L)、94.5%(H)、95.2%(HN)、92.8%(HCN)、 94.7%(HCC)でり、Osaka05 株と 111 株は、100.0%(L)、97.6%(H)、98.6% (HN)、100.0%(HCN)、94.3%(HCC)であり、また Okra 株と 111 株は、99.5% (L)、93.6%(H)、95.5%(HN)、92.8%(HCN)、91.3%(HCC)であった。 BoNT/B C 末端領域の塩基配列に基づく分子系統解析: BoNT/B subtype を効率よく区別する方法を検討するため、供試菌株 15 株 および第 I 群 B 型菌のリファレンス配列 13 株(Table 6)、計 28 株について、 boNT/B 遺伝子塩基配列間の多様性が最も高いと考えられた BoNT/B HCCサ
ブドメイン塩基配列の一部(400 bp)を比較し、分子系統解析を行った(Figure 6B.)。28 株は、boNT/B 塩基配列全長からの分子系統解析の結果と同様、3 つの第I 群 subtype クラスター(B1、B2 および B3)に分類されたが、Osaka05 株はどのクラスターにも属さなかった。B1 subtype には米国で発生した乳児 ボツリヌス症分離株 5 株、豚肉分離株 2 株、保存株 3 株、リファレンス配列 4 株が属し、B2 subtype にはわが国で発生した乳児ボツリヌス症分離株 2 株、 ショウガ分離株、ハチミツ分離株、リファレンス配列8 株が属していた。リ ファレンス配列CDC795 株および Osaka05 株は B1 もしくは B2 subtype クラ スターには属さず、また両者は同じクラスターに分類されなかった。
PFGE および Southern blot hybridization:
供試菌株15 株の Sma I 切断 PFGE パターンおよびその類似性に基づき構 築したデンドログラムを Figure 8 に示した。すべての菌株は異なるパターン を示し、パターン間の類似性は 25.8%~94.1%であった。Osaka05 株は、他 の 14 株と著しく異なるパターンを示し、B1 subtype 株 10 株との類似性は 28.6%~45.7%、B2 subtype 株 4 株との類似性は 25.8%~54.6%であった。 B1 subtype 株と B2 subtype 株との類似性は、29.4%~60.0%であった。同一 subtype に属する菌株間の類似性は、B1 subtype 株間は 41.2%~94.1%、B2 subtype 株間は 35.7%~53.9%であった。B1 subtype 10 株は 89E00061-2 株を
除いて、類似性 80%以上の 3 つのクラスター(Okra 株および 407 株、326
株、9B 株および 67B 株、89E00067-4 株、89E00123-1 株、90E00001-3 株お
よび3129-2-77 株)に分類された。
Sma I 切断 PFGE から Southern blot hybridization 法を用いて検出した
boNT/B 遺伝子が存在するフラグメントを arrowhead で示した(Figure 8)。
3129-2-77 株、326 株、9B 株および 67B 株からは約 110 kbp、Okra 株および 407 株からは約 150 kbp、7H215S 株からは約 175 kbp、Osaka06 株からは約 175 kbp、89E00061-2 株からは約 260 kbp、111 株からは約 275 kbp、Osaka05
株からは約 280 kbp のフラグメントが検出された。
制限酵素処理を行わない PFGE から、111 株、Osaka05 株、89E00061-2 株、
Okra 株および 407 株において、染色体 DNA(>970 kbp)の他に、約 150 kbp
~280 kbp のフラグメントが各 1~3 本検出された(Figure 9A)。boNT/B 遺伝
子の存在位置を Southern blot hybridization で確認した結果、Okra 株および
407 株では約 150 kbp、89E00061-2 株では約 260 kbp、111 株では約 275 kbp、 Osaka05 株では約 280 kbp のフラグメント上に検出された(Figure 9B)。一
方、これら 5 株の 16S rRNA 遺伝子はすべて染色体の位置に検出された
