Pathological studies on the cell
characteristics and roles of glial cells in
myelin mutant rats
著者
Izawa Takeshi
内容記述
学位授与大学: Osaka Prefecture University(大阪
府立大学), 学位の種類: 博士(獣医学), 学位記番
号: 論獣第140号, 学位授与年月日: 2010-03-31,
指導教員: 山手丈至.
大阪府立大学博士(獣医学)学位論文
Pathological studies on the cell characteristics and roles of
glial cells in myelin mutant rats
(ミエリン疾患ミュータントラットにおける
グリア細胞の特性と役割に関する病理学的研究)
井澤 武史
Contents
Preface... 1
Chapter 1 Glial pathology during development of hypomyelination in the myelin vacuolation (mv) rat... 6
Section 1-1. Analysis of glial pathology in the spinal cord of the mv rat ...7 ...
Introduction 7
...
Materials and Methods 8
... Results 10 ... Discussion 11 ... Summary 14 ... Figure Legends 15 ... Plate I 17 ... Section 1-2. Identification of attractin-expressing cells in the normal spinal cord 20
...
Introduction 20
...
Materials and Methods 21
... Results 22 ... Discussion 23 ... Summary 24 ... Figure Legends 25 ... Plate II 26
Chapter 2 Microglial activation and gray matter lesions in the mv rat... 27
Section 2-1. Analysis of expression patterns of microglia-related factors in the mv rat ...28 ...
Introduction 28
...
Materials and Methods 29
... Results 31 ... Discussion 31 ... Summary 34
... Table 1 35 ... Figure Legends 36 ... Plate III 37
Section 2-2. Analysis of expression patterns of myelin proteins in the mv rat ...38 ...
Introduction 38
...
Materials and Methods 39
... Results 42 ... Discussion 45 ... Summary 48 ... Tables 2, 3 49 ... Figure Legends 50 ... Plate IV 53
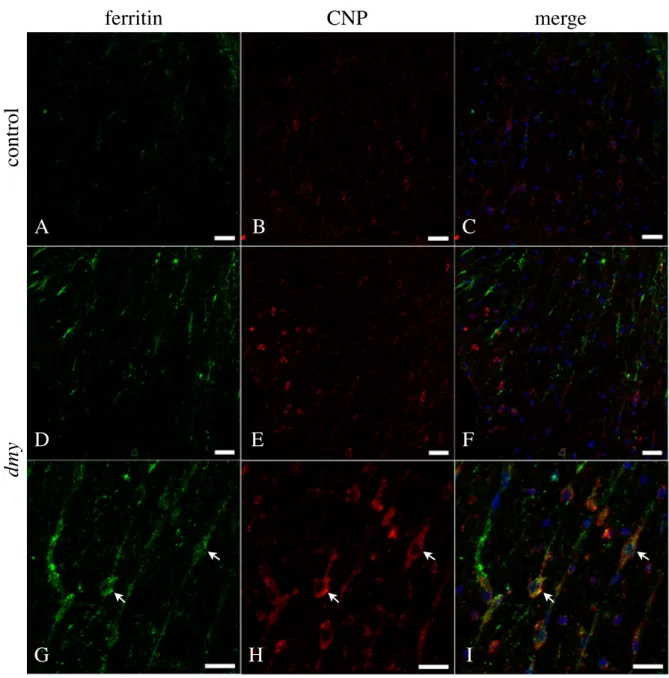
Chapter 3 Iron metabolism in the two different myelin mutant rats... 57 ...
Introduction 58
...
Materials and Methods 59
... Results 61 ... Discussion 63 ... Summary 66 ... Table 4 67 ... Figure Legends 68 ... Plate V 71 Conclusions... 76 References... 78 Acknowledgements... 97
Preface
The rapid saltatory conduction along vertebrate axons requires the formation of
insulating myelin sheaths, which are formed by oligodendrocytes in the central nervous
system (CNS) and Schwann cells in the peripheral nervous system (PNS). Myelin diseases are
a heterogenous group of neurological diseases in which myelin structure is primarily affected.
Primary diseases of CNS myelin in humans are devided into the two major types: 1)
demyelination (myelin breakdown after it is formed) such as multiple sclerosis (MS) and
acute disseminated encephalomyelitis, 2) dysmyelination/hypomyelination (failure to form
normal amount of myelin during development) such as adrenoleukodystrophy and
Pelizaeus-Merzbacher disease (Powers, 2004). The causes of myelin diseases are various (e.g.,
hereditary, immune-mediated, infectious and metabolic) and there are many clinical
phenotypes even in one myelin disease. Because of this heterogeneity, the pathogenesis of
human myelin diseases is still obscure. Therefore, suitable animal models are needed to
elucidate the mechanism of disease processes.
In experimental animals such as mice and rats, various mutant models with CNS
myelin disorders have been reported ("myelin mutants"). These myelin mutants are largely
devided into two types according to their mutated genes. The first are mutants deficient in
myelin-constituting proteins, such as the shiverer (shi) mouse, jimpy (jp) mouse, myelin
deficient (md) rat and Long Evans shaker (les) rat (Baumann and Pham-Dinh, 2001). Myelin
abnormalities of these mutants are mainly due to structural defects of myelin sheaths. The
second are mutants with defects in proteins that are not structural components of myelin but
responsible for the formation and/or maintenance of myelin, such as the tremor rat (Kitada et
al., 2000) and taiep (tremor, ataxia, immobility, epilepsy and paralysis) rat (Li et al., 2003;
pathogenesis of myelin diseases in this type of mutants. Myelin mutants can be suitable
models for studying the cellular and molecular mechanisms of myelination and maintenance
of myelin.
Homeostasis of the central nervous system is maintained by a balanced interaction
between glial cells, which constitute the great majority of cells in the nervous system (65%
and 90% in rodents and humans, respectively) (Baumann and Pham-Dinh, 2001).
Oligodendrocytes are the myelin-forming cells of the CNS and thus seem to be primarily
affected in the myelin diseases. Extensive loss of oligodendrocytes is a hallmark of MS, an
autoimmune demyelinating disease in humans (Lassmann, 2004a), whereas it is much less
severe in experimental autoimmune encephalomyelitis (EAE), an animal model of MS
(Lassmann, 2004b). Astrocytes are the supporting cells in the CNS and mainly involved in the
maintenance of tissue homeostasis. In MS and EAE, astrocytes play a protective role in
demyelination by releasing anti-inflammatory cytokines such as interleukin (IL)-4 and IL-10,
and cytokines that promote oligodendrogenesis such as insulin-like growth factor-1 and
leukemia inhibitory factor, whereas they also play a destructive role by producing
inflammatory cytokines such as IL-12 and IL-23, and by forming the glial scar, a physical
barrier for myelin repair (Nair et al., 2008). Microglia are the CNS-resident macrophages and
involved in immune functions and the clearance of tissue debris. Activated microglia in
demyelinating lesions release proinflammatory cytokines such as tumor necrosis factor-α
(TNF-α) and IL-1β, and reactive oxygen and nitrogen species, all of which are detrimental to
myelin (Jack et al., 2005; Raivich and Banati, 2004). These findings suggest that not only
oligodendrocytes but also astrocytes and microglia play crucial roles in the pathogenesis of
myelin diseases.
It is well known that iron is enriched in the CNS and is essential for its normal
oligodendrocytes are the principal cells that contain a high amount of iron in the CNS,
suggesting functional relationship between iron accumulation and myelin production
(MacKenzie et al., 2008; Meguro et al., 2008; Todorich et al., 2009). In humans, iron
deficiency is the most common nutritional disorder in the world, and the affected children
suffer from neurological deficits in addition to anemia (Beard, 2008; Lozoff et al., 2006). The
most common neurological signs are poor school performance, decreased cognitive abilities
and behavior problems, which are likely to be consistent with alterations in myelination and
neurotransmitter functions. Studies using rat models of iron deficiency demonstrated that
restriction of dietary iron during gestation and early postnatal period results in
hypomyelination with a decrease in myelin proteins, lipids and cholesterol in the CNS
(Todorich et al., 2009). These findings suggest that iron acquisition during prenatal and
postnatal period is necessary for normal CNS myelination.
