報 文 1.緒 言 小麦粉を主原料としたパンや饅頭をはじめとする発酵食 品は,世界各地で多種類が存在し,各国独自の食文化を築 いている.伝統的な製法で作られた小麦粉発酵食品は,酵 母のみならず,自然界の様々な微生物の発酵を利用した風 味豊かな食品も多く存在し,人々の暮らしを豊かにしてき た1-4).小麦は,主食となる穀物であり,栄養源として重 要であるが,食物アレルギーに関する課題も多い.日本に おける食物アレルギーの食品別頻度は,鶏卵(38.3%), 乳製品(15.9%),小麦(8.0%)であり,三大アレルゲン として発症割合が高いことが知られている5).小麦アレル ゲンの低アレルゲン化には,これまで市販酵素であるセル ラーゼ・アクチナーゼを用いた方法6)や,小麦のエピトー プ解析とその配列を示した報告7-8)等があり,小麦アレル ゲン中の IgE 結合のエピトープ構造の解明が進歩してき た9).さらに,また,低アレルゲン小麦の製品開発10)に よる低アレルゲン化食品も模索されてきた. 一方,微生物を利用した伝統的な発酵法には,果実汁な どを用いた微生物により膨化する自然発酵法でスターター を作る方法がある.スターターを用いてドウを作り,ドウ の一部は次のスターターに引き継ぐ製法を老麺法といい, 中国では古くからこの老麺法を利用した饅頭や酒等があ る.著者らは,中国浙江省おいて伝統的な製法が継承され, * To whom correspondence should be addressed
E-mail:[email protected]
小麦粉発酵食品中の微生物による
小麦たんぱく質への作用
堀光代
1*,鈴木徹
2,長野宏子
2,3(
1岐阜市立女子短期大学,
2岐阜大学教育学部,
3岐阜大学連合農学研究科)
原稿受付 平成 24 年 2 月 27 日;原稿受理 平成 24 年 9 月 8 日Influence on Wheat Proteins of Microorganisms
in Traditional Fermented Food Products
Mitsuyo HORI
1* Tohru SUZUKI
2and Hiroko NAGANO
2, 3 1Gifu City Women’s College, Gifu 501-01922 The United Graduate School of Agriculture Science, Gifu 501-1193 3 Faculty of Education, Gifu University, Gifu 501-1193
SDS electrophoresis and antigen-antibody reactions were carried out in order to investigate the effects of bacterial enzymes isolated from Indian nan and chapati on the decomposition of allergenic proteins in wheat flour.
The salt-soluble and -insoluble fractions of the allergenic proteins were treated with the crude enzymes produced by microorganisms isolated from Nan and chapati. The results of antigen-antibody reactions indicated that only three microorganisms isolated from chapati, 00-IND-016-1, 00-IND-016-3 and 00-IND-016-4, could decompose the allergenic proteins.
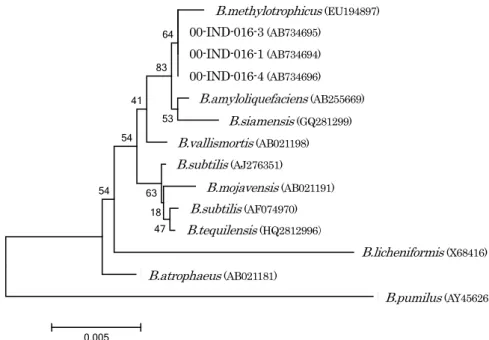
These three also exhibited 99% similarity to B. subtilis and B. amyloliquefaciens when analyzed with an API 50 CHB/E medium kit. A phylogenetic tree based on the 16SrRNA sequences showed that the three microorganisms were close to B. methylotrophicus and B. amyloliquefaciens.
