学位論文
パーキンソン病における痛覚過敏と
その薬物療法に関する基礎研究
大阪大学大学院歯学研究科
研究逐行 : 口腔科学専攻 顎口腔病因病態制御学講座
( 所属 : 口腔科学専攻 高次脳口腔機能学講座 )
成清 綾
目次
緒言 ……… 1 実験方法 ……… 2 1. 実験動物 ……… 3 2. 薬物 ……… 3 3. 6-OHDA 誘発片側線条体破壊マウスの作製 ……… 4 4. 自発運動量測定試験 ……… 4 5. ロータロッド試験 ……… 5 6. ホットプレート試験 ……… 5 7. プランター試験 ……… 5 8. カプサイシン誘発疼痛試験 ……… 5 9. von Frey 試験 ……… 6 10 免疫組織化学的解析 ……… 6 11 統計解析 ……… 9 結果 ……… 10 1. 6-OHDA 誘発片側線条体破壊マウスの運動機能の評価 ……… 10 2. 6-OHDA 誘発片側線条体破壊マウスの熱刺激,化学刺激および機械刺激 に対する痛み感受性の変化 ……… 10 3. 6-OHDA 誘発片側線条体破壊マウスの痛覚関連部位におけるカプサイシン 誘発 c-Fos 発現の変化 ……… 11 4. 6-OHDA 誘発片側線条体破壊マウスの痛覚過敏に対する PXT 急性投与 の効果 ……… 11 5. 6-OHDA 誘発片側線条体破壊マウスの痛覚過敏に対する PXT 慢性投与の効果 ……… 12
考察 ……… 19
謝辞 ……… 25
緒言
Parkinson 病 (PD) は,中脳黒質から線条体に投射するドパミン (DA) 作動性神経細 胞の選択的な変性・脱落により,振戦,筋固縮,無動,姿勢反射障害の運動機能障害を 発現する神経変性疾患である.PD の罹患率は 0.18%とされ,発症年齢は 50~65 歳に 多いが, 高齢になるほど発症率が増加し,70 歳以上では約 100 人に 1 人の頻度で発 症していると言われている (Yamawaki et al., 2009).一方,PD 患者では,運動機能障 害に加えて,痛覚異常,認知機能障害,気分障害などの非運動機能障害がしばしば認め られる (Rana et al., 2015).なかでも,40%以上の PD 患者において診られる痛覚過敏 (Broen et al., 2012) は,QOL を低下させる大きな一要因と考えられている.PD 患者の薬物治療では,運動機能障害の改善を目的として DA 前駆薬であるL-DOPA
や DA 受容体作用薬が適用されている (LeWitt, 2015; Alonso Cánovas et al., 2014) が, 非運動機能障害に対する薬物治療は施されておらず,なかでも発現率の高い痛覚過敏に 対する薬物療法の確立が待ち望まれている.このような背景のもと,最近,PD モデル 動物において,L-DOPA が痛覚過敏を部分的に改善すること (Park et al., 2015),セロ トニン (5-HT) ・ノルアドレナリン (NA) 再取り込み阻害薬であるデュロキセチンと深 部脳刺激の併用が痛覚過敏を抑制すること (Kaszuba et al., 2017) が報告されたが,PD における痛覚過敏に関わる神経化学基盤は未だ明らかとされていない. そこで本研究は,PD における痛覚過敏に関わる神経化学基盤を明らかとし,また, その神経化学基盤を標的とする新たな薬物療法を確立することを目的とした.まず, DA 神経毒である 6-ヒドロキシドーパミン臭化水素酸塩 (6-OHDA) の脳局所注入によ り片側線条体を破壊したマウス (6-OHDA 誘発片側線条体破壊マウス) を用いて,痛覚 過敏を行動学的に確認し,次に,免疫組織学的手法により痛み刺激の受容と制御に関わ る上行性痛覚伝達経路ならびに下行性痛覚抑制経路の応答変化を検討した.さらに,変
化を見いだした神経系を標的とする薬物を用いて,PD の痛覚過敏に対する新規薬物療 法の可能性を追究した.