(Figure 9C)。その他の供試菌株 10 株においては、制限酵素を行わない PFGE
から染色体 DNA しか検出されず、Southern blot hybridization の結果、boNT/B
遺伝子および 16S rRNA 遺伝子は染色体 DNA 上に存在していた。
BoNT/B subtype を識別する Multiplex PCR 法:
boNT/B1、boNT/B2 および boNT/Osaka05 遺伝子を検出する Multiplex PCR
を検討した。供試菌株 15 株および他の Clostridium 属菌 11 株を用いた結果
を Figure 10 に示した。10 株(Okra 株、89E00061-2 株、89E00067-4 株、
89E00123-1 株、90E00001-3 株、3129-2-77 株、326 株、407 株、9B 株および 67B 株)からは、585 bp の boNT/B1 遺伝子断片が増幅され、4 株(111 株、 Osaka06 株、Ginger 株および 7H215S 株)からは、370 bp の boNT/B2 遺伝子 断片が増幅され、Osaka05 株からは 317 bp の boNT/Osaka05 遺伝子断片が、 そ れ ぞ れ 特 異 的 に 増 幅 さ れ た 。 本 PCR の 検 出 限 界 は 、 9.5×10~ 2.5×102 cells/ml であった。B 型菌以外の陰性コントロール菌株 11 株;A 型菌 2 株(62A
株および Kyoto-F 株)、C 型菌 1 株(CB-19 株)、D 型菌 1 株(1873 株)、E
型菌1 株(Iwanai 株)、F 型菌1株(Langeland 株)、BoNT/E 産生 C. butyricum
1株(5262 株)、C. sporogenes1株(ATCC19404 株)、C. bifementas1株
(ATCC638 株)、C. perfringenes1株(ATCC13124 株)および C. difficile1株 (ATCC43593 株);からは、いずれの遺伝子断片も増幅されなかった。
考察
わが国で 2005 年および 2006 年に連続して発生した B 型乳児ボツリヌス
症分離株 Osaka05 株および Osaka06 株について、boNT/B 遺伝子塩基配列全
長を決定し、GenBank に登録されている boNT/B 遺伝子塩基配列と比較した。 BoNT subtype は、かつては同一毒素型内でアミノ酸配列が 2.6%以上異なる 場合を指していた (74) が、現在では boNT 遺伝子塩基配列全長に基づく分 子系統解析によって別クラスターに分類された配列をsubtype とすることが 一般的になっている (13, 29)。本章で行った boNT/B 遺伝子の比較から、 Osaka05 株は特異的な boNT/B 遺伝子塩基配列を有すること、および既存の 5 つの BoNT/B subtype クラスターには属さないことが明らかになった。こ
れよりOsaka05 株は新たな BoNT/B subtype と定義される可能性が示唆され
た。一方、Osaka06 株の boNT/B 遺伝子塩基配列は B2 subtype 株の CDC6291
株 (29) の配列と完全に一致し、また分子系統解析からもB2 subtype に分類 された(Figure 6A)。 Subtype 間の BoNT/B アミノ酸変異は重鎖、特に重鎖 C 末端、中でも HCC サブドメインに集中していた。このことを利用し、BoNT/B C 末端領域(HCC サブドメインの一部)の塩基配列に基づく分子系統解析による subtyping を 試みた結果、boNT/B 塩基配列全長を用いて解析した場合と同様の結果が得
られた(Figure 6B)。また、同領域塩基配列の多様性から boNT/B1、B2 およ び Osaka05 遺伝子をそれぞれ特異的に検出する Multiplex PCR 法を検討した (Figure 10)。すべての供試菌株において、本 PCR の結果と boNT/B 遺伝子 塩基配列の分子系統解析から決定したBoNT/B subtype は一致していた。従 って、この方法は簡便な BoNT/B subtyping 法として適用が可能であると考 えられる。
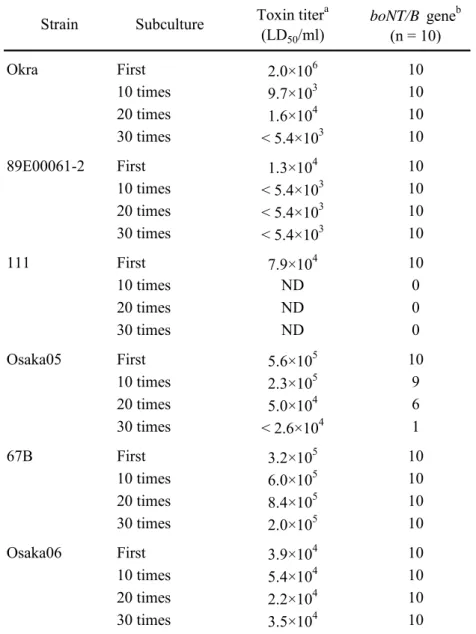