Besides iron deficiency, abnormal accumulation of iron is considered to be involved in
the pathogenesis of myelin diseases. Histochemical studies revealed that abnormal iron
deposits are observed in infiltrating macrophages/reactive microglia, axons/neurons and
oligodendrocytes in MS patients (LeVine, 1997; Levine and Chakrabarty, 2004). In mice with
EAE, the iron deposits are detected in macrophages and astrocytes (Forge et al., 1998). These
data suggest that glial cells may participate in altered iron metabolism of demyelinating
diseases.
The myelin vacuolation (mv) rat is an autosomal recessive mutant characterized by
hypomyelination and vacuole formation in the myelin throughout the CNS, which is caused
by a null mutation in the attractin (Atrn) gene on rat chromosome 3 (Kuwamura et al., 2002).
ATRN was first discovered as a secreted serum glycoprotein, released by activated T cells
with a role in chemotaxis regulation in humans (Duke-Cohan et al., 1995, 1998). The Atrn
(Kuramoto et al., 2001; Tang et al., 2000), whereas only transmembrane form has been found
in mice (Gunn et al., 1999). Atrn mRNA is widely expressed in various organs (Gunn et al.,
1999; Tang et al., 2000), and ATRN protein has multiple biological functions including
immune cell interaction, hair pigmentation, energy homeostasis, and CNS myelination
(Dinulescu et al., 1998; Duke-Cohan et al., 2000; He et al., 2001; Kuramoto et al., 2001;
Nagle et al., 1999). Mutations in the Atrn gene have been reported in mice (mahogany,
Atrnmg), rats (zitter, Atrnzi; myelin vacuolation, Atrnmv) and Syrian hamsters (black tremor,
Atrnbt) (Gunn et al., 1999; Kuramoto et al., 2001, 2002); all of these mutations result in
abnormal hair pigmentation and vacuole formation in the CNS (Gunn et al., 2001; Nunoya et
al., 1985; Rehm et al., 1982). However, the detailed glial pathology during development of
myelin disorders in the Atrn mutant animals has not been investigated to date.
The demyelination (dmy) rat is an autosomal recessive mutant which exhibits hind
limb ataxia and severe myelin breakdown in the CNS during the late stage of postnatal
myelination (Kuwamura et al., 2004). Genetic analysis revealed that the dmy locus is located
on rat chromosome 17, homologous to human chromosome 6 and mouse chromosome 13
(Kuramoto et al., 1996). To date, no myelin-related gene has been reported in the homologous
regions of humans or mice, suggesting that the dmy rat is a novel myelin mutant.
Immunohistochemical and morphometrical studies have been reported in the dmy rat
(Kuwamura et al., 2004), however, the exact pathogenesis of demyelination remains to be
elucidated.
In this thesis, the cellular and molecular changes of glial cells during development of
myelin disease were investigated in the hypomyelinating mutant mv and demyelinating
mutant dmy rats. In the first chapter, for understanding the pathogenesis of hypomyelination in
the mv rat, glial cell kinetics were investigated in the mv rat, and the Atrn-expressing cells in
microglia-related factors and myelin-constituting proteins were examined to determine the
relationship between microglial activation and myelin alterations in the gray matter of the mv
rat. In the last chapter (chapter 3), changes in iron metabolism and expression of iron
regulatory proteins in the mv and dmy rats were investigated for understanding the role of iron
Chapter 1
Glial pathology during development of hypomyelination
in the myelin vacuolation (mv) rat
Section 1-1. Analysis of glial pathology in the spinal cord of the mv rat
Introduction
It is well known that glial abnormalities are critically involved in the pathogenesis of
myelin mutant animals. Premature cell death of oligodendrocytes accounts for a severe CNS
myelin deficit in the jimpy mouse, a myelin mutant with a X-linked mutation in the
proteolipid protein (PLP) /DM20 gene (Skoff, 1976; Knapp et al., 1986). Unlike the jimpy,
functional abnormalities of oligodendrocytes are responsible for the myelin alterations in the
les and taiep rats, which are characterized by dysmyelination with an insertional mutation in
the myelin basic protein (MBP) gene (Delaney et al., 1995; Kwiecien et al., 1998; O’Connor
et al., 1999), and by early hypomyelination and subsequent demyelination associated with
microtubule alterations in oligodendrocytes (Duncan et al., 1992; Holmgren et al., 1989;
Lunn et al., 1997; Song et al., 1999), respectively.
Besides oligodendrocyte abnormalities, changes in astrocytes and microglia are
commonly observed in the myelin mutants. The jimpy mutant mice show a striking
hypertrophy of astrocytes (Omlin and Anders, 1983; Skoff, 1976) and an intense microglial
reaction that is more pronounced in the white matter than in the gray matter and is related to
apoptotic oligodendrocytes (Vela et al., 1995 and 1996). In the les rat, astrogliosis and
microgliosis are localized to the white matter (Kwiecien et al., 1998; Zhang et al., 2001). In
the taiep rat, progressive astrogliosis is developed in the brain lesions where hypomyelination
is more prominent (Leon Chavez et al., 2001), and the number of CD11b-positive microglia/
macrophages is increased in the cerebellar white matter (Leon Chavez et al., 2006). The
pattern and degree of glial abnormalities vary in each mutant, but are closely associated with
In this section, glial changes during development of hypomyelination in the spinal
cord of the mv rat were investigated.
Materials and Methods
Animals
The mv rats were bred and maintained in a mixed background of Sprague-Dawley
and Donryu. Homozygous mv (mv/mv) rats were obtained by mating heterozygous (mv/+)
females with heterozygous males. The genotype of each rat was determined by polymerase
chain reaction (PCR) using diagnostic primer sets (Tokuda et al., 2004). Rats were maintained
in a room with controlled temperature and 12-hour light-dark cycle. Food and water were
provided ad libitum. Rats were handled according to the Guidelines for Animal
Experimentation of Osaka Prefecture University.
Histology and Immunohistochemistry
For histological analysis, control (+/+ or mv/+) and mv homozygous (mv/mv) rats at
2, 4, 6, and 10 weeks of age were deeply anesthetized and perfused transcardially with 4%
paraformaldehyde (PFA) in 0.1 M phosphate buffer (PB; pH 7.4). Removed lumbar spinal
cords were stored in 2% PFA and 2.5% glutaraldehyde in 0.1 M PB, postfixed with 1% osmic
acid for 2 h, and embedded in epoxy resin. One μm semi-thin sections were cut and stained
with toluidine blue. Ultrathin sections were stained with uranyl acetate and lead citrate and
examined in a Hitachi H-7500 electron microscope (Hitachi, Japan). For
transcardially with 4% PFA in 0.1 M PB. Lumbar spinal cords were then removed and
postfixed in 4% PFA in 0.1 M PB at 4°C overnight. After 2- or 3-day treatment with 30%
sucrose in phosphate-buffered saline (PBS) at 4°C, spinal cords were transversely sliced and
frozen at –80°C. Ten μm frozen sections were cut using a cryostat. The following primary
antibodies were used: mouse anti-2', 3'-cyclic nucleotide-3'-phosphodiesterase (CNP) for
oligodendrocytes (1:1,000; Sigma, USA), rabbit anti-glial fibrillary acidic protein (GFAP;
1:1,000; Dako, Denmark) for astrocytes, mouse anti-CD11b (clone OX-42; 1:200; Millipore,
USA) for microglia/macrophages, and mouse anti-MHC class II (rat RT-1B; clone OX-6;
1:100; Serotec, UK) for activated microglia/macrophages. Horseradish peroxidase-conjugated
secondary antibody (Histofine simplestain MAX PO; Nichirei, Japan) was used for GFAP,
CD11b and MHC class II immunohistochemistry, while the avidin-biotinylated enzyme
complex Kit (Vector Laboratories, USA) was used for CNP immunohistochemistry. Signals
were visualized with 3,3-diaminobenzidine (DAB) substrate kit (Vector Laboratories).
