ミツバチ科学18(4):165-174 HoneybeeScience(1997)
ニホ ン ミツバチ-生態 とその飼育法
Ⅳ-吉田 忠晴
7.
ニホ ンミツパテとセイヨウミツパ テの 種間相違点 ニホ ンミツバチとセイ ヨウ ミツパ テの種間の 相 違 に つ い て は,Tokuda(1924),岡 田 (1955,1958,1986,1991),Sakagamietal. (1958-1960),Sasakietal.(1995)によっ て報告 されている.既 に前述 の5.ニホ ンミツ バチの生態, 6. 可動巣枠式巣箱 による飼育法 と採蜜 の項 でニホ ン ミツバチの特徴的な諸行動 について解説 したが,上記 の報告やその後 の研 究成果 を基 に,(1)形態 ・生理,(2)行動 ・生 悲,(3)訪花,(4)害敵 ・病気,(5)生産物 の 各項 目につ いてニホ ンミツパ テとセイ ヨウ ミツ パ テの相違点 についてま とめた. (1)形態 ・生理 ニホ ンミツバチ とセイ ヨウ ミツパテの形態 ・ 生理 に関す る相違点 を衰 2に示 した. セイ ヨウ ミツバチの自然分布域 は, ヨー ロッ ノ1., アフ リカ, マグカスカル,地 中海東方 はイ ラン西部 まで,さ らに ロシア,アフガニスタン, 中国北部, そ してモ ンゴルが東方 の南限であ っ た.現在で はセイ ヨウ ミツバ チの人為 的な導入 によ って 自然分布域 か ら生息地域 はさ らに拡大 し,南北 アメ リカ大陸, ニ ュージー ラン ド, オ ース トラ リアや中国, 日本 な どのアジア地域 に 広が っている (Ruttner,1988). 青森県下北半 島を北限 と して本州以南 に生息 しているニホ ン ミツバチは,北海道南部地域で の生息 は未確認であ り (岡田,1957;1990), 筆者 の最近 の調査か らも,分布域 は本州以南で あることが確 め られた. 図.79に示 した女王蜂 と働 き蜂, さ らに雄蜂 図79 ニホンミツバチ (上)とセイヨウミツバチ (下)の女王蜂と働 き蜂 図80 ニホンミツバチ (右)とセイヨウミツパテ (左)の後週の廼脈 ニホンミツバチでは後麹の中脈(M3.1)が顕 著 にみ られ る166
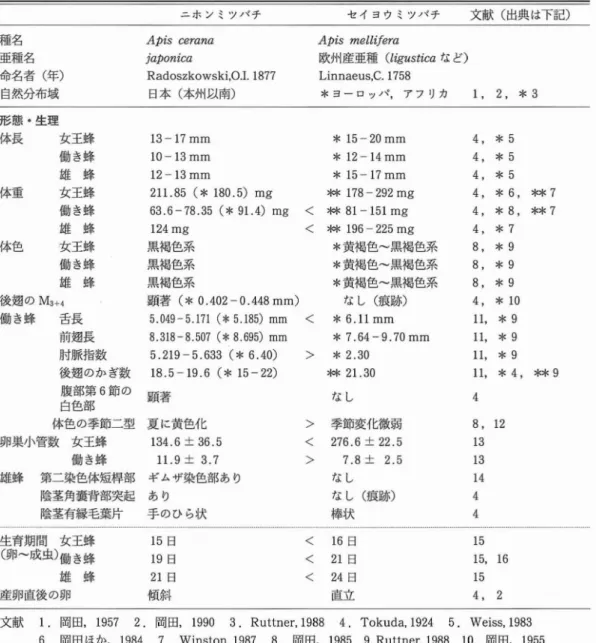
表2ニホンミツバチとセイヨウミツバチの形態 ・生理の相違点
ニホンミツバチ セイヨウミツバチ 文献 (出典は下記)
種名 Aptscerana ApismelLifera
亜種名 )'aponica 欧州産亜種 (ligusticaなど)
命名者 (年) Radoszkowski,0.Ⅰ.1877 Linnaeus,C.1758
自然分布域 E]本 (本州以南) *ヨーロッパ,アフI)カ 1, 2, *3 形態 ・生理 体長 女王蜂 13-17mm 働 き蜂 10-13mm 雄 蜂 12-13mm 体重 女王蜂 211.85(*180.5)mg 働 き蜂 63.6-78.35(*91.4)mg 雄 蜂 124mg 体色 女王蜂 黒褐色系 働 き蜂 黒褐色系 雄 蜂 黒褐色系 後逸のM3+4 顕 著 (*0.402-0.448mm) *15-20mm *12-14mm *15-17mm * 178-292mg < * 81-151mg
<
**196-225mg *黄褐色∼黒褐色系 *黄褐色∼黒褐色系 *黄褐色∼黒褐色系 な し (痕跡) 働 き蜂 舌長 5.049-5.171(*5.185)mm<
*6.llmm 前延長 8.318-8.507(辛8.695)mm 肘脈拍数 後延のかぎ数 腹部第6節の 白色部 体色の季節二型 卵巣小管数 女王蜂 働 き蜂 雄蜂 第二染色体短梓部 陰茎角褒背部突起 陰茎有縁毛莞片 5.219-5.633(*6.40) 18.5-19.6(* 15-22) 顕著 夏 に黄色化 134.6±36.5 ll.9± 3.7 ギムザ染色部あり あ り 手 のひら状 *7.64-9.70mm>
*2.30 * 21.30 な し コhP 5 5 Ttk ・ ) 徴 22 2 . 跡 靴 .J は 鳩 節 6 . 7 . L L 状 季 27 な な 棒>
<
>