実験方法
1. 実験動物 実験には,8 週齢の C57BL/6J 系雄性マウス (日本エスエルシー株式会社,浜松) を 購入した.実験動物は,室温 22±1℃,明暗サイクル 8:00~20:00,プラスチック ケージ (28 cm×17 cm×12 cm) に 1 ケージ 4‒5 匹にて飼育し,水と固形飼料は自由 に摂取させた.1 週間環境に馴化させた後,実験に用いた.なお,動物実験は,「大阪 大学大学院歯学研究科動物実験規定」ならびに「大阪大学大学院薬学研究科動物実験規 定」に従って倫理的に行った. 2. 薬物 L-アスコルビン酸ナトリウムはナカライテスク株式会社 (京都) より購入した.デシ プラミン塩酸塩,6-ヒドロキシドーパミン臭化水素酸塩 (6-OHDA),パロキセチンマレ イン酸塩は Sigma-Aldrich Co. (St. Louis, MO, USA) より購入した.(R)-(-)-アポモルフ ィン塩酸塩 0.5 水和物 (アポモルヒネ),カプサイシンは和光純薬工業株式会社 (大阪) より購入した. デシプラミン塩酸塩は生理食塩水 (0.9% w/v,大塚製薬株式会社,東京) に溶解し た.6-OHDA は 0.1%アスコルビン酸ナトリウム含有溶液に溶解し,2.0 mg/mL の濃度 で溶解した.アポモルヒネは生理食塩水に溶解した.カプサイシンは 1%エタノール含 有生理食塩水に 80 µg/mL の濃度で溶解した. パロキセチンマレイン酸塩 (PXT) は生理食塩水に溶解し 3 または 10 mg/kg の用量 で腹腔内投与した (Gai et al., 2014).単回投与実験では,PXT 投与 30 分後に行動解析 を行った.慢性投与実験では,PXT を 2 週間 (1 回/1 日) 投与し,PXT 最終投与 24 時 間後に行動解析を行った.3. 6-OHDA 誘発片側線条体破壊マウスの作製
6-OHDA 誘発片側線条体破壊マウスは Coulomb らの方法 (2016) に従って作製した. 6-OHDA 注入 30 分前に NA 神経保護のためにデシプラミン塩酸塩 (25 mg/kg) を腹腔 内投与した (Tanaka et al., 2017 ; Parikh et al., 2016).ペントバルビタールナトリウム (50 mg/kg, i.p.) 麻酔下,左側線条体 (ブレグマを基準として AP: +1.0 mm, ML: +1.7 mm, DV: ‒3.0 mm) (Paxinos and Franklin, 2004) に 4 分間かけて 6-OHDA 溶液を (2.0 mg/mL) を 2.0 µL 注入した (流速:0.5 µL/min).注入後 5 分間針を静置した. 6-OHDA 注入 1 週間後にマウスをプラスチックケージ (24 cm×17 cm×12 cm) に 入れて,10 分間馴化させた後,アポモルヒネ (0.5 mg/kg) を皮下投与して,線条体非 破壊側への回転運動を 60 分間計測した.Iancu らの基準 (2005) に従い,1 分間あた り 4 回以上回転運動した (12.0 ± 1.10 回/分) マウスを 6-OHDA 誘発片側線条体破壊 マウスとして使用した.一部の 6-OHDA 誘発片側線条体破壊マウスでは,免疫組織化 学染色により,ドパミン神経の破壊を確認した. 4. 自発運動量測定試験 マウスの自発運動量はHasebeらの方法 (2016) に従って,自発運動量測定システム (Supermex®; 室町機械株式会社,東京) を用いて測定した.マウスをプラスチックケー ジ (28 cm×17 cm×12 cm) に入れ,サウンドアテニュエイティング・チャンバ (PYB-003; 室町機械株式会社) に静置し,90分馴化させた.その後,自動的に5分間隔 で赤外線センサー (Supermex® センサー PYS-001; 室町機械株式会社) の下を横切っ た回数を測定して,開始から120分後に終了とした.スーパーメックスセンサーから得 られたデータはCompACT AMS Ver.3 (室町機械株式会社) を用いて解析した.