Sma I 切断 PFGE の結果、B1 subtype 株、B2 subtype 株および Osaka05 株
間の遺伝学的多様性が明らかになった。B2 subtype 株間では、B1 subtype 株 間よりも遺伝学的に多様性があることが分かった。わが国で分離された B 型菌7 株(乳児ボツリヌス症分離株 3 株および食品分離株 4 株)は、互いに パターンが明らかに異なっていた。一方、米国で発生した B 型乳児ボツリ ヌス症分離株間は比較的高い相同性を有し、わが国の症例とは背景が異なる と考えられた。 従来、boNT 遺伝子およびその上流に位置する無毒成分遺伝子の存在位置 は、毒素型によって異なると考えられていた。boNT/A、B、E および F 遺伝 子は染色体上に、一方 boNT/C および D 遺伝子はバクテリオファージ上に、 boNT/G 遺伝子はプラスミド上に存在することが報告されてきた (10, 57, 86, 91)。しかし最近、同一毒素型内で boNT 遺伝子の存在位置が異なるケース が報告されている。B 型菌だけでなく、A 型菌においても、2007 年に Loch Maree 株(BoNT/A3)、657Ba 株(BoNT/A4)の boNT/A 遺伝子がそれぞれ約 270 kbp のプラスミドに存在することが報告されている (49, 73)。本章では、
それぞれ由来も subtype も異なる B 型菌 5 株の boNT 遺伝子が染色体 DNA
ではない DNA フラグメントに存在することを示した(Figure 9)。このフラ
グメントには rRNA 遺伝子が存在しないため、プラスミドである可能性が高
約260 kbp(89E00061-2 株)、約 275 kbp(111 株)および約 280 kbp(Osaka05 株)であった。この結果は、boNT/B 遺伝子の存在位置と BoNT/B subtype (boNT/B 遺伝子塩基配列の多様性)とは関連しないことを示している。 boNT/B 遺伝子の存在位置が異なることの意義を遺伝学的および生物学的に 解明することで、ボツリヌス菌の毒素産生機構、遺伝子の獲得および進化の 過程についての新たな知見が得られると思われる。 BoNT は無毒成分との複合体毒素として産生されるが、生体内における無 毒成分の役割は、BoNT の安定性を高めること、BoNT を胃酸やプロテアー ゼから守ること、BoNT の小腸上皮からの吸収を促進することなどであると 考えられている (57)。これまでの報告から、boNT/B 遺伝子の上流には、ヘ
マ グ ル チ ニ ン 遺 伝 子 ( ha70, ha33, ha17 )、 転 写 調 節 遺 伝 子 ( botR )、 nontoxic-nonhemagglutinin 遺伝子(ntnh)が存在することが分かっている (46, 57, 73)。BoNT/B subtype 間でアミノ酸配列を比較すると、他の無毒成分およ
びBoNT/B に比べ HA33 の相同性が著しく低かった(Table 9)。HA33 は毒素
複合体が小腸上皮細胞および赤血球に付着する際の接着因子として働くこ と が 報 告 さ れ て い る (24, 25, 50) こ と か ら 、 乳 児 ボ ツ リ ヌ ス 症 に お い て HA33 のアミノ酸変異が本症の発生にどのような影響を与えているのかを 明らかにすることは興味深いと思われる。
BoNT HCCサブドメインにおいて、BoNT/Osaka05 と BoNT/Okra とのアミ
ノ酸置換は 15 残基であった。BoNT/Osaka05 と BoNT/111 では 14 残基、
BoNT/Okra と BoNT/111 は 23 残基であった。精製毒素を用いた研究から、 BoNT/111 は BoNT/Okra よりも毒力が低く、それは BoNT/111 の神経細胞受
容体(ガングリオシド GT1b およびシナプトタグミン 2/GTb1 複合体)への
結合が BoNT/Okra よりも弱いことに起因し、変異アミノ酸の解析から、受
ことが明らかにされてきた (31, 42, 44)。他の報告では、HCCサブドメイン
中の8 残基(4残基はガングリオシド GT1b への、4 残基はシナプトタグミ
ン 2/GTb1 複合体への結合に関与)が、BoNT/B の受容体結合に必須である