饅頭や酒などを作る際にタデ科植物(ヤナギタデ)の汁を 用いたヤナギタデスターターに着目した.このヤナギタデ スターターから分離した微生物の産生する酵素は,小麦ア レルゲンたんぱく質を分解することを報告11)している. また Leu らは,モンゴルの小麦粉発酵食品からたんぱく 質分解酵素を産生する B.subtilis M2-4 を分離し,その諸 性質について報告12)している.このように,市販酵素で はなく,伝統発酵食品中の微生物作用により小麦アレルゲ ンたんぱく質を分解の可能性を見出すことは意義深い. これらの伝統発酵食品に関与している微生物の報告を積 み重ねることは,伝統の継承と科学的な解明につながり, さらに,近年進歩している遺伝学的な手法を用いた微生物 の分類も可能になってきている. そこで本研究は,インドのナンとチャパティのドウに存 在する微生物について,これまでに報告した伝統発酵食品 中の普遍性を確認し,プロテアーゼ活性の高い,低アレル ゲン化につながる酵素を産生する微生物の分離を行い,微 生物の産生する酵素の小麦たんぱく質へ分解作用を検討し た.微生物の同定を行い,今後の応用につながる知見を得 ることを目的とした. 2.実験試料および実験方法 (1)実験試料 小麦たんぱく質は,日清カメリア(日清製粉㈱製)の強 力粉を用いた.伝統的小麦粉発酵食品の微生物は,2000 年 8 月にインドで採取したナンとチャパティのドウを用 い,0.5%グルコースを含んだ NA(普通寒天)培地から分 離したものを用いた. (2)小麦たんぱく質の抽出および分画と反応条件 小麦たんぱく質の抽出は,Watanabe らの方法13)にて 行った.強力粉 5g に 0.5M 食塩溶液 25ml 加え,一晩(4℃) 抽出した後,遠心分離(10,000rpm 4℃ 60 分)し,上澄 液を塩可溶性画分とした.残渣は,4M 尿素溶液にて 60 分室温放置後,遠心分離 10,000rpm 4℃ 60 分)を行い, 塩不溶性画分とした.両画分を分画分子量 3,000 のセン トリプラス YM-3(Millipore 社製)を用い,遠心分離(3,000 × g,4℃ 290min)を行い,高分子部分,および低分子 部分に分画した.これらを凍結乾燥し,試料とした. (3)微生物検査法 1)微生物の形態観察 ナンおよびチャパティ製品中の微生物の観察は,ライト 染色14)を行った.ライト染色は,試料の内部から一欠片 採取し,アルコールで除菌したスライド上におき,滅菌 水に浸し,滅菌楊枝にて固定後,ライト染色液(MERCK, Wright’s eosin methylene blue solution, stabilized MERCK from microscopy 50% methanol)を数滴滴下し,1 分後,
蒸留水で余分な染色液を流し,乾燥させたものを光学顕 微鏡にて観察を行った.また,分離菌株のグラム染色も 行った. 2)微生物の生育試験 ナン及びチャパティに存在する微生物の確認は,光岡 の 方 法15)に よ り 行 っ た. す な わ ち,NA,LBS,DHL, TATAC,PEES,Potato dextrose の 6 種類の寒天培地を選 択培地とした.0.9%生理食塩水に懸濁した希釈溶液 1ml を各培地に塗布し,培養(37℃ 48 時間)後に微生物の生 育を観察した. 3)プロテアーゼ産生微生物の分離法 ナン(00-IND-005)およびチャパティ(00-IND-016) のドウからプロテアーゼ活性のある微生物の分離法を以下 に示した.NA(普通寒天)培地に生育した微生物をコラ ゲナーゼ誘導液体培地16)(りん酸水素 2 カリウム 0.7%, りん酸 2 水素カリウム 0.2%,硫酸マグネシウム 7 水和物 0.01%,クエン酸 2 水和物 0.05%,イーストエキス 0.1%, ゼラチン 0.3%,ポリペプトン 1.0%,グルコース 1.0%, pH7.4)に一白金耳植菌し,培養(37℃ 48 時間)した. その後,コラーゲンを含んだ選択培地に一白金耳植菌し, 培養(37℃ 48 時間)し,培地周辺に透明なハローを示す 独立したコロニーから微生物の分離を行った. (4)分離菌の特徴および小麦たんぱく質への作用 1)使用菌株 コラーゲンプレート培地にハローを示した分離菌株は, ナンの 8 株(00-IND-005-1 ~ 00-IND-005-8)とチャ パティの 8 株(00-IND-016-1 ~ 00-IND-016-8)であり, これらを実験に用いた. 2)微生物の生育確認とプロテアーゼ活性等微生物選択 試験 10% NaCl 生育試験は,普通ブイヨン(栄研化学㈱)に NaCl を加えた 10% NaCl 生育試験培地(普通ブイヨン 1.8%,寒天 1.5%,塩化ナトリウム 10%)を調整し,分 離菌株を一白金耳植菌し,培養(37℃ 24 時間)後,微生 物の生育を確認した. ゼラチン液化試験は,17%ゼラチン培地(普通ブイヨ ン 1.8%,寒天 1.5%,塩化ナトリウム 10%)に分離菌株 を白金線にて窄刺し,培養(37℃ 72 時間)した.培養後 冷蔵し,溶解性を確認した.再凝固しないものはゼラチン 分解活性を持つ菌株とした. 3)遊離アミノ酸定量 上記のゼラチン分解活性を持つ微生物については,分解 能試験を行い,基質(コラーゲン)に対する遊離アミノ酸 を定量した.前述のコラゲナーゼ誘導液体培地にて培養 (37℃ 48 時間)後,遠心分離した上清を粗酵素として用 いた.コラーゲン(Type Ⅰ ,Insoluble From Bovine