7 7 9 Ei Ei Ei 5 5 5 6 8 7 9 9 9 10 9 9 9 4 * * * * * * * * * * * * * * 2 , , , , , , , , , , l 1 1 l , 3 3 4 4 4 4 4 4 4 8 8 8 4 1 1 1 1 4 8 1 1 1 4 4 生育期間 女王蜂 (卵∼成虫)働 き蜂 雄 蜂 産卵直後の卵 日 日 日 止 T 15 19 21 儲 日 日 日 立 16 21 24 直く
く
く
61 2 5 5 5 . 1 1 1 4文献 1.岡田,1957 2.岡田,1990 3.Ruttner,1988 4.Tokuda,1924 5.Weiss,1983 6.岡田ほか,1984 7.Winston.1987 8.岡田,1985 9.Ruttner,1988 10.岡田,1955 11.岡田ほか,1956 12.Tsurutaeta1.,1989 13.Sasakieta1..1995 14.HoshibaandOkada,1986 15.Singh,1962 16.MishraandDogra,1983
の 体 長,体 重 はTokuda (1924), 岡 田 は か (1984),岡 田 (1985)の報 告 か らセ イ ヨウ ミツ バ チ (W eiss,1983;W inston,1987)より小 型 で あ る. ニホ ン ミツパ テの体色 は女 王蜂,働 き蜂 ,雄 蜂 と も黒褐 色系 で (岡 田,1985),特 に 雄 蜂 は黒色 に近 い. セ イ ヨウ ミツバ チで は亜種 間 で体色 は異 な り, イ タ リア ン種 は黄 褐色 で あ るが, カーニオ ラ ン種 や コ- カ シア ン種 はニホ ン ミツバ チ に近 い黒褐 色 で (Ruttner,1988), 見 間違 え る ことさえ あ る. ニ ホ ン ミツバ チ とセ イ ヨウ ミツバ チの形 態上 の違 い と して顕 著 な点 は,斑 80に示 した働 き 蜂 の後麹 の遡 脈 で あ る. こ ホ ン ミツバ チで は後 麹 に0.402-0.448mm の中脈 (M3+4) が顕著 にみ られ るの に対 して, セ イ ヨウ ミツバ チで は 存 在 し な い か,痕 跡 程 度 で あ る (Tokuda, 1924;岡 田, 1955). この点 は トウ ヨウ ミツバ チ の亜種 間で共通 で あ り,女 王蜂,堆蜂 に もみ られ る. この廼脈 の特徴 は低倍 率 のル ーペで確 認 す ることが で きる.
図81 ニホンミツバチ働き蜂の体色変化 蜂群内に混成する黒色型と黄色型 働 き蜂 の 舌 長 は5.049- 5.171mm (岡 田, 1956),5.185mm (Ruttner,1988)であ り,セ イ ヨウ ミツバチの6.llmm (Ruttner,1988) に比較 して短 く,前廼長 は採集 した蜂群 によっ て差があるが, 二ホ ン ミツバ チの方がやや短 い 傾向がある.前週 の肘脈指数 (cubitalindex) は,5.219-5.633 (岡田,1956),6.40 (Rut -tner,1988)のニホ ン ミツバチの値 よ りセイ ヨ ウ ミツパテは2.30 (Ruttner,1988) と有意差 が認 め られた.後廻前縁 にある鈎数 について は, ニホ ンミツパテが18.5-19.6(岡田,1956), 15-22(Tokuda,1924)に対 してセイ ヨウ ミ ツバチは21,30(Ruttner,1988)と多少差があ る程度であ った.働 き蜂 は腹部第6節 の白色部 がセイ ヨウ ミツバチに比較 して顕著である点が 特徴的である(Tokuda,1924),また働 き蜂 は 季節的に二型 の体色がみ られ る (図81).同一 蜂群内で8月∼10月 に黄色型 が, 10月下旬∼ 翌 年 の5月 に は黒色型 が多 く出現 す る傾 向が ある (岡田,1985;Tsurutaeta1.,1989).体 色変化 は腹部小循板,腹部第3・4背節片節間 膜,腹部腹面 にみ られ, その変化 は嫡期 に受 け る25℃∼38℃ の温度 に関係 してお り, 34℃以 上 で黄色が, それ以下 で は黒色が出現す ること が確かめ られた (Tsurutaeta1.,1989). 