ことが明らかにされている (63, 77)。これら HCCサブドメイン中の 10 残基
において、BoNT/Osaka05 は BoNT/Okra と一致していた(Figure 7)。今後 BoNT/Osaka05 の構造と活性を詳細に検討することで、BoNT/B の受容体結 合機構に関し、新しい情報が得られると期待される。
本章では 2 つの新たな BoNT/B subtyping 法、boNT/B 遺伝子塩基配列の一
部を用いて分子系統解析を行う方法およびMultiplex PCR 法を構築した。前
者は新たな BoNT/B subtype を検出するのに優れ、後者はより簡便性が高い。
BoNT/B subtype の世界的な分布は、B1 subtype が主に米国で分離され、B2 subtype が主にヨーロッパで分離されることが報告されており、B2 subtype 株は韓国の土壌からも分離されている (29)。わが国では第 I 群 B 型菌は環
境中から分離されていない (53, 87, 89) が、本章の結果から、乳児ボツリヌ
ス症からは B2 subtype および Osaka05 株、食品からは B1 subtype および B2
subtype が分離されたことが分かった。今後、輸入食品なども含めたボツリ ヌス検査および分離菌の分子型別を継続して行うことで、ボツリヌス症の感 染源解明や、詳細なリスク分析が可能になると期待される。
第 3 章 Osaka05 株が産生する B 型毒素の抗原性の解析 緒言 一般的に、毒素遺伝子の変異に伴うアミノ酸配列の変異は、本来毒素が 有している抗原性の変化を引き起こす可能性が高いと考えられる。毒素の抗 原性の変化は、現在使用あるいは検討されているボツリヌス症診断法、予防 法および治療法の感度や有効性に影響を与える可能性がある。ボツリヌス症 の検査においては、動物を用いたバイオアッセイに替わる、免疫学的手法を 用いた in vitro 毒素検出法(Enzume-Linked Immunosorbent Assay [ELISA]法、 ELISA-Enzume-Linked Coagulation Assay [ELCA]法、Immuno PCR 法、イムノ クロマト法など)が開発されている (67, 68) 他、従来の予防ワクチン(精 製毒素のトキソイド)の抗原を変更する利点や、治療剤としてのウマ抗血清 をヒト型モノクローナル抗体に代替しようとする方向性が論じられている (9, 71, 72, 90)。そのため、同一毒素 subtype 間における抗原性の差異につい ての詳細な知見を得ることが必要である。またBoNT subtype の分類は、毒 素遺伝子の塩基配列や毒素タンパクの一次構造の変異のみならず、抗原性の 差異も考慮する必要があると考えられる。これまで B 型毒素では、BoNT/B2 はBoNT/B1 と異なる抗原性を有することが報告されている (31, 44)。また、 その抗原決定基はBoNT/B subtype 間のアミノ酸置換が最も多い HCドメイン 上に存在することが明らかにされている。 本章では、第2章で boNT/B 遺伝子塩基配列の分子系統解析から既存の BoNT/B subtype には属さない新たな変異株であると考えられた乳児ボツリ
ヌス症分離株 Osaka05 株が産生する BoNT/B の抗原性を、既存の BoNT/B1
株(B2 subtype)の、リコンビナント BoNT/HCドメイン(rHC)をそれぞれ
調製し、これらの特異抗体を用いた Sandwich ELISA 法によって、各抗体の
rHC/Osaka05、rHC/Okra、rHC/111 および Osaka05 株、Okra 株、111 株の PYG 培養上清への反応性を比較し、Osaka05 株の抗原特異性を明らかにした。 材料と方法 リコンビナント BoNT/HCドメイン(rHC)の作製: ヒスチジンタグ融合 rHC(rHC/Osaka05、rHC/Okra および rHC/111)の発現 は、既報 (31) に準じて行った。HC/Osaka05 をコードする DNA フラグメン トをPCR 増幅するためのプライマーを新たに設計した。使用したプライマ ーの配列を以下に示す。Forward, 5’-CACGGATCCAAAAAATATAATAGCGAAATTTTAAA-3’; reverse, 5’-ATTAAGCTTTTATTCAATCCATCCTTCATCTTT-3’。発現した各 rHCは、
