九州大学学術情報リポジトリ
Kyushu University Institutional Repository
アレルギー関連因子の評価法の開発とその利用に関 する研究
高杉, 美佳子
九州大学農学研究科食糧化学工学専攻
https://doi.org/10.11501/3135077
出版情報:Kyushu University, 1997, 博士(農学), 課程博士 バージョン:
権利関係:
第6章 食品のアレルゲン活性評価法の検討
第1節 緒 日
食物アレルギーは小児だけでなく成人にも重要な問題となっている。 食物ア レルギーは食品中のアレルゲンと呼ばれるタンパク質によって引き起こされる。
アレルゲンは加工食品中にも合まれている場合があるため、 食品中のアレルゲ ンの検出はアレルギ一治療法の一つであるアレルゲン除去のための重要な情報 となる。 また、 低アレルゲン食品や代替食品の開発を行う場合において、 m vÍtroにおけるアレルゲン活性の測定は臨床試験を行う前の予備試験としても重 要である。 In vÍtroでの食物アレルゲンの検出には、 サンドイツチELISA (Levieux and Venien, 1994; Lovegrove et a1., 1993; Makinen-Kijunen and Palosuo,
1992)、 inhibition ELISA (Kilshaw et a1., 1982)、 radioimmunoassay (Paganelli and Levinsky, 1980 ) などの方法が用いられている。
卵、 大豆および牛乳はアレルギーを引き起こす主要食品と言われており、 卵 ではオボアルブ、ミン(OVA)、 オボムコイド、 オボトランスフェリン、 リゾチー ムなどが(Ebbehoj et a1., 1995)、 大豆ではG1ymBd 30K などが(Ogawa et a1.,
1991) アレルゲンタンパクとして同定されている。 食物アレルギーの発症を予防 する最も確実な方法はアレルゲンを合む食品の摂取を避けることであり、 食品 中のOVAあるいは大豆タンパクの合量を知ることは卵あるいは大豆アレルギー の予防に重要である。
そこで、 低アレルゲン食品あるいは代替食品の開発のための基礎情報の獲得 および抗原活性評価法の開発を目的として、 分離大豆タンパク質をペプシンお よび微生物由来プロテアーゼで分解して得られる非消化性画分、 UDFの抗原活 性をELISAおよびinhibition ELISAを用いて検討した。 また、 大豆あるいは卵
を使用したマヨネーズ製品のOVA合量についても比較検討した。
第2節 材料と方法
第1項 一般試薬
分離大豆タンパク質(ニューフジ プ口氏Soy)、 大豆タンパク質非消化性画分 (UDF)およびサボニン除去処理をほどこしたUDF(UDF-T)は不二製油( 大阪,
日本)より提供された。
ELISA用として、 Soy、 UDFおよびUDF-Tを50mM炭酸緩衝液(pH 9.6)に 1 mg/mlとなるように懸濁し、 12,500 x gで20分間遠心分離した上清を抗原溶液 として調製した。 Inhibition ELISA用には、 Soy、 UDFおよびUDF-Tを生理リ ン酸緩衝液(PBS; pH 7.4 )に 10mg/mlとなるように懸濁し、 12,500 x gで20分間 遠心分離した上清を用いた。 卵黄使用マヨネーズ(EY-M )および、全卵使用マヨ 不一ズズ、(但E
(東京'日本)より提供されたものを用いたo これらのマヨネーズ製品はPBS で
10倍に希釈して 12,500 x gで20分間遠心分離後、 中間層をパスツールピペット で回収し、 0.22μm フィル
タ
ーでろ過したものをサンプルとして実験に使用した。OVAはSigma社(St.lρuis, MO)より購入し、 50mM炭酸緩衝液で 1mg/mlと なるように調製したものをinhibition ELISAの抗原溶液として用いた。
1ρwry法のために、 炭酸ナトリウム2g 、 2N水酸化ナトリウム、 4%酒石酸ナ トリウム0.5mlおよび2%硫酸銅・五水和物0.5mlを混合し、 液量を50mlにし た溶液(試薬A)を調製した。 また、 タンパク標準液として、 ウシ血清アルブミ ンを200mg/mlの濃度で0.4 N水酸化ナトリウム に溶解したものを使用した。
ヒト大豆抗血清として、 以前に大豆アレルギーと診断された本学科の学生の 血清を使用した。 インフォームドコンセントを得た後、 真空採血管を用いて血 液を採取し、 室温に30分間放置した後に1,500 x gで15分間遠心分離して血清を 調製した。
ELISAおよびinhibition ELISAには、 西洋ワサビベルオキシダーゼ(HRP)標 識抗ヒトIg G およびHRP標識アビジンはDako社(Glostrup, Denmark )より、
ビオチン標識抗ヒトIgEはKPL社(Gaithersburg , MD )より、 HRP標識抗ラツ
トI gGは C app el社(West Chest er, PA)より購入したものを用いた。
ポリアクリルアミドゲル電気泳動(PA GE)用として 、 アクリルアミドモノ
マ一、 トリス(ヒドロキシメチル)アミノメタンおよびグリセロールをナカライ テスク(京都フ日本)より、 N,N'-メチレンビスーアクリルアミド 、 ドデ、シル硫酸ナ
トリウム(SDS)、 ブロモフェノールブツレーおよび2-メルカプトエタノールを和 光純薬(大阪,日本)より、 過硫酸アンモニウム(APS)、 N,N, N', N'-テトラメチ ルエチレンジアミン(TEMED)および分子量マーカーであるSDS-PAGE
standar d s low range をBio Rad 社(H ercules, C A)より、 クマシーブリリアントブ ルー(CBB )を Sigma社より購入した。 また、 電気泳動装置としてミニプロテイ アンII 1-D、 電源としてモデ、ル200 /2.0 パワーサプライをBio Rad 社より購入し て使用した。
第2項 ラットOVA抗血清の調製
4週齢のB row nNo rway系雄ラットをセアック吉富 (吉富,日本)より購入し、
飼育中は カゼイン20%を合むOVA無添加食を与えた。 oおよび21日目に3%
A1 (OH)3 -0.15 M NaCIに懸濁したOVA を50μg/ratの用量で腹腔内に投与した。
24日自にジエチルエーテルで麻酔後、 腹部大動脈より採血を行った。 採取した 血液は3,0∞xgで15分間遠心するこ とにより血清を調製した。
第3項 タンパクの定量
