九州大学学術情報リポジトリ
Kyushu University Institutional Repository
口腔扁平上皮癌の浸潤・転移におけるΔNp63を介し た上皮-間葉転換に関する研究
後藤, 雄一
九州大学大学院歯学府
https://doi.org/10.15017/21984
出版情報:Kyushu University, 2011, 博士(歯学), 課程博士 バージョン:
権利関係:
- 29 -
1-2. ΔNp63ノックダウンが OSCC細胞の増殖、分化、遊走およびアポトーシ
ス与える影響
次に、ΔNp63ノックダウンによるOSCC細胞の増殖、分化、遊走およびアポトー シスへの影響について検討した。
HSC-2細胞へのΔNp63 siRNAを導入により、ΔNp63の発現は著明に抑制された
(図3)。ΔNp63 siRNA導入48時間後の細胞形態を観察したところ、多角形であっ
たHSC-2の形態はΔNp63ノックダウンにより紡錘形へと変化した(図4)。WST-1
assayにおいて、ΔNp63 siRNA導入群は対照群と比べ細胞増殖が有意に抑制されて
いた(図5 A; ANOVA, *p<0.05)。BrdU incorporation assayでは、ΔNp63 siRNA導入 群のBrdU陽性細胞率は対照群と比較して有意に低下していた(図5 B;
Mann-Whitney U-test, *p<0.05)。アポトーシスが誘導されていることを示すannexin V 陽性細胞は、有意差は認められなかったものの、対照群と比較してΔNp63 siRNA 導入群において多く認められた(図6)。また、Bcl-2の発現量をreal-time PCRによ り検索した結果、ΔNp63ノックダウンによりBcl-2の発現量は有意に低下した(図 7; Mann-Whitney U-test, *p<0.05)。wound healing assayによる遊走能の比較では、
ΔNp63 siRNA導入群は間隙形成後24時間で間隙がほぼ消失したが、対照群では間 隙が残存し、ΔNp63ノックダウンにより遊走能の亢進が認められた(図8)。さら
に、real-time PCRにおいてEMT関連遺伝子の発現量を検索したところ、ΔNp63ノ
ックダウンによりCK5およびCK14の発現量は低下し、vimentinとN-cadherinの発 現量は増加したが(Mann-Whitney U-test, *p<0.05)、E-cadherinの発現量に変化は認 められなかった(図9)。
図3 ΔNp63 siRNA導入によるΔNp63の発現
HSC-2細胞へのΔNp63 siRNA導入により、ΔNp63のmRNAおよびタンパク質の発現 はともに低下した。
siCtrl: scrambled siRNA導入HSC-2細胞 siRNA: ΔNp63 siRNA導入HSC-2細胞
図4 ΔNp63ノックダウンがOSCC細胞の形態に与える影響
ΔNp63ノックダウンにより、多角形であったHSC-2の細胞形態は紡錘形に変化した。
scale bar, 20 μm
siCtrl siRNA
Δ Np63 β-actin
siCtrl siRNA
Western blotting
Δ Np63 GAPDH
siCtrl siRNA
RT-PCR
- 30 -
siCtrlsiRNA
scale bar, 50 μm 0
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
0 24 48 72 96
吸光度 ( A
450 nm-A
620 nm)
時間( h )
*
siCtrl siRNA
A
B BrdU PI merge
0 5 10 15 20 25 30 35 40 45
50
*
BrdU 陽性細胞率 (%)
siCtrl siRNA
図5 ΔNp63ノックダウンがOSCC細胞の増殖活性に与える影響(A; WST-1 assay, B; BrdU incorporation assay)
ΔNp63ノックダウンにより、ΔNp63 siRNA群は対照群に比べ細胞増殖が有意に 抑制された。
WST-1におけるグラフは、独立した3回のデータの平均および標準偏差を示して
おり、2群間に統計学的有意差を認めた。BrdU incorporation assayにおけるグラフは、
無作為に選択した5視野のBrdU陽性細胞率の平均および標準偏差を示しており、2群 間に統計学的有意差を認めた(WST-1 assay; ANOVA *p<0.05, BrdU incorporation assay; Mann-Whitney U-test *p<0.05)。
- 31 -
annexin V hoechst PI merge
siRNA siCtrl
annexin V 陽性細胞率 (%)
0 5 10 15 20
0 1 2 3 4
相対的発現量
siCtrl siRNA
scale bar, 50 μm
siCtrl siRNA
図6 ΔNp63ノックダウンがOSCC細胞のアポトーシスに与える影響
annexin V陽性細胞率は、ΔNp63 siRNA導入群において有意な差を認めなかったもの
の高かった。
グラフは、無作為に選択した5視野におけるannexin V陽性細胞率の平均および標準 偏差を示している(Mann-Whitney U-test)(N.S.: not significant)。
*
図7 ΔNp63ノックダウンがOSCC細胞のBcl-2発現に与える影響 ΔNp63ノックダウンにより、Bcl-2の発現量は有意に低下した。
グラフは、独立して行った3回のデータの平均および標準偏差を示しており、
2群間に統計学的有意差を認めた(Mann-Whitney U-test *p<0.05)。
Bcl-2
N.S.
- 32 -
siCtrl siRNA
0h
24h
図8 ΔNp63ノックダウンがOSCC細胞の遊走能に与える影響 ΔNp63 siRNA群の遊走能は対照群と比較して亢進していた。
scale bar, 200 μm
- 33 -
図9 ΔNp63ノックダウンがOSCC細胞におけるEMT関連遺伝子の発現に与える影響 ΔNp63ノックダウンにより、上皮系マーカーであるCK5およびCK14の発現量は有 意に低下したが、間葉系マーカーであるvimentinおよびN-cadherinの発現量は有意に 増加した。E-cadherinの発現量に変化は認められなかった。グラフは、独立して 行った3回のデータの平均および標準偏差を示しており、2群間に統計学的有意差を 認めた(Mann-Whitney U-test *p<0.05)(N.S.: not significant)。
0 0.2 0.4 0.6 0.8 1 1.2
ΔNp63
*
89.0%
相対的発現量
siCtrl siRNA
00.2 0.4 0.6 0.8 1 1.2
E-cadherin
相対的発現量
siCtrl siRNA
0 0.2 0.4 0.6 0.8 1 1.2
CK5
0 0.2 0.4 0.6 0.8 1 1.2
CK14
* *
相対的発現量 相対的発現量
siCtrl siRNA siCtrl siRNA
0 1 2 3 4 5 6 7 8
vimentin
*
0 5 10 15 20
N-cadherin
*
siCtrl siRNA siCtrl siRNA
相対的発現量 相対的発現量
N.S.