Achil-les Tendon, SIGMA 社 製 )0.01g に 0.1M リ ン 酸 緩 衝 液 (pH7.4)300μℓを加え均一に懸濁し,各菌株の粗酵素 を 200μℓ加え,30℃で 6 時間反応後,1 M 塩酸 1500 μℓで反応を停止した.これを 10 分遠心分離(10,000rpm 4℃)し,上清 100μℓと蒸留水 900μℓをニンヒドリン 反応17)にて吸光度(570nm)を測定した.対照には蒸留 水を,標準アミノ酸溶液は,ロイシンを用いた.遊離アミ ノ酸含量(mg)は,使用した粗酵素 1ml/1 時間あたりの 値とした. 4)小麦たんぱく質の分解試験 小麦たんぱく質試料は,凍結乾燥した塩可溶,塩不溶性 画分試料を 0.1M リン酸緩衝液(pH7.4)にて溶解後,ナ ンとチャパティから分離した微生物の粗酵素を加え,培養 (37℃ 6 時間)した.試料溶液は実際の調理操作を想定 し,加熱(100℃ 2 分)処理したものを遠心分離(13,000 rpm 4℃ 5 分)した.その後各上澄みを採取し,SDS 試料 用試薬と 2- メルカプトエタノールを加え,煮沸(3 分) 後に急冷した.さらに BPB 溶液を加え,SDS 電気泳動用 試料とした. 5)小麦たんぱく質の低アレルゲン化確認試験 分離菌株の中からプロテアーゼ活性の強い菌株の粗酵素 を用い,Laemmli の方法18)に従い,SDS 電気泳動用試料 を 15%ポリアクリルアミドゲルにて 30mA/ プレートに て泳動した.泳動後,CBB-R250(ナカライテスク㈱)で の染色を行った.抗原抗体反応は,SDS 電気泳動後,ゲル をニトロセルロースメンブラン(GE ヘルスケア㈱製)を 用いてウエスタンブロッティング(150mA/ プレート 1 時間)を行った.その後,0.3%スキムミルク入り PBS-T (Phosphate buffered saline-Tween 20)にてブロッキン
グ(4℃ 一晩)した.翌日,メンブランに PBS-T で 100 倍希釈した一次抗体(小麦アレルギー患者プール血清)を 反応(37℃ 1 時間)させた.PBS-T にて洗浄後,PBS-T で 500 倍希釈した二次抗体(抗ヒト IgE・ ウサギポリクロー ナル抗体パーオキシダーゼ標識,ダコ・ジャパン㈱製)を 反応(37℃ 1 時間)させた.PBS-T にて洗浄後,化学発 光試薬 ECL(GE ヘルスケア㈱製)を用い化学発光検出し, 暗室にて X 線フィルムに露光した. 小麦アレルギー患者プール血清の使用に際しては,岐阜 大学医学研究等倫理委員会の承認を得た. 6)分離菌の同定と系統樹作成 菌株は,生化学的特徴をみるため,API50CHB/E メディ ウムキット(シメックス・ビオメリュー㈱社製)を用い, 37℃で 24 時間および 48 時間培養し,色調の変化をみた. 得られた生化学プロファイルは,ソフトウエア(apiwebTM 細菌名検索用)を用いて同定した.また,遺伝的特徴につ いては,16SrRNA 遺伝子解析法19)を用いた.菌体からの
DNA 抽出は,Ultra Clean Microbial DNA Isolation Kit(MO BIO 社製)を用い,PCR 反応は Thermo-Start DNA Poly-merase(Thermo SCIENTIFIC 製)を使用した.プライマー は,f1L,f3L,r1L,r3L,1525r を 用 い た. 精 製 さ れ た PCR 産物は,Big Dye Terminator Cycle Sequencing Kit (Applied Biosystems)を用い,シークエンス反応を行っ た後,DNA Sequencer ABI PRISM Analyzer 3130(Applied Biosystems)にて約 1500 の塩基配列を決定した.得られ た塩基配列データは,データバンク(BLAST)に照合し, 相同性が 99%以上のものを推定菌属および菌種とした. これらの塩基配列データと,これまでに明らかになってい る遺伝子解析データと比較するため,ソフトウエア MEGA (Version 5.0)を用いて系統樹を作成した. 3.結果と考察 (1)インドのナンとチャパティの微生物について パンは紀元前 4000 年頃エジプトで食された記録があ り,発酵パンと無発酵パンがつくられている.生地を発酵 させて焼くナンや,膨化源を用いず,生地を薄く伸ばして 焼く無発酵パンとして平焼きパンがある.平焼きパンの チャパティは,中近東やインド等で好まれている20,21). 今回の試料は,2000 年 8 月にインドで採取したナンと チャパティとそれらのドウであり,その写真とドウ中の微 生物を Fig. 1 に示した.発酵パンのナンでは,球菌と短桿 菌が,無発酵パンとされるチャパティでは,ナンと同様に 球菌と短桿菌に加え細長い桿菌も観察された.しかし,酵 母は,顕微鏡下において両試料ともにみられなかった. 帰国後,ドウ中の微生物菌叢を 6 種類の選択培地を用 いて生育観察した.ナンは Bacillus 属などの一般細菌が生 育する NA 培地と Streptococcus 属が生育する TATAC 培 地上に生育がみられた.