5. ロータロッド試験
ロータロッド試験は Ago らの方法 (2011) に従って,マウス用ロータロッド (Acceler Rota-Rod, Model 7650; Ugo Basile, Varese, Italy) を用いて測定した.マウスを 一定速度で回転するロッドの上に置き,強制的に歩行させ,ロッドから落下するまでの 時間を測定した.回転速度は 5 rpm および 16 rpm とし,カットオフ値を 5 分とした.
6. ホットプレート試験
ホットプレート試験は Horiguchi らの方法 (2013b) に準じて行った.マウスを 49℃ (±0.5℃) もしくは 55℃ (±0.5℃) に設定したプレート (Model 35100; Ugo Basile) の上に置き,licking するまでもしくは熱刺激から逃避するためにジャンプするまでの 時間を測定した.全てのマウスは痛み刺激に対するストレスを取り除くために,ハンド リングを行った.肢の組織の損傷を避ける為に,カットオフ値は低温の場合 30 秒,高 温の場合 15 秒と設定し,全てのマウスの使用は 1 回とした. 7. プランター試験 プランター試験は Horiguchi らの方法 (2013a) に準じて行った.マウスを透明なプ ラスチックケージ (10 cm×18 cm×10 cm) に入れ,90 分馴化させた.プランター式 鎮痛効果測定装置 (Ugo Basile) を用いて熱光源を後肢にあて,逃避反応を起こすまで の時間を測定した.対照群のマウスが逃避反応を起こすまでの時間が 8‒11 秒となるよ うに刺激値を設定し,15 秒をカットオフ値とした. 8. カプサイシン誘発疼痛試験 カプサイシン誘発疼痛試験は Horiguchi らの方法 (2013a) に従って行った.マウス をプラスチックケージ (10 cm×18 cm×10 cm) に入れ,90 分テスト環境に馴化させ
た.30 ゲージの BD ロードーズ (日本ベクトン・ディッキンソン株式会社,福島) を用 いてカプサイシン溶液 (1.6 µg/20 µl) を後肢足底に投与した.対照群には溶剤である 1%エタノール含有生理食塩水 (vehicle) を 20 µL 注射した.注射後の 5 分間の licking 及び lifting 時間を測定した. 9. von Frey 試験
von Frey 試験は Horiguchi らの方法 (2013b) に準じて,Touch Test™ Sensory Evaluators (Stoelting Co., Wood Dale, IL, USA) を用いて測定した.マウスを網の上に置 き,その上から透明なプラスチックゲージ (10 cm×18 cm×10 cm) をかぶせ,60 分 馴化させた.機械刺激値は加圧力が 0.008,0.02,0.04,0.07,0.16,0.4,0.6,1.0, 1.4,2.0 g のフィラメントを用いて up-down 法 (Bonin et al., 2014) で測定した.フィ ラメントが曲がるまで後肢に負荷をかけ,最大 3 秒間逃避反応を観察した.刺激は加圧 力が 0.16 g のフィラメントから開始し,逃避反応を認めると 1 段階加圧力が小さいフ ィラメントに変更し,逃避反応が認められなくなるまで繰り返した.逃避反応の消失が 認められた後,1段階加圧力が大きいフィラメントでの刺激に対する逃避反応と再度 1 段階加圧力が小さいフィラメントでの逃避反応の消失を確認し,逃避反応を引き起こし たフィラメントの最少機械刺激値を逃避反応閾値とした. 10. 免疫組織化学的解析
チロシンヒドロキシラーゼ (TH) 免疫染色は既報の方法 (Ago et al., 2011; Maegawa et al., 2015) に準じて,一部のマウスで行った.ペントバルビタールナトリウム (50 mg/kg, i.p.) で麻酔し,左心室より,生理食塩水を全身に灌流させ脱血後,4%パラホル ムアルデヒド (PFA) / 0.1M リン酸緩衝液 (PB) を灌流させ組織を固定した. 全脳を摘 出し,4% PFA/0.1 M PB 中で 4 ℃,24 時間浸潤固定を行った後,20%スクロース溶