- 34 -
- 35 -
1-3. ΔNp63過剰発現が OSCC細胞の増殖、分化、遊走およびアポトーシスに
与える影響
さらに、ΔNp63過剰発現がOSCC細胞の増殖、分化、遊走およびアポトーシスに 与える影響について検討した。
ΔNp63の発現を認めなかったSQUU-B細胞にΔNp63発現ベクターを導入し、
ΔNp63過剰発現細胞株(SQUU-BO)を樹立した。陰性対照としてemptyベクター を導入した細胞株(SQUU-BC)を用いた。RT-PCRおよびWestern blottingにより、
SQUU-BO細胞はΔNp63を発現していることが確認された(図10)。ΔNp63過剰発
現による細胞形態の違いを観察したところ、SQUU-BC細胞は紡錘形を示していた
が、SQUU-BO細胞では多角形の細胞を多く認め、敷石状の増殖を示した(図11)。
WST-1 assayでは、播種後120時間までは両群間に差を認めなかったが、144時間
後以降において、SQUU-BO細胞はSQUU-BC細胞と比較して有意に細胞増殖が亢 進していた(図12 A; Student’s t-test, *p<0.05)。またBrdU incorporation assayにおい
て、SQUU-BO細胞はSQUU-BC細胞と比較してBrdU陽性細胞率が有意に高かっ
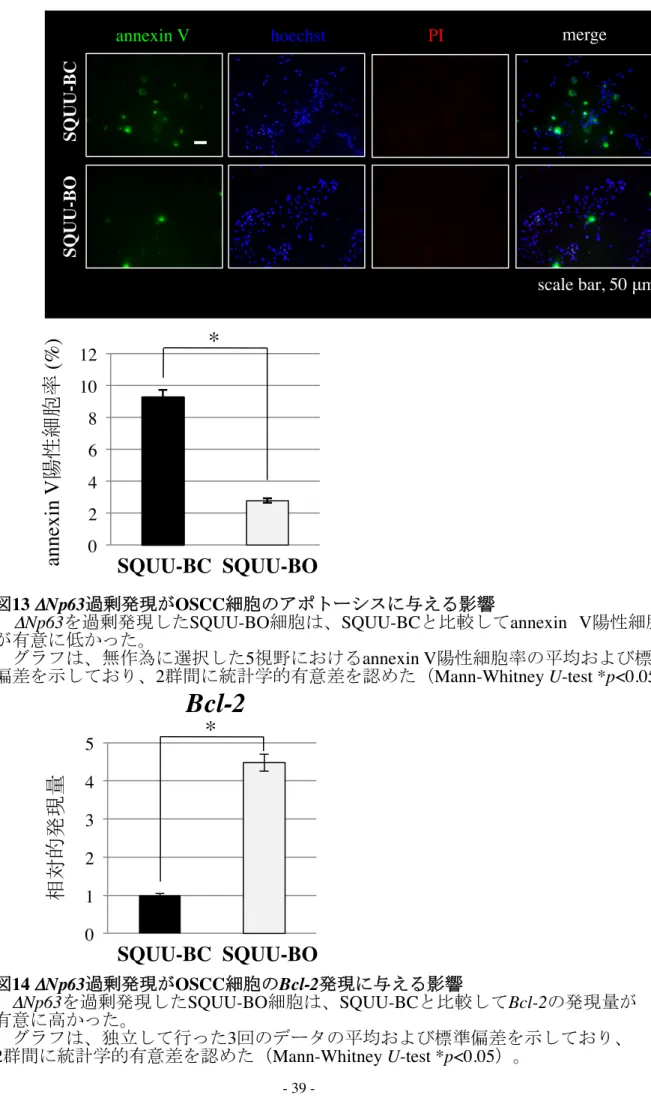
た(図12 B; Mann-Whitney U-test, *p<0.05)。annexin V 陽性細胞率の比較では、
SQUU-BO細胞はSQUU-BC細胞と比較して陽性率が有意に低く、アポトーシスが
抑制されていた(図13; Mann-Whitney U-test, *p<0.05)。さらに、real-time PCRにて Bcl-2の発現量を比較したところ、SQUU-BO細胞ではSQUU-BC細胞に比べBcl-2 の発現量が有意に高かった(図14; Mann-Whitney U-test, *p<0.05)。wound healing
assayによる遊走能の比較では、間隙形成24時間後において、SQUU-BO細胞は
SQUU-BC細胞と比較し間隙の幅が大きく、遊走能の低下が認められた(図15)。
real-time PCRを用いてEMT関連遺伝子の発現量を検索したところ、SQUU-BO細胞
はSQUU-BC細胞と比較してCK5およびCK14の発現量が高く、vimentinと
N-cadherinの発現量は低かった(Mann-Whitney U-test, *p<0.05)。E-cadherinの発現
- 36 -
量に有意な差は認められなかった(図16)。
Δ Np63 β-actin
SQUU-BC SQUU-BO
Western blotting
Δ Np63 GAPDH
SQUU-BC SQUU-BO
RT-PCR
scale bar, 20 μm
SQUU-BC SQUU-BO
図10 ΔNp63発現ベクター導入によるΔNp63の発現
ΔNp63発現ベクターを導入したSQUU-BOにおいてΔNp63の発現を認めた。
SQUU-BC: emptyベクター導入SQUU-B細胞 SQUU-BO: ΔNp63発現ベクター導入SQUU-B細胞
図11 ΔNp63過剰発現がOSCC細胞の形態に与える影響
SQUU-BC細胞は紡錘形を示したが、SQUU-BO細胞は多角形を示し、敷石状に増
殖していた。
- 37 -
0 10 20 30 40 50 60
SQUU-BCSQUU-BO
0 0.1 0.2 0.3 0.4
0 24 48 72 96 120 144 168
*
SQUU-BO SQUU-BC
*
scale bar, 50 μm
吸光度 ( A
450 nm-A
620 nm)
時間( h )
A
B BrdU PI merge
*
BrdU 陽性細胞率 (%)
SQUU-BC SQUU-BO
図12 ΔNp63過剰発現がOSCC細胞の増殖活性に与える影響(A; WST-1 assay, B; BrdU incorporation assay)
WST-1 assayおよびBrdU incorporation assayにおいて、SQUU-BO群は対照群に比 べ、細胞増殖が有意に亢進していた。
WST-1におけるグラフは、独立した3回のデータの平均および標準偏差を示して
おり、2群間に統計学的有意差を認めた。BrdU incorporation assayにおけるグラフ は、無作為に選択した5視野のBrdU陽性細胞率の平均および標準偏差を示してお り、2群間に統計学的有意差を認めた(WST-1 assay; Student’s t-test *p<0.05, BrdU incorporation assay; Mann-Whitney U-test *p<0.05)。
- 38 -
0 1 2 3 4 5 0 2 4 6 8 10 12
scale bar, 50 μm
SQUU-BCSQUU-BO
annexin V hoechst PI merge
annexin V 陽性細胞率 (%) 相対的発現量
SQUU-BC SQUU-BO
SQUU-BC SQUU-BO
図13 ΔNp63過剰発現がOSCC細胞のアポトーシスに与える影響
ΔNp63を過剰発現したSQUU-BO細胞は、SQUU-BCと比較してannexin V陽性細胞数 が有意に低かった。
グラフは、無作為に選択した5視野におけるannexin V陽性細胞率の平均および標準 偏差を示しており、2群間に統計学的有意差を認めた(Mann-Whitney U-test *p<0.05)。
*
図14 ΔNp63過剰発現がOSCC細胞のBcl-2発現に与える影響
ΔNp63を過剰発現したSQUU-BO細胞は、SQUU-BCと比較してBcl-2の発現量が 有意に高かった。
グラフは、独立して行った3回のデータの平均および標準偏差を示しており、
2群間に統計学的有意差を認めた(Mann-Whitney U-test *p<0.05)。
Bcl-2
*
- 39 -
SQUU-BC SQUU-BO
0h
24h
図15 ΔNp63過剰発現がOSCC細胞の遊走能に与える影響
SQUU-BO細胞の遊走能は対照群と比較して低下していた。
scale bar, 200 μm
- 40 -