女王蜂 の卵巣小管数 (左右 に存在す る卵巣 の 合計) は, ニホ ンミツパ テで は134.6本 に対 し てセイ ヨウ ミツバチで は276.6本で (図82), ニホ ン ミツバチはセイ ヨウ ミツバチの約半分で ある(Sasakieta1.,1995).ニホ ン ミツバチの 蜂群 サイズはセイ ヨウ ミツバチよ り小 さいが, 図82 ニホンミツバチ (右)とセイヨウミツバチ (左)の卵巣 (撮影 ;小野正人) 図83 ニホンミツバチ (上) とセイヨウミツバチ (下)の陰茎有縁毛葉片 (矢印) これはニホ ン ミツバチ女王蜂 の産卵数が少 ない ことに起因 していると考 え られ る. ニホ ン ミツ バ チ働 き蜂 の卵巣小管数 の合計 は11.9本, セ イ ヨウ ミツバチは7.8本 であ る (Sasaki,eta1., 1995). ニホ ン ミツバチの方 が卵巣小管数 は多 く,大 きな卵巣 を もっているため,働 き蜂 によ る産卵が起 きやす い理 由に もな っている. ミツバチ属9種 の中で,セイ ヨウ ミツバチ, トウヨウ ミツパテ, オオ ミツバチ, コ ミツバチ の4種 につ いて は,雄 蜂 の染色体数 はn-16 であることが確かめ られて いる.Hoshibaand
168 図84 ニホンミツバチ (左)とセイヨウミツバチ (右)の女王蜂による産卵直後の卵 Okada(1986)は,染色体 の核型分析の結果
,ニ
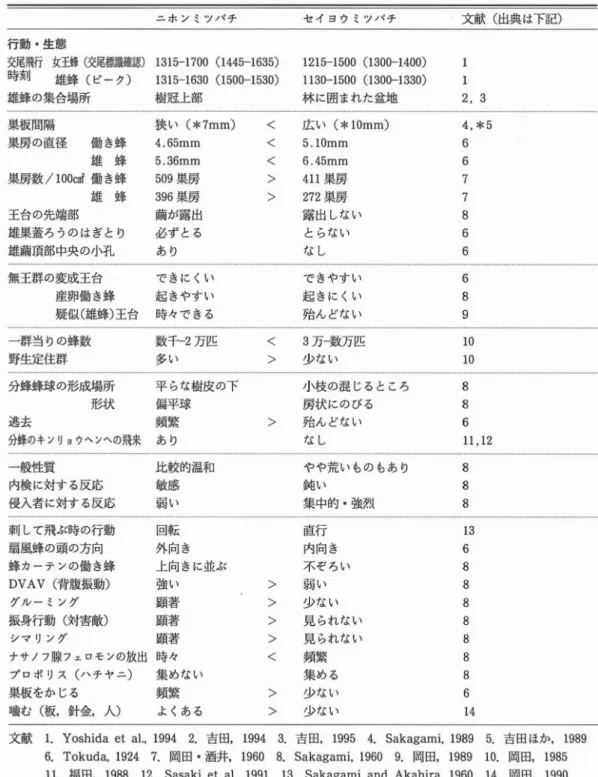
ホ ンミツバチとセイ ヨウ ミツバチの核型 は両種 ともほぼ同 じであるが, ニホ ンミツバチ雄蜂の 第二染色体短梓部 はギムザ染色 で薄染 され る が,セイ ヨウ ミツバチには認め られないことを 報告 した.雄蜂の生殖器 について は陰茎角蛮背 部 にニホ ン ミツパ テで は突起 が あ るのに対 し て,セイヨウ ミツバチでは存在 しないか,痕跡 程度である (Tokuda,1924).図 83に示 した 陰茎有縁毛葉片(fimbriatelobe)は,ニホ ンミ ツバチは手のひ ら状であるのに対 して,セイ ヨ ウ ミツパ テ は棒 状 を示 して い る (Tokuda, 1924). ニ ホ ン ミツバ チ の生 育 期 問 は イ ン ド亜 種 (Apisceranaindica)と同様で,女王蜂 は15日 (Singh,1962),働 き蜂 は19日 (Singh,196 2;Mishra and Dogra,1983), 雄蜂 は21日 (Singh,1962)であ り,セイ ヨウ ミツパテより 短 いことが観察 された. セイ ヨウ ミツバチ女王蜂が産卵 した直後 の働 き蜂の卵 は,巣房の底 に直立 してお り,3日間 の卵期 の終 わ り頃 にな ると傾斜 した状態 にな る. しか しニホ ンミツパテでは産卵直後か ら既 に 傾 斜 し て お り (Tokuda,1924; 岡 田, 1990), セイ ヨウ ミツバチと異 なることが観察 される (斑 84). (2)行動 ・生態 ニホ ンミツバチとセイ ヨウ ミツパテの行動 ・ 生態に関す る相違点 を表3に示 した. 日本 には在来種のニホ ンミツバチと導入種の セイ ヨウ ミツパテの2種が同所的に生息 してい 1100 12叩 1300 140D 1500 1680 1700 令 SL r再 生 撃 た だ 令 S LJ 点 せ 撃 た 群 11t)0 1200 1300 1400 1500 1600 1700 時 刻 図85 ニホンミツバチとセイヨウミツバチの女王蜂 と雄蜂の交尾飛行時刻 る. そのような環境の中で,ニホ ンミツバチ女 王蜂 の飛行時刻 は13:15-17:00の問で,雄蜂 と の 交 尾 が 成 立 し た 印 で あ る交 尾 標 識 (mating sign)を確認 で きた時刻 は 14: 45-16:35であった.一方, セイヨウ ミツバチ女王 蜂の飛行時刻 は,12:15-15:00の間で,交尾標 識 の確認時刻 は13:00-14:00とニホ ン ミツバ チ女王蜂 はセイ ヨウ ミツバチよ り1-1.5時間 遅 いことが認 め られた (図85), (Yoshidaet a1.,1994). ニホ ン ミツバ チ雄蜂 の 出巣 時刻 は13:1 5-16:30の間で,飛行 の ピー クは 15:00-15:30 であった.