それぞれの試料溶液と10%トリクロロ酢酸を混合し、 650xgで5分間遠心分 離を行った。 エタノール・エーテル・酢酸混液(74 : 24 : 1 ) 1 mlに沈澱物を再懸 濁後、 650xgで15分間遠心分離した。 沈澱物に0.8NNaOH 溶液および超純水 を1 mlずつ加えて溶解した後、 この溶液0.5 mlを試薬 A1 mlと 混合して室温に 10分間放置した。 さらにフェノール試薬0.2 mlを添加して暗所に30分間放置し
た後、 750 n mの吸光度を測定し、 タンパク標準液よりタンパク質濃度を計算し た。
62
第4項 ELISA法による抗原特異的IgEおよび:lgGの検出
Immuno-p late Maxi sorp C96はNunc社(Roskilide,Denmark)より購入した。
洗浄液はTween 20をナカライテスクより購入し、 0.05% Tween 20合有生理リン 酸緩衝液(TPBS)を調製して使用した。 魚ゼラチンは Sigma社(St. 1ρuis, MO)
より購入し、 TPBSで2%になるように希釈してブロッキング溶液として用いる とともにHRP標識抗体、 サン プルおよび標準抗体の希釈液として使用した。 発 色基質溶液は、 0.0069るHの2合有0.2Mク エン酸緩衝液(pH 4.0)、 超純水および
6 mg/ml 2, 2'-アジノービス(3-エチルベン ゾチアゾリンー6-スルホン酸 ) ジアンモニ ウム塩(ABTS)溶液を10: 9: 1で混合したも のを用い、 発色反応 停止液には 1.59るシュウ酸溶液を用いた。
IgGは通常の ELISA法により、 IgEはアビジンービオチン法により 測定した。
抗原溶液を50 mM炭酸緩衝液で30'"'-'38倍に段階希釈し、 これらの溶液を150μl ずつ 96 well ELISA プレートに添加し、 370Cで1時間保温して抗原をプレート
に固定した。 ブロッキング溶液を300μiずつ添加し、 40Cで一晩ブロッキング した後、 IgG測定の場合は ブロッキング溶液で100倍に希釈したヒト抗血清を 50μIずつ添加し、 370Cで1時間反応させた。 ブロッキング溶液で1,000倍希釈 したHRP標識抗ヒトIgG抗体を100μlずつ添加して1時間反応させた後に発色 基質溶液を100μiずつ添加して370Cで15分間反応させた。 1.59るシュウ酸溶液 を100μl添加して反応を停止させ、 マイクロプレート分光光度計で 415 nm の 吸 光度を測定した。 各反応過程の聞は TPBSで3回ずつ洗浄 した。
IgE 測定の場合、 ブロッキング溶液で10倍に希釈したヒト抗血清を50μiずつ 添加して 40Cで1時間反応させた後、 ブロッキング溶液で1,000倍希釈したど オチン標識抗 ヒトIgEを100μlずつ添加して40Cで1時間反応させた。 つぎに 、 ブロッキング溶液で5,000倍に希釈したHRP標識 アビジンを100μlずつ添加し て40Cで30分間保温した後、 発色反応を行った。 抗血清添加以後の洗浄は 4回 ずつ行った。 IgGに対する抗原活性は吸光度の最大値の 1/2となる希釈率を50%
inhibiting dilution (IDso)として表し、 IDsoを用いて比較した。
第5項 Inhibition ELISA
プレート、 洗浄液、 ブロッキング溶液、 HRP標識抗体 希釈液、 発色基質溶液 および発色反応停止液は ELISAと同じものを用いた。
PBSで希釈したサンプル溶液とPBSで10倍 に希釈した抗血清を1:1に混合 し、 40Cで一晩放置した。 50 mM炭酸緩衝液に溶解した1 mg/ml Soyまたは OVA抗原溶液を 96 well ELISA プレートに100μlずつ添加して370Cで 1時間保 温して抗原をプレートに固定した後、 ブロッキング溶液を300μlずつ添加して 40Cで一夜ブロッキングを行った。 つぎ、に、 サンプル溶液ー抗血清 混合液を100μ
iずつ添加し、 370Cで1時間反応させ、 ブロッキング溶液で1,000倍に希釈した HRP標識抗ラットIgG抗体を100μlずつ添加してさらに1時間反応させた後に 発色反応を行った。 各反応過程の聞はTPBSで3回ずつ洗浄した。
第6項 SDS-PAGE
サンプル緩衝液は超純水、 0.5Mト リスー塩酸緩衝液(pH6.8)、 50 9る グリセロー ル、 1 %ブロモフェノールブルー、 2-メルカプトエタノールを20: 5 : 8: 1 : 2に 混合して調製した。 泳動 用緩衝液は、 ト リス(ヒドロ キシメチル )アミノメタン
6 mg/ml、 グリシン28.8 mg/mlおよび SDS1mg/mlとなるように超純水に溶解し た。
SDS-PAGE はLaemmli(1970)の方法に基づき、 4%濃縮ゲルおよび149る分離 ゲルを用いて 泳動を行った。 サンプル はサンプル緩衝液と容量比1:1で 混合し て1000Cで 5分間加熱し、 1ウエルあたり10μlずつ添加して200Vで30分間泳 動し、 泳動終了後、 0.04%CBBを用いて室温で1時間染色した。
第3節 結 果
第1項 大豆タンパクの抗原性に及ぼす食品加工の影響
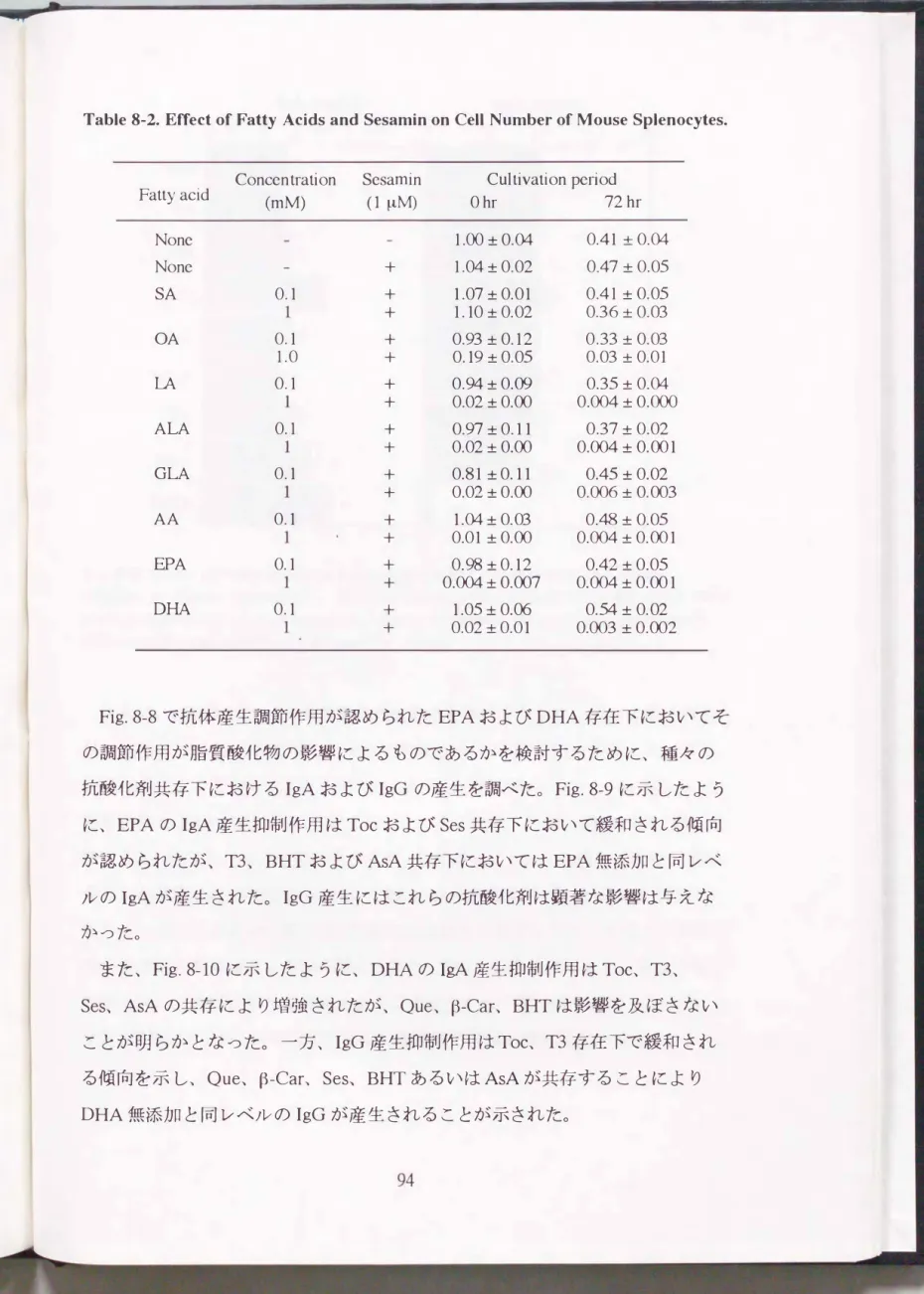
Table 6-1に大豆タンパク質加工物可溶性タンパク濃度を定量した結果を示し た。 Soyに比べてUDFおよびUDF-Tは可 溶性タンパク量が1/5� 1/6に減少し
64
Table 6・1. Determination of Proteins Concentration in Soy and UDF Dissolved in 50 mM Sodium Carbonate-Sodium Bicarbonate Buffer by Lowry's Method.
Concentration (μg/ml)
320 52 63 Protein
Soy UDF UDF-T
0.4 、3
白
0.2
O.O�
10 0.8
0.4 0.6
0.2
0.0
0.6
(出マ吋)恒国£ロ。ω同凶同
(23刊)苫£口。ω。2
10 4
ミJハU1i
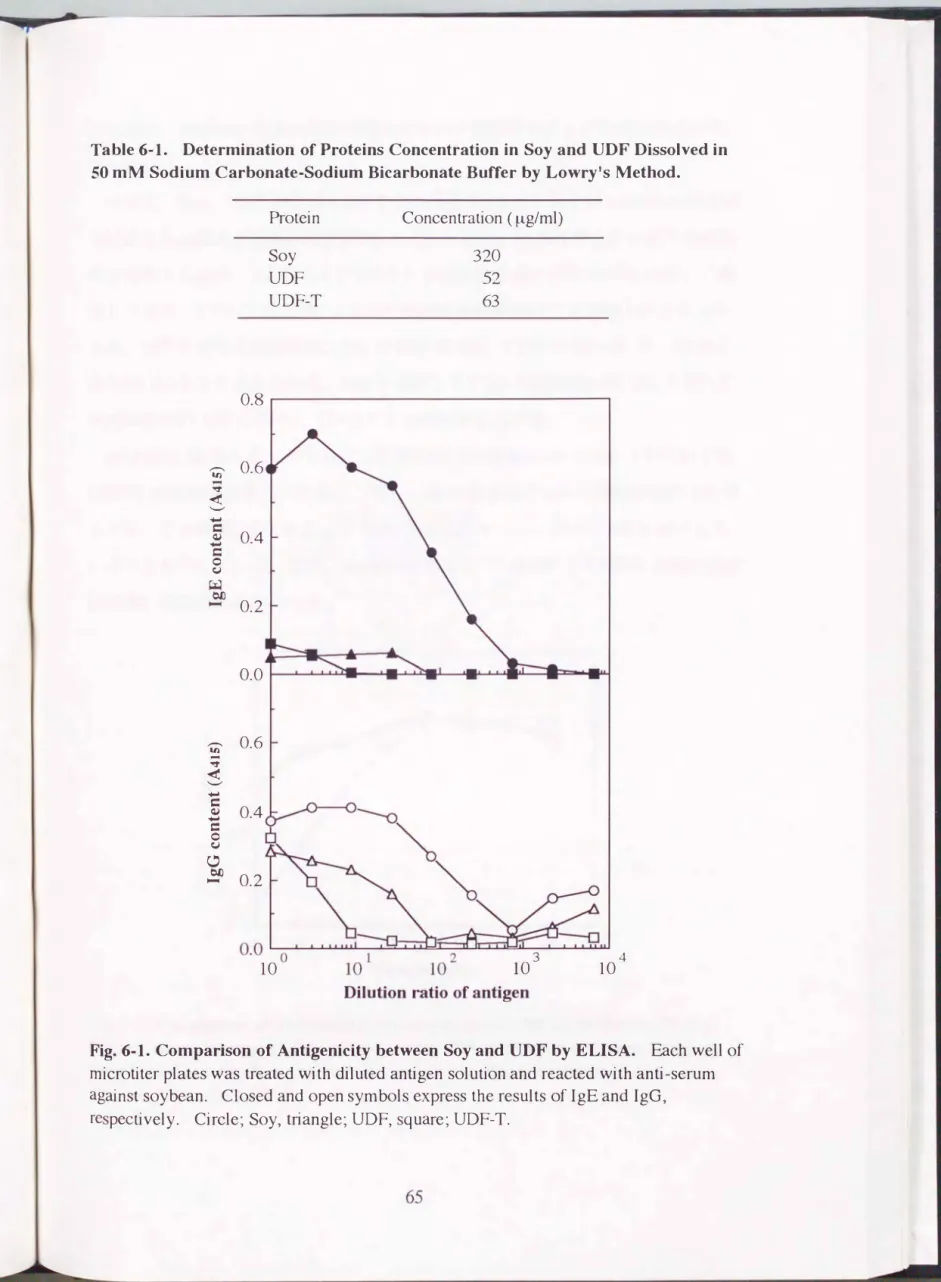
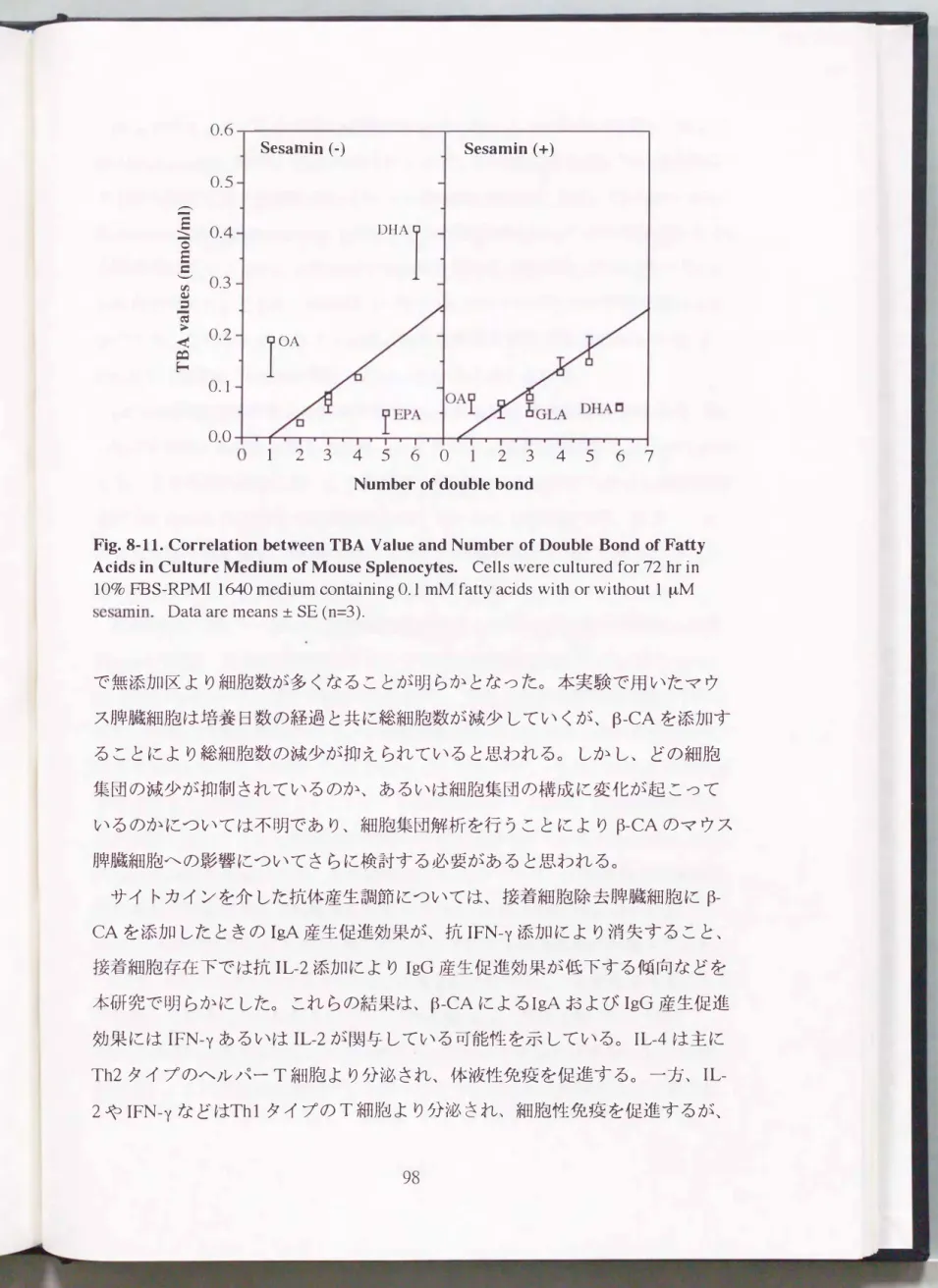
Fig.6・1. Comparison of Antigenicity between Soy and UDF by ELISA. Each well of mícrotiter plates was treated with diluted antigen solution and reacted with anti-serum against soybean. Closed and open symbols express the results of IgE and IgG,
respectively. Circle� Soy, triangle� UDF, square� UDF-T.