Cell counts
The number of CNP-positive oligodendrocytes was counted in the whole dorsal
funiculus of 10-μm transverse sections of the lumbar spinal cord under microscopic
observation. Three different fields from three different animals were evaluated. The sections
were scanned by a digital camera (Coolpix990; Nikon, Japan), and the area of the dorsal
funiculus was measured using a software (WinRoof; Mitani Corporation, Japan). The data are
presented as the number of CNP-positive cells/mm2.
Data are presented as means ± SD. Statistical analysis was performed using one-way
analysis of variance followed by Tukey’s test or Scheffe’s test. A value of P less than 0.05 was
considered statistically significant.
Results
Myelin abnormalities increase within spinal cord white and gray matter during postnatal development
To understand the progression of myelin pathology in the mv rat, the temporal
changes in the myelin during postnatal development of the spinal cord were examined.
Compared with age-matched control rats (Fig. 1A-C), fewer myelinated axons and thinner
myelin sheaths were observed in the white matter of the mv rats (Fig. 1D-F). This
hypomyelination became more prominent with age. Also present in the mv rats were
variously-sized vacuoles and abnormal myelin sheaths (Fig. 1F, arrows), which were
frequently observed in the white matter from 6 weeks. In the gray matter, fewer myelinated
axons and vacuolated myelin sheaths (Fig. 2B, arrows) were observed in the mv rats from 6
weeks. In electron microscopy, naked and hypomyelinated axons were observed in the mv rats
(Fig. 3, arrows and arrowheads, respectively), whereas no obvious ultrastructural abnormality
was found in oligodendrocytes (Fig. 3).
Myelin pathology is associated with increased astrogliosis but no changes in oligodendrocyte numbers or morphology, while microglial activation is mainly confined to the gray matter
with specific glial markers. There were no apparent changes in the morphology of
CNP-positive oligodendrocytes between control and mv rats at any of the ages examined (Fig. 4B,
C), neither did the number of CNP-positive oligodendrocytes differ significantly (Fig. 4A).
Compared with control rats (Fig. 5A, B), GFAP immunoreactivity in the white matter of the
spinal cord was increased in the mv rats from 2 weeks (Fig. 5E) and was more pronounced at
8 weeks (Fig. 5F). In the gray matter, the morphology of astrocytes was almost similar both in
control and mv rats at 2 weeks (Fig. 5C, G), but at 8 weeks the astrocytes in the mv rats
showed hypertrophy of their processes, a common feature of activated astrocytes (Fig. 5H,
arrows). Microglial morphology in the white matter of the mv rats was almost similar to
controls at all ages (Fig. 6A, D). In the gray matter, the morphology of microglia was not
prominently changed either in control and mv rats up to 4 weeks, but from 6 weeks the
microglia in the mv rats had an activated morphology, characterized by swollen cell bodies
and shortened processes (Fig. 6E). Furthermore, MHC class II immunohistochemistry
revealed marked microglial activation in the mv rats from 6 weeks, which was exclusively
found in the gray matter (Fig. 6F).
Discussion
In the mv rat, hypomyelination and abnormal myelination were observed in the white
matter of the spinal cord from 2 weeks, the severity of which progressed with age. Despite
progressive myelin pathology, the number and morphology of oligodendrocytes in the mv rat
were almost similar to those in controls at all ages examined. In the jimpy mouse,
oligodendrocytes proliferate but die rapidly through apoptosis (Knapp et al., 1986), which
may result from the accumulation of misfolded PLP in the endoplasmic reticulum (Gow et al.,
life and increase with age, whereas CNS myelin is gradually lost, indicating an inability of
oligodendrocytes to maintain myelin (Kwiecien et al., 1998). The les rat also has an abnormal
accumulation of vesicular organelles in the cytoplasm of oligodendrocytes, which may reflect
a failure of myelin production and formation. In the taiep rat, the myelin deficit has been
directly associated with microtubule alterations in oligodendrocytes (Song et al, 1999), and
with alterations in the expression and intracellular distribution of myelin gene products
(O’Connor et al., 2000). In the mv rat, no degenerating oligodendrocytes were seen under
light microscope, and no ultrastructural abnormality of oligodendrocytes was observed. The
results of this study show that oligodendrocytes in the mv rat do not undergo direct cell injury
or death, but may have some dysfunction in formation and maintenance of CNS myelin.
The results of this study showed increased GFAP immunoreactivity in the white
matter of the spinal cord in the mv rat from 2 weeks, which progressed with age. Hypertrophic
astrocytes also appeared in the gray matter of the mv rat from 6 weeks. The regional and
temporal patterns of astrocyte activation are consistent with those of myelin alterations in the
mv rat, and these astrocytic changes are commonly observed in myelin mutants. In the les rat,
the temporal pattern of astrocyte hypertrophy corresponds to that of dysmyelination, and the
distribution of reactive astrocytes is dominant in the white matter, indicating an astrocyte
response to the white matter lesions (Zhang et al., 2001). Similarly, the progressive
astrogliosis is considered as a reactive change in the taiep rat, since the regional and temporal
patterns correlate with the process of dysmyelination (Leon Chavez et al., 2001). Unlike the
les and taiep rats, astrocyte abnormalities in the jimpy mouse are observed not only during
myelination but also in early postnatal development, before mature oligodendrocytes appear,
supporting but not proving a causative effect of astrocytes on myelin deficit (Omlin and
Anders, 1983; Skoff, 1976). The results of this study demonstrate that the progressive
reactive changes of astrocytes to the myelin alterations.
CD11b and MHC class II immunohistochemistry demonstrated the presence of
numerous activated microglia in the gray matter of the spinal cord in the mv rat from 6 weeks.
The time course of microglial activation was consistent with the progression of myelin
pathology, whereas the distribution of activated microglia did not match with the myelin
lesions that were predominant in the white matter. This distribution was clearly different from
other myelin mutants such as the jimpy mouse (Vela et al., 1995 and 1996), les rat (Zhang et
al., 2001) and taiep rat (Leon Chavez et al., 2006), where the microglial reaction is limited to
the white matter. Despite the numerous activated microglia in the gray matter, there were no
phagocytic microglia or degenerating neurons in the mv rat, indicating that the microglial
activation in this mutant is unlikely to be a mere reaction to myelin disruption or neural
degeneration. Further study is required to determine why microglial activation is limited to
Summary
Glial pathology during the development of hypomyelination in the spinal cord of the
mv rat was investigated. The severity and extent of myelin disorder increased during postnatal
myelination in the mv rat, whereas no abnormality was found in the number or morphology of
oligodendrocytes at any of the ages examined. Coincident with the myelin abnormalities,
there was progressive astrogliosis both in the white and gray matter of the mv rat from 2
weeks. Marked microglial activation was mainly confined to the gray matter of the mv rat
from 6 weeks, which was consistent temporally but not spatially with the morphological
abnormalities of myelin sheaths. The glial abnormalities in the mv rat were closely associated
with the myelin lesions, suggesting that, like other myelin mutants, glial cells are involved in
Figure Legends
Fig. 1
Toluidine blue-stained sections of the white matter (ventral funiculus) of the lumbar spinal
cord in control (A-C) and mv (D-F) rats. Compared with control rats (A-C), fewer myelinated
axons and thinner myelin sheaths are found in the mv rats (D-F). From 6 weeks, abnormal
myelin sheaths are frequently observed in the mv rats (F, arrows). Bar: 20 μm.
Fig. 2
Toluidine blue-stained sections of the gray matter of the lumbar spinal cord in control (A) and
mv (B) rats at 6 weeks. The number of myelinated fibers are markedly decreased in the mv
rats (B) compared with control rats (A). Vacuolated myelin sheaths are frequently observed in
the mv rats (B, arrows). Bar: 20 μm.
Fig. 3
Electron micrograph of the white matter of the lumbar spinal cord of the mv rat at 6 weeks.