図16 ΔNp63過剰発現がOSCC細胞におけるEMT関連遺伝子の発現に与える影響 上皮系マーカーであるCK5およびCK14の発現量は有意に高く、間葉系マーカー であるvimentinおよびN-cadherinの発現量は有意に低下した。E-cadherinの発現量に 変化は認められなかった。グラフは、独立して行った3回のデータの平均および標 準偏差を示しており、2群間に統計学的有意差を認めた(Mann-Whitney U-test
*p<0.05)(N.S.: not significant)。
0 200 400 600 800 1000
ΔNp63
0 0.2 0.4 0.6 0.8 1 1.2 1.4
E-cadherin
0 2 4 6 8
CK14
0 2 4 6 8 10 12
CK5
0 0.5 1 1.5 2
N-cadherin
0 0.5 1 1.5 2 2.5
vimentin
*
相対的発現量
SQUU-BC SQUU-BO
相対的発現量
SQUU-BC SQUU-BO
相対的発現量 相対的発現量
SQUU-BC SQUU-BO SQUU-BC SQUU-BO
SQUU-BC SQUU-BO SQUU-BC SQUU-BO
相対的発現量 相対的発現量
*
* *
*
N.S.
- 41 -
- 42 -
研究 2
OSCC生検材料における ΔNp63、vimentinおよび E-cadherinの免疫組織化学的検 討
2-1. OSCC生検材料における ΔNp63、vimentinおよび E-cadherinの発現様式 OSCC生検材料を用いてΔNp63、E-cadherinおよびvimentinの発現を免疫組織 化学的に二重染色を用いて検索した。OSCCの腫瘍中心部では、ΔNp63および
E-cadherinはほぼ全ての癌細胞に発現を認めたが、vimentinの発現は認めなかっ
た(図17 b, c)。一方、浸潤先端部では、ΔNp63およびE-cadherinの発現が減弱
している癌細胞を認め、これらはvimentinの発現が亢進していた(図17 e, f)。
2-2. 浸潤先端部におけるvimentinの発現および ΔNp63発現強度の減弱の有無
と臨床病理組織学的所見との関連
浸潤先端部の癌細胞のvimentin陽性細胞率を算出し、臨床病理組織学的所見との 関連について検討した。vimentin陰性群(<5%)に比べ、陽性群(≥5%)では、頸 部リンパ節転移と遠隔転移の発生頻度が有意に高かった。また、Stage分類では
vimentin陰性群に比べ陽性群では病期が進行した症例が多かった。さらに、浸潤先
端部の癌細胞におけるΔNp63発現強度の減弱の有無についても同様に検討を行っ た。ΔNp63発現強度の減弱を有する群では頸部リンパ節転移と遠隔転移の発生頻度 が有意に高かった(表5)。その他の臨床病理組織学的所見には関連が認められなか った。
疾患特異的累積5年生存率は、vimentin陽性群で69.8%、vimentin陰性群では88.6%
であり、vimentin陽性群は陰性群と比較し有意に低かった。同様に浸潤先端部で
ΔNp63発現強度の減弱を有する群の疾患特異的累積5年生存率は67.6%、認めない
群は87.7%であり、ΔNp63の発現強度の減弱を有する群は認めない群と比較して有
- 43 -
意に低く予後不良であった(図18; log-rank test *p<0.05)。
a b c d e f
a d
HE Δ Np63 / vimentin E-cadherin / vimentin
腫瘍中心部 浸潤先端部
scale bar , 50 μ m scale bar , 200 μ m
HE
図17 OSCC生検材料におけるΔNp63、vimentinおよびE-cadherinの発現様式(A; 弱拡大像、B; 強拡大像、a-c; 腫瘍中心部、d-f; 浸潤先端部) ΔNp63とvimentinの二重染色では、腫瘍中心部の癌細胞のほぼすべてにΔNp63の発現を認めたが、vimentinの発現は認めなかった(b)。 一方、浸潤先端部の癌細胞ではΔNp63の発現強度の減弱を認め、これらの細胞ではvimentinの発現が亢進していた(e)。 E-cadherinとvimentinの二重染色では、ΔNp63と同様に浸潤先端部の癌細胞でE-cadherinの発現強度の減弱を認め、vimentinの発現が亢進していた (f)。 bとeの青色はΔNp63、茶色はvimentinを示し、cとfの青色はE-cadherin、茶色はvimentinを示す。B A
- 44 -
- 45 -
症例数 (%) 陽性 陰性 p-value 有 無 p-value
54 24 42 24 9 3 8 13 57 16 34 8 20 15 27 12 24 59 14 5 2 16 41 15 4 67 11 41 37 70 8
vimentinの 発現 ΔNp63発 現強 度の 減 弱
(69.2) (30.8) (53.9) (30.8) (11.5) (3.8) (10.2) (16.7) (73.1) (20.5) (43.6) (10.3) (25.6) (19.2) (34.6) (15.4) (30.8) (75.6) (18.0) (6.4) (2.6) (20.5) (52.6) (19.2) (5.1) (85.9) (14.1) (52.6) (47.4) (89.7) (10.3) 性 別
原 発部 位
臨 床発 育様 式
T分 類
臨 床病 期分 類
分 化度 (Grade分 類)
浸 潤様 式(YK分 類)
局 所再 発
頸 部リ ンパ 節 転移
遠 隔転 移 男性 女性 舌 歯肉 口底 頬粘膜 表在型 外向型 内向型 T1 T2 T3 T4
Ⅰ
Ⅱ
Ⅲ
Ⅳ Grade 1 Grade 2 Grade 3 Grade 1 Grade 2 Grade 3 Grade 4C Grade 4D 無
有 無 有 無 有
32 11 18 14 8 3 2 6 35 4 19 7 13 3 14 9 17 28 11 4 1 6 19 13 4 38 5 15 28 35 8
22 13 24 10 1 0 6 7 22 12 15 1 7 12 13 3 7 31 3 1 1 10 22 2 0 29 6 26 9 35 0
N.S.
N.S.
N.S.
N.S.
p<0.05
N.S.
N.S.