ているが、 サボニン除去処理は可溶性タンパク濃度にほとんど影響を及ぼさな かった。
つぎに、 Soy、 UDFおよび UDF-Tの抗原性を ELISAおよび inhibition ELISA で比較した結果をFig.6-1 およびFig.6-2に示した。 ELISAでIgEに対する反応
性を比較した結果、 UDFおよびUDF-Tの抗原性はSoyの約1/1000に著しく低 下しており、 UDF-Tは UDFよりわずかに抗原性が低いことが明らかとなった。
また、 IgGに対する抗原性は、 SoyのID50は 110、 UDFのID50は 30、 UDF-T のID50は3.5であることから、 Soyと比較してUDFの抗原性は約1β、 UDF-T の抗原性は約1β0に低下していることが明らかとなった。
Inhibition ELISAによるIgGに対する抗原性の比較においては、 UDFおよび UDF-Tの試料原液を用いた場合と等しいA415値を示すSoyの希釈率は約120で あった。 この結果はUDFおよびUDF-Tの抗原性がSoyの約1/120に低下して いることを示している。 また、 inhibition ELISAではUDFと UDF-Tの聞に抗原 性の違いは認められなかった。
1.5
1.0
同同マ吋
0.5
O.O�
10 10 10 2 10 3 10 4 10 5 10 6 10 7 10 8 Dilution ratio
Fig.6・2. Comparison of Antigenicity between Soy and UDF by Inhibition ELISA.
Original sample solution (10うwere diluted with PBS to 107 times and reacted with 10 times diluted anti-serum against soybean, and then applied to inhibition ELISA. (0);
Soy, (ð); UDF, (ロ);UDF-T.
Table 6・2. Determination of Proteins in Mayonnaise Products Dissolved in PBS by Lowry's恥lethod.
Protein
EY-M E-M Soy-M
MW kDa
104.0 ... 、h 82.0 ..
48.3 ..
33.4 ..
28.3 ..
19.4 ..
1
Concentration (μg/ml)
190 290 not detected
2 3 4
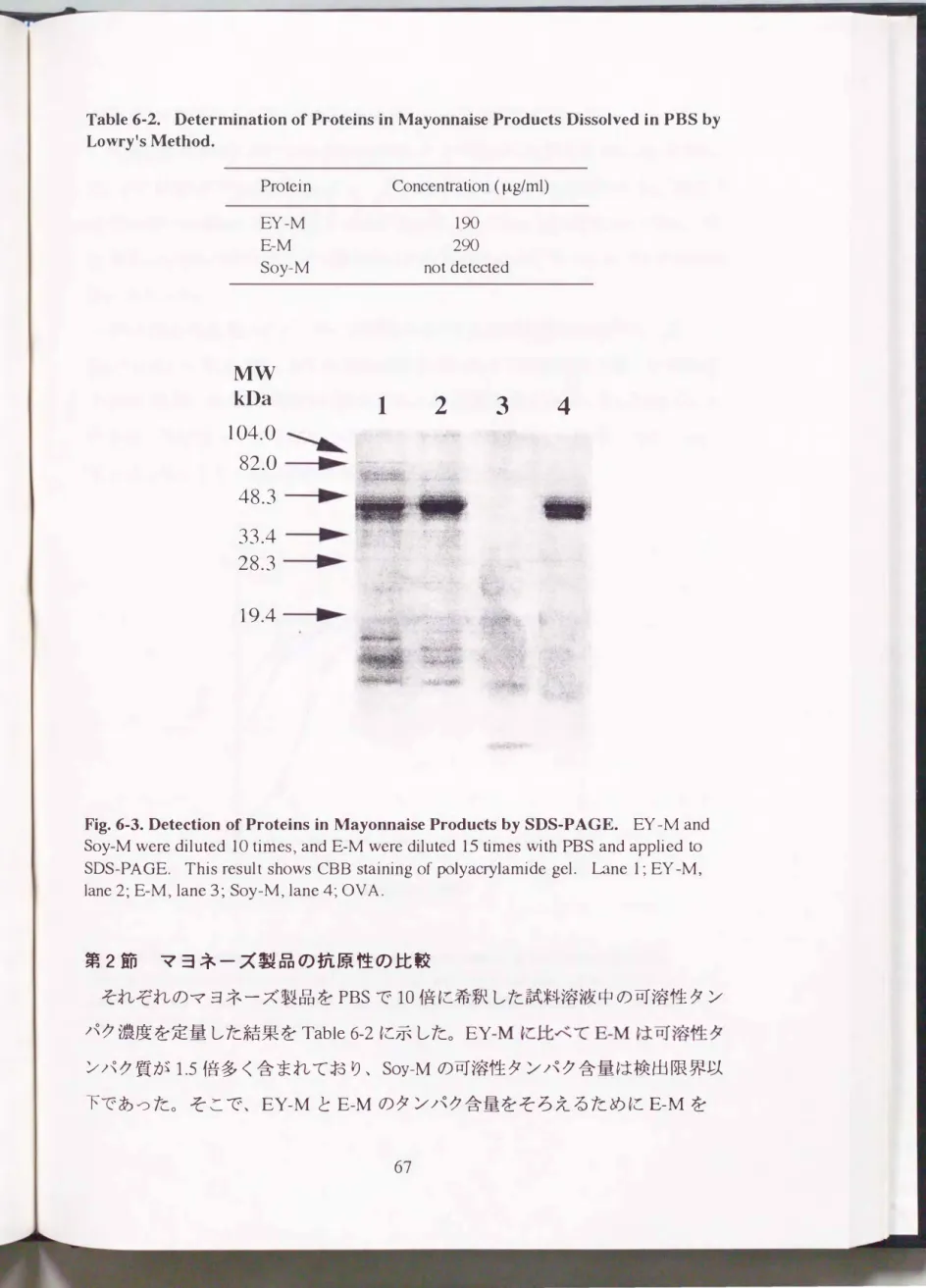
Fig.6・3. Detection of Proteins in Mayonnaise Products by SDS-PAGE. EY -M and Soy-M were diluted 10 times, and E-M were diluted 15 times with PBS and applied ω SDS-PAGE. This result shows CBB staining of polyacrylamide gel. Lane 1; EY -M,
lane 2; E-M, lane 3; Soy-恥土lane4;OVA.