There are many unmyelinated (arrows) or thinly-myelinated axons (arrowheads), whereas
no ultrastructural abnormality is found in oligodendrocytes. Bar: 1.4 μm.
Fig. 4
The graph represents the number of CNP-positive oligodendrocytes in the dorsal funiculus of
the lumbar spinal cord (A). Data are presented as the number of positive cells/mm2 (n=3 in
each group). CNP immunohistochemistry in the white matter of the lumbar spinal cord in
control (B) and mv rats (C). There are no significant differences in the cell number or
Fig. 5
GFAP immunohistochemistry in the lumbar spinal cord of control (A-D) and mv (E-H) rats.
Compared with control rats (A, B), immunoreactivity for GFAP is increased in the white
matter of mv rats from 2 weeks (E, F). Hypertrophic astrocytes (arrows) are found in the gray
matter of the mv rats at 8 weeks (H). Bar: 40 μm.
Fig. 6
Immunohistochemistry for CD11b (A, B, D, E) and MHC class II (C, F) in the lumbar spinal
cord of control (A-C) and mv (D-F) rats at 6 weeks. There are no apparent morphological
changes of microglia in the white matter of the mv rats (D), whereas there are many activated
microglia with swollen cell bodies in the gray matter of the mv rats (E). MHC class II
immunohistochemistry shows that microglial activation is confined to the gray matter of the
Plate I
Fig. 1
F
E
D
C
B
2 wks
4 wks
6 wks
A
control
mv
Fig. 2
A
B
Fig. 3
Fig. 4
A
Fig. 5
A
G
F
E
C
B
D
H
control
mv
2 wks
8 wks
2 wks
8 wks
white matter
gray matter
Fig. 6
F
C
E
D
B
white matter
MHC class II
A
control
mv
gray matter
CD11b
Section 1-2. Identification of attractin-expressing cells
in the normal spinal cord
Introduction
It was demonstrated in the previous section that the Atrn defect does not affect
oligodendrocyte numbers and morphology, however results in progressive myelin vacuolation
in the spinal cord, implying a functional abnormality of oligodendrocytes in myelin
formation.
A transgenic rescue experiment revealed that membrane-type but not secreted-type
Atrn complemented neurological alteration in the Atrn mutant zitter rat, indicating a critical
role of membrane-type Atrn in normal myelination and myelin maintenance in the CNS
(Kuramoto et al., 2001). An in situ hybridization analysis demonstrated a broad distribution of
Atrn mRNA throughout the CNS of normal adult rats (Lu et al., 1999). However, in that study
the cell type expressing Atrn was not identified, and the distribution of Atrn was not examined
in young rats. Since myelin abnormalities in the mv rat are observed from 2 weeks of age, an
early stage of postnatal myelination, analysis for the Atrn expression should be performed
using rats at early myelination.
Therefore in this section, the expression of Atrn mRNA during early postnatal
development of normal rat spinal cord was investigated for exploring the role of Atrn in
Materials and Methods
Preparation of digoxigenin (DIG)-labeled riboprobe
For riboprobe preparation, a 788-bp fragment of exon 29 of rat membrane-type Atrn
(GenBank accession no. AB038387) was amplified by PCR (sense primer, 5’-GGC TCC CAC
CTA CCT GTT TAT G-3’, nucleotide position 6454-6475; antisense primer, 5’-TTT GCC
TGT TCG TGC TGT G -3’, nucleotide position 7223-7241). The PCR product was subcloned
into pGEM T-easy vector (Promega, USA). DIG-labeled antisense or sense riboprobe was
synthesized with T7 or SP6 RNA polymerase (Roche, Switzerland), respectively.
In situ hybridization for Atrn
Lumbar spinal cords of wild-type and mv rats at 2 and 4 weeks were sampled as
previously described (see Section 1-1, Materials and Methods). Ten-μm frozen sections were
cut using a cryostat. Before hybridization, sections were pretreated as follows: 1) 4% PFA in
PBS for 15 min, 2) 10 μg/ml proteinase K (Invitrogen, USA) at 37°C for 12 min, 3) 0.2 M
HCl for 10 min, 4) 0.25% acetic anhydride in 0.2 M triethanolamine (pH 8.0) for 10 min.
DIG-labeled riboprobes were diluted in hybridization buffer (50% formamide, 10 mM
Tris-HCl, pH 8.0, 200 μg/ml yeast tRNA, 10% dextran sulfate, 1×Denhardt’s solution, 600 mM
NaCl, 0.25% SDS, 1 mM EDTA), and were placed on each slide. Sections were then
coverslipped and incubated at 65°C for 16 h. After hybridization, they were rinsed in
2×sodium saline citrate (SSC) containing 50% formamide at 65°C for 30 min, treated with 20
μg/ml RNase A (Roche) at 37°C for 30 min, and rinsed in 2×SSC, 0.2×SSC, and 0.1×SSC
phosphatase-conjugated anti-DIG antibody (Roche) at 4°C overnight and were visualized with
5-bromo-4-chloro-3-indolyl phosphate (BCIP) and nitro blue tetrazolium chloride (NBT) substrates
(Roche).
Identification of Atrn-expressing cells
For double labeling, immunohistochemistry was performed following in situ
hybridization. RNA hybrids were visualized with 2-hydroxy-3-naphthoic acid-2-phenylanilide
phosphate (HNPP)/Fast Red TR (Roche). Sections were then incubated with mouse anti-CNP
(Millipore) and rabbit anti-GFAP (Dako) antibody and reacted with fluorescein isothiocyanate
(FITC)-conjugated secondary antibodies (Jackson Immunoresearch, USA). Signals were
detected with a confocal imaging system (C1Si; Nikon) .
Results
Atrn mRNA is mainly expressed by CNP-positive oligodendrocytes in the white matter
To explore the role of Atrn in the normal rat spinal cord, Atrn-expressing cells was
investigated by in situ hybridization using DIG-labeled riboprobe specific to the
membrane-type Atrn mRNA. Atrn antisense probe labeled a large number of positive cells both in the
white and gray matter of wild-type spinal cord at 2 weeks (Fig. 7A, D), whereas there was no
labeling with the sense probe (Fig. 7B, E). No specific signal was detected in the mv rats with
the antisense probe (Fig. 7C, F). For identification of Atrn-expressing cells, double labeling
by in situ hybridization and immunohistochemistry was performed. Atrn mRNA was
arrowheads). In the gray matter, GFAP-positive astrocytes were occasionally positive for Atrn
(Fig. 8D-F; arrow). Some neurons were also positive for Atrn in the gray matter (Data not
shown).
Discussion
The results of this study revealed that Atrn mRNA is mainly expressed by oligodendrocytes in the spinal cord white matter of wild-type rats at early postnatal ages,
which suggests that the membrane-type Atrn is involved in oligodendrocyte function.
Recently, a biochemical study showed the progressive loss of lipid-raft domains and reduction
in cellular cholesterol in splenocytes of the Atrn-deficient mouse, proposing that the
juvenile-onset hypomyelination and neurodegeneration represent a defect in attractin-mediated
raft-dependent myelin biogenesis (Azouz et al., 2007). Lipid rafts are detergent-insoluble
glycolipid-enriched membrane microdomains that participate in protein sorting and trafficking
(Simons and Ikonen, 1997) and may take a critical role in forming the myelin membrane
(Simons et al., 2000). Together with these findings, the results of this study show that Atrn
may be directly involved in the function of oligodendrocytes in postnatal myelination.