Ni-NTA agarose(Qiagen Inc.)を使用してアフィニティー精製した後、 rHC/Osaka05 は pH 7.0、rHC/Okra および rHC/111 は pH 6.5 で CM-Sephadex C-25
(GE Healthcare UK Ltd.)カラムに結合後、プロトコールに従ってイオン交
換クロマトグラフィー精製を行った。
抗 rHCアフィニティーポリクローナル抗体の作製:
各 rHCに対するウサギアフィニティーポリクローナル抗体を調製した。日
本白色種ウサギ(2.5~3.0 kg、オス)に、精製した 50 μg の各 rHCにFreund’s
complete adjuvant(Difco, Becton Dickinson and Co.)を混合して皮下接種し、
初回免疫とした。初回免疫後、2 週間隔で 50 μg の各 rHCにFreund’s incomplete
疫を行った。さらに 2 週間後、50 μg の各 rHCを接種し、2 週間後に全採血
を行い、血清を採取した。硫酸アンモニウム沈殿にてIgG 分画を回収し、
各rHCをカップリングした Hi-Trap NHS-activated HP column(GE Healthcare UK Ltd.)を用いてアフィニティー精製を行った。抗体のビオチン化には EZ-Link Sulfo-NHS-LC- Biotinylation Kit(Pierce, Rockford, U.S.A.)を使用し た。
供試菌株、培養条件および毒力測定:
第I 群 B 型菌 15 株(Table 6)を供試した。各菌株を PYG 培地 [2% Proteose
peptone(Oxoid), 0.5% Yeast extract (Mikuni peptone, Japan), 0.5% glucose, 0.025% Sodium thioglycolate(Kishida chem, Japan), pH 7.0]で 3 日間嫌気培養
した培養液を 10,000×g、5 分間遠心分離し、得られた上清を Type IX-S Trypsin(Sigma-Aldrich, USA)で 37℃、30 分処理した。毒力測定はマウス 静脈内注射法で行った (43)。適当な濃度に希釈した毒素 0.1 ml を静脈内に 投与し死亡するまでの時間を測定し、この際に毒力(腹腔内LD50)の対数 と致死時間の対数間で直線的な相関が成立することから、この関係をもとに 作製した標準曲線から毒素量(腹腔内LD50/ml)を算出した。また、上清を
口径が0.45μm の Cellulose acetate filter(Advantec, Japan)に通し、適宜 0.2% gelatin および 5 mM EDTA 添加 0.05 M phosphate buffer (pH 7.0)で希釈し
たものをSandwich ELISA 法の抗原として用いた (43)。
Sandwith ELISA 法:
96 穴マイクロタイタープレート(IWAKI, Japan)に、0.3 M NaCl 添加 10 mM phosphate buffer (pH 7.0)(ELISA 用バッファー)で希釈した 100 μl の非
2 時間静置した。0.05% Tween20 加 ELISA 用バッファー(洗浄用バッファー) で3 回洗浄後、150 μl の Block Ace(Dainippon Sumitomo Pharma Co. Ltd., Japan)を添加し、4°C、18 時間のブロッキングを行った。洗浄用バッファ ーで1 回洗浄後、100 μl の抗原溶液を添加し、37°C、1 時間インキュベーシ ョンした。抗原溶液として、10~5,000 ng/ml の各 rHCおよび供試菌株培養 上清を使用した。洗浄用バッファーで5 回洗浄後、100 μl のビオチン標識抗 体(1 μg/ml)を添加し、37°C、1 時間インキュベーションした。洗浄用バッ ファーで5 回洗浄後、100 μl の streptavidin-conjugated HRP(Pierce)(0.2 μg/ml) を添加し、37°C、1 時間インキュベーションした。洗浄用バッファーで 5 回 洗浄後、ABTS[2,2-azinobis (3-ethylbenzthiazoline sulfonic acid)](Roche Diagnostics)を添加し、37°C、1 時間インキュベーションし、反応を停止さ せた後、マイクロプレートリーダーLabsystem Multiscan MS (Labsystem, Finland) で 415 nm の吸光度を測定した。統計的な差は Student’s t test により 確認した。