N.S.
p<0.01 p<0.01
23 14 20 11 4 2 5 5 27 10 15 4 8 9 12 5 11 31 3 3 2 2 20 10 3 31 6 11 26 30 7
31 10 22 13 5 1 3 8 30 6 19 4 12 6 15 7 13 28 11 2 0 14 21 5 1 36 5 30 11 40 1
N.S.
N.S.
N.S.
N.S.
N.S.
N.S.
N.S.
N.S.
p<0.01
p<0.05
表 5 OSCC浸潤先端部 における vimentinの発現および ΔNp63発現強度の
減弱の有無 と臨床病理組織学的所 見との関連
OSCC 患者において、vimentin陰性群に比べ陽性群は頸部リンパ節転移および遠隔 転移の発生頻度が有意に高かった。ΔNp63発現強度の減弱を有する群においても同様 にこれらの発現頻度が有意に高かった。なお、統計処理には𝑥2 testまたはFisher’s exact testを用いた(N.S.: not significant)。
- 46 -
浸潤先端部における ΔNp63発現強度の減弱
無(n=41) 有(n=37)
A
B
図 18 疾患特異的 累積 5 年生存率(A; vimentin 陽性群と陰 性群の比較、
B; ΔNp63発現強度の 減弱の有無による比較)
浸潤先端部におけるvimentin陽性群は、陰性群と比較し生存率が有意に低か った。同様に、ΔNp63発現強度の減弱を有する群は、減弱がない群と比較し生 存率が有意に低く、予後不良であった(log-rank test *p<0.05)。
vimentin陰性群(n=35) vimentin陽性群(n=43)
- 47 -
考 察
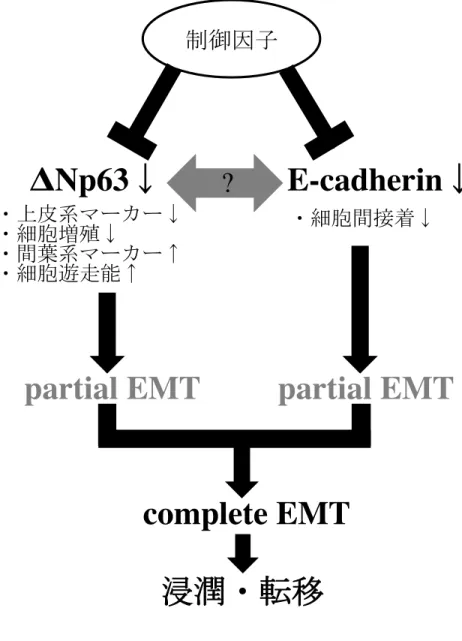
近年、癌の浸潤・転移において器官形成に必須の現象であるEMTが関与してい ることが明らかになってきた(21, 22, 33)。EMTが誘導された細胞は線維芽細胞 様の形態変化、上皮系マーカーの発現量減少、間葉系マーカーの発現量増加、遊 走能の亢進、増殖能の低下および細胞間接着の減弱などの表現型を示し、このよ うな細胞特性が癌の浸潤・転移に寄与していることが示唆されている(24, 25, 60)。 癌抑制遺伝子p53のホモログであるΔNp63は癌の増殖や分化に関与していること が報告されているが、EMTに関連している可能性も示されている(61)。しかし ながら、その詳細については不明な点が多い。本研究ではΔNp63に着目し、EMT と癌の浸潤・転移との関連について検討を行った。
Hay(1995)により初めて提唱されたEMTという概念は、多くの研究者により 実証され、様々なEMTの形質が報告されてきた(26, 27, 29, 30)。そのため、近年 では、EMTの定義についても議論がなされており、2008年のCold Spring Harbor で開催されたEMTのコンセンサス会議の後、Zeisberg(2009)らは以下の特徴を EMTを獲得した細胞の形質として定義した(62)。
・vimentinの発現
・E-cadherin が消失し N-cadherin が発現する cadherinスイッチ
・Snailや Slugなどの転写因子の発現
・cytokeratinなどの上皮系マーカーの発現消失
・紡錘形の細胞形態
・遊走能の亢進
・アポトーシスへの抵抗
本研究で用いたSQUU-B細胞やΔNp63をノックダウンしたHSC-2細胞は、上
- 48 -
記の形質の多くを有しており、EMTが誘導されている可能性が示された。しかし、
上記の定義に合致しない事象が2点認められた。まず第1点は、SQUU-B細胞に
おいてE-cadherinの発現を認め、ΔNp63をノックダウンしたHSC-2細胞において
もE-cadherinの発現量に変化を認めなかったこと、すなわちE-cadherinの消失が 認められなかった点である。E-cadherinは、上皮細胞に特異的に発現している細胞 間接着因子であり、その消失は代表的なEMTの形質と考えられている(26, 63, 64)。 E-cadherinは、増殖抑制因子である形質転換増殖因子β(transforming growth factor β:
TGFβ)との関連が最も良く知られており、TGFβが発現すると下流の転写因子で
あるSnailおよびSlugの発現が誘導され、その結果E-cadherinの発現が低下するこ
とが判っている(65-69)。また、TGFβにより誘導されたSnailやSlugはΔNp63 の発現も抑制すると考えられており、実験的にSnailなどの転写因子を癌細胞に過 剰発現させると、E-cadherinやΔNp63は消失し、EMTが誘導されることが示され
ている(70-72)。しかしながら、今回の研究においてE-cadherinの発現消失が認め
られなかったことは、E-cadherinがΔNp63とは異なる因子により制御されている か、ΔNp63の上流で制御されている可能性が示された。また、E-cadherinの発現が 消失しなかったことを別の観点から考えると、SQUU-B細胞が均一な集団ではな く、様々なEMTの段階を獲得した細胞が混在した不均一な細胞集団であることが 推察される。これは、免疫細胞化学的染色において、vimentinの発現が亢進してい る細胞の一部でE-cadherinの発現が減弱している細胞を認めたことから裏付けら
れる。Grunert(2003)らは、癌細胞におけるEMTの形質は段階的に獲得されると
し、一部の形質のみを示す場合を“部分的なEMTの獲得”と述べている(73)。ま
た、Klymkowsky(2009)らは、癌細胞におけるEMTの獲得はその種類や固有の
微小環境の違いにより獲得する形質が異なるとし、癌におけるEMT獲得細胞を
“EMT-like phenotype”と称している(74)。これらの報告からも、SQUU-B細胞や
- 49 -
ΔNp63をノックダウンしたHSC-2細胞でE-cadherinの発現が認められたことは、
完全な(complete)EMTの獲得ではなく、部分的な(partial)EMTの獲得であっ
た可能性が示された(図19)。
第2点は、ΔNp63ノックダウンによりアポトーシスが亢進した点である。ΔNp63
はTAp63やp53に対し、ドミナントネガティブに作用し、TAp63やp53の標的遺
伝子の発現を抑制することで抗アポトーシスに働く(40-47, 75)。ゆえに、本研究 で得られた結果のように、ΔNp63ノックダウンによりアポトーシスが誘導された 細胞が増加することは想像に難くない。しかしながら、前述したとおり、complete EMTの形質を獲得した細胞ではアポトーシスに抵抗性を示すとされている(76)。 これらの相反する結果は、complete EMTにおけるアポトーシス抵抗性が、ΔNp63 とは異なる因子により制御されている可能性を示している。
ΔNp63の発現低下がEMTを獲得した細胞に与える他の影響として、細胞増殖活 性の低下がある。Vega(2004)らはSnailの発現がサイクリン依存性キナーゼ
(cyclin-dependent kinase: CDK)インヒビターであるp21cip1を誘導し、細胞周期の G1からS期への移行が阻害され、細胞増殖が低下するとしている(60)。ΔNp63 に関しても、その発現低下がp21cip1、p27kip1、およびp57kip2の発現を亢進し細胞増 殖が低下するとされている(77-79)。今回の研究においても、ΔNp63ノックダウ ンにより細胞増殖の抑制が認められたことから、細胞増殖活性の低下は、EMT獲 得細胞の形質の1つである可能性が示されたと同時に、ΔNp63がOSCC細胞の増 殖に関与していることも示唆された。