Recently, Nakadate et al. (2008) investigated ATRN expression in the mouse and rat
CNS at 8 weeks using a specific antibody against the membrane-type ATRN. ATRN is widely
expressed by neurons, microglia, astrocytes and oligodendrocytes, and localized in the
cytoplasmic membrane of Golgi apparatus, endoplasmic reticulum and mitochondria of
neurons and glial cells. The broad cellular and subcellular localization of ATRN suggests that
ATRN may serve multiple functions in the CNS. Further study is needed to determine the role
of ATRN in CNS myelination and the relationship between oligodendrocytes and other cell
Summary
The expression of Atrn mRNA in the spinal cord of normal rats at early postnatal ages
was investigated. Atrn expression was widely distributed both in the white and gray matter of
the spinal cord in wild-type rats, whereas no signal was detected in mv rats. In situ
hybridization combined with immunohistochemistry revealed that Atrn mRNA is mainly
expressed by mature oligodendrocytes in the white matter, and also by astrocytes and neurons
in the gray matter of normal spinal cord. These data suggest an important role of Atrn in
Figure Legends
Fig. 7
In situ hybridization for Atrn mRNA in the lumbar spinal cord of wild-type (A, B, D, E) and mv (C, F) rats. A large number of Atrn-positive cells are labeled with the antisense probe both
in the white (A) and gray matter (D) of wild-type rats, whereas there is no labeling with the
sense probe (B, E). No specific signal is detected in the mv rats with the antisense probe (C,
F). Bar: 50 μm
Fig. 8
Atrn in situ hybridization combined with immunohistochemistry for CNP (A-C) and GFAP
(D-F) in the lumbar spinal cord of wild-type rats. In the white matter, Atrn-positive cells are
mainly CNP-positive oligodendrocytes (A-C, arrowheads). In the gray matter, astrocytes are
occasionally positive for Atrn (D-F, arrowheads). Green; immunohistochemistry (FITC),
Plate II
Fig. 7
gray matter
white matter
antisense
sense
antisense
wild type
mv
F
C
E
D
B
A
Fig. 8
white matter
gray matter
CNP
/
Atrn
/
DAPI
GFAP
/
Atrn
/
DAPI
Atrn
CNP
Atrn
GFAP
F
C
E
D
B
A
Chapter 2
Microglial activation and gray matter lesions
in the mv rat
Section 2-1. Analysis of expression patterns of microglia-related factors
in the mv rat
Introduction
It was demonstrated in section 1-1 that microglial activation is confined to the gray
matter whereas morphological abnormalities of myelin sheaths were more apparent in the
white matter of the mv rat. These results raise the possibility that the microglial activation is
more than a simple reaction to the myelin destruction in the mv rat.
Microglia/macrophages are involved in the pathology of multiple sclerosis (MS) and
experimental autoimmune encephalomyelitis (EAE) by the production of various kinds of
cytokines. Activated microglia and infiltrating macrophages in demyelinating lesions of MS
and EAE show a strong upregulation of TNF-α, IL-1β and IL-6 (Benveniste, 1997; Raivich
and Banati, 2004). These cytokines have been proved to have detrimental effects on CNS
demyelination, one of which is the production of reactive oxygen and nitrogen species (Jack
et al., 2005; Raivich and Banati, 2004). Inducible nitrogen oxide synthase (iNOS) is
upregulated in astrocytes and macrophages in MS patients (Jack et al., 2007), and its product
nitric oxide (NO) is thought to be toxic to myelin (Raivich and Banati, 2004; Smith and
Lassmann, 2002). Transforming growth factor-β1 (TGF-β1) plays an important role as an
immunosuppressive cytokine in MS and EAE (Prud'homme and Piccirillo, 2000) and is
upregulated in infiltrating macrophages, activated microglia and reactive astrocytes in MS (De
In this section, the expression of several cytokines which are known to be related with
microglial activation in human and animal myelin diseases, was investigated to understand
the role of activated microglia in the gray matter of the mv rat.
Materials and Methods
Reverse transcription PCR (RT-PCR) for microglia-related factors
Cervical spinal cords of wild-type and mv rats at 4 and 6 weeks of age were removed
and dissected into the white and gray matter. Total RNA was isolated using SV Total RNA
isolation system (Promega) according to the manufacturer’s instructions. One μg of total
RNA was transcribed with Superscript II reverse transcriptase using random hexamers
(Invitrogen). To examine gene expression of TNF-α, IL-1β, and IL-6, first-strand cDNA was
amplified by a thermal cycler (PC 707; Astec, Japan) with GoTaq DNA polymerase
(Promega). To determine relative expression levels of iNOS and TGF-β1 genes, quantitative
real-time PCR was performed with a SYBR Green PCR master mix (Toyobo, Japan) in a
Linegene system (Bioflux, Japan). Details of specific primers are listed in Table 1. β-Actin
was used as an internal control. The cycling condition for real-time PCR was as follows: 1
cycle of 95°C for 1 min, followed by 45 cycles of 95°C for 15 sec, 60°C for 15 sec, and 72°C
for 30 sec. Relative expression levels were calculated based on threshold cycle (Ct) value
(comparative Ct method).
Western blot for iNOS protein
homogenized in a cell lysis reagent (CelLytic MT; Sigma). After centrifugation at 13,000 g for
10 min, protein concentrations were determined by the Bradford Protein Assay (BioRad,
USA). Ten μg of total proteins were separated on 7.5% polyacrylamide gels and transfered to
polyvinylidene difluoride membranes (BioRad). Membranes were incubated with mouse
anti-iNOS antibody as described elsewhere (Nakamura et al., 2006) and mouse anti-β-actin
antibody (Sigma) at 4°C overnight, and were treated with Histofine simplestain MAX PO for
30 min. Signals were visualized with ECL reagent (GE Healthcare, USA) and quantified with
a luminescent image analyzer (LAS-3000; Fujifilm, Japan). β-Actin was used as an internal
control.
Immunohistochemistry for iNOS
Lumbar spinal cords of control and mv rats at 6 weeks were sampled as previously
described (see Section 1-1, Materials and Methods). Ten-μm frozen sections were cut using a
cryostat. Sections were incubated with 10% normal goat serum in PBS for 30 min, treated
with mouse ant-iNOS antibody (Nakamura et al., 2006) at 4°C overnight, and incubated with
Histofine simplestain MAX PO for 45 min. Signals were visualized with DAB.
Statistical analysis
Data are presented as means ± standard deviation. Statistical analysis was performed
using one-way analysis of variance followed by Tukey’s test. A value of P less than 0.05 was
Results
Microglial activation is associated with increases in TGF-β1 and iNOS expression
To gain further insights into the role of activated microglia in the mv rat, the
expression of several factors associated with microglial activation was examined. Expression
levels of TGF-β1 mRNA were significantly increased in the gray matter of the mv rats
compared with wild-type rats at 6 weeks (Fig. 9A). Expression levels of iNOS mRNA were
not significantly changed between wild-type and mv rats at either age (Fig. 9B). There were
no obvious changes in the expression of TNF-α, IL-1β, and IL-6 mRNA between wild-type
and mv rats (data not shown). Western blot analysis demonstrated a significant increase in the
expression of iNOS protein in the spinal cord of the mv rats at 10 weeks (Fig. 10A).
Immunoreactivity for iNOS was weakly detected in astrocytes in the white matter of control
rats at 6 weeks (Fig. 10B, D), whereas it was markedly increased in activated astrocytes of
both the white and gray matter of mv rats (Fig. 10C, E).
Discussion
The results of this study demonstrated increased expression of TGF-β1 mRNA in the
spinal cord gray matter of the mv rat, which is coincident with the microglial activation.
TGF-β1 is normally present at low levels in the normal CNS tissues, while it is highly upregulated
in various cell types, such as activated microglia and astrocytes, and neurons in many
neurodegenerative diseases (Finch et al., 1993; Kiefer et al., 1998; Vivien and Ali, 2006; Zhu
et al., 2000). An in vivo analysis of TGF-β1-deficient mice revealed that lack of TGF-β1
microglial activation (Brionne et al., 2003). This finding is further supported by in vitro
studies demonstrating that TGF-β1 regulates microglial activity by inhibiting iNOS
expression (Lieb et al., 2003; Vincent et al., 1997) and NO production (Herrena-Molina and
von Bernhardi, 2005; Lieb et al., 2003). These data suggest that TGF-β1 may play a key role
in microglial activation in the mv rat.