チャパティでは,一般細菌が生育 する NA のみに生育がみられた.酵母等が生育する Potato dextrose 寒天培地では,ナン,チャパティともに生育が みられなかった. 世界の伝統発酵食品中の微生物を観察した報告4)では, ナン中に酵母の存在を確認できた試料と酵母をまったく確 認できない試料がみられた.今回の試料の原材料は不明で あるが,酵母の存在割合が少なく,顕微鏡下において観察 できなかったことも考えられる.また,現地のフィールド 調査では,試料採取に限られた培地を用いることもやむを 得ない.今回,現地にて試料ドウは 0.5%グルコースを含 む NA(普通寒天)斜面培地に塗布したため,酵母の生育 に適さなかった可能性や酵母が死滅した可能性も考えら れる. 一般に小麦粉発酵食品には,酵母が必要であると考え られるが,今回の結果から,一般細菌や Streptococcus 属
などが存在しており,複合的な微生物の存在が明らかと なった. (2)分離菌の特徴とプロテアーゼ産生株の選択 試料のナンとチャパティのドウ中の微生物についてコ ラーゲンを含む培地上に生育させ,周囲にハローを形成し た独立したコロニーを,プロテアーゼ活性のある微生物と して選択分離し,各 8 菌株を分離した. 分離菌株のプロテアーゼ活性をみるために,ゼラチン分 解能試験および塩分含有を可能とする食品への応用を考 え,10% NaCl による耐塩性試験を行い,結果を Table 1 に示した.ナンから分離した微生物は,ゼラチン液化試験 において菌株により異なる結果を示し,10% NaCl 生育試 験では,すべての菌株に耐塩性がみられた.プロテアー ゼ活性および 10% NaCl 耐塩性のある菌株は,2 株(00-IND-005-4,00-IND-005-6)であった.チャパティから 分離した微生物では,ゼラチン液化試験において全ての 菌株にプロテアーゼ活性がみられた.10% NaCl 生育試験 では,分離した 8 株中 5 株(00-IND-016-2,00-IND-Fig. 1 Product (A), dough (B) and Wright’s stain (C) of Nan and Chapati
Table 1 Results of gelatin-liquefaction test and growth in 10% NaCl solution of microorganisms isolated from Nan and Chapati
Nan Chapati
(00-IND-005) (00-IND-016)
Strain Gelatin-liquefaction 10% NaCl Strain Gelatin-liquefaction 10% NaCl 00-IND-005-1 - + 00-IND-016-1 + - 00-IND-005-2 - + 00-IND-016-2 + + 00-IND-005-3 - + 00-IND-016-3 + + 00-IND-005-4 + + 00-IND-016-4 + + 00-IND-005-5 - + 00-IND-016-5 + + 00-IND-005-6 + + 00-IND-016-6 + - 00-IND-005-7 - + 00-IND-016-7 + + 00-IND-005-8 - + 00-IND-016-8 + - Geratin-liquefacitonl: + Liquefying 10% NaCl: + Growth
- Not liquefying - No growth Gelatin- liquefying gelatin and growth in 10% NaCl solution
抗体反応では,両画分とも小麦コントロールより濃いバ ンドが出現していた.塩可溶性画分における小麦たんぱ く質の主要アレルゲンは,15 kDa ~では,α-amylase / trypsin inhibitor family が知られ,当初は baker’s asthma
の原因物質としてあげられていた23)が,現在は,アトピー
性皮膚炎を主徴とするアレルギーにおいても主要アレルゲ
ンとしても知られている24).また,同じく塩可溶性画分
の主要アレルゲン Acyl-CoA oxidase は,27 kDa 付近にみ られる24).本実験結果の塩可溶性画分には 15 kDa 付近お よび 27 kDa 付近にバンドが出現していた.また,塩不溶 性画分における小麦たんぱく質の主要アレルゲンは,グリ アジンであり,30 kDa,44 ~ 55 kDa,60 kDa の各分子