液に置換し,2 日間脱水処置を行った.組織をティシュ・テック OCT コンパウンド (サ クラファインテックジャパン株式会社,東京) に包埋し,凍結させた後,クライオスタ ット (CM1520; Leica Microsystems GmbH, Wetzlar, Germany) を用いて,線条体を含 む厚さ 20 µm の冠状切片を作製した.得られた切片を MAS コートスライドガラス (松 浪硝子工業株式会社,大阪) に貼付し,風乾後,クエン酸バッファー (pH 6.0) 中で 10 分間マイクロウェーブを照射した.室温中 5%ウマ血清 (Vectastain ABC kit ; Vector Laboratories, Burlingame, CA, USA) で 1 時間ブロッキングした。ブロッキング後,マ ウス抗 TH 抗体 (1,000 倍希釈;Sigma-Aldrich Co.) で 12 時間,4℃でインキュベート した.つづいて,抗マウス IgG 抗体 (50 倍希釈;Vectastain ABC kit) で 1 時間,室温 でインキュベートした.切片を 0.03% PBST で洗浄し,アビジン・ビオチン標識酵素 複合体液 (25 倍希釈;Vectastain ABC kit) を 1 時間室温で反応させた。さらに,0.05% ジアミノベンチジンテトラハイドロクロライド (DAB) / 1 M Tris-HCl (pH 7.4)を反応さ せた.その後,100%エタノール,キシレンで脱水処理して,Mount Quick (コスモ・バ イオ株式会社,東京) を用いて封入し,顕微鏡で観察確認した (Fig. 1).
Fig. 1. 6-OHDA 誘発片側線条体破壊マウスの TH 免疫組織化学染色の典型的な顕微鏡像. Bar は 1 mm.(A) Sham 群. (B) 6-OHDA 投与群.
c-Fos 免疫染色は,既報の方法 (Horiguchi et al., 2013a; Hara et al., 2015, 2016) に 準じて行った.マウスの右後肢足底部表面にカプサイシン溶液 (1.6 µg/20 µL) または
vehicle を注射した 2 時間後に,ペントバルビタールナトリウム (50 mg/kg, i.p.) で麻 酔し,左心室より,生理食塩水を全身に灌流させ脱血後,4% PFA/0.1 M PB を灌流さ せ組織を固定した. 全脳を摘出し,4% PFA/0.1 M PB 中で 4 ℃,24 時間浸潤固定を 行った後,20%スクロース溶液に置換し,2 日間脱水処置を行った. 体性感覚野 (SSC; ブレグマを基準に‒0.7 mm~‒1.0 mm),扁桃体基底外側核 (BLA ; ブレグマを基準に‒1.3 mm~‒1.5 mm),前部帯状回皮質 (ACC; ブレグマを基準に+0.1 mm~+0.5 mm),中脳水道灰白質 (PAG; ブレグマを基準に‒3.5 mm~‒4.2 mm),青斑 核 (LC ; ブレグマを基準に‒ 5.3mm~‒5.5 mm) および延髄吻側腹内側部 (RVM; ブレ グマを基準に‒5.8 mm~‒6.1 mm) の脳各領域を含む厚さ 20 µm の冠状切片を作製し た.得られた切片を MAS コートスライドガラス (松浪硝子工業株式会社) に貼付し, 風乾後,クエン酸バッファー (pH 7.0) 中で 10 分間マイクロウェーブを照射した.0.3% H2O2に 5 分間浸し,内因性ペルオキシダーゼ不活性化させた後,室温中 5%ヤギ血清
(Vectastain ABC kit) で 1 時間ブロッキングした。ブロッキング後,ウサギ抗 c-Fos 抗 体(1,000 倍希釈;sc-253, Santa Cruz Biotechnology, Dallas, TX, USA)で 12 時間,4℃ でインキュベートした.つづいて,ビオチン化ヤギ抗ウサギ抗体 (100 倍希釈; Vectastain ABC kit) で 1 時間,室温でインキュベートし,アビジン・ビオチン標識酵 素複合体液 (50 倍希釈;Vectastain ABC kit) を 1 時間室温で反応させた.さらに,0.05% DAB / 1 M Tris-HCl (pH 7.4)を反応させた.その後,100%エタノール,キシレンで脱 水処理して,Mount Quick (コスモ・バイオ株式会社) を用いて封入した.
c-Fos 陽性細胞数の計数は,冷却 CCD デジタルカメラシステムを装備した正立顕微鏡 (Axio Imager.M2/AxioCam MRc5; Carl Zeiss, Jena, Germany) を用いて各脳領域を含む 5 枚の独立した切片から画像を取り込んだ.SCC,BLA と ACC では左脳で計数し,1×
1 mm2あたりの c-Fos 陽性細胞数の平均値を算出した.LC では左脳の領域全体で c-Fos
の c-Fos 陽性細胞数の平均値を算出した.PAG は左右脳を合わせた領域全体で c-Fos 陽 性細胞数の平均値を算出した.