第2節 マヨネーズ製品の抗原性の比較
それぞれのマヨネーズ製品をPBSで10倍に希釈した試料溶液中の可溶性タン パク濃度を定量した結果をTable 6-2に示した。 EY
ンパク質が1.5倍多く合まれており、 Soy-Mの可溶性タンパク合量は検出限界以 下であった。 そこで、 EY-MとE-Mのタンパク合量をそろえるためにE-Mを
67
PBSで1.5倍希釈したものを試料溶液として以後の実験に用いた。
試料溶液中のタンパク質をSDS-PAGEにより検出した結果をFig.6-3に示し
た。 EY-MおよびE-Mにおいては、 分子量45 kDaのOVAの濃いバンドおよび 15 kDa以下の薄いバンドが各3本検出された。 しかし、 Soy-Mにおいては、 分
子量約2.5 kDaの薄いバンドが検出された以外にははっきりとしたバンドは検出 されなかった。
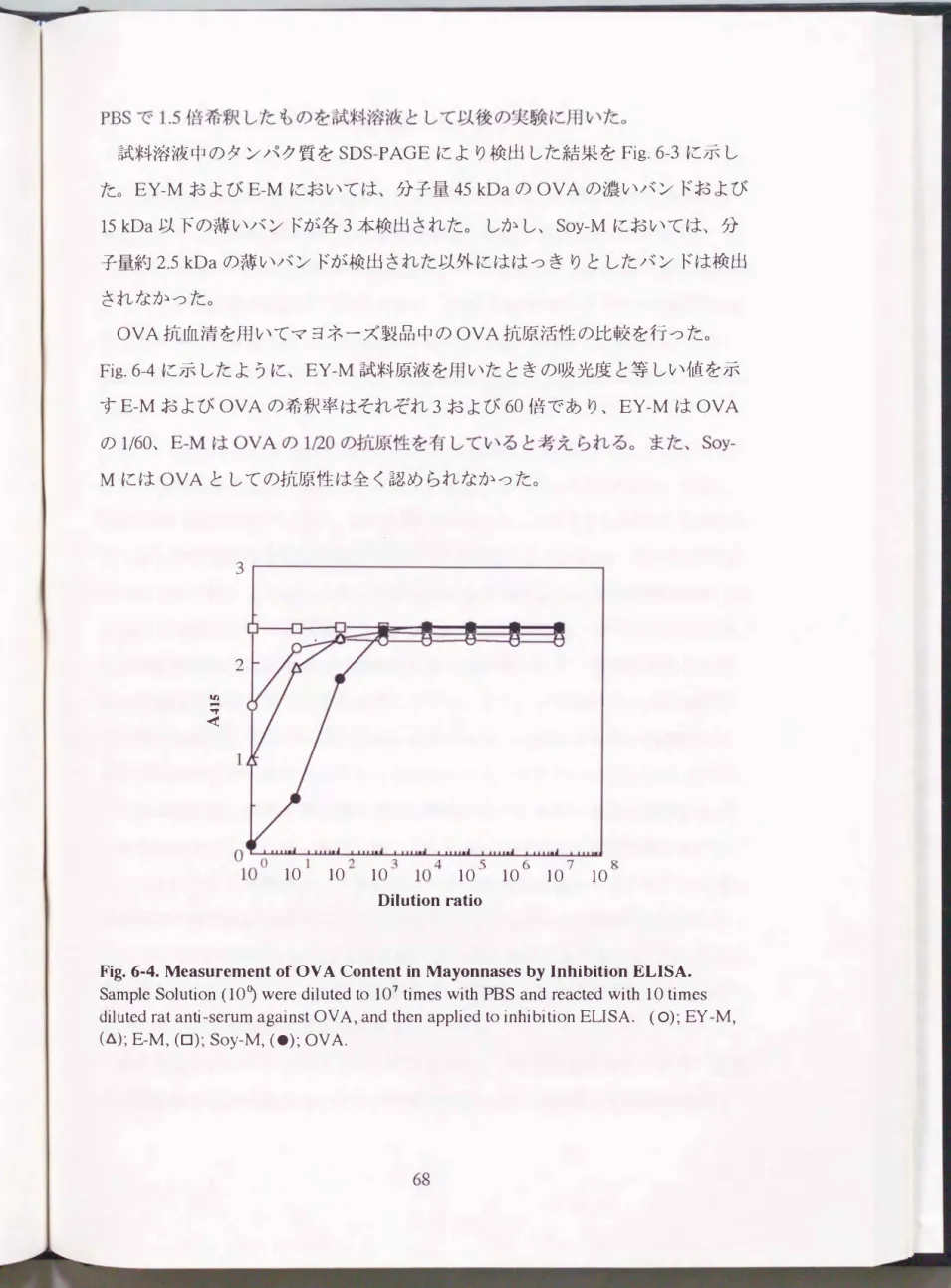
OVA抗血清を用いてマヨネーズ製品中のOVA抗原活性の比較を行った。
Fig.6-4に示したように、 EY-M試料原液を用いたときの吸光度と等しい値を示 すE-MおよびOVAの希釈率はそれぞれ3および60倍であり、 EY-MはOVA の1/1焔60仏、 E
MにはOVAとしての抗原性は全く認められなかつたo
3
同同守司
2
3 4 5 6 7 8
10 10 10 10 ' 10 - 10 - 10 10 � Dilution ratio
Fig. 6-4. Measurement of OV A Content in Mayonn俗的by Inhibition ELISA.
Sample Solution (10うwere diluted to 107 times with PBS and reacted with 10 times diluted rat anti-serum against OV A, and then applied to inhibition ELISA. (0); EY -M,
(6); E-M, (ロ);Soy-M, (・);OVA.
第4節 考 察
加熱や加圧などの食品加工により食品の物性が変化し、 抗原性や栄養価に影 響を及ぼすことが知られているほilshaw et a1., 1982; Nakamura et a1., 1993;
Varshney et al., 1991)0 UDFは分離大豆タンパク質のペプシンおよび微生物由来 プロテアーゼ分解の非消化性画分であり、 UDFには降コレステロール効果があ ることがすでに報告されている(Ogawa et al., 1992)。 今回、 ELISAおよび inhibition ELISAを用いてこのUDFの抗原性の変化をSoyとの比較により評価
した。
まず、 ELISAによりIgEに対する反応性を比較した結果、 UDFおよびUDF- TではIgEとの反応性が著しく低下していることが明らかとなったが、 このこ
とはUDF標品ではアレルゲン性がかなり低下していることを示唆するものであ る。 IgGとの反応性もUDFおよびUDF-Tでは低下していたが、 その低下の度 合いはIgEの場合より軽度であり抗原抗体反応における抗原分子量依存性がIgE とIgGでは異なることが示唆された。 Burks et a1. (1992) は、 ピーナツおよび大 豆抽出物を熱処理した場合には抗原性の変化は認められず、 酵素消化によりは じめて抗原性が低下することを報告している。 また、 ある種のチーズや納豆な どの発酵食品ではタンパク質の分解が顕著で、あり、 それにともない抗原性が大 きく低下することも報告されている(Hassan et a1., 1997; Yamanishi et a1., 1995)。
これらの結果は、 酵素や微生物の適切な利用が低アレルゲン食品の開発に有用 であることを示している。 また、 IgGよりもIgEに対する反応性が強く低下し ていたことから、 食物タンパク質のプロテアーゼ消化においてはI型アレルギー 抑制効果が優先的に発現することが示唆された。 Inhibition ELISAにおいても UDFおよびUDF司TのIgG反応性の低下が示されたが、 低下率はELISAよりも 高い値が得られ、 どちらの方法が生体内での抗原性をより強く反映しているか
についてはさらに検討する必要がある。
卵の主要アレルゲンとしてOVAやオボムコイドなどがあげられており、 抗体 や実験動物を用いたこれらのタンパグ質のアレルゲン性に関する研究が幅広く
69
行われている(Cooke and Sampson, 1997; Kahlert et a1., 1992; Renz et a1., 1993)。
また、 アレルギー患者用食品の栄養価や晴好性の改善のために代替食品を利用 することも多い。 卵の加工食品であるマヨネーズとその代替食品として大立タ
ンパク質を用いて調製されたマヨネーズ製品のOVAとしての抗原性を
inhibition ELISAにより評価した。 その結果、EY-MよりE-Mの方が抗原性が 強いことが明らかとなった。 これはOVAは卵白中に合まれているタンパク質で あるが、E
とがこの違いをもたらしたと考えられる。 しかし、EY-MにおいてもSDS-
PAGEでOVAが検出され、inhibitionELISAにより抗原性を確認されたことか ら、 卵黄を使用した加工食品中にもかなりの量のOVAが混入している可能性が あることが明らかとなった。 これに対し、Soy-MにはSDS-PAGEにおいて OVAやオボムコイドなどの卵アレルゲンは全く検出されず、inhibitionELISA においてもOVAとしての抗原性は認められなかったことから、 マヨネーズの代 替食品として卵アレルギー患者に利用可能であると考えられる。
今回の研究より、ELISAおよびinhibition ELISAは抗原性測定のための有用 な手段であることが明らかとなったが、 より確実な情報を得るためには、 抗血 清やモノクローナル抗体を用いたイムノプロッテイングなどの方法と併せて評 価する必要があると思われる。
第7章 マウス牌臓細胞を用いた免疫調節機能因子 検定系の開発
第1節 緒 言
われわれの体は常に細菌、 ウィルス、 食品抗原などの異物にさらされており、
これらの生体異物に対する防御機構として免疫系が機能している。 免疫系は大 別して体液性免疫と細胞性免疫とよばれる二つの機構を介して作用する。 体液 性免疫は抗原となる生体異物が体内に侵入するとその抗原と特異的に結合する 抗体がBリンパ球より産生され、 抗原と抗体との反応を介して抗原を除去する。
ヒト抗体には五種類のクラスが存在し、 なかでも生体防御に関連する抗体は IgA、 IgGおよびIgMである。
IgAは血中だけでなく腸管や唾液、 初乳などの分泌液中にも合まれており
(Shorter and Tomおし1982; Watanabe et a1., 1984)、 分泌型IgAは微生物やアレル ゲンなどの生体異物の吸収を阻害する(Shorter and Tomasi, 1982)0 IgGは血中に 最も多量に合まれている抗体であり、 終生免疫の獲得に関与している。 IgGはア レルギーの減感作療法により誘導され、 I型アレルギーの原因抗体であるIgEと 競合することによりアレルギー抑制的に働く(Aalberseet a1., 1993)。 また、 IgM は初期免疫に関与し、 抗原感作を受けると一番最初に産生される抗体である。
生体内ではBリンパ球はTリンパ球やマクロファージなどの免疫関連細胞と 相互作用している。 Bリンパ球の抗体産生は、 Tリンパ球などから産生、 放出さ れるインターロイキン-4(lL- 4)、 IL-5、 インターフエロンy(IFN-y) およびIFN-α
といったサイトカインによってクラス特異的に制御されている(Gauchatet a1.,
1991; Ochel et a1., 1991; Pene et a1., 1988; Rousset et a1., 1991)。 また、 マクロ ファージや抗原提示細胞などの接着細胞はIL-1を産生し(Borish and
Rosenwasser, 1996)、 Tリンパ球の刺激を通じて、 抗体産生に影響を及ぼしてい る。
これまで、 種々の生体および食品成分が抗体産生調節機能を有することが報
告されているが(L im et a1., 1994a, 1996; Kaku et a1., 1997)、 これらの実験はラッ
トリンパ球を用いたもの であり、 抗体産生調節機構の解明には抗体などの免疫 関連試薬が充実しているマウスやヒトの系を用いることが望ましい。 また、 こ れ ら因子の抗体産生調節機能がリンパ球への直接作用だけでなく、 マクロ
ファー ジなどの接着細胞への間接作用を通じて発現する可能性が考えられるの で、 接着細胞存在下におけるマウス牌臓細胞の 抗体産生応答について検討した。
第2節 材料と方法
第1項 一般試薬
マイトジェンは、 コンカナバリンA (Con A)およびポークウィードマイトジェ ン(PWM)を EY Lab. (San Mat eo, CA)より、 リポポ リサッカラ イド(LPS)を Difco Lab. (Detro it, MI)より、フィトヘマグルチニ ン (PHA)をVector Lab.