量であることが報告されている24).本実験結果の塩不溶 性画分では,30 kDa 付近と,43 kDa 付近にバンドがみら 016-3,00-IND-016-4,00-IND-016-5,00-IND-016-7)に耐塩性がみられた.これらの分離した菌株のうち, ゼラチン活性および 10% NaCl 耐塩性が見られた 7 株に ついてプロテアーゼ活性を測定した.すなわち,コラーゲ ンを基質とし各微生物から産生する粗酵素を作用させ遊 離アミノ酸含量を測定し,その結果を Fig. 2 に示した.遊 離アミノ酸含量は,菌株により,2.4 ~ 29.6mg/ml と差 がみられ,チャパティから分離した菌株 00-IND-016-8 と 016-1 およびナンから分離した菌株 00-IND-005-6 は,基質コラーゲンに対する活性が強く,プロテ アーゼ活性の強い酵素を産生する菌株であることが示され た. 著者らは,中国浙江省に伝わるヤナギタデスターターか ら饅頭や酒を作る伝統的製法に着目し,ヤナギタデスター ターから分離した微生物のうち,遊離アミノ酸含量が高く, コラーゲンに対する活性の強い菌株は,小麦アレルゲンの 低減化がみられたことを報告11)しており,本実験試料に おいても,低アレルゲン化の可能性を推察できた. (3)分離菌産生酵素の小麦たんぱく質への作用 塩可溶および塩不溶たんぱく質を基質として,ナンと チャパティから分離した微生物(ナン 2 株,チャパティ 8 株)の産生する酵素を作用させた.酵素を作用させ,小麦 たんぱく質の分解と小麦アレルギー患者血清との抗原抗体 反応を検討した結果を Fig. 3,Fig. 4 に示した.小麦アレ ルギー患者により,小麦アレルゲン物質の塩可溶性画分へ の反応,塩不溶性画分への反応は異なるとの報告22)があ るため,塩可溶性画分および塩不溶性画分の両画分につい て検討を行った. ナンから分離した 2 菌株について SDS 電気泳動と抗原 抗体反応の結果(Fig. 3)では,塩可溶・塩不溶画分とも に小麦コントロールとほぼ同様のバンドがみられた.抗原
Fig. 3 SDS electrophoresis and antigen-antibody patterns of salt soluble and insoluble fractions of wheat proteins Wheat proteins were treated with crude enzymes produced by the microorganisms isolated from Nan. For antigen-antibody reaction, serum of wheat allergy patients was used. A: SDS-PAGE, B: Immunoblot
Lane M: Molecular weight marker, Lane C: Control (wheat flour), Lane 1: 00-IND-005-4, Lane 2: 00-IND-005-6
Fig. 2 The content of free amino acids produced in reaction of collagen to crude enzymes obtained from the microorganisms isolated from Nan and Chapati
いる可能性がみられた.抗原抗体反応の結果は,レーン 1, レーン 3,レーン 4 において,他のレーンより陰影が薄く, アレルゲンの分解を裏付ける結果を示した. 上述のように,SDS 電気泳動パターンおよび抗原抗体反 応バンドの結果から,小麦アレルゲンたんぱく質を分解し, また,基質コラーゲンに対する遊離のアミノ酸含量(Fig. 2) の結果より,レーン 1(00-IND-016-1)株,レーン 3 (00-IND-016-3)株,レーン 4(00-IND-016-4)株の 3 株をプロテアーゼ活性の強い菌株と判断し,微生物の同定 を行った. (4)分離菌の同定 分 離 し た 3 菌 株,00-IND-016-1,00-IND-016-3, 00-IND-016-4 を API50CHB/E メディウムキットを用い, 糖またはアミノ酸の菌による分解能等の菌株の生化学的 特徴について結果を Table 2 に示した.グラム染色結果 れた.したがって,本実験試料のナンでは,両画分ともに 小麦アレルゲンの分解はほとんどみられないことが明らか になった. チャパティから分離した 8 株の SDS 電気泳動と抗原抗 体反応の結果(Fig. 4)では,SDS 電気泳動では,塩可溶 性画分のすべての菌株において,15 kDa 付近に濃いバン ドが出現していた.レーン 2 は,小麦コントロール(レー ン C)と分解パターンが近似していた.抗原抗体反応にお いては,レーン 1,レーン 3,レーン 4 は,他の菌株に比 べ,17 kDa,27 kDa 付近のバンド薄くなっており,菌株 によって異なることが示唆された.また,塩不溶性画分で は,SDS 電気泳動結果のレーン 1,レーン 2,レーン 4 に おいて,他の菌株に比べてバンドが薄くなっていた.特に レーン 3,レーン 4 は,43 kDa ~ 14.4 kDa の広範囲に わたりバンドが消失しており,小麦アレルゲンを分解して
Fig. 4 SDS electrophoresis and antigen-antibody patterns of salt soluble and insoluble fractions of wheat proteins Wheat proteins were treated with crude enzymes produced by the microorganisms isolated from Chapati. For antigen-antibody reaction, serum of wheat allergy patients was used.