11. 統計解析
全てのデータは平均値±標準誤差で示した.統計解析は解析ソフト Prism 6 for (GraphPad Software, San Diego, CA, USA) を用いて行い,有意水準はP < 0.05 とした. 自発運動量の経時変化は two-way repeated measures ANOVA にて,ロータロッド試験 での落下時間,痛覚感受性変化 (2 群比較) および c-Fos 発現変化は unpaired t-test に て,ホットプレート試験での逃避潜時および痛覚感受性変化に対するパロキセチンの効 果は two-way ANOVA ならびにpost hoc Sidak's multiple comparisons test にて解析を 行った.
結果
1. 6-OHDA 誘発片側線条体破壊マウスの運動機能の評価
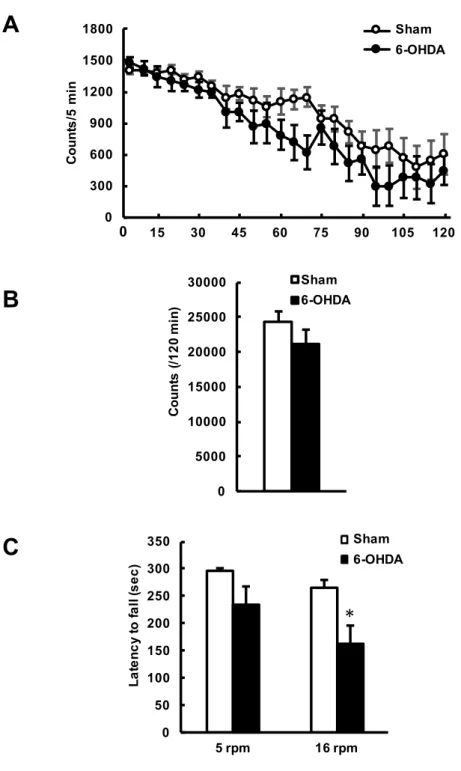
6-OHDA 誘発片側線条体破壊マウスの運動機能変化を自発運動量測定試験およびロ ータロッド試験で評価した (Fig. 2). Sham 群と 6-OHDA 群の自発運動量の経時変化 (operation: F1,7 = 1.605, P = 0.2458; time: F23,161 = 25.50, P < 0.0001; interaction: F23,161
= 0.7097, P = 0.8312 by two-way repeated measures ANOVA) (Fig. 2A) および 120 分 間の総運動量 (P = 0.1319 by unpaired t-test) (Fig. 2B) の間に有意な差違は認められ なかった.また,ロータロッド試験の 5 rpm での試行においても,Sham 群と 6-OHDA 群の間に有意な差違は認められなかった (P = 0.0940 by unpaired t-test) (Fig. 2C, left). 一方,回転数を 16 rpm に上げた試行では,6-OHDA 群は Sham 群と比べてロッドから の落下時間が有意に短縮していた (P < 0.05 by unpaired t-test) (Fig. 2C, right).