ATRN is expressed on the cell surface of peripheral blood monocytes and involved in
their function in cell adhesion and release of cytokines including TGF-β1 (Wrenger et al.,
2006). Since microglia are believed to be derived from bone marrow during embryogenesis, it
is likely that ATRN may also be involved in microglial function, and that the lack of ATRN
may affect the function of microglia in the mv rat. Recent studies demonstrated abnormal
recruitment of macrophages/microglia during very early postnatal development and
subsequent activation of microglia in the brain of the zitter rat, suggesting that macrophage/
microglial lineage may contribute to myelin alterations in the zitter rat (Kadowaki et al.,
2007; Sakakibara et al., 2008). Further studies on the cytokine network should be needed to
understand the molecular mechanism and roles of microglial activation in the Atrn-mutant
animals.
The results of this study showed that expression of iNOS protein is significantly
increased in activated astrocytes both in the white and gray matter of the mv rat. However, the
author failed to detect a difference in iNOS mRNA levels between wild-type and mv rats. This
may be due to the sensitivity of the real-time PCR method, since the expression levels of
iNOS in the spinal cord of both groups were very low (more than 30 in Ct value). In the
demyelinating lesions of MS and EAE, iNOS is induced mainly in reactive astrocytes and
microglia/macrophages (Hill et al., 2004; Jack et al., 2007; Liu et al., 2001; Raivich and
Banati, 2004). iNOS produces a large amount of NO, and NO and its reactant with
and Benjamins, 2006; Smith et al., 1999) and mediators of damage to myelin (Bizzozero et
al., 2004; Boullerne and Benjamins, 2006; Jack et al., 2007; Smith et al., 1999). The results of
this study suggest that the activated astrocytes in the mv rat may be involved in the
Summary
The expression of microglia-related cytokines in the spinal cord of the mv rat was
investigated to explore the role of activated microglia in the gray matter. Increased expression
of TGF-β1 was closely associated with the microglial activation, which may indicate a key
role of this cytokine in the regulation of microglial activation in the mv rat. iNOS expression
was markedly increased in activated astrocytes both in the white and gray matter of the mv
rat, suggesting that the activated astrocytes in the mv rat may be involved in the progression
Table 1. Primers for RT-PCR analysis
Gene Primer sequence Accession number
TNF-α sense: 5’- TGTCTACTGAACTTCGGGGTG -3’ X66539
antisense: 5’-GAGGCTGACTTTCTCCTGGTA-3’
IL-1β sense: 5’-AAGCCTCGTGCTGTCGGACCC-3’ M15131
antisense: 5’-TCCAGCTGCAGGGTGGGTGTG-3’
IL-6 sense: 5’-ATGAAGTTTCTCTCCGCA-3’ M26744
antisense: 5’-GGGGTAGGAAGGACTATT-3’
iNOS sense: 5’-CCCTAAGAGTCACAAGCATCAAAAT-3’ D44591
antisense: 5’-GGTTCCTGTTGTTTCTATTTCCTTTGTTAC-3’
TGF-β1 sense: 5’-CTTCAGCTCCACAGAGAAGAACTGC-3’ X52498
antisense: 5’-CACGATCATGTTGGACAACTGCTCC-3’
β-Actin sense: 5’-TAAAGACCTCTATGCCAACAC-3’ BC063166
antisense: 5’-CTCCTGCTTGCTGATCCACAT-3’
TNF-α; tumor necrosis factor, IL-1β; interleukin-1β, IL-6; interleukin-6, iNOS; inducible nitric oxide synthase, TGF-β1; transforming growth factor-β1
Figure Legends
Fig. 9
Expression levels of TGF-β1 (A) and iNOS (B) mRNA in the cervical spinal cord of
wild-type (WT; white column) and mv rats (black column) at 4 and 6 weeks. Data are presented as
the mean ratio of target to reference gene (n=3 in each group). β-Actin is used as an internal
control. Expression levels of TGF-β1 mRNA are significantly increased in the gray matter of
mv rats at 6 weeks compared with control rats (A). Expression levels of iNOS mRNA do not
differ significantly between wild-type and mv rats at either age (B). *P<0.05 by Tukey's test.
Fig. 10
Expression levels of iNOS protein in the thoracic spinal cord of wild-type (WT; white
column) and mv rats (black column) at 10 weeks (A). Data are presented as the mean ratio of
iNOS to β-actin levels (n=3 in each group). β-Actin is used as an internal control. Expression
levels of iNOS protein are significantly increased in the mv rats (A). Immunohistochemistry
for iNOS in the lumbar spinal cord of control (B, D) and mv rats (C, E). Compared with
control rats (B, D), iNOS immunoreactivity is markedly increased in activated astrocytes of
Plate III
Fig. 9
gray matter white matter*
TGF-β1
iNOS
gray matter white matterB
A
0 1 2 3 4 wks 6 wks 4 wks 6 wks relative expression WT mv 0 0.5 1 1.5 2 2.5 4 wks 6 wks 4 wks 6 wks relative expression WT mvFig. 10
0 0.5 1 1.5 2 2.5 cont, 10 wks mv, 10 wks relative expression iNOS β-actin*
A
B
E
D
C
control mv white matter gray matterSection 2-2. Analysis of expression patterns of myelin proteins in the mv rat
Introduction
Myelin proteins are specific components of myelin and oligodendrocytes and are
involved in formation and maintenance of myelin. Proteolipid protein (PLP) and its splice
variant DM20 are the most abundant proteins in CNS myelin, which constitute nearly 50% of
myelin proteins in the CNS but less than 0.5% in the PNS (Baumann and Pham-Dinh, 2001;
Garbay et al., 2000). In the CNS, PLP is localized in compact myelin (Schwob et al., 1985)
and subserves a structural function in maintaining the compaction of the myelin sheath (Greer
and Lees, 2002 ; Klugmann et al., 1997). The second most abundant protein in the CNS is
myelin basic protein (MBP), which comprises 30% and 5-15% of total myelin proteins in the
CNS and PNS, respectively (Boggs, 2006; Garbay et al., 2000). MBP is localized in major
dense line of both CNS and PNS myelin (Omlin et al., 1982) and is essential for myelin
compaction in the CNS (Boggs., 2006; Kwiecien et al., 1998). 2', 3'-Cyclic
nucleotide-3'-phosphodiesterase (CNP) represents 4% of CNS myelin proteins and less than 0.5% of PNS
myelin proteins (Braun et al., 2004; Garbay et al., 2000). CNP is expressed by
oligodendrocyte lineage from an early stage of CNS myelination, and is localized in
non-compact myelin (Braun et al., 1988; Trapp et al., 1988). Myelin oligodendrocyte glycoprotein
(MOG) is a quantitatively minor component of myelin, which is specific for the CNS and is
selectively localized at the outermost surface of the myelin sheath (Brunner et al., 1989; Johns
and Bernard, 1999). This protein is well known as an important candidate autoantigen in MS
and to induce EAE (Iglesias et al., 2001).
Recent immunohistochemical studies for myelin proteins have revealed extensive gray
Stadelmann et al., 2008). Gray matter lesions in MS patients are characterized by less
extensive inflammation, less gliosis, and more efficient myelin repair than the white matter,
leading to the proposal that pathogenesis is different between the white and gray matter. In the
mv rat, marked microglial activation is found exclusively in the gray matter of the spinal cord
from 6 weeks, whereas morphological anbormalities of myelin sheaths are more apparent in
the white matter (see section 1-1). Kondo et al. reported slightly delayed and weakened
immunoreactivity of some CNS myelin proteins during early myelination in the Atrn-mutant
zitter rat; however, there is no description of myelinogenesis in the gray matter (Kondo et al.,
1992).
To explore the role of Atrn in myelin protein production and to gain further insights
into the relationship bewteen gray matter lesions and microglial activation, the expression
patterns of major CNS myelin proteins in the mv rat was investigated.