セイ ヨウ ミツバチ雄蜂の出巣時刻 は ll:30-15:00の間で, ピークは 13:00-13:30 とニホ ンミツバチ雄蜂 は女王蜂 と同様 にセイヨ ウ ミツバチよ り約2時間遅 い ことが認 め られ た (図85),(Yoshidaeta1.,1994). 空中の特定な空間に存在す る女王蜂 と雄蜂の 交尾場所である雄蜂の集合場所 に関 しては, ニ ホ ンミツバチは周囲の地形 の中で目立つ木の樹 冠上を集合場所 に してお り,セイヨウ ミツバチ の林 に囲まれた盆地状の地形 の場所 とは異 なる ことが認 め られ (吉田,1994),2種間には時間 と空間の双方の違 いによる生殖隔離機構が存在表3ニホ ン ミツバチ とセイ ヨウ ミツバチの行動 ・生態の相違点 ニホ ンミツバチ セイ ヨウ ミツバチ 文献 (出典 は下記) 行動 ・生態 交尾飛行 女王蜂 (交尾標識確認)1315-1700(1445-1635) 1215-1500(1300-1400) 時刻 雄蜂 (ピーク) 1315-1630(1500-1530) 1130-1500(1300-1330) 雄蜂 の集合場所 樹冠上部 林 に囲まれた盆也 3 1 1 2 巣板間隔 狭 い (*7mm) < 広 い (*10mm) 巣房の直径 働 き蜂 4.65mm 雄 蜂 5.36mm 巣戻数/100CⅡf働 き蜂 509巣房 雄 蜂 396巣房 王台の先端部 繭が露 出 雄巣蓋 ろ うのはぎとり 必ず とる 雄繭頂部中央 の小孔 あ り 無王群の変成王台 で きに くい 産卵働 き蜂 起 きやすい 疑似 (雄蜂)王台 時々で きる
<
5.10mm<
6.45mm>
411巣房>
272巣房 露 出 しない と らない な し 5 * 4 6 6 7 7 8 6 6 で きやす い 起 きに くい 殆 んどない 一群当 りの蜂数 数千-2万匹<
3万一数万匹 野生定住群 多 い>
少 ない 分蜂蜂球 の形成場所 平 らな樹皮 の下 小枝 の混 じるところ 形状 偏平球 房状 にのびる 逃去 頻繁>
殆 ん どない 分蜂のキンリョウへンへの飛来 あ り な し 一般性質 比較的温和 やや荒 い もの もあ り 内検 に対す る反応 敏感 鈍 い 侵入者 に対す る反応 弱 い 集中的 ・強烈 刺 して飛ぶ時の行動 回転 扇風蜂 の頑の方向 外 向 き 蜂 カーテ ンの働 き蜂 上 向きに並ぶ DVAV(背腹振動) 強 い グルー ミング 振身行動 (対害敵) シマ リング ナサノフ腺フェロモンの放出 プロポ リス (-チャニ) 巣板をか じる ー_V な 著 著 着 々 め 繁 顧 顧 鋸 時 集 頻 噛む (板,針金,人) よ くある 直行 内向 き 不 ぞろい > 弱 い > 少 ない > 見 られない > 見 られない < 頻繁 集 め る>
少 ない>
少 ない 3 4 1 6 8 8 8 00 8 8 8 6 1文献 1.Yoshidaeta1.,1994 2.吉 田,1994 3.吉 田,1995 4.Sakagami,1989 5.吉田はか,1989 6.Tokuda,1924 7.岡田 ・酒井.1960 8.Sakagami,1960 9.岡田,1989 10.岡田,1985 11.福 田,1988 12.Sasakieta1.,1991 13.
し, そ の態 果 ,両 程 の共 存 が可 能 とな って い る こ とが 示 さ れ †こ (志 田,1995).
巣 板 と巣 板 の 間 隔 (beespace)は,ニ ホ ン ミ ツバ チ はセ イ ヨウ ミツバ チ に比 較 して狭 い こ と が 報 告 さ れ (Sakagami,1959), 吉 田 は か
SakagamiandAkahira,1960 14.岡臥 1990
(1989)紘, セ イ ヨ ウ ミツ バ チ の巣 坂 間 隔 で あ る 10mmよ Y)ち - ホ ン ミツ パ テ ほ 30%穐 め ,
7mmと して良 好 な結 果 を得 て い る.