2. 6-OHDA 誘発片側線条体破壊マウスの熱刺激,化学刺激および機械刺激に対する痛み 感受性の変化
片側線条体破壊マウスにおける痛み感受性変化を 6-OHDA 注入 2 週間後に評価した (Fig. 3).ホットプレート試験の 49℃および 55℃の両試行において,Sham 群と 6-OHDA 群の逃避潜時の間に有意な差違は認められなかった (operation: F1,36 = 2.205, P =
0.1463; temperature: F1,36 = 279.1, P < 0.0001; interaction: F1,36 = 1.531, P = 0.2239 by
two-way ANOVA) (Fig. 3A).つぎに,破壊線条体と同側後肢 (左後肢) に痛み刺激を負 荷する群を ipsilateral 群,反対側後肢 (右後肢) に痛み刺激を負荷する群を contralateral 群とし,片側線条体破壊マウスにおける熱刺激 (Fig. 3B),化学刺激 (Fig. 3C) および機 械刺激 (Fig. 3D) に対する痛み感受性変化を検討した.Ipsilateral 群での検討では,プ ランター試験での逃避潜時 (P = 0.0791 by unpaired t-test) (Fig. 3B, left),カプサイシ
ン誘発疼痛試験での licking および lifting 時間 (P = 0.4496 by unpaired t-test) (Fig. 3C, left) ならびに von Frey 試験での逃避閾値 (P = 0.8838 by unpaired t-test) (Fig. 3D, left) のいずれにおいても,Sham 群と 6-OHDA 群の間に有意な差は認められなかった. 一方,contralateral 群での検討では,6-OHDA 群は Sham 群と比べて,プランター試験 での逃避潜時の短縮 (P < 0.01 by unpaired t-test) (Fig. 3B, right),カプサイシン誘発疼 痛試験での licking および lifting 時間の増大 (P < 0.001 by unpaired t-test) (Fig. 3C, right) および von Frey 試験での逃避閾値の低下 (P < 0.001 by unpaired t-test) (Fig. 3D, right)が認められた.
3. 6-OHDA 誘発片側線条体破壊マウスの痛覚関連部位におけるカプサイシン誘発 c-Fos 発現の変化
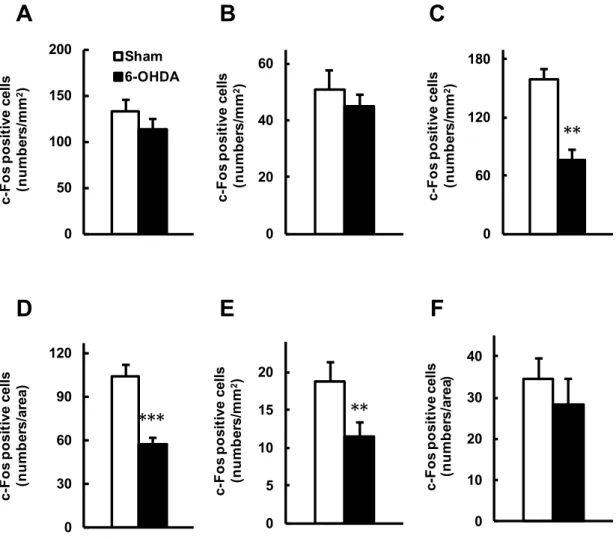
破壊線条体と反対側後肢足底 (右後肢足底) へのカプサイシン (1.6 µg/ 20 µl) 刺激に よる脳内での c-Fos 発現を指標として,片側線条体破壊マウスの痛覚関連部位における 神経応答変化を解析した (Fig. 4, 5).SSC (P = 0.2565 by unpaired t-test) (Fig. 5A), BLA (P = 0.4929 by unpaired t-test) (Fig. 5B) および LC (P = 0.1592 by unpaired t-test) (Fig. 5F) における c-Fos 陽性細胞数において,Sham 群と 6-OHDA 群の間に有意な差違は認 められなかった.一方,ACC (P < 0.01 by unpaired t-test) (Fig. 5C), PAG (P < 0.001 by unpaired t-test) (Fig. 5D) および RVM (P < 0.01 by unpaired t-test) (Fig. 5E) において は,6-OHDA 群は Sham 群と比べて c-Fos 陽性細胞数が有意に減少していた.