Materials and Methods
Immunohistochemistry
Lumbar spinal cords of control and mv rats at 2, 4 and 6 weeks were sampled as
previously described (see Section 1-1, Materials and Methods). Tissues were routinely
processed and embedded in paraffin. Five-μm sections were dewaxed, pretreated in a
microwave with distilled water for 10 min, and incubated in 3% hydrogen peroxide for 10
min. Sections were then incubated overnight at 4°C with the following antibodies: mouse
anti-PLP (1:1,000; Millipore), mouse anti-MBP (1:1,000; Millipore), and mouse anti-MOG (a
kind gift from Dr. Morgan B.P.; University of Wales College of Medicine, UK) (Piddlesden et
anti-mouse IgG antibody (Histofine Simplestain MAX PO; Nichirei) for 45 min. Signals were
visualized with a DAB substrate kit (Vector Laboratories). Sections were lightly
counterstained with hematoxylin.
Western blot analysis
Cervical spinal cords of SD (wild-type) and mv rats at 6 weeks of age were dissected
into the white and gray matter, and homogenized in a cell lysis reagent (CelLytic MT; Sigma).
After centrifugation at 13,000 g for 10 min, the pellets were lysed in SDS sample buffer
(0.0625 M Tris-HCl, pH6.8, 2% SDS, 15% glycerol and 5% 2-mercaptoethanol) and then
boiled for 5 min. Samples were separated on 15% polyacrylamide gels and transferred to
polyvinylidene difluoride membranes (BioRad). Membranes were incubated with mouse
anti-PLP (1:2,500), mouse anti-MBP (1:10,000), mouse anti-MOG (1:1,000), mouse anti-CNP
(1:20,000; Millipore) and mouse anti-β-actin (1;10,000; Sigma) antibodies at 4°C overnight,
and were then treated with Histofine simplestain MAX PO for 30 min. Signals were
visualized with ECL or ECL plus reagents (GE Healthcare), and quantified with a luminescent
image analyzer (LAS-3000; Fujifilm). β-Actin was used as an internal control.
Real-time reverse transcriptase polymerase chain reaction (RT-PCR)
Cervical spinal cords of wild-type and mv rats at 4 and 6 weeks of age were dissected
into the white and gray matter. Total RNA was isolated with the SV Total RNA isolation
system (Promega) according to the manufacturer’s instructions. One μg of total RNA was
transcribed with Superscript II reverse transcriptase using random primers (Invitrogen).
a Linegene system (Bioflux). Details of specific primers are listed in Table 2. β-Actin was
used as an internal control. The relative expression levels were calculated using the
comparative Ct method.
In situ hybridization for PLP mRNA
For riboprobe preparation, a 568-bp fragment of the rat PLP cDNA (GenBank
accession no. NM_0309906539) was amplified by PCR (forward primer, 5’-TTT GGA GTG
GCA CTG TTC TG-3’, nucleotide position 201-220; reverse primer, 5’-GAA AAG CAT TCC
ATG GGA GA-3’, nucleotide position 749-768). The PCR product was subcloned into pGEM
T-easy vector (Promega). DIG-labeled sense or antisense riboprobe was synthesized with SP6
or T7 RNA polymerase (Roche), respectively. For in situ hybridization, lumbar spinal cords
were fixed in 4% PFA in 0.1 M PB at 4°C overnight, treated with 30% sucrose in PBS at 4°C
for 2 or 3 days, and frozen at -80°C. Ten-μm transverse sections were cut using a cryostat. In
situ hybridization was performed as previously described (see section 1-2, Materials and
Methods). RNA hybrids were immunostained with alkaline phosphatase-conjugated anti-DIG
antibody (1:1,000; Roche) at 4°C overnight and were visualized with BCIP/NBT substrate
(Roche).
Image analysis
Tissue sections were captured with a light microscope (BX41; Olympus, Japan) and a
digital camera (DS-Fi1; Nikon). Transverse lumbar spinal cord sections immunostained for
PLP, MBP and MOG were collected from three different animals in each experimental group.
gray matter was measured using a WinRoof software (Mitani Corporation). Data are
expressed as a percentage of positive area in each white or gray matter.
Cell counts
The number of PLP-positive oligodendrocytes in the white (dorsal funiculus) or gray
matter of the lumbar spinal cord was measured using a WinRoof software. Three different
transverse sections from three different animals were evaluated in each experimental group.
The data are presented as the number of PLP-positive cells/mm2.
Statistical analysis
Data are presented as means ± standard deviation. Statistical analysis was performed
using one-way analysis of variance followed by Tukey’s test. A value of P less than 0.05 was
considered statistically significant.
Results
Expression of major CNS myelin proteins is decreased both in the white and gray matter
To clarify the distribution patterns of major CNS myelin proteins in the mv rat,
immunohistochemistry was performed using transverse spinal cord sections. In the white
matter of control rats, PLP immunoreactivity was observed from 2 weeks of age and gradually
increased with age (Fig. 11A-C). In control gray matter, PLP-positive myelin sheaths rapidly
distributed throughout the gray matter, showing a complex network of myelinated axons (Fig.
11D). Compared with control rats, PLP immunoreactivity was markedly decreased in the
white matter of mv rats from 2 weeks, and was more pronounced from 4 weeks (Fig. 11E-G).
PLP-positive myelin sheaths in the gray matter of mv rats were fewer than in control rats (Fig.
11E-G), which was most prominent at 6 weeks (Fig. 11H). PLP-positive vacuolated myelin
sheaths were diffusely observed in the gray matter of the mv rats (Fig. 11H, arrows).
Immunoreactivity for MBP in control rats gradually increased both in the white and gray
matter during postnatal myelination (Fig. 12A-D). Compared with control rats, reduced MBP
immunoreactivity was observed both in the white and gray matter of mv rats (Fig. 12E-H). As
observed in PLP immunohistochemistry, MBP-positve vacuolated myelin sheaths were
diffusely found in the gray matter of the mv rats (Fig. 12H, arrows). MOG immunoreactivity
in control white matter was weakly detected at 2 weeks and rapidly increased with age (Fig.
13A-C), while that in control gray matter was found from 4 weeks (Fig. 13A-C) and became
more intense at 6 weeks, showing a complex network of myelinated nerve fibers (Fig. 13D).
Compared with control rats, MOG immunoreactivity was markedly decreased in the white
matter of mv rats (Fig. 13E-G). MOG-positive myelin sheaths in the gray matter of mv rats
were fewer than in control rats (Fig. 13E-G); the difference was more apparent at 6 weeks
(Fig. 13H).
To evaluate the altered distribution of myelin proteins in the mv rat, the percentage of
positive area for myelin proteins was measured in the white or gray matter of control and mv
rats at 6 weeks, when the myelin lesions become most severe both in the white and gray
matter of the mv rat as previously described (see section 1-1). The results are shown in Table
3. In the white matter of the mv rats, there were significant decreases in the percentage of
positive area for PLP, MBP and MOG, of which the most severely affected was PLP
significant decreases in the percentage of positive area for PLP (16.9±0.9% in control vs.
5.1±0.5% in mv) and MOG (8.1±0.6% in control vs. 1.0±0.4% in mv), whereas only slight
decrease was observed in MBP immunostaining. These data correlated well with the
immunohistochemical findings described above.
To further investigate the changes in myelin proteins expression, Western blot analysis
was performed using samples from the white and gray matter of the cervical spinal cord at 6
weeks. Compared with wild-type rats, expression levels of PLP, MBP, CNP and MOG in mv
rats were slightly decreased in the white matter, and significantly decreased in the gray matter
(Fig. 14). These results strongly suggest gray matter hypomyelination in the spinal cord of the
mv rat.