A: SDS-PAGE, B: Immunoblot
Lane M: Molecular weight marker, Lane C: Control (wheat flour), Lane 1: 00-IND-016-1, Lane 2: 00-IND-016-2, Lane 3: 00-IND-016-3, Lane 4:00-IND-016-4, Lane 5: 00-IND-016-5, Lane 6: 00-IND-016-6, Lane 7: 00-IND-016-7, Lane 8: 00-IND-016-8
は,3 菌株すべてグラム陽性,桿菌であった.分離した 3 菌株は,Salicin,D-sucrose,D-turanose の反応では,菌 株により違いがみられたが,他の指標は,3 株すべての反 応は一致していた.API50CHB の判定結果は,3 菌株とも B.subtilis および B.amyloliquefaciens と 99%の相同性を 示した.この API50CHB/E メディウムキットによる生化 学的同定は,基質に対する酵素作用により色調が変化し, その陽性率からソフトウエァで判定する方法である.この キットによる B.subtilis と B.amyloliquefaciens の区別は困 難であったため,16SrRNA 遺伝子解析法により同定を行っ た.その結果,分離した 3 菌株の配列は,API50CHB の 判定とは異なり,B.subtilis のみ 99%相同性を示しため, Table 2 Fermentation of carbohydrates by isolated microorganisms that were determined by API 50 CH
Strain
00-IND-016-1 00-IND-016-3 00-IND-016-4
Gram stain + + +
Form Rods Rods Rods
Glycerol + + + Erythritol - - - D-arabinose - - - L-arabinose + + + D-ribose + + + D-xylose + + + L-xylose - - - D-adonitol - - - Methyl-βD-xylopyranoside - - - D-galactose - - - D-glucose + + + D-fructose + + + D-mannose + + + L-sorbose - - - L-rhamnose - - - Dulcitol - - - Inositol + + + D-mannitol + + + D-sorbitol + + + Methyl-αD-mannopyranoside - - - Methyl-αD-glucopyranoside + + + N-Acetylglucosamine - - - Amygdalin - - - Arbutin - - -
Esculin ferric citrate + + +
Salicin + + -
D-cellobiose + + +
D-maltose + + +
D-lactose (bovine origin) + + +
D-melibiose - - - D-saccharose (sucrose) + - - D-trehalose + + + Inulin - - - D-melezitose - - - D-raffinose + + + Amidon (starch) + + + Glycogen - - - Xylitol - - - Gentiobiose - - - D-turanose + - - D-lyxose - - - D-tagatose - - - D-fucose - - - L-fucose - - - D-arabitol - - - L-arabitol - - - Potassium gluconate - - - Potassium 2-ketogluconate - - - Potassium 5-ketogluconate - - -
99.8%の相同性を示した.
これまでに伝統的小麦粉発酵食品中から分離した菌 株についての研究報告は,モンゴルの饅頭から分離し た B.subtilis M2-4(Accession No.AB379850)12), タ イ の 小 麦 粉 ド ウ か ら 分 離 し た B.subtilis DB(Accession No.AB440269)25-26)などがある.これらは,プロテアー ゼ活性が比較的高い菌株であり,小麦アレルゲンたんぱく 質の分解がみられた. 本研究において分離した 3 菌株も,同様にプロテアー ゼ活性が比較的高く,小麦アレルゲンたんぱく質の分解が 示唆された.従ってこれらの菌株が,小麦アレルゲンの低 減化への応用に新たな可能性を示す菌株であり,意義深い. さらに今後は最新の遺伝子解析手法も試みることが必要か もしれない. 4.まとめ インドのナンとチャパティから分離した微生物が産生す る粗酵素を用いて小麦粉抽出液によるアレルギーたんぱく 質の分解について以下の結果を得た. (1) インドのナンとチャパティから分離した微生物は, Bacillus 属などの一般細菌や Streptococcus 属が生育 していた.コラーゲンに対するプロテアーゼ活性の強 い酵素を産生する微生物の菌株は,チャパティから分 離した 1(00-IND-016-1)株と 8(00-IND-016-8) 株の 2 菌株であった. (2) 微生物が産生する粗酵素を用いての小麦たんぱく質の 系統解析を行った.系統解析に使用した菌株は,今回分離 した 3 菌株,00-IND-016-1(Accession No.AB734694), 00-IND-016-3 (Accession No.AB734695),00-IND-016-4(Accession No.AB734696) と,B.subtilis の 基 準株 B.subtilis(Accession No.AJ276351)および基準株 と 99%以上の相同性を示す菌株を選択した.すなわち, B.subtilis subsp.spizizenii(Accession No.AF074970), B.mojavensis(Accession No.AB021191),B.vallismortis (Accession No.AB021198),B.atrophaeus(Accession No.