巣 房 の直 径 は,ニ ホ ン ミツバ チ働 き蜂 は4.65
170 図86 ニホンミツバチ (左)とセイヨウミツバチ (右)の王台 ニホンミツバチは茶褐色の繭が琵出する ミツバチの方が大 きく,雄蜂 について もそれぞ れ5.36mm,6.45mm と同様の傾向である.ニ ホ ンミツバチ雄蜂巣房の大 きさはセイ ヨウ ミツ バチ働 き蜂巣房 と同程度の大 きさであると考え られる (Tokuda,1924). 100cm2当た りの巣 房数の調査 において も, ニホ ンミツパテ働 き蜂 では509巣房であるのに対 して,セイ ヨウ ミツ バチでは411巣房である (岡田・酒井,1960). 同 じサイズの ラングス トロス式巣枠 に造 られた 巣房数 は, ニホ ンミツバチでは約4,400, セイ ヨウ ミツバ チで は約3,400と両 者 に 1,000巣 房 の近 くの差がある (吉 田ほか,1989). ニホ ンミツバチでは王台先端のろう部分が働 き蜂 によって取 り除かれ,茶褐色の繭が露出す るが,セイ ヨウ ミツパテで は繭の露出はみ られ ない (図86),(Sakagami,1960),筆者の観察 では,王台が塞がれたてか ら約3日後 に繭が露 出す る. ニホ ンミツバチ雄蜂児の巣房 は蓋がけされた 級,必ず表面のろうがはぎとられ,その雄繭の 頂部中央 に小孔があるのは, セイ ヨウ ミツバチ には見 られない特徴である (図87),(Tokuda, 1924). 蜂群が無王状態になった場合には, セイ ヨウ ミツバチに比較す るとこホ ンミツバチでは変成 王台 はできに くい傾向があ り(Tokuda,1924), 働 き蜂産 卵 の開始 が早 く起 こる (Sakagami, 1960). また, ニホンミツパテでは雄性の桶の ある疑似王台が時々観察 される (岡田,1989). 1群当た りの蜂数 は, ニホ ンミソバチでは数 図87 ニホンミツバチ (左)とセイヨウミツバチ (右)の雄蜂巣房 千か ら上限が2万匹程度で,セイ ヨウ ミツバチ の3万-数万 匹 に比 べ る と半分以下 と考 え ら れ る (岡乱 1985).野生 的に定住 している群 は,ニホ ンミツバチの方が圧倒的に多 く (岡田, 1985), 前述 の野生群の生息場所の観察か らも 自然巣を造 って生息 している. 分蜂 による蜂球 は, ニホ ンミツ/ヾチは太 い枝 が分かれた平 らな樹皮の下 に偏平球状 に形成 さ れるのに対 して,セイ ヨウ ミツバチは小枝の混 じる場 所 に枝 を包 む よ うに房 状 に造 られ る (Sakagami,1960). これ らの形状 は母女王蜂 と共 に蜂群が飛 び出す第一分蜂でよ くみ られ る が,働 き蜂の数が多い強群 の場合に起 こる未交 尾女王が分蜂す る第二分蜂 では第-分蜂のよう に一定 していない. ニホンミツパテは貯蜜が減少 した り,-チノ スッヅ リガの幼虫 によ って巣板が食 い荒 らされ た り,セイ ヨウ ミツパテの盗蜂を受 けた りす る と頻繁に逃去す るが, セイ ヨウ ミツバチでは殆 ど起 こらない (Tokuda,1924). 図88 ニホンミツパテ (左)とセイヨウミツバチ (右)の分蜂
図 89 ニホンミツバ チ (上)とセイ ヨウミ ツバチ (下)の
扇風
巣門に対する向き
が異な る 東洋 ランの一種であるキ ンリョウへ ンの花 に ニホ ンミツバチが特異的 に誘引され,分蜂群が 飛 来 す る現 象 が 発 見 さ れ た (福 田,1988; Sasakieta1.,1991). キ ンリョウヘ ンには働き蜂が餌 として採取す る蜜や花粉 はな く,誘引 物質 として放 出され る花香 の研究が進め られて いるが,セイ ヨウ ミツバチはこの ランに全 く興 味を示 さない. ニホ ンミツバチの一般性質 は,セイ ヨウ ミツ バチよ り温和であるため (Sakagami,1960), 内検時に煉煙器 は必要 と しない. しか し巣板 を 移動 した り,持 ち上 げた りす ると神経質 に巣坂 上を走 り回 ることが多 い(Sakagami,1960). セイ ヨウ ミツバチは巣箱への侵入者 に対す る攻 撃が強 いが, ニホ ンミツパテはその点 は弱 いよ うである(Sakagami,1960).そのためクロゴ キブ リ, カマ ドウマの昆虫類か らナメクジ, ム カデなど20数種 の小動物 がみ られ る. 働 き蜂が人畜を刺 し飛 び上が る時,刺針部 は 身体 と連結 している筋肉部分か らちぎれ,毒歪 が付 いた状態で針が残 る. その際ニホ ンミツバ チは刺 した針を中心 に回転 しなが ら飛 び上が る が, セイ ヨウ ミツバチは回転す ることはな く, 刺 して か ら直線 的 に飛 び上 が る (Sakagami andAkahira,1960). 