(Burlingam e, CA)より購入した。 これらのマイトジェンはPBSに溶解し、
0.22μmフィルターで滅過滅菌したものを細胞実験に用いた。
RPMI 1640培地は日水製薬(東京,日本 )より、 2 - [4 - (2-ヒドロキシエチル)- 1 -ピベラ ジニル]エタンスルホン酸(HEPES)は同仁化学(熊本,日本)より、 ペ ニシリンGカリウムおよび、硫酸ストレプトマイシンは明治製菓(東京,日本)よ
り、 炭酸水素ナトリウムは和光純薬(大阪,日本)よりそれぞれ購入した。 RPMI 1640を10.4 mg/ml、IlliPES を1.05 mg/ml、 ペニシリンGカリウムを 50UIl、 硫
酸ストレプトマイシンを 20 UIl の濃度で、超純水に溶解し、 さ らに炭酸ガス でpH を6.8 "-' 7.0に調整後、 漉過滅菌してリンパ球用培地 として使用した(RPMI 1640培地) 。 ディッシュはすべてB ecton Dickinson社(Franklin Lakes, NJ)より
購入し、 培養は24穴ディッシュで、行った。 リンパ球 の遠心分離用のプ ラスチッ クチ ューブはNunc社(Roskilide,Denmark)より購入した。 また、 ウシ胎児血清 (FBS)はInterg en社(Purchas e,NY)より購入した。
ELISA 用試薬ではブロックエースを大日本製薬(大阪,日本)より購入し、 超 純水 で 4 倍に希釈したものを ブロッキン グ溶液として、10倍に希釈したものを
サンプルおよび抗体希釈液として使用した。 マウスIgA、 IgG、 IgM 、 ウサギ抗 マウスIgA、 ヤギ抗マウスIgG、 西洋ワサビベルオキシダーゼ( HRP)標識ウサ ギ抗マウスIgAおよびH RP標識ヤギ抗マウスIgGはZymed社.( S an Francisco,
CA)より購入し、 ヤギ抗マウスIgM およびHRP標識ヤギ抗マウスIgM は Capp el社(WestChester, P A)より購入した。
第2項 細胞調製および細胞培養
リンパ球採取 用として、 8"'--'9週齢BAL B /cマウスをセアック吉富(吉富 ,日 本)より購入した。 エーテル麻酔下で、マウスを屠殺後、 採取した牌臓からRPMI 1640培地に 細胞を分離し、 金属メッシュで鴻過することにより組織片を除去し
た。 この細胞懸濁液を370Cで 0"'--'6時間静置して、 プラスチックディッシュに 細胞を接着させることにより接着細胞を除去した。 非接着細胞懸濁液を回収し、
細胞をRPMI1640培地で 3 回洗浄した。 各洗浄過程の遠心分離は3500x g、 5分 間行った。 血球計測板を用いて総細胞数を 2.5X 106 cell s/mlとなるように 調整し、
FBSを10%添加した細胞懸濁液を24穴ディッシュに 添加して 59るC02通気下、
37 0Cで培養を行った。 マイトジェン溶液は培養液 1 /10量を添加した。 培養終了 後、 培養液を回収し、 400x gで 5分間遠心分離し培養上清を回収した。 牌臓細 胞はコール ターインダストリアルDカウンタ一(C oulterE lectronics L td., L uton,
Eng land)を用いて細胞数を測定し、 培養上清はELISAにより抗体の定量を行う まで -300Cで凍結保存した。
第3項 ELISA;,去による抗体の定量
I mmuno-plate Maxi sorp C96をNunc社(Roskilid e,Denmark)より、 ELISA プ レート洗浄液はTween20をナカライテスク(京都,日本)より購入し、 0.05%
Tween 20合有生理リン酸緩衝液(TPBS)を調製して使用した。 発色基質溶液は 、 O.006%H2Ü2合有0.2M クエン酸緩衝液(pH 4.0)、 超純水および 6 mgJml 2 , 2'-ア
ジノービス(3-エチルベンゾチアゾリン-6-スルホン酸)ジアンモニウム塩( ABTS)
溶液を10: 9 : 1で混合して用い、 反応停止液には 1.59るシュウ 酸溶液を用いた。
また、 吸光度の測定に は東ソ一社製(東京,日本)のマイクロプレート分光光度計、
MRP-A4iを使用した。
50mM炭酸緩衝液で1,000倍に希釈した固相抗体を各ウエノレに100μlずつ添加
し、 370Cで1時間保温した。 ブロッキング溶液を各ウェルに 300μlずつ添加し、
40Cで一晩放置してプレート のブロッキングを行った後、 培養上清および標準抗 体を50μlずつ加え、 370Cで1時間保温した。 さらに、 1,000倍に希釈したHRP 標識抗体を各ウェルに 100μlずつ添加し、 370Cで1時間保温した。 各操作の聞 はTPBSで3回洗浄を行った。 発色基質溶液を100μlずつ添加して370Cで15分 間保温した後、 1.5%シュウ酸を100μiずつ添加して発色反応を停止させ、 マイ クロプレート分光光度計に より 415 nmの吸光度を測定した。
第3節 結 果
第1項 抗体産生に及ぼす接着細胞の影響
牌臓細胞を接着細胞存在下あるいは 370Cで30分間静置して接着細胞を除去 した条件下で、 25μgJml のLPSを添加して 培養したときの培養上清中の抗体価
の経時的 変化をEL1SAで測定した結果をFig.7-1 に示した。 接着細胞を除去し た場合、 培養上清中の19A濃度は全体的に低く、 培養 10日目までゆるやかに増
加した。 IgG濃度は最初の5日間はゆるやかに増加した後、 7日目に急激に上昇 したが、 7日目以降はIgG濃度はほぼ一定に保たれた。 IgMの蓄積もゆるやか なも のであり、 培養 5日目まで少しずつ上昇した後、 ほぼ一定 値を与えた。
接着細胞存在下では、 IgA濃度は 培養開始7日固まで速やかに上昇し、 他のク ラスの抗体よりも急速に 蓄積された。 IgG濃度は 5日目までは 接着細胞除去区の 場合と同様の値を与えたが、 7日目以降の濃度は接着細胞除去区より高い値を与 えた。 19M濃度は5日目に急激に上昇したが、 5日日以降は 抗体濃度はほぼ一定 値を与えた。
つぎに、 抗体産生に及ぼす接着細胞の影響を検討するため、 接着細胞を除去 時間 を変えて調製した牌臓細胞を25μg/ml のLPS存在下で7日間培養したとき
400 1.2
300
百E2曲
E
百0.8〈ha可e 。� 0.4 1∞
2 4 6 8 10 12 2 4 6 8 10 12
Cultivation period (days)
400 300
自
主2∞E
ol) 100
2 4 6 8 10 12
Fig.7・1. EtTect of Adhesive Cells on Immunoglobulin Production by Mouse
Splenocytes. Cells were cultured in the presence (・) or absence (0) of adhesive cells.
25μg/ml of lipopolysaccharide was added to the culture medium. Values are mean :t SE (n=3 or 4).
120 100
‘恒5回。ω
�
802ω
-畠z
qb C電,40
20
2 4 6 8
Adhesion period (hr)
Fig.7・2. EtTect of Adhesion Period on Immunglobulin Production by Mouse
Splenocytes Stimulated with Lipopolysaccharides. Cells were cultured for 7 days in the presence of 25μg/ml of lipopolysaccharides. Values are mean:t SE (n=3 or 4).
(0); IgA, (口);IgG, (ð.); IgM.
の抗体産生量を測定した。 Fig.7-2に示すように、 2時間以内の接着細胞除去時 間では、 19A、 IgGおよびIgM濃度は接着時間の延長にともない急速に減少し、
6時間の除去 により抗体産生はほとんど認められなくなった。 抗体濃度の減少率 は抗体のクラスによって異なり、 IgM濃度の減少が最も急速であり、 IgA濃度 の減少がもっともゆるやかであった。 接着細胞合有牌臓細胞の抗体濃度の1/2 を 与える接着細胞除去時間は、 IgAで1.23、 IgGで0.58、 IgMで0 .35時間であっ た。 以上の結果に従い、 以後の実験では、 接着細胞除去時聞は6時間とした。
第2項 抗体産生に及ぼすマイトジェンの影響
牌臓細胞を接着細胞存在下および非存在下で、 種々の濃度のLPSを添加して 7日間培養したときの培養上清中の抗体量を定量した。 Fig.7-3 に示したように、
接着細胞非存在下では、 LPS添加濃度2.5μg/mlでIgA濃度が3.6倍に増加した。
接着細胞存在下では、 添加濃度25μg/mlでIgA濃度が4.0倍に増加し、 最高濃 度を与えた。 IgG濃度は、 接着細胞非存在下の場合にはLPS濃度2.5μg/ml(110 倍増)、 接着細胞存在下ではLPS濃度25μg/ml(42倍増) で最大値を与えた。 ま
た、 IgM濃度は、 接着細胞非存在下ではLPS濃度2.5μg/ml (21倍増)、 接着細胞 存在下ではLPS濃度25μg/ml(67倍増) で最大値を与えた。 これらの結果より、
マイトジェンの添加濃度は最大の抗体産生増強効果が得られた25μg/mlとし、
種々のマイトジェンの抗体産生調節効果について検討した。
接着細胞存在下および非存在下におけるマイトジェンの影響を検討するため、
370CでOあるいは6時間静置した牌臓細胞を終濃度25μgfmlのLPS、 ConA、
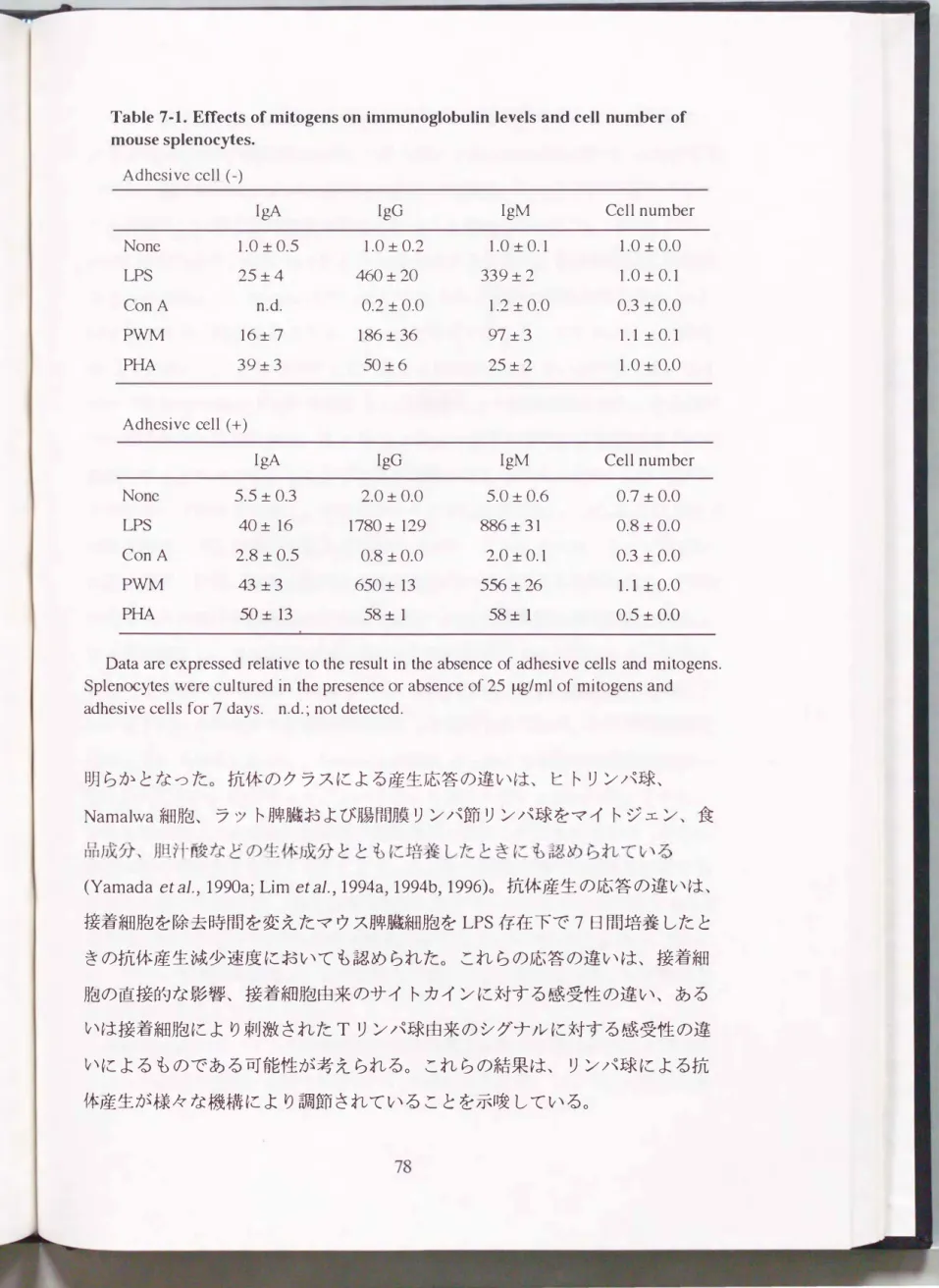
PWMもしくはPHAの存在下で7日間培養した。 Table7-1にマイトジェン添加 区の抗体濃度を接着細胞およびマイトジェンがいずれも存在しない場合の各ク ラスの抗体濃度を1としたときの相対抗体濃度として示した。 接着細胞および マイトジェン非存在下の各クラスの抗体濃度は、 IgAが1.1ng/ml、 IgGが5ng/