AB021181),B.licheniformis(Accession No.X68416), B.methylotrophicus(Accession No.EU194897), B.siamensis(Accession No.GQ281299),B.tequilensis (Accession No.HQ223107)の 8 菌株とした.また,先 の API50CHB/E メディウムキット判定結果に示された B.amyloliquefaciens(Accession No.AB255669) を 加 え, アウトグループは,B.pumilus (Accession No.AY456263) とした.これらを Clastal W を用いて系統樹を作成し た(Fig. 5).その結果,インドのチャパティから分離 し た 3 菌株(IND-016-1,IND-016-3,IND -016-4) は,B.methylotrophicus(Accession No.EU194897), B.amyloliquefaciens(Accession No.AB255669)と近縁で あることが明らかとなった. 分離した3株間では,00-IND-016-1(Accession N o . A B 7 3 4 6 9 4 ) と 0 0 - I N D - 0 1 6 - 3 ( A c c e s s i o n No.AB734695)の 2 株は 100%一致していた.00-IND -016-4(Accession No.AB734696) は, 上 記 2 株 と
Fig. 5 Phylogenetic tree based on 16SrRNA sequences. Tree was constructed by the neighbor-joining method. The type strain of B.pumilus was used for out group
; and Watanabe M. A major wheat allergen has a Gln-Gln-Gln-Pro-Pro motif identified as an IgE-binding epitope. Biochem. Biophys. Res. Commum. 1996, vol.219, p.290-293
8) Matsuo H. ; Kohno K. ; Morita E. Molecular cloning, re-combinant expression and IgE-binding epitope of ω-5 gliadin, a major allergen in wheat-dependent exercise-induced anaphylaxis. FEBS. J. 2005, vol.272, p.4431-4438 9) 中村丁次,板垣康治,池沢善郎,栗原和幸,手島玲子, 高松伸枝,鈴木志保子,杉山久仁子,土橋朗.食物アレ ルギーA to Z.第一出版,2010, p.64 10) 田辺創一,渡辺純,園山慶,渡辺道子;難治性の小麦ア レルギーに挑む.科学と生物,2001,vol.39,no.7, p.440-447 11) 堀光代,長野宏子,;タデ科植物の中国伝統食品への利 用.日本調理科学会誌.2005,vol.38. no.1,p.51-57 12) LIU C. ; and Nagano H. Purification and Characterization
of a Priteolytic Enzyme from Bacillus subtilis M2-4. J. Home Econ Jpn. 2008, vol. 59, no.8, p.565-573
13) Watanabe M. ; Suzuki T. ; Ikezawa Z. ; and Arai S. Con-trolled enzymatic treatment of wheat proteins for pro-duction of hypoallergenic flour. Biosci. Biotechnol. Bio-chem. 1994, vol.58, no.2, p.388-390
14)佐野豊(1972),組織学研究法,南山堂,東京,p302 15) 光岡知足(1972),腸内細菌の世界改訂版,叢文社,
東京,p53
16) Hisano T. ; Abe, S. ; Wakashiro M. ; Kimura A. ; andMu-rata K. Isolation and Properties of Collagenese with Ca-seinolytic Activity from Psudomoonas sp. J. Ferment and Bioeng. 1989, vol.68, no.6, p.399-403
17) H. Rosen. A modified ninhydrin colorimetric analysis for amino acid. Achive. Biochem. Biophys. 1957, vol.67, p.10-15