扇風蜂の頭 の向 きは, ニホンミツバチは巣門 に対 して外側 に向けて風 を送 り込む扇風である が,セイ ヨウ ミツパテは内側 に向け,空気を排 出す る (図89),(Tokuda,1924).扇風行動 は 2種問で大 きな違 いがある. 巣坂上 をカーテン状 に覆 っている多数の働 き 蜂 は,ニホ ンミツバチでは上向 きに整然 と並ぶ 特徴があるが,セイ ヨウ ミツバチでは巣坂上で 色々な方向を示 している. 働 き蜂 は他 の働 き蜂や女王蜂の体 の上 に乗 っ た り, しがみついた りして腹部を背腹方向にす ばや く動かすDVAV (dorsoventralabdomト natvibration)と呼ばれ る背腹振動を行 う.こ の行動 はニホ ンミツバチとセイ ヨウ ミツバチの 両種で観察で きるが, ニホ ンミツバチの方がそ の行動が強 く行われ る. 働 き蜂 は中脚で胸部の毛繕 いを行 った り,サ ルの社会でみ られるような個体問での身繕 いと 同様 に,他の働 き蜂が胸部 と腹部の柄部や超の 付 け根の毛を大顎です く, グルー ミングが行わ れ る. グルー ミング行動 は両種 で観察で きる が,ニ ホ ン ミツバ チ の方 が頻 繁 に行 わ れ る (Sakagami,1960). 振身行動 はニホ ンミツバチに顕著 にみ られ る (Sakagami ,1960).飛和 しなが ら巣門に接近 す るスズメバチに対 して門番蜂 は前傾姿勢を と り,腹部を30-40度持 ち上 げて 40-60度左 図90 セイヨウミツバチ働き蜂によるナサノフ腺か らの集合フェロモンの放出
172 表 4ニホンミツパテとセイヨウミツバチの訪花及び害敵 ・病気の相違点 ニホンミツバチ セイヨウミツバチ 文献 (出典は下記) 訪花 訪花植物の選択 多岐に亘 る 採餌圏 狭い 主要植物に集中 1,2
<
広い 3 円舞 40m以内 (*50-70m) 林 80-100m 4,*5,**6 8*蒜論 議 て露 ′S) 約700- < 約1000- 3 雄蜂のキンリョウヘンへの訪花 あり な し 7 害敵 ・病気 盗蜂 対天敵行動 オオスズメバチ 同巣層状物質よる忌避 キイロスズメバチ ハチノスッヅ リガ ウスグロツヅ リガ スムシヒメコマユバチ ミツバチへギイタダニ トウヨウミツパテホコリダニ 腐姐病 時々/個体単位 発達 巣箱内に誘導 顕著 蜂球により熱殺 蜂群内で被害 多発 多い 被害少ない 見 られる 希 頻繁 /集団 未発達 巣門前で応戦 見 られない 刺針行動 被害/抵抗性あり 少ない 少ない 被害多い 見 られない 時々 2 4 4 4 0 -1 3 8 2 8 9 1 8 9 2 1 1 2文献 1.Miyamoto,1958 2.岡田,1991 3.佐々木はか,1993 4.Sakagami,1960 5.沖本 ・佐々木,
1990 6.FrlSCh,1965 7.Sasakieta1.,1991 8.Tokuda,1924 9.岡田,1990 10.Onoeta1..1987 11.Yoshidaeta1.,1989 12.Yoshidaetal‥ 1995 13.酒井 ・佐々木,1989
右 に激 しく数 回振 る行動 を行 な う. シマ リング (Shimmering)は振身行動 と同 様 にニホ ン ミツバ チで顕著 にみ られ る行 動 で あ る (Sakagami,1960).振 動 な どの物 理 的要 因 や巣坂上 の働 き蜂 に息 を吹 きか けた り, スズメ バ チが接近 した場合 に は,一 斉 に シ ャワー とい う特異 な羽音 を発 す る. シマ リングは巣坂上 の 多数 の働 き蜂 が一定方 向 に連鎖 的 に行 うため, 波 を うって い るよ うにみ え る. ナ サ ノ フ腺 か らの 集 合 フ ェ ロ モ ンの 放 出 (scenting)は,セ イ ヨウ ミツバ チで は頻繁 にみ られ るが (図90), ニホ ン ミツバ チで はセ イ ヨ ウ ミ ツ バ チ の よ う に 多 く な い (Sakagami, 1960). ニホ ン ミツバ チ は植物 の樹脂 で あ るプ ロポ リ ス (ハ チャ ニ)を集 め る ことはな く(Sakagmi, 1960), 巣箱 の隊 問 な どの修繕 に はろ うが使 わ れて い る. 巣板 を頻繁 にか じる事 はセ イ ヨウ ミツバ チで はあま りみ られ ないが (Tokuda,1924),生 態 の項 の巣造 りで述 べて よ うに, ニ ホ ン ミツバ チ は巣 板 をか じる ことに よ って保 温効果 を高 めて い るの で は な いか と考 え られ て い る ( Sakag-amiandKouta,1958). 