ΔNp63の発現低下がEMTを獲得した細胞に与える影響として、cytokeratinの発 現低下も認められた。Yang(1999)らは、p63ノックアウトマウスで表皮形成異 常が生じたことから、上皮の分化にΔNp63が重要であることを示した(80)。Koster
(2004)らもΔNp63が基底層に発現し、有棘層や顆粒層に分化した際に発現が消
- 50 -
失することを示している(81)。また、ΔNp63はCK5の5’末端に位置し、エンハ ンサーとして機能するDNase I Hypersensitive siteやCK14のプロモーターおよびエ ンハンサー領域に結合することによって、これらの発現を誘導していることが示 されている(82-84)。本研究においても、ΔNp63の発現とCK5あるいはCK14の 発現の間に相関性が認められた。ことから、ΔNp63はCK5やCK14の発現を直接 的に制御することで、重層扁平上皮細胞としての形質維持に役割を担っていると 考えられた。
研究1でのin vitroの結果をもとに、研究2ではin vivoにおいてEMTが誘導さ れた癌細胞を同定することが可能かどうかについて免疫組織化学的に検索を行っ た。その結果、OSCCの浸潤先端部において、vimentin発現亢進とΔNp63および
E-cadherinの発現が減弱した癌細胞を認め、これらはpartial EMTを獲得している
と考えられた。しかし、in vivoにおいて完全に間葉系細胞に形質転換した癌細胞 は病理組織学的に特定が難しく、その特異的な分子マーカーもないことから、
complete EMTを獲得した細胞の存在を今回の研究では確認出来なかった。臨床病
理組織学的所見との関連についての検討では、浸潤先端部においてpartial EMTを 生じた細胞の存在が転移ならびに予後と強く関連していることが示された。また、
本研究で使用したOSCC細胞株のうちMorifuji(2000)らが樹立したSQUU-B細 胞は、ヌードマウスの舌へSQUU-B細胞を同所性移植した結果、86.7%の頸部リン パ節転移を認め、他の細胞株と比較し極めて高い転移能を有することが分かって
いる(54-57)。これらの結果を考慮すると、ΔNp63発現減弱および消失がEMTを
誘導し、OSCCの転移に関与していることが明らかとなった。
近年、Mani(2008)らは不死化した乳腺上皮細胞にEMTを誘導した結果、これ らの細胞が幹細胞のマーカーであるCD44high/CD24lowの発現様式を示し、in vitroに おいて幹細胞の特徴であるmammosphereを形成したことを報告している(85)。
- 51 -
また、Santisteban(2009)らは、乳癌細胞にEMTを誘導させた細胞は
CD44high/CD24low/-を示すことを報告している(86)。これらの結果は、EMTが誘導 された細胞と組織幹細胞が極めて類似した細胞形質を持つことを意味し、さらに は癌細胞において極めて少数存在すると言われている癌幹細胞とも関連している ことを示唆している。現在、様々な癌で癌幹細胞が同定されつつあるが、OSCC においてはその存在が示唆されているものの、同定には至っていない(87-93)。今 後は、OSCCの浸潤・転移の分子機構におけるΔNp63を介したEMTの関与につ いてさらに解明を進めるとともに、OSCCにおける癌幹細胞の同定についても検 索を行う予定である。
図19 浸潤・転移におけるΔNp63を介したEMTの関与
!17 !Np63"#$%EMT&E-cadherin"#$%EMT!
'
"Np63 (! E-cadherin (!
)*+,-.(!
)/012343(! )*+56(!
),7123438! )*+9:;8!
<=)>?!
@ABC!
complete EMT !
partial EMT ! partial EMT !
? !
- 52 -
- 53 -
謝 辞
稿を終えるにあたり、このような研究の機会を与えて頂きましたとともに終始御 懇篤なる御指導、御校閲を賜りました九州大学大学院歯学研究院口腔顎顔面病態学 講座顎顔面腫瘍制御学分野中村誠司教授に深甚なる謝意を表します。さらに、直接 御指導、御校閲を頂きました九州大学大学院歯学研究院口腔顎顔面病態学講座顎顔 面腫瘍制御学分野川野真太郎助教に謹んで感謝の意を表します。また、常に研究の 協力ならびに励ましの言葉を頂きました九州大学大学院歯学研究院口腔顎顔面病 態学講座顎顔面腫瘍制御学分野の教室員の皆様に深く感謝いたします。最後に研究 を蔭ながら支えてくれた家族に感謝いたします。
- 54 -
参考文献
1. Capote-Moreno A., Naval L., Munoz-Guerra M. F., Sastre J., and
Rodriguez-Campo F. J., Prognostic factors influencing contralateral neck lymph node metastases in oral and oropharyngeal carcinoma. J Oral Maxillofac Surg, 2010. 68 (2): 268-75.
2. Sano D. and Myers J. N., Metastasis of squamous cell carcinoma of the oral tongue. Cancer Metastasis Rev, 2007. 26 (3-4): 645-62.
3. Kowalski L. P., Bagietto R., Lara J. R., Santos R. L., Silva J. F., Jr., and Magrin J., Prognostic significance of the distribution of neck node metastasis from oral carcinoma. Head Neck, 2000. 22 (3): 207-14.
4. Statistics and Information Dept. M. s. S., Ministry of Health, Labour and Welfare, Vital Statistics of Japan.
5. Goerres G. W., Schmid D. T., Gratz K. W., von Schulthess G. K., and Eyrich G.
K., Impact of whole body positron emission tomography on initial staging and therapy in patients with squamous cell carcinoma of the oral cavity. Oral Oncol, 2003. 39 (6): 547-51.
6. Teknos T. N., Rosenthal E. L., Lee D., Taylor R., and Marn C. S., Positron emission tomography in the evaluation of stage III and IV head and neck cancer.