結果
rHC/Osaka05、rHC/Okra および rHC/111 の抗原性の比較:
精製 rHC/Osaka05、rHC/Okra および rHC/111 の SDS-PAGE 像を Figure 11
に示した。それらの各 rHCに対する特異抗体への反応性を Sandwich ELISA
法によって測定した(Figure 12)。抗 rHC/Osaka05 抗体を用いた Sandwich
ELISA の結果、rHC/Okra および rHC/111 の反応性は rHC/Osaka05 と比較して
有意に低く、また rHC/Okra は rHC/111 よりも低い反応性を示した(Figure
12A)。抗 rHC/Okra 抗体を用いた Sandwich ELISA では、rHC/Okra と比較し
抗rHC/111 抗体を用いた Sandwich ELISA では、rHC/Osaka05 および rHC/Okra
の反応性は rHC/111 と比較して有意に低く、rHC/Okra は rHC/Osaka05 よりも
低い反応性を示した(Figure 12C)。
Osaka05 株、Okra 株および 111 株培養上清の抗原性の比較:
Osaka05 株、Okra 株および 111 株の PYG 培地培養上清を抗原として、各
抗 rHC特異抗体への反応性を Sandwich ELISA 法によって測定した(Figure
13)。抗 rHC/Osaka05 抗体を用いた Sandwich ELISA の結果、Okra 株および
111 株培養上清の反応性は Osaka05 株培養上清と比較して有意に低く、また Okra 株培養上清は 111 株培養上清よりも低い反応性を示した(Figure 13A)。
抗 rHC/Okra 抗体を用いた Sandwich ELISA では、Okra 株培養上清と比較し
て Osaka05 株および 111 株培養上清は顕著に低い反応性を示した(Figure
13B)。抗 rHC/111 抗体を用いた Sandwich ELISA では、Osaka05 株および Okra
株培養上清の反応性は 111 株培養上清と比較して有意に低く、Okra 株培養
上清はOsaka05 株培養上清よりも低い反応性を示した(Figure 13C)。
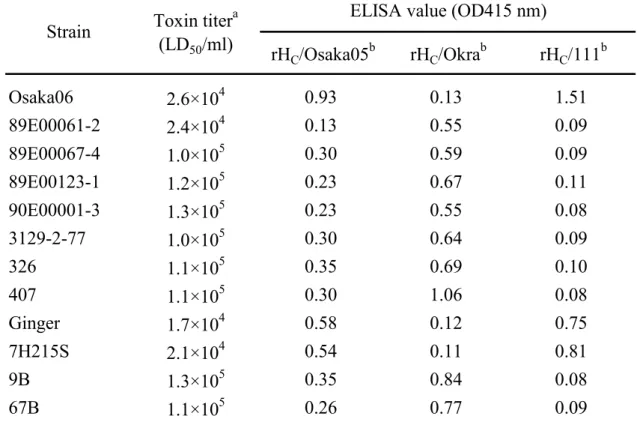
第 I 群 B 型菌培養上清の抗原性の比較:
第I 群 B 型菌 12 株の PYG 培地培養上清を抗原として、各抗 rHC特異抗体
への反応性を、前項と同様Sandwich ELISA 法によって測定した(Table 10)。
89E00061-2 株、89E00067-4 株、89E00123-1 株、90E00001-3 株、3129-2-77
株、326 株、407 株、9B 株および 67B 株の培養上清は、抗 rHC/Okra 抗体へ
の反応性が最も高く、Osaka06 株、Ginger 株および 7H215S 株の培養上清は、
抗rHC/111 抗体への反応性が最も高かった。



![Table 9. Percentage of identities of nucleotide and amino acid sequences among strains Osaka05, Okra (B1) and 111 (B2) in the BoNT/B and nontoxic components NTNH 3594 / 1197aBoNT/B3876 / 1291aHA70 1881 / 626aHA17441 / 146aHA33885[879] / 294[292]aBotR537 /1](https://thumb-ap.123doks.com/thumbv2/123deta/8512943.1805628/62.892.173.340.243.1046/table-percentage-identities-nucleotide-sequences-strains-nontoxic-components.webp)