18) Leammli, U. K. ; Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature. 1970, vol.227, p.680-685 19) 篠田吉史,加藤暢夫,森田直樹.16SrRNA遺伝子解析 による細菌の系統分類法.島津評論.2000,vol.57, no.1・2, p121-132 20) 岡田哲.“パン-6000年の歴史をたどる.”コムギの食文 化を知る事典.東京堂出版,2001,p.84-86 21) 有村誠,大田正次,河原太八,加藤鎌司,笹沼恒男,佐 藤洋一郎,武田和義,丹野研一,辻本壽,冨永達,長野 宏子,西田英隆,森直樹,森川利信,吉村作治,渡部 武.“フィールドからみた世界のパン.”麦の自然史.佐 分解は,抗原抗体反応における小麦たんぱく質の塩可 溶性・塩不溶性の両画分の結果から,チャパティから 分離菌株,1(00-IND-016-1)株,3(00-IND-016-3) 株,4(00-IND-016-4)株にアレルゲンの分解が示 された. (3) アレルゲンの分解が示された分離菌株の 1(00- IND -016-1)株,3(00- 016-3)株,4(00- IND-016-4)株は,API50CHB/E メディウムキットでは, B.subtilis お よ び B.amyloliquefaciens と も に 99 % の 相同性を示した.また,16SrRNA 遺伝子解析結果で は,B.subtilis と 99%相同性を示した.系統樹では, B.methylotrophicus,B.amyloliquefaciensと近縁で あった. 謝 辞 本研究の一部は,平成 20 年度文部省科学研究費補助金 (基盤 C 20500698)によって遂行されました.ここに付 記して謝意を表します.また,分離菌株の 16SrRNA 遺伝 子解析についてお世話になりました岐阜大学遺伝子セン ター脇原祥子氏に心より御礼申し上げます. 引 用 文 献
1) Nagano, H. ; Omori, M. ; Shoji, Z. ; Iibuti, S. ; Arai, M. Iden-tification and Characteristics of Microorganisms Isolated from Traditional Wheat Flour Dough in Thailand. J. Home Econ. Jpn. 1994, vol. 45, no. 3, p.219-226
2) Okada S. ; Ikezawa M. ; Yoshida I. ; Uchimura T. ; Ohara N. ; and Kozaki M. Identification and Characteristics of Lactic Acid Bacteria Isolation from SourDough Sponges. Biosci. Biotech. Biochem. 1992, vol 56, no.4, p.572-575
3) 武田泰輔,岡田早苗,小崎道雄.パン生地,生イースト およびドライイースト中の乳酸菌の特性.日本食品工業 学会誌.1984, vol. 31, no.10, p.642-648 4) 長野宏子,説田佑子,粕谷志郎.伝統的な小麦粉発酵食 品中の微生物とその働き.日本家政学会誌.2003, vol. 54, no.9, p.713-721 5) 海老沢元宏.“食物等によるアナフィラキシー反応の原因 物質(アレルゲン)の確定,予防,予知法の確立に関す る研究.”平成15年厚生労働省科学研究費補助金,免疫ア レルギー疾患予防・治療等研究事業研究報告書,2004, p.89-91
6) Watanabe M. ; Suzuki T. ; Ikezawa Z. ; and Arai S. Con-trolled enzymatic treatment of wheat proteins for produc-tion of hypoallergenic flour. Biosci. Biotechnol. Biochem. 1994, vol.58, no.2, p.388-390
藤洋一郎,加藤鎌司編著.北海道大学出版会,2010, p.262-263
22) Kimoto M. ; Yoshikawa M. ; Takahashi K. ; BandoN. ; Oki-ta M. ; and Tsuji H. Identification of Allergens in Cereals and Their Hypoallergenization, I. Screening of Allergens in Wheat and Identification of an Allergen, Tri a Bd 17K. Ann. Rep. Interdiscipl. Res. Inst. Environ Sci. 1998, vol.17, p.53-60
23) J. M. James. ; J. P. Sixbey. ; R. M. Helm. ; G. A. Bannon. ; and A. W. Burks. Wheat alpha-amylase inhibitor : a second route of allergic sensitization. J. Allergy Clin. Im-minol. 1997, vol.99, p.239-244
24) 小川正,篠原和毅,新本洋士.“アレルギーの発症機 構”抗アレルギー食品開発ハンドブック.㈱サイエンス フォーラム,2005,p.23
25) Phromraksa P. ; Nagano H. ; Boonmars T. ; and Kam-boonrung C. Indentification of proteolytic bacteria from Thai traditional fermented foods and theirallergenic re-ducing potentials. J. Food Sci. 2008, vol.73, nr.4, M189-195
26) Phromraksa P. ; Nagano H. ; Kanamaru Y. ; Izumi H. ; Yamada C. ; and Kamboonrung C. Characterzation of Ba-cillus subtilis Isolated from Asain Fermented Food. Food