Head Neck, 2001. 23 (12): 1056-60.
7. Araki Y., Okada Y., Izumi M., and Katagiri M., An Analysis of Distant
Metastases in Oral Squamous Cell Carcinoma. J Hard Tissue Biol, 2010. 19 (1):
27-32.
8. Matsuda T., Marugame T., Kamo K., Katanoda K., Ajiki W., and Sobue T.,
- 55 -
Cancer incidence and incidence rates in Japan in 2005: based on data from 12 population-based cancer registries in the Monitoring of Cancer Incidence in Japan (MCIJ) project. Jpn J Clin Oncol, 2011. 41 (1): 139-47.
9. Paget S., The distribution of secondary growths in cancer of the breast. 1889.
Cancer Metastasis Rev, 1989. 8 (2): 98-101.
10. Ewing J., Neoplastic disease. A treatise on tumors. 1928: 1046.
11. Levin I., The mechanisms of metastasis of metastasis formation in experimental cancer. J Exp Med, 1913. 18 (4): 397-405.
12. Fidler I. J., Critical determinants of metastasis. Semin Cancer Biol, 2002. 12 (2):
89-96.
13. Fidler I. J., Selection of successive tumour lines for metastasis. Nat New Biol, 1973. 242 (118): 148-9.
14. Fidler I. J., The pathogenesis of cancer metastasis: the 'seed and soil' hypothesis revisited. Nat Rev Cancer, 2003. 3 (6): 453-8.
15. Woodhouse E. C., Chuaqui R. F., and Liotta L. A., General mechanisms of metastasis. Cancer, 1997. 80 (8 Suppl): 1529-37.
16. Liotta L. A., Rao C. N., and Barsky S. H., Tumor invasion and the extracellular matrix. Lab Invest, 1983. 49 (6): 636-49.
17. Hanahan D. and Weinberg R. A., The hallmarks of cancer. Cell, 2000. 100 (1):
57-70.
18. Stacker S. A., Achen M. G., Jussila L., Baldwin M. E., and Alitalo K.,
Lymphangiogenesis and cancer metastasis. Nat Rev Cancer, 2002. 2 (8): 573-83.
19. Hanahan D. and Folkman J., Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell, 1996. 86 (3): 353-64.
- 56 -
20. Sahai E., Illuminating the metastatic process. Nat Rev Cancer, 2007. 7 (10):
737-49.
21. Thiery J. P., Epithelial-mesenchymal transitions in tumour progression. Nat Rev Cancer, 2002. 2 (6): 442-54.
22. Potenta S., Zeisberg E., and Kalluri R., The role of endothelial-to-mesenchymal transition in cancer progression. Br J Cancer, 2008. 99 (9): 1375-9.
23. Hugo H., Ackland M. L., Blick T., Lawrence M. G., Clements J. A., Williams E.
D., and Thompson E. W., Epithelial--mesenchymal and mesenchymal--epithelial transitions in carcinoma progression. J Cell Physiol, 2007. 213 (2): 374-83.
24. Kalluri R. and Weinberg R. A., The basics of epithelial-mesenchymal transition. J Clin Invest, 2009. 119 (6): 1420-8.
25. Polyak K. and Weinberg R. A., Transitions between epithelial and mesenchymal states: acquisition of malignant and stem cell traits. Nat Rev Cancer, 2009. 9 (4):
265-73.
26. Hay E. D., An overview of epithelio-mesenchymal transformation. Acta Anat (Basel), 1995. 154 (1): 8-20.
27. Morabito C. J., Dettman R. W., Kattan J., Collier J. M., and Bristow J., Positive and negative regulation of epicardial-mesenchymal transformation during avian heart development. Dev Biol, 2001. 234 (1): 204-15.
28. Acloque H., Adams M. S., Fishwick K., Bronner-Fraser M., and Nieto M. A., Epithelial-mesenchymal transitions: the importance of changing cell state in development and disease. J Clin Invest, 2009. 119 (6): 1438-49.
29. Sharpe P. T., Neural crest and tooth morphogenesis. Adv Dent Res, 2001. 15:
4-7.
- 57 -
30. Arana-Chavez V. E. and Massa L. F., Odontoblasts: the cells forming and maintaining dentine. Int J Biochem Cell Biol, 2004. 36 (8): 1367-73.
31. Sun D., McAlmon K. R., Davies J. A., Bernfield M., and Hay E. D.,
Simultaneous loss of expression of syndecan-1 and E-cadherin in the embryonic palate during epithelial-mesenchymal transformation. Int J Dev Biol, 1998. 42 (5): 733-6.
32. Bischof P., Aplin J. D., Bentin-Ley U., Brannstrom M., Casslen B., Castrillo J. L., Classen-Linke I., Critchley H. O., Devoto L., D'Hooghe T., Horcajadas J. A., Groothuis P., Ivell R., et al., Implantation of the human embryo: research lines and models. From the implantation research network 'Fruitful'. Gynecol Obstet Invest, 2006. 62 (4): 206-16.
33. Christiansen J. J. and Rajasekaran A. K., Reassessing epithelial to mesenchymal transition as a prerequisite for carcinoma invasion and metastasis. Cancer Res, 2006. 66 (17): 8319-26.
34. Yang J. and Weinberg R. A., Epithelial-mesenchymal transition: at the crossroads of development and tumor metastasis. Dev Cell, 2008. 14 (6): 818-29.
35. Yang A., Kaghad M., Wang Y., Gillett E., Fleming M. D., Dotsch V., Andrews N.
C., Caput D., and McKeon F., p63, a p53 homolog at 3q27-29, encodes multiple products with transactivating, death-inducing, and dominant-negative activities.
Mol Cell, 1998. 2 (3): 305-16.
36. Ikawa S., Nakagawara A., and Ikawa Y., p53 family genes: structural comparison, expression and mutation. Cell Death Differ, 1999. 6 (12): 1154-61.
37. Augustin M., Bamberger C., Paul D., and Schmale H., Cloning and chromosomal mapping of the human p53-related KET gene to chromosome 3q27 and its
- 58 -
murine homolog Ket to mouse chromosome 16. Mamm Genome, 1998. 9 (11):
899-902.
38. Trink B., Okami K., Wu L., Sriuranpong V., Jen J., and Sidransky D., A new human p53 homologue. Nat Med, 1998. 4 (7): 747-8.
39. Osada M., Ohba M., Kawahara C., Ishioka C., Kanamaru R., Katoh I., Ikawa Y., Nimura Y., Nakagawara A., Obinata M., and Ikawa S., Cloning and functional analysis of human p51, which structurally and functionally resembles p53. Nat Med, 1998. 4 (7): 839-43.
40. Jost C. A., Marin M. C., and Kaelin W. G., Jr., p73 is a simian [correction of human] p53-related protein that can induce apoptosis. Nature, 1997. 389 (6647):
191-4.