ml、 IgMが460 ng/mlであった。 マイトジェンを接着細胞除去牌臓細胞に添加し たとき、 LPSは19A濃度を25倍、 IgGおよびIgM濃度を数百倍に増加させたこ とから、 LPSはIgA産生増強効果よりIgGおよびIgM産生増強効果の方が大き
いことが示唆された。 ConA存在下では、 IgAおよびIgG濃度が減少したが、
細胞数も30%に減少した。 P京市4はIgA、 IgGおよびIgM濃度を上昇させたが、
80 3 30
60
h
旦E 40
�
ヨ2主
4
炉ω圃4 o 胸a圃a 同2 凶2
1 0
勺3nu l
勾,ゐnu l
l nu l
nu nu l
nu l q4 nu -EA
LPS conωntration (時/凶)
Fig.7・3. Dose-dependent Effect of Lipopolysaccharides on Immunoglobulin
Production by Mouse Splenocytes. Cells were cultured for 7 days in the presence (・) or absence (0) of adhesive cells. Values are mean:t SE (n=3 or 4).
19A産生増強効果は他のクラスの抗体 への効果より小さいものであったo PHA は今回用いた マイトジェンのなかで最も強いIgA産生増強効果(39倍増)を示し たが、 IgGおよびIgM濃度 もそれぞれ50倍および25倍に増加させた。
接着細胞はマイトジェン非存在下で も 抗体濃度を上昇させた。 接着細胞はLPS およびPWMのIgGおよびIgM産生促進効果をさらに増強し、 接着細胞 および LPS存在下で最も高いIgG濃度が得られた。 LPSおよびPWMがIgA産生に及 ぼす影響はIgGおよびIgM産生へ の影響 より小さかった。 ConA 存在下では、
細胞数が30%に減少 したが、 IgAおよびIgM濃度はわずかに増加した。 PHA 存在下では、 接着細胞の存在によりIgA、 IgGおよびIgM濃度が上昇したが、
その効果は LPSおよびP\\ゐfより低いものであった。 また 、 PHAおよび接着細 胞の存在下では細胞数の減少が認められた。
第4節 考 察
LPS存在下において、 接着細胞が抗体産生の経時的変化に及ぼす影響を検討 した結果 、 19Aが最も早く培地中に蓄積され、 抗体産生が開始されるまでの時間 や抗体濃度が一定レベルと なるまでの日数が抗体のクラス により異なることが
Table 7・1. EtIects of mitogens on immunoglobulin levels and cell number of mouse splenocytes.
Adhesi ve cell (-)
IgA IgG IgM Cell number
None 1.0 :!: 0.5 1.0 :!: 0.2 1.0 :!: 0.1 1.0 :!: 0.0
LPS 25:!: 4 460:!: 20 339:!: 2 1.0 :!: 0.1
ConA n.d. 0.2 :t0.0 1.2:!: 0.0 0.3 :!: 0.0 P\\札4 16:!: 7 186:t 36 97 :!: 3 1.1 :!: 0.1
PHA 39 :!: 3 50:!: 6 25:!: 2 1.0 :!: 0.0
Adhesive cell (+)
I会生 IgG IgM Cell number
None 5.5 :!: 0.3 2.0:!:0.0 5.0 :!: 0.6 0.7:!: 0.0
LPS 4O:!: 16 1780:!: 129 886:!: 31 0.8 :!:O.O
ConA 2.8 :!:0.5 0.8 :!: 0.0 2.0:!: 0.1 0.3 :!: 0.0
PWM 43 :!: 3 65O:!: 13 556:!: 26 1.1 :!: 0.0
PHA 50 :!: 13 58:t 1 58:!: 1 0.5 :!: O.O
Data are expressed relative to the result in the absence of adhesive cells and mitogens.
Splen∞ytes were cultured in the presence or absence of 25 �/ml of mitogens and adhesive cells for 7 days. n.d.; not detected.
明らかとなった。 抗体のクラスによる産生応答の違いは、 ヒトリンパ球、
Namalwa細胞、 ラット牌臓および腸間膜リンパ節リンパ球をマイトジェン、 食 品成分、 胆汁酸などの生体成分とともに培養したときにも認められている (Yamada etal., 1990a; Lim etal., 1994a, 1994b, 1996)。 抗体産生の応答の違いは、
接着細胞を除去時聞を変えたマウス牌臓細胞をLPS存在下で7日間培養したと きの抗体産生減少速度においても認められた。 これらの応答の違いは、 接着細 胞の直接的な影響、 接着細胞由来のサイトカインに対する感受性の違い、 ある いは接着細胞により刺激されたTリンパ球由来のシグナルに対する感受性の違 いによるものである可能性が考えられる。 これらの結果は、 リンパ球による抗 体産生が様々な機構により調節されていることを示唆している。
マイトジェンがリンパ球の増殖および抗体産生を調節することが報告されて
いる(Gianniet al., 1980; Newberry et al., 1973; Yamada et aJ., 1990a)。 本研究にお いても、 種々のマイトジェンを培地に添加した場合、 ConA添加区以外ではマ ウス牌臓リンパ球の抗体産生が増強されることを明らかにした。 LPSおよび PWMは抗体産生、 特にIgGおよびIgMの産生を促進し、 接着細胞はこの効果
をさらに増強した。 Bucala(1992) はLPSによるB細胞の細胞分裂促進および IgM産生促進は接着性のマクロファージに依存すること、 マクロファージ由来
のIL-1が補助因子として作用していることを報告している。 本研究では、 IgM だけでなく19AおよびIgGの産生もLPS刺激により促進されており、 今回認め られた抗体産生促進効果は、 IL-1やIL-1により刺激を受けたT細胞由来の他の 種類のサイトカインによるものである可能性が考えられる。 また、 LPSやブド ウ球菌のペプチドグリカンがマクロファージ非依存的に19A、 IgGおよびIgMの 分泌を誘導したとの報告もある(Dziar ski, 1980)。 これらマクロファージ依存性
に関する不一致は、 細胞の調製方法や細胞集団の違いによると思われる。 P\\市f によるヒト末梢血単核細胞の抗体産生細胞への分化誘導能は接着細胞を除去し たときに減少し、 その原因は細胞間相互作用の阻害によるものであると報告さ れている(Rosenberg and lipsky, 1979)。 本研究では、 接着細胞存在下において、
LPSとP\\市fの抗体産生促進効果は類似した傾向を示したが、 その制御機構は 異なっているかもしれない。 ConAは終濃度25μg/mlで顕著な細胞数の減少が 認められており、 他のマイトジェンに比ベて毒性が強いと思われる。 しかし、
ConA添加によりわずかに抗体産生促進効果が認められたものもあり、 さらに 細胞集団の解析などを行うことにより、 ConAの牌臓細胞への影響を検討する 必要があると思われる。 PHAは今回使用したマイトジェンのなかで最も19A産 生促進活性が強かった。 PHAはT細胞のマイトジェンであることから、 PHA がヘルパーT細胞を刺激し、 19A産生を促進するようなサイトカインが放出さ れたためであると考えられる。
本研究において、 マウス牌臓細胞の初代培養系を用いて接着細胞およびマイ トジェンが抗体産生に及ぼす影響について検討した結果、 マイトジェンは抗体
産生調節機能を有しており、 接着細胞はその効果を促進することが明らかとなっ た。 今回確立した免疫調節因子の機能検定系は免疫調節機能を有する食品成分 の作用機構の解明に有用な手段を与えることが期待される。
第8章 マウス牌臓細胞の抗体産生に及ぼす食品成分の影響
第1節 緒 言
われわれの体は常に細菌、 ウィルス、 食品抗原などの異物にさらされており、
これらの生体異物に対する防御機構として免疫系が機能している。 免疫系は大 別して体液性免疫と細胞性免疫とよばれる二つの機構を介して作用する。 体液 性免疫は抗原となる生体異物が体内に侵入するとその抗原と特異的に結合する 抗体がBリンパ球より産生され、 抗原と抗体との反応を介して抗原を除去する。
ヒト抗体には五種類のクラスが存在し、 なかでも生体防御に関連する抗体は 19A、 IgGおよびIgMである。
19Aは血中だけでなく腸管や唾液、 初乳などの分泌液中にも合まれており
(Shorter and Tomasi, 1982; Watanabe et a1., 1984)、 分泌型19Aは微生物やアレル ゲンなどの生体異物の吸収を阻害する(Shorter and Tomasi, 1982)0 IgGは血中に 最も多量に合まれている抗体であり、 終生免疫の獲得に関与している。 IgGはア レルギーの減感作療法により誘導され、 I型アレルギーの原因抗体であるIgEと 競合することによりアレルギー抑制的に働く(Aalberseet a1., 1993)。 また、 IgM は初期免疫に関与し、 抗原感作を受けると一番最初に産生される抗体である。
生体内でBリンパ球はTリンパ球やマクロファージなどの免疫関連細胞と相 互作用している。 Bリンパ球の抗体産生は、 Tリンパ球などから産生、 放出され るインターロイキン-4(IL-4)、 IL-5、 インターフエロン y(IFN-y) およびIFN-α
といったサイトカイン によってクラス特異的に制御されている(Gauchatet a1.,
1991; Ochel et a1., 1991; Pene et a1., 1988; Rousset et a1., 1991)。 また、 マクロ ファージや抗原提示細胞などの接着細胞はIL-1を産生し(Borish and