大顎 を使 って噛 む力 はニ ホ ン ミツバ チで は強 く (岡 田, 1990), 観察 時 に手 な どを噛 まれ た り,巣 門 な どの木部 や巣枠 の針 金 を よ く噛 む行 動 が観察 され る. (3)訪 花 ニ ホ ン ミツパ テ とセ イ ヨウ ミツバ チの訪花 に 関す る相 違点 を衰 4に示 した, セ イ ヨウ ミツバ チの訪花 は主 要 な蜜 源植物 に 集 中す る傾 向が あ るが, ニ ホ ン ミツバ チで は多 岐 に亘 る こ とが報 告 され て い る (Miyamoto, 1958).岡 田 (1991) は これ まで の報告 を もと に104種 の訪花 植物 を記 載 して い る. 採 餌 圏 の広 さの違 い は, 収穫 ダ ンスの発音 時 間 と採餌 距離 の関係 か ら考 察 す る と, ニ ホ ン ミ ツバ チ に比 較 して セ イ ヨ ウ ミツバ チ の方 が2 倍 近 い面 積 を採 餌 対 象 に して い る (佐 々木 ほ か,1993). 一 般 に蜜 源 まで の距離 が近 い場 合 に踊 る収穫
ダンスの円舞 は,セイ ヨウ ミツバチでは蜜源ま で の距 離 が80- 100m の場 合 にみ られ るが (Frisch,1965), 二ホ ン ミツバチでは40m以 内(Sakagami,1960),あ るいは50-70m (沖 本 ・佐 々木,1990)の報告がある. 蜜源 までの距離 と方 向 の情報 を示 す8の字 ダンスでは,尻振 り時の中心線上でニホ ンミツ バ チとセイ ヨウ ミツバチ は約250Hzの音信号 を発信す る.両種 とも蜜源が近 いほど信号音 は 短 く,遠 くなるに従 い音 の持続時間は長 くなる (佐 々木 ほか,1993).その結果,1秒 の音信号 はニホ ンミツバチでは約700m, セイ ヨウ ミツ パテで は約1000mと採餌距離 に差が認 め られ た (佐 々木 ほか,1993). 雄蜂が訪花活動を行 うことは, これまで全 く 報告 されていなか った. ところがニホ ンミツバ チの雄蜂が東洋 ランの一種であるキ ンリョウへ ンに訪花 し,花粉媒介を行 っている新知見が確 認 され ことは,セイ ヨウ ミツバチと大 きく異な る点である (Sasakieta1.,199
1
)
.
(4)害敵 ・病気 ニホ ンミツバチとセイ ヨウ ミツバチの害敵 ・ 病気 にに関す る相違点を衰 4に示 した. 他 の蜂群 に入 り込 み,貯蜜 を盗 み出す盗蜂 は, ニホ ンミツバチでは個体単位で時々起 こる が,セイ ヨウ ミツバチは集団で頻繁 に発生す る (Tokuda,1924;Sakagami,1960).特 に蜜源 が少 ない越冬開けや梅雨 の時期 にニホ ンミツバ チに対 してセイ ヨウ ミツバチの激 しい盗蜂が起 こることがある. 天敵であるスズメバチ類 に対す る行動 は, ニ ホ ンミツパテではその防衛行動がよ く発達 して お り, セイヨウ ミツバチでは未発達である (岡 田, 1991). オオスズメバチの攻撃では, ニホ ンミツバチは捕食者 の餌場 マークフェロモ ンを 傍受 して巣門付近で大騒 ぎとな り,蜂の出入 り が減少す る(Onoeta1.,1996).さ らに巣箱内 に誘導す るように, その侵入を許すが,巣緒内 部で蜂球を造 り熟殺す る (図91).一方, セイ ヨウ ミツバチは巣門前で激 しく応戦 し,集団攻 撃を受 けると壊滅的な被害 を受 ける (Tokuda, 1924;Sakagami,1960).オオスズメバチが飛 図91 オオスズメバチに対する巣箱内での熱殺蜂球 釆す ると,ニホ ンミツパテでは粘性のある巣層 状の物質が巣門周辺 に塗 られるが, これは害敵 の侵入防止用 と考え られている (岡乱 1990). セイ ヨウ ミツバチではこのような物質の塗布 は み られない. キイロスズメバチの攻撃に対 して は, ニホ ンミツパテでは蜂球 による熟殺が行わ れるが,セイヨウ ミツパテでは針を盛んに出す 刺針行動がみ られ る (Onoeta1.,1987). ハチノスッヅ リガの被害 はニホ ンミツバチで は大 きく,弱小群,無王群での巣板の食 い荒 し 方 は激 しいが,セイヨウ ミツバチの蜂群内での 被害 は殆 どみ られない (Tokuda,1924;Saka -gami,1960).小型 のウスグロツヅ リガはニホ ンミツパテでは多 く発生す るが,セイ ヨウ ミツ バチでは少 ない (岡田, 1990). ウスグロツヅ リガは巣板 よりも巣箱 の底 に堆積 した巣屑内に み られる.ハチノスッヅ リガとウスグロツヅ リ ガの幼虫に寄生す る小型 の寄生蜂であるスムシ ヒメコマユバチの成虫 は, ニホ ンミツバチの巣 箱 の中で は4
月∼1