41. Hagiwara K., McMenamin M. G., Miura K., and Harris C. C., Mutational analysis of the p63/p73L/p51/p40/CUSP/KET gene in human cancer cell lines using intronic primers. Cancer Res, 1999. 59 (17): 4165-9.
42. Bourdon J. C., Fernandes K., Murray-Zmijewski F., Liu G., Diot A., Xirodimas D.
P., Saville M. K., and Lane D. P., p53 isoforms can regulate p53 transcriptional activity. Genes Dev, 2005. 19 (18): 2122-37.
43. Nylander K., Coates P. J., and Hall P. A., Characterization of the expression pattern of p63 alpha and delta Np63 alpha in benign and malignant oral epithelial lesions. Int J Cancer, 2000. 87 (3): 368-72.
44. Ratovitski E. A., Patturajan M., Hibi K., Trink B., Yamaguchi K., and Sidransky D., p53 associates with and targets Delta Np63 into a protein degradation
pathway. Proc Natl Acad Sci U S A, 2001. 98 (4): 1817-22.
45. Wu G., Nomoto S., Hoque M. O., Dracheva T., Osada M., Lee C. C., Dong S. M.,
- 59 -
Guo Z., Benoit N., Cohen Y., Rechthand P., Califano J., Moon C. S., et al., DeltaNp63alpha and TAp63alpha regulate transcription of genes with distinct biological functions in cancer and development. Cancer Res, 2003. 63 (10):
2351-7.
46. Yang A. and McKeon F., P63 and P73: P53 mimics, menaces and more. Nat Rev Mol Cell Biol, 2000. 1 (3): 199-207.
47. Mills A. A., p63: oncogene or tumor suppressor? Curr Opin Genet Dev, 2006. 16 (1): 38-44.
48. Hibi K., Trink B., Patturajan M., Westra W. H., Caballero O. L., Hill D. E., Ratovitski E. A., Jen J., and Sidransky D., AIS is an oncogene amplified in squamous cell carcinoma. Proc Natl Acad Sci U S A, 2000. 97 (10): 5462-7.
49. Crook T., Nicholls J. M., Brooks L., O'Nions J., and Allday M. J., High level expression of deltaN-p63: a mechanism for the inactivation of p53 in
undifferentiated nasopharyngeal carcinoma (NPC)? Oncogene, 2000. 19 (30):
3439-44.
50. Rocco J. W. and Ellisen L. W., p63 and p73: life and death in squamous cell carcinoma. Cell Cycle, 2006. 5 (9): 936-40.
51. Matsubara R., Kawano S., Kiyosue T., Goto Y., Hirano M., Jinno T., Toyoshima T., Kitamura R., Oobu K., and Nakamura S., Increased DeltaNp63 expression is predictive of malignant transformation in oral epithelial dysplasia and poor prognosis in oral squamous cell carcinoma. Int J Oncol, 2011. 39 (6): 1391-9.
52. Barbieri C. E., Tang L. J., Brown K. A., and Pietenpol J. A., Loss of p63 leads to increased cell migration and up-regulation of genes involved in invasion and metastasis. Cancer Res, 2006. 66 (15): 7589-97.
- 60 -
53. Carroll D. K., Carroll J. S., Leong C. O., Cheng F., Brown M., Mills A. A., Brugge J. S., and Ellisen L. W., p63 regulates an adhesion programme and cell survival in epithelial cells. Nat Cell Biol, 2006. 8 (6): 551-61.
54. Momose F., Araida T., Negishi A., Ichijo H., Shioda S., and Sasaki S., Variant sublines with different metastatic potentials selected in nude mice from human oral squamous cell carcinomas. J Oral Pathol Med, 1989. 18 (7): 391-5.
55. Morifuji M., Taniguchi S., Sakai H., Nakabeppu Y., and Ohishi M., Differential expression of cytokeratin after orthotopic implantation of newly established human tongue cancer cell lines of defined metastatic ability. Am J Pathol, 2000.
156 (4): 1317-26.
56. Aida T., Irie T., and Tachikawa T., Establishment and charactalization of human oral squamous cell carcinoma cell line with highly lymph node metastatic potential. J Jpn Soc Oral Tumor, 2001. 13 (4): 301-6.
57. Takahashi K., Kanazawa, H., Akiyama, Y., Tazaki, S., Takahara, M., Muto, T., Tanzawa, H., Sato, K., Establishment and characterization of a cell line (SAS) from poorly differentiated human squamous cell carcinoma of the tongue. Jpn Stomatol Soc, 1989. 38 (1): 20-8.
58. Brown M. and Petric M., Evaluation of cell line 293 for virus isolation in routine viral diagnosis. J Clin Microbiol, 1986. 23 (4): 704-8.
59. Akobeng A. K., Understanding diagnostic tests 3: Receiver operating characteristic curves. Acta Paediatr, 2007. 96 (5): 644-7.
60. Vega S., Morales A. V., Ocana O. H., Valdes F., Fabregat I., and Nieto M. A., Snail blocks the cell cycle and confers resistance to cell death. Genes Dev, 2004.
18 (10): 1131-43.
- 61 -
61. Higashikawa K., Yoneda S., Tobiume K., Saitoh M., Taki M., Mitani Y., Shigeishi H., Ono S., and Kamata N., DeltaNp63alpha-dependent expression of Id-3 distinctively suppresses the invasiveness of human squamous cell carcinoma.
Int J Cancer, 2009. 124 (12): 2837-44.
62. Zeisberg M. and Neilson E. G., Biomarkers for epithelial-mesenchymal transitions. J Clin Invest, 2009. 119 (6): 1429-37.
63. Yoshida C. and Takeichi M., Teratocarcinoma cell adhesion: identification of a cell-surface protein involved in calcium-dependent cell aggregation. Cell, 1982.
28 (2): 217-24.
64. Takeichi M., Molecular basis for teratocarcinoma cell-cell adhesion. Dev Biol (N Y 1985), 1986. 2: 373-88.
65. Miettinen P. J., Ebner R., Lopez A. R., and Derynck R., TGF-beta induced transdifferentiation of mammary epithelial cells to mesenchymal cells:
involvement of type I receptors. J Cell Biol, 1994. 127 (6 Pt 2): 2021-36.
66. Nieto M. A., The snail superfamily of zinc-finger transcription factors. Nat Rev Mol Cell Biol, 2002. 3 (3): 155-66.
67. Cano A., Perez-Moreno M. A., Rodrigo I., Locascio A., Blanco M. J., del Barrio M. G., Portillo F., and Nieto M. A., The transcription factor snail controls
epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat Cell Biol, 2000. 2 (2): 76-83.
68. Medici D., Hay E. D., and Olsen B. R., Snail and Slug promote
epithelial-mesenchymal transition through beta-catenin-T-cell factor-4-dependent expression of transforming growth factor-beta3. Mol Biol Cell, 2008. 19 (11):
4875-87.