Rosenwasser, 1996)、 Tリンパ球の刺激を通じて、 抗体産生に影響を及ぼしてい る。
ラクトフェリンやカゼインなどの乳タンパク質はin vitroにおいて抗体産生調 節機能を発現することがヒト型ハイブリドーマ実験を用いて報告されているが
81
(Yamada et a1., 1990a, 1991a)、 その作用機構は不明な点が多い。 また、 不飽和脂 肪酸はリンパ球の増殖を抑制す る こと により免疫応答に影響を及ぼすことが報 告されているが(Calderet a1., 1992; Soyland et a1., 1993; Tonkin and B rostoff,
1978)、 抗体産生に及ぼす影響についての研究は非常に限られており、 いずれも ラットリンパ球を用いたものである (Limet a1., 1996; Yamada et a1., 1996)。 そこ で、 前章で確立したマウス牌臓細胞培養系を用いて 、 乳タンパクおよび脂肪酸 の抗体産生に及ぼす影響およびその作用機構について検討を行った。
第2節 材料と方法
第1項 一般試薬
乳タンパク質であるß- ラクトグロプリン(ß-LG)、 αーカゼ、イン(α-CA)、Pーカ ゼイン(ß- CA)、Kーカゼ、イン(K-CA)およびトランスフエリン(T下)はSigma社
(St.lρusi, MO)より、 ラクトフェリン(LF)は和光純薬(大阪,日本)より購入し た。 脂肪酸ナトリウム塩は 、 ステアリン酸(SA)、 オレイン酸( OA)、 y- リノレン 酸( GLA)、 αーリノレン酸をフナコシ(東京,日本)より、 アラキドン酸(AA)、 エ イコサベンタエン酸( EPA)、 ドコサヘキサエン酸(DHA)をSigma社より、 リノー ル酸(LA)をナカライテスク(京都,日本)より購入した。 抗酸化剤であるアスコ ルビン酸(AsA)およびbutylated hydroxytoluene ( BHT)はSigma社より、 ケルセ チン(Que)はナカライテスクより、Pーカロチン( ß-Car)は ICN社(Costa Mesa,
CA)より購入し、 αートコフエロール(Toc)およびトコトリエノール(T3)はエー ザ、イ(東京,日本)より、 セサミン(Ses)はサントリー(大阪,日本)より提供され
た。 乳タンパク質および脂肪酸は生理リン酸緩衝液(PBS; pH 7.4)に 、 抗酸化剤 はエタノールにそれぞれ溶解し、 0.22μmフィルターで漉過滅菌したものを実験 に用いた。
RPMI1640培地は日水製薬(東京,日本)より、 2-[4-( 2-ヒドロキシエチノレ)- 1 -ピベラジニル]エタンスルホン酸(HEPES)は同仁化学(熊本,日本)より、 ペ ニシリン Gカリウムおよび硫酸ストレプト マイシンは明治製菓(東京,日本)よ
り、 炭酸水素ナト リウムは和光純薬よりそれぞれ購入した。 RPMI1640 を
10.4 mg/ml、 HEPESを1.05 mg/ml、 ペニシリン Gカリウムを50 U/l、 硫酸スト レプトマイシンを20 U/l の濃度で超純水 に溶解し、 炭酸ガスで pHを6.8� 7.0 に調整後、 減過滅菌してリンパ球用培地として使用した(RPMIl640培地)。
ディッシュはすべて Becton Dickinson社(FranklinL akes, NJ)より購入し、 細胞 培養は24穴ディッシュで行った。 リンパ球の遠心分離 用の プラスチックチュー
ブはN unc社(Roskilide, Denmark)より購入し、 ウシ胎児血清(FBS)はIntergen 社(Purchase, NY)より購入した。
ELISA試薬ではブロックエースを大日本製薬(大阪3日本)より購入し、 超純 水で 4倍に希釈したものをブロッキ ング溶液として、 10倍に希釈したものをサ
ンプルおよび抗体希釈液として使用した。 マウスIgA、 IgG、 IgM 標準抗体 、 ウ サギ抗マウス19A、 ヤギ抗マウスIgG、 西洋ワサビベルオキシダーゼ(HRP)標 識ウサギ抗マウスIgAおよびHRP標識ヤギ抗マウスIgGはZymed社(San
Francis∞, CA)より、 ヤギ抗マウスIgM およびHRP標識抗マウスIめ4は Cappel社(West Chester, PA)より購入した。
サイトカ イン中和抗体は、 抗IL-1α、 抗IL-1ß、 抗IL-2 、 抗IL-4および抗 IFN-y 抗体を Genzyme社(Cambridge, M A)より購入した。 それぞれ PBSで 抗
IL-1α、 抗IL-1ß、 抗IL-2 は50倍、 抗IL-4 は25倍、 抗IFN-yは1,0∞倍に希釈 して1 ml の細胞懸濁液に 50μiずつ添加した。
第2項 細胞調製および細胞培養
リンパ球採取用として、 8�9週齢BALB/cマウスをセアック 吉富(吉富3日 本)より購入した。 エーテル麻酔下で、マウスを屠殺後、 採取した牌臓からRPMI
1640培地に細胞を分離 し、 金属メッシュで鴻過することにより組織片を除去し た。 この細胞懸濁液を 370CでOある いは6時間静置してプラスチックディッシユ に細胞を接着させることにより接着細胞を除去した。 非接着細胞懸濁液を回収 し、 細胞をRPMI1640培地で 3回洗浄した。 各洗浄過程の遠心分離は3,500 x g、
5分間行ったo 血球計測板を用い て総細胞数を2.5 X 106 cells/ml となるように調
整し、 FBSを10%添加した細胞懸濁液を24穴ディッシュにま いて5% C02通気 下、 37 0Cで培養を行った。 培養終了後、 培養液を回収し、 400 xgで5分間遠心 分離し培養上清を回収した。 沈殿した牌臓細胞はコールターインダストリアル
Dカウン ター(Coulter Electronics Ltd., Luton, England)を用いて細胞数を測定し、
培養上清は分析を行うまで-300Cで凍結保存した。
第3項 生細胞率の測定
細胞懸濁液 に 1%トリパンブ、ルー溶液を1/10容量添加して5分間静置し、 染 色された死細胞数および染色されていない生細胞数を計数 し、 生細胞率を算出
した。
第4項 ELISA法による抗体量の定量
Immuno-plate Maxi sorp C96をNunc社(Roskilide, Denmark)より購入した。
プレート洗浄液はTween 20をナカライテスク(京都,日本)より購入し、 0.05%
Tween 20合有PBS(TPBS)を調製 して使用した。 発色基質溶液は、 0.006%
Hの2合有0.2 Mク エン酸緩衝液(pH 4.0)、 超純水および6 mg/ml 2- 2'-アジノービ ス(3-エチルベンゾチ アゾリンー6-スルホン酸)ジアンモニウム塩(ABTS)溶液を 10: 9: 1で混合して用い、 反応 停止液には 1.59るシュウ酸溶液を 用いた。 ま た、
吸光度の測定には東ソ一社製(東京,日本)のマイクロプレート分光光度 計 、 MRP-A4iを使用した。
50 mM炭酸緩衝液で1,000倍に希釈した固相抗体を各ウエルに100μiずつ添加
し、 370Cで1時間保温した。 ブロッキング溶液を各ウェルに 300μlずつ添加し、
4 0Cで一晩放置してプレートのブロッキングを行った後、 培養上清および標準 抗体を50μlずつ加え、 370Cで1時間保温した。 さらに、 1,000倍に希釈した HRP 標識抗体を各ウェルに100μiずつ添加し、 370Cで1時間保温した。 各過程 の聞は TPBSで3 回洗浄を行った。 発色基質溶液を100μlずつ添加して370Cで 15分間保温した後、 1.5%シュウ酸を100μlずつ添加して発色反応を停止させ、
マイクロプレート分光光度計により415 nmの吸光度を測定した。
84
第5項 ELISA法によるサイトカインの定量
培養上清中のIL-4、 IL-5およびIFN-yの定量はPerSeptive社(Framingham,
MA) よりEIAキットを購入し、 キットの試薬を用い、 キットの方法に準じて実 験を行った。 プレートに50μ!の培養上清またはIL-4、 IL-5、 IFN-y標準液を添
加しておだやかに撹持し、 室温で1時間振とうしながら放置した。 ビオチン標 識抗IL-4、 IL-5または IFN-y抗体を50μiずつ添加して 室温で1時間振とうしな がら放置した。 さらに、 HRP標識ストレプトアビジンを50μiずつ添加し、 室 温で30分間振とうしながら放置した。 各過程の聞は洗浄液で3�4回洗浄を行っ
た。 TMB試薬を50μiずつ添加して室温で15分間振とうしながら反応させ、 反 応停止液を50μlずつ添加した後、 450 nmの吸光度を測定した。
第6項 TBA値の測定
和光純薬より過酸化脂質ーテストワコーを購入し、 キットの方法に準じて実験 を行った。 培養上清 100μ!と1/12 N硫酸4.0 mlを混合して10%リンタングステ ン酸水溶液0.5 mlを加えた。 撹持して室温に5分間放置し、 1,500x gで10分間
遠心分離した沈殿物に1/12 N硫酸2.0 mlおよび10%リンタングステン酸水溶液 0.3 mlを加えた。 沈殿物をよく懸濁し、 1,500x gで10分間遠心分離を行った。
沈殿物に超純水を4.0 ml加えて懸濁し、 TBA試薬1.0 mlを添加し よく 撹持し、
遠沈管上にガラス玉を置き、 沸騰水浴中で60分間加熱した後、 流水で冷却した。
n-ブタノールを5.0 ml加え、 20秒間十分に混合して抽出し、 1,5∞xgで10分間 遠心分離した。 上層のn-ブタノール層について励起波長515 nm、 蛍光波長553 nmの蛍光光度を測定した。
第7項 統計解析
結果の統計解析は Duncan (1955 )の new multiple range testもしくはt検定を 用いて 行った。
第3節 結 果
第1項 マウス牌臓細胞による抗体産生に及ぼす乳タンパクの影響
終濃度o � 1 mg/mlの乳タンパクを培地に添加し、 牌臓細胞を接着細胞存在下 および非存在下で7日間培養したときの培養上清中のIgAおよびIgG濃度をFig.