1月の間み られ,その数 は セイ ヨウ ミツバチよ り多い (岡田,1991). セイ ヨウ ミツバチでは ミツバチへギイタダニ の防除が必須であるが, ニホ ンミツバチでは蜂 群内か ら発見す ることは希で,寄生率 は極 めて 低 い (Yoshidaeta1.,1989;1995). トウヨウ ミツバチホコ リダニはニホ ンミツバチ働 き蜂の 頭部後面 の頚部両側に陥入 している小孔の中に 生息 している体長0.2mm 弱のダニであるが, セイヨウ ミツバチでの寄生 は確認 されていない (酒井 ・佐 々木,1989). 家畜法定伝染病 に指定 されている ミツバチの 蜂児の病気である腐姐病 は,セイ ヨウ ミツパテiliJF 表5ニホンミツ/ヾチとセイヨウミツパテの生産物の相違点 ニホンミソバチ セイヨウミツバチ 文献 (出典 は下記) 生産物 蜂毒中のメリチン 108±34〃g/毒のう
<
310±59〟g/毒のう アパ ミン 1.6±0.6pg/毒のう<
6.8±1.5pg/毒のう 蜂ろうの融点 65℃ 酸価 5-7 < 炭化水素炭素数31 多い>
炭化水素炭素数35 少ない<
脂肪酸炭素数24 少ない<
63-65℃ 17-20 少ない 多い 多い プラスチ ック人工王椀の口径 6.7mm<
9.3mm 守_諾 完孟 a(蒜壷 78'品 ) 90・2± 23・6-g
<
187・3±27・3-g 4 ローヤルゼリー中の タンパク質 多 い>
少ない 5 ローヤルゼリー中の炭水化物 少 ない<
多い 5 文献 1.井上 ・中嶋,1985 2.Tokuda,1924 3.吉田 ・佐々木,1995 4.吉田はか,1993 5.TakenakaandTakenaka,1996で時々発病 す るが, ニホ ン ミツパ テで の発生 は 希 で あ り, その抵抗性 につ いて注 目されて い る (岡 田, 1991). (5)生産物 ニホ ン ミツバ チ とセイ ヨウ ミツバ チの生産物 に関す る相違点 を表5に示 した. 蜂毒 の主成分 で溶血活性 ペ プチ ドで あ るメ リ チ ンは, ニホ ン ミツバ チで の毒 の う中 の含量 は 108/上gで あ るの に対 して, セ イ ヨウ ミツパ テ で は310〟g,神経毒 で あ るアパ ミンはニ ホ ン ミツパ テで1.6FLg,セ イ ヨウ ミツパ テで は 6.8 〃gとセ イ ヨウ ミツバ チ の 1/3の量 で あ った (井上 ・中嶋, 1985). 蜂 ろ うの融点 はニ ホ ン ミツバ チで は65℃, セ イ ヨウ ミツバ チ は63-65℃ と大 きな差 は認 め られなか った (Tokuda,1924).遊離脂肪酸 の測定 によ る酸化 は, ニ ホ ン ミツパ テで は 5-7, セ イ ヨウ ミツバ チで は 17-20とニ ホ ン ミ ツバ チの方 が低 い値 で あ った. ガスク ロマ トグ ラフィーによる主要構成成分 の比較 で は,全炭 化水素 中 の炭素数31の割 合 はニ ホ ン ミツバ チ は多 く, セイ ヨウ ミツバ チで は少 なか った. ま た炭素数35は反対 にニ ホ ン ミツバ チで は少 な く,セイ ヨウ ミツバ チで は多 か った.炭素数24 の遊離脂肪酸 の割合 はニホ ン ミツバ チで は少 な く, セイ ヨウ ミツバ チで は多 い とい うよ うに, 両 者 の 違 い が 認 め られ た (吉 田 ・佐 々 木, 1995). セイ ヨウ ミツバ チでの ローヤルゼ リー生産 や 女 王養成 に用 い られて い る市販 の プ ラスチ ック 製人工王椀 の 口径 は9.3mm で あ る. ニホ ン ミ ツバ チで はプ ラスチ ック管 で 自作 した王椀 の検 討 か ら, 口径 が6.7mm の王椀 で良好 な受入 率 を示 した. またその王椀 によ る移 虫48時 間後 の ローヤルゼ リー採乳量 は, ニホ ン ミツバ チで は90.2mg, セ イ ヨウ ミツバ チ で は 187.3mg とニ ホ ン ミツバ チ は約半 分 の採 乳 量 で あ った (吉 田 はか, 1993). ローヤルゼ リー成分 中の タンパ ク質 は, ニホ ン ミツパ テで はセイ ヨウ ミツバチよ り多 く,戻 水化物 につ いて はその逆 の関係 で あ ることが明
ら か に な っ た (Takenaka and Takenaka,
1996).
一 つづ く一 参考文献 は最終号 にまとめて掲載 す る. (〒194 町田市玉川学園 6-1-1
玉川大学 ミツバチ科学研究施設) YosHIDA,TADAHARU.Japanese honeybee,ecol -ogyanditsrearingmethodsⅣ.HoneybeeSci -ence(1997)18(4):165-174.HoneybeeSci.Res. Center,Tamagawa Univ.,Machida-shi,Tokyo,