- 62 -
69. Kokudo T., Suzuki Y., Yoshimatsu Y., Yamazaki T., Watabe T., and Miyazono K., Snail is required for TGFbeta-induced endothelial-mesenchymal transition of embryonic stem cell-derived endothelial cells. J Cell Sci, 2008. 121 (Pt 20):
3317-24.
70. Herfs M., Hubert P., Suarez-Carmona M., Reschner A., Saussez S., Berx G., Savagner P., Boniver J., and Delvenne P., Regulation of p63 isoforms by snail and slug transcription factors in human squamous cell carcinoma. Am J Pathol, 2010. 176 (4): 1941-9.
71. Takkunen M., Grenman R., Hukkanen M., Korhonen M., Garcia de Herreros A., and Virtanen I., Snail-dependent and -independent epithelial-mesenchymal transition in oral squamous carcinoma cells. J Histochem Cytochem, 2006. 54 (11): 1263-75.
72. Higashikawa K., Yoneda S., Tobiume K., Taki M., Shigeishi H., and Kamata N., Snail-induced down-regulation of DeltaNp63alpha acquires invasive phenotype of human squamous cell carcinoma. Cancer Res, 2007. 67 (19): 9207-13.
73. Grunert S., Jechlinger M., and Beug H., Diverse cellular and molecular
mechanisms contribute to epithelial plasticity and metastasis. Nat Rev Mol Cell Biol, 2003. 4 (8): 657-65.
74. Klymkowsky M. W. and Savagner P., Epithelial-mesenchymal transition: a cancer researcher's conceptual friend and foe. Am J Pathol, 2009. 174 (5):
1588-93.
75. Sun Q., Ming L., Thomas S. M., Wang Y., Chen Z. G., Ferris R. L., Grandis J. R., Zhang L., and Yu J., PUMA mediates EGFR tyrosine kinase inhibitor-induced apoptosis in head and neck cancer cells. Oncogene, 2009. 28 (24): 2348-57.
- 63 -
76. Franco D. L., Mainez J., Vega S., Sancho P., Murillo M. M., de Frutos C. A., Del Castillo G., Lopez-Blau C., Fabregat I., and Nieto M. A., Snail1 suppresses TGF-beta-induced apoptosis and is sufficient to trigger EMT in hepatocytes. J Cell Sci, 2010. 123 (Pt 20): 3467-77.
77. Chiang C. T., Chu W. K., Chow S. E., and Chen J. K., Overexpression of Delta Np63 in a human nasopharyngeal carcinoma cell line downregulates CKIs and enhances cell proliferation. J Cell Physiol, 2009. 219 (1): 117-22.
78. Westfall M. D., Mays D. J., Sniezek J. C., and Pietenpol J. A., The Delta Np63 alpha phosphoprotein binds the p21 and 14-3-3 sigma promoters in vivo and has transcriptional repressor activity that is reduced by Hay-Wells syndrome-derived mutations. Mol Cell Biol, 2003. 23 (7): 2264-76.
79. Beretta C., Chiarelli A., Testoni B., Mantovani R., and Guerrini L., Regulation of the cyclin-dependent kinase inhibitor p57Kip2 expression by p63. Cell Cycle, 2005. 4 (11): 1625-31.
80. Yang A., Schweitzer R., Sun D., Kaghad M., Walker N., Bronson R. T., Tabin C., Sharpe A., Caput D., Crum C., and McKeon F., p63 is essential for regenerative proliferation in limb, craniofacial and epithelial development. Nature, 1999. 398 (6729): 714-8.
81. Koster M. I. and Roop D. R., The role of p63 in development and differentiation of the epidermis. J Dermatol Sci, 2004. 34 (1): 3-9.
82. Romano R. A., Birkaya B., and Sinha S., A functional enhancer of keratin14 is a direct transcriptional target of deltaNp63. J Invest Dermatol, 2007. 127 (5):
1175-86.
83. Romano R. A., Ortt K., Birkaya B., Smalley K., and Sinha S., An active role of
- 64 -
the DeltaN isoform of p63 in regulating basal keratin genes K5 and K14 and directing epidermal cell fate. PLoS One, 2009. 4 (5): e5623.
84. Candi E., Rufini A., Terrinoni A., Dinsdale D., Ranalli M., Paradisi A., De Laurenzi V., Spagnoli L. G., Catani M. V., Ramadan S., Knight R. A., and
Melino G., Differential roles of p63 isoforms in epidermal development: selective genetic complementation in p63 null mice. Cell Death Differ, 2006. 13 (6):
1037-47.
85. Mani S. A., Guo W., Liao M. J., Eaton E. N., Ayyanan A., Zhou A. Y., Brooks M., Reinhard F., Zhang C. C., Shipitsin M., Campbell L. L., Polyak K., Brisken C., et al., The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell, 2008. 133 (4): 704-15.
86. Santisteban M., Reiman J. M., Asiedu M. K., Behrens M. D., Nassar A., Kalli K.
R., Haluska P., Ingle J. N., Hartmann L. C., Manjili M. H., Radisky D. C., Ferrone S., and Knutson K. L., Immune-induced epithelial to mesenchymal transition in vivo generates breast cancer stem cells. Cancer Res, 2009. 69 (7):
2887-95.
87. Yu F., Yao H., Zhu P., Zhang X., Pan Q., Gong C., Huang Y., Hu X., Su F., Lieberman J., and Song E., let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell, 2007. 131 (6): 1109-23.
88. O'Brien C. A., Pollett A., Gallinger S., and Dick J. E., A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature, 2007. 445 (7123): 106-10.
89. Dalerba P., Dylla S. J., Park I. K., Liu R., Wang X., Cho R. W., Hoey T., Gurney A., Huang E. H., Simeone D. M., Shelton A. A., Parmiani G., Castelli C., et al.,
- 65 -
Phenotypic characterization of human colorectal cancer stem cells. Proc Natl Acad Sci U S A, 2007. 104 (24): 10158-63.
90. Suetsugu A., Nagaki M., Aoki H., Motohashi T., Kunisada T., and Moriwaki H., Characterization of CD133+ hepatocellular carcinoma cells as cancer
stem/progenitor cells. Biochem Biophys Res Commun, 2006. 351 (4): 820-4.
91. Singh S. K., Clarke I. D., Terasaki M., Bonn V. E., Hawkins C., Squire J., and Dirks P. B., Identification of a cancer stem cell in human brain tumors. Cancer Res, 2003. 63 (18): 5821-8.
92. Singh S. K., Hawkins C., Clarke I. D., Squire J. A., Bayani J., Hide T., Henkelman R. M., Cusimano M. D., and Dirks P. B., Identification of human brain tumour initiating cells. Nature, 2004. 432 (7015): 396-401.
93. Kiyosue T., Kawano S., Matsubara R., Goto Y., Hirano M., Jinno T., Toyoshima T., Kitamura R., Oobu K., and Nakamura S., Immunohistochemical localization of p75 neurotrophin receptor (p75NTR) in oral leukoplakia and oral squamous cell carcinoma. Int J Clin Oncol, 2011 (in press).