8-1およびFig.8-2に示した。 LFは接着細胞存在下では終濃度100μgfmlで、 接 着細胞非存在下では終濃度1 mg/mlで強い19A産生促進活性を示した。 ß-LGは 添加濃度依存的に19A産生を促進する傾向を示し、 接着細胞の有無にかかわら ず終濃度1∞μg/mlでIgA産生促進効果が最大となった。 α-CAおよびβCAも
30 Lactoferrin
!
固人l
10 ト j I \
30 α-Casein
E
30 30
ß-Lactoglobulin I Transferrin
20 20
10 ト / \ 10
00プF平ー�一・・・・....一一
3 • � 4 -� .. _ � -2 _ �-1
o 10 -10 . 10- 10. 10- 10-10' 0 1O-�1O-' 10v 10' 10� lOJ 10
30 30
ß-Casein
I
K-Casein20
/
2010卜 iν 10
Protein concentration (μg/mJ)
Fig.8・1. EtTect of Milk Proteins on IgA Production by Mouse Splenocytes. Cells were cultured for 7 days in the presence (・) or absence (0) of adhesive cells. Various concentration of milk proteins were added to the culture medium.
濃度依存的にIgA産生を促進し、 終濃度1 mgJmlで促進効果が最大となった。
特に接着細胞存在下、 1 mgJmlのβCA添加において強いIgA産生促進効果が認 められた。 K-CAおよびLFと同じ鉄結合性タンパク質であるTFには19A産生 調節活性はほとんど認められなかった。
LFは弱いIgG産生促進傾向を示すにすぎなかったが、 ß-LGおよびα-CAは 強い産生促進効果を示した。ß-LGは終濃度100μg/mlで、 α-CAは終濃度1 mgJ mlで接着細胞の有無にかかわらず最大の促進効果を与えた。 ß-CAは終濃度1 mgJmlにおいてIgG濃度を数百倍に上昇させ、 非常に強いIgG産生促進活性を 有することが明らかとなった。 K-CAおよびTFにはIgAと同様にIgG産生調節
80 80
Lactoferrin I IβLactoglobulin 80 Transferrin
トr � Iト
60
60卜 1 60
20� cf 1. 20
3ゐ百40
。?ロ写 - d
ハU o IM 唱EA ハU 勺& 'EA nu ハu nU ハu 'EA 内,Lnu ,EA ハU qJ ハu 4 。
20
6 ß-Casein
4
聖
百
2
K-Casein
80 80
α-Casein
A『 ハU 唱EA q3 ハU 今,ゐ ハU 唱aA ハU 唱EA nu ハU 唱1A ハU 今,h ハU 'EA --〕す ハu nu
60 60
判 (-gh回) 判 (-ghロ)
む制,....-:,
o
10 -10 -10- 10- 10- 10- 10、
目 、 円 ':J AProtein concentration (陪Iml)
Fig. 8-2. Effect of Milk Proteins on IgG Production by Mouse Splenocytes. Cells were cultured for 7 days in the presence (・) or absence (0) of adhesive cells. Various concentration of milk proteins were added to the culture medium.
活性はほとんど認められなかった。
つぎに、 19AおよびIgG産生促進活性が認められた乳タンパク質の抗体産生 促進効果発現の時間的経過を示した。 なお、 培養10日目におけるリンパ球の生 細胞率は全て889る以上であった。 Fig.8-3に示したように、 100μg/mlのß-LG 添加区では、 接着細胞存在下においては培養5日目まで、 接着細胞非存在下に おいては7日目まで19A濃度がゆるやかに増加した後に一定値を与えた。
1∞μg/mlのLF添加区では接着細胞存在下においては培養5日目まで、 接着細 胞非存在下においては7日固まで19A濃度が増加した。 また、 1 mgfmlのLF添 加区においては接着細胞の有無にかかわらず培養10日目まで19A濃度が増加し た。 1 mg/mlのα-CA添加区では、 接着細胞存在下では、 培養5日目まで19A濃
2∞
150
1∞
百
三E恥 50
宮帽冨EEω回。ω 。 o
r;
・白壁画(1 mg/凶)
150
司固。君圃£
1∞
50
0
� Lactoglobulin (1ω時Iml)
�Casein (1 mg/凶)
Lactρferrin (1∞時Iml)
Lactoferrin (1 mg/m1)
o 2 4 6 8 10 0 2 4 6 8 10 0 2 4 6 8 10 12
Cultivation perion (day)
Fig.8・3. Time Courses of IgA Contents in Culture Medium of Mouse Splenocytes Cultivated with九1iIk Proteins. Cells were cultured in the presence (・) or absence (0)
of adhesive cells. Values are means ::t SE (n=3 or 4).
88
度が増加した後に一定値を与えたが、 接着細胞非存在下では、 培養10日目まで 19A濃度が継続的に増加した。 1 mg/mlのβCA添加区では、 接着細胞存在下で は、 培養7日目まで、19A濃度が大幅に増加したのに対して、 接着細胞非存在下で は5日目までにわずかしか19A濃度が増加していなかった。
Fig.8-4に示したように、 乳タンパク質非存在下では培養開始後1日自にIgG 濃度が上昇し、 その後顕著な変化は認められなかったo 100μg/mlのβLGもし
くは LF、 1 mg!mlのLFもしくはα-CAを添加した区では、 培養開始後3'"'-'5日 目までIgG濃度が増加した後、 ほぼ一定値を与えた。 また、 これら乳タンパク 質添加区では、 接着細胞のIgG産生増強効果は認められなかったo これに対し て、 1 mg/mlのß-CA添加では、 培養3日目までIgG濃度が大幅に増加したが、
接着細胞存在下ではさらに強いIgG産生促進効果が認められた。
ヨω
。l)z C
101
。
吻4
510
回ω ω 図
。ω
。凶
同圃
Con仕01
同郡山
2
4 6 810 0 2
4 6 810 0
Cultivation perion (day)
Lactoferrin (1ω時Iml)
Lactoferrin (1 mgI凶)
2
4 6 810 12
Fig.8・4. Time Courses of IgG Contents in Culture Medium of Mouse Splenocytes Cultivated with九1ilk Proteins. Cells were cultured in the presence (・) or absence ( 0)
of adhesive cells. Values are means :t SE (n=3 or 4).
第2項 マウス牌臓細胞数に及ぼす乳タンパク質の影響
前項でいくつかの乳タンパク質が抗体産生促進活性を有することを示したが、
その作用機構のーっとして細胞数の変化が考えられる。 そこで、 抗体産生促進 活性が認められた乳タンパクの細胞数への影響を検討し、 培養5日目の無添加 区に対する相対細胞数をFig.8-5に示した。 100μg/mlß-LG添加区において接着 細胞存在下の細胞数が有意に高い値を与えた。 100μg/mlLF添加区では無添加区 より細胞数が減少したが、 1 mg/ml LF添加区では無添加区より細胞数が多いこ とが明らかとなった。 1 mg/mlα-CA添加区では接着細胞存在下の細胞数が無添 加区より多く、 1 mg/ml ß-CA添加区でも有意に高い値が得られることが示され た。 特に接着細胞存在下では無添加区に比べて3倍以上の細胞数が得られた。
4
d
3
」呂田ロ&2 ω •
3ω 2
E ー
・‘ω国国B圃d , ω
。
None ß-LG LF LF α-CA ß-CA (100問Iml) (1∞μg/ml) (lmg/ml) (1mg/ml) (1mg/ml)
恥1ilk protein
Fig. 8-5. EtTect of Milk Proteins on Cell Number of Mouse Splenocytes. Cells were cultured for 5 days in the presence (圏) or absence (口)of adhesive cells. Data are means ::t SE (n=3 or 4) and values without a common letter are significantly different at pく0.05.
第3項 乳タンパク質の抗体産生調節活性とサイトカインの関与
乳タンパク質の抗体産生調節機構を明らかにするために、 1 mgfmlβCA存在 下でサイトカイン中和抗体として抗IL-1α、 抗IL-1ß、 抗ILム抗IL-4および抗
IF1吋-yを培地に添加して7日間培養したときの19AおよびIgG濃度をFig.8-6に 示したo ß-CAの19A産生促進効果の発現は抗IL-1α、 抗IL-1ß、 抗ILム抗IL- 4共存下では接着細胞存在下および 非存在下のどちらにおいても影響を受けなかっ た。 しかし、 抗IFN-y添加区では接着細胞存在下でのIgA産生には影響を及ぼ さなかったが、 接着細胞非存在下では19A産生促進効果の発現が阻害され、ß-
CA無添加と同レベルのI�生濃度が得られた。 また、 βCAのIgG産生促進効果 は、接着細胞存在下において抗IL-2添加区で若干低下する傾向が認められたが
その他は顕著な変化は認められなかった。
つぎに、 100μgfmlLF添加区において 培養上清中に放出されるサイトカイン量 の定量を行った。 LFを添加しただけでは培養上清中のサイトカイン濃度は検出
。
None ß-CA ß-(つA+ anti・IL-lα ß-CA + anti-IL-l ß
PベコA+ anti・IL-2
PベゴA+ anti-IL-4 ß-CA + anti・IFN吋
IgA (ng/ml)
・ 20 40 。
IgG (時Iml)
2 4 6 8 10
B
Fig.8・6. E釘'ect of Neutralizing Antibody against Cytokines on Immunoglobulin Production by Mouse Splenocytes in the Presence of ß-Casein. Splen∞ytes with
(圏) or without (口) adhesive cells were cultured for 7 days in the presence of 1 mg/ml of ß-casein. Data are means::t SE (n=3 or 4) and values without a common letter are significantly different at pく0.05.