Expression levels of PLP mRNA significantly decreases both in the white and gray matter
To analyze the expression patterns of myelin genes in the mv rat, real-time PCR for
four major CNS myelin genes was performed using samples from the white and gray matter
of the spinal cord at 4 and 6 weeks, when the gray matter lesions become more pronounced in
the mv rat as previously described (see section 1-1). Compared with wild-type rats, expression
levels of PLP mRNA significantly decreased in the white matter of mv rats at both 4 and 6
weeks (Fig. 15A). In the gray matter, expression levels of PLP mRNA were not substantially
changed between wild-type and mv rats at 4 weeks, and then significantly decreased in the mv
rats at 6 weeks. There were no significant changes in the expression of MBP, CNP and MOG
mRNA between wild-type and mv rats at either age, although overall there could be minor
The morphology and cell number of mature oligodendrocytes are not changed either in the white or gray matter
To determine whether Atrn mutation affects oligodendrogenesis in the gray matter of
the spinal cord, the morphology and cell number of oligodendrocytes in the mv rat were
examined by in situ hybridization with riboprobe for PLP mRNA, a marker for mature
oligodendrocytes. The PLP antisense probe labeled numerous oligodendrocytes in the white
and gray matter of control rats (Fig. 16A-D), whereas there was no labeling with the sense
probe (data not shown). No apparent difference was observed between control and mv rats in
the morphology of PLP-positive oligodendrocytes (Fig. 16). Furthermore, there was no
significant difference between control and mv rats in the number of PLP-positive
oligodendrocytes either in the white or gray matter (Fig. 17). These results confirm the
previous findings that Atrn mutation does not affect the cell number of CNP-positive
oligodendrocytes (see section 1-1).
Discussion
The results of this study demonstrated a marked reduction in the expression of myelin
proteins and PLP mRNA both in the white and gray matter of the mv rat during postnatal
development. Of the four major CNS myelin proteins examined, PLP was the most severely
affected in the mv rat. Since PLP is the most abundant and highly specific to the CNS
(Baumann et al., 2001; Garbay et al,. 2000), the decreased PLP expression in the mv rat can
be closely associated with the myelin alterations that are confined to the CNS (Kuwamura et
al., 2002). PLP is a highly hydrophobic tetraspan protein, localized in compact myelin.
normally and manage to assemble large amount of myelin (Hudson, 2004 ; Klugmann et al.,
1997). However, there are subtle but widespread ultrastructural abnormalities in the CNS
myelin; an increased numer of unmyelinated and thinly myelinated axons, disturbance of the
radial component of myelin ("diagonal" component), and most strikingly, irregular
interlamellar spaces of variable width (Rosenbluth et al., 2006; Yool et al., 2002). These
findings of "looser" myelin sheaths in the PLP-null mouse indicate a structural role of PLP in
maintaining myelin compaction along the external surface of the myelin lamella. In the mv
rat, myelin vacuolation is due to splitting of the myelin sheath and its frequency increases
with age, suggesting impaired maintenance of CNS myelin (Kuwamura et al., 2002 and this
study). It is possible that the severe reduction of PLP impairs the maintenance of myelin
compaction, resulting in myelin vacuolation in the mv rat.
MOG is regarded as an marker of developmental maturation of oligodendrocytes,
since it is not expressed by immature oligodendrocytes but by mature myelinating
oligodendrocytes (Trapp et al., 2004). Despite its importance as a target for autoimmune
attack in demyelinating diseases, the physiological function of MOG is still obscure.
Interestingly, MOG-deficient mice exhibit no morphological abnormalities in the CNS myelin
and axons, suggesting that MOG itself is not essential for normal CNS myelination (Delarasse
et al., 2003). Therefore the decrease of MOG in the mv rat seems to reflect an impairment of
oligodendrocyte maturation or of the production of MOG, rather than to cause structural
alterations in myelin sheaths. A recent histological study in the brain of the zitter rat revealed
severe morphological abnormalities of oligodendrocytes during postnatal myelination,
indicating an impairment of oligodendrocyte differentiation (Sakakibara et al., 2008). The
results of this study show that the composition and distribution of major myelin proteins are
oligodendrocytes are not affected. These data suggest an impairment of oligodendrocyte
differentiation and/or a failure of oligodendrocytes to produce myelin proteins in the mv rat.
The results of this study revealed severe hypomyelination in the gray matter of the mv
rat as well as the white matter. Recent studies in human MS have focused on the pathogenesis
of gray matter demyelination since it is thought to be more related with clinical disability than
that in the white matter (Pirko et al., 2007; Stadelmann et al., 2008). Gray matter lesions in
MS are characterized by a relative lack of lymphocyte infiltration, astrogliosis and microglial
activation, and more extensive remyelination compared with the white matter lesions (Albert
et al., 2007; Bo et al., 2003; Peterson et al., 2001). In murine cuprizone model of
demyelination, the time corse and dynamics of demyelination, and cellular responses differ
between the corpus callosum and cerebral cortex, of which the most striking is a considerable
reduction in the number of activated microglia in the cortex (Gudi et al., 2009).
Hypomyelination of the cerebral cortex is noted in the jimpy mutant mouse, a mutant with
myelin deficiency and oligodendrocyte cell death caused by a PLP mutation (Baracskay et al.,
2002). Oligodendrocyte number in the jimpy mouse is reduced in the cerebral cortex, but
increased in the white matter, supporting the hypothesis that oligodendrogenesis is
differentially regulated in the white and gray matter. Together, the results of this study suggest
that the microglial activation in the gray matter of the mv rat is closely associated with the
severe gray matter lesions, and that glial responses to myelin alterations could differ between
the white and gray matter. It remains to be elucidated whether this impairment is caused by an
intrinsic defect of Atrn-deficient oligodendrocytes or by a loss of interaction between
Summary
The expression patterns of major CNS myelin proteins in the spinal cord of the mv rat
were examined to explore the role of Atrn in myelin production and to examine the
relationship between the gray matter lesions and microglial activation in the mv rat.
Immunohistochemical and Western blot analyses demonstrated a considerable reduction in the
expression of major CNS myelin proteins both in the white and gray matter of the mv rat.
Real-time PCR analysis revealed a significant decrease in the expression of PLP mRNA both
in the white and gray matter of the mv rat. However, there was no significant difference in the
cell number of PLP mRNA-positive oligodendrocytes between control and mv rats either in
the white or gray matter, suggesting an impairment of oligodendrocytes in the myelin protein
production. The results of this study indicate that myelinogenesis but not oligodendrogenesis
is severely altered both in the white and gray matter of the mv rat, and that microglial
activation is closely associated with the severe hypomyelination in the gray matter of the mv
Table 2. Primers for real-time RT-PCR
Gene Primer sequence Primer location Accession number Note
PLP forward: 5’- TTGTTTGGGAAAATGGCTAGGA -3’ at the end of exon3 NM_0309906539 specific to PLP,
reverse: 5’-GCAGATGGACAGAAGGTTGGA-3’ exon5 excluding its splice variant DM20
MBP forward: 5’- TCACAGAAGAGACCCTCACAGC -3’ exon1 NM_001025291 common for all 6 splice variants
reverse: 5’-GAGTCAAGGATGCCCGTGTC-3’ exon1 of MBP
CNP forward: 5’- TCCGAGGAGTACAAGCGTCT -3’ exon2 NM_012809
reverse: 5’-ACAGCTGCCATTGGTTCTTC-3’ exon2
MOG forward: 5’- TCCGTGCAGAAGTCGAGAA -3’ between exon 4 and 5 NM_022668
reverse: 5’-CCATCACTCAAAAGGGGTTTC-3’ exon8
β-Actin forward: 5’-TAAAGACCTCTATGCCAACAC-3’ exon5 NM_031144
reverse: 5’-CTCCTGCTTGCTGATCCACAT-3’ exon6
PLP; proteolipid protein, MBP; myelin basic protein, CNP; 2',3'-cyclic nucleotide 3'-phosphodiesterase, MOG; myelin oligodendrocyte glycoprotein
Table 3. Percentage of positive area for myelin proteins
white matter gray matter
control mv control mv
PLP 55.5±4 20.6±3 * 16.9±0.9 5.1±0.5 *
MBP 45.8±4 29.8±2 * 6.4±0.6 3.7±1
MOG 36.2±2 24.3±1 * 8.1±0.6 1.0±0.4 *
PLP; proteolipid protein, MBP; myelin basic protein, MOG; myelin oligodendrocyte glycoprotein