4. 6-OHDA 誘発片側線条体破壊マウスの痛覚過敏に対する PXT 急性投与の効果
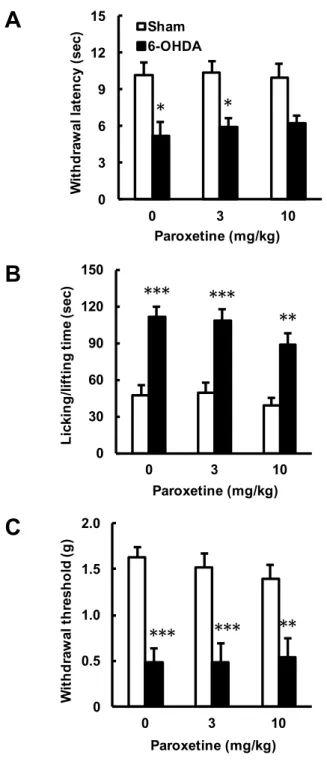
6-OHDA 注入 2 週間後に認められた片側線条体破壊マウス右後肢 (contralateral後肢) での痛み感受性増大に対して,PXT 急性投与の効果を検討した (Fig. 6).PXT の単回投 与は,3 および 10 mg/kg のいずれの用量においても,6-OHDA 群が示すプランター試
験での逃避潜時の短縮 (drug: F2,42 = 0.1216, P = 0.8858; operation: F1,42 = 29.94, P <
0.0001; interaction: F2,42 = 0.1857, P = 0.8312 by two-way ANOVA) (Fig. 6A),カプサ
イシン誘発疼痛試験での licking および lifting 時間の増大 (drug: F2,42 = 2.043, P =
0.1424; operation: F1,42 = 66.01, P < 0.0001; interaction: F2,42 = 0.3451, P = 0.7101 by
two-way ANOVA) (Fig. 6B) ならびに von Frey 試験での逃避閾値の低下 (drug: F2,42 =
0.1585, P = 0.8539; operation: F1,42 = 66.88, P < 0.0001; interaction: F2,42 = 0.4371, P =
0.6488 by two-way ANOVA) (Fig. 6C) のいずれに対しても効果を示さなかった.
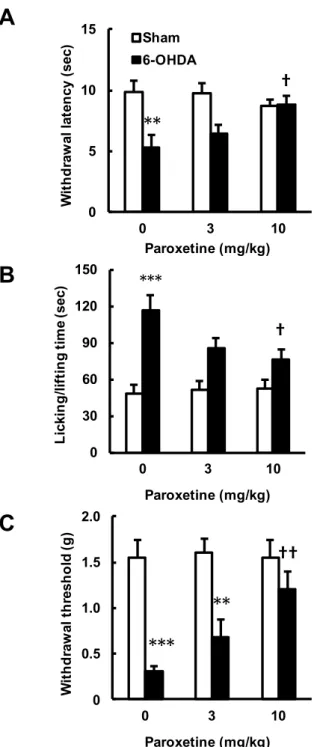
5. 6-OHDA 誘発片側線条体破壊マウスの痛覚過敏に対する PXT 慢性投与の効果
さらに,6-OHDA 注入 2 週間後に認められた片側線条体破壊マウスの右後肢 (contralateral 後肢) での痛み感受性増大に対して,PXT 慢性投与の効果を検討した (Fig. 7).6-OHDA 注入 2 週間後より 2 週間の PXT 反復投与 (10 mg/kg, 1 回/1 日) は, 最終投与の 24 時間において,6-OHDA 群が示すプランター試験での逃避潜時の短縮 (drug: F2,42 = 1.068, P = 0.3527; operation: F1,42 = 15.62, P = 0.0003; interaction: F2,42 =
4.626, P = 0.0153) (Fig. 7A),カプサイシン誘発疼痛試験での licking および lifting 時 間の増大 (drug: F2,42 = 2.563, P = 0.0891; operation: F1,42 = 39.07, P < 0.0001;
interaction: F2,42 = 3.954, P = 0.0267) (Fig. 7B),ならびに von Frey 試験での逃避閾値の
低下 (drug: F2,42 = 3.831, P = 0.0296; operation: F1,42 = 40.44, P < 0.0001; interaction:
F2,42 = 3.930, P = 0.0273) (Fig. 7C) の全てに対して改善効果を示した (for all
comparisons, saline/sham vs. vehicle/6-OHDA: P < 0.01; saline/6-OHDA vs. 10 mg/kg PXT/6-OHDA: P < 0.05 by post hoc Sidak's multiple comparisons test).なお,3 mg/kg の用量では,PXT 慢性投与は片側線条体破壊マウス右後肢での痛み感受性増大に対して 影響を示さなかった (saline/6-OHDA vs. 3 mg/kg PXT/6-OHDA: P > 0.1144 by post hoc Sidak's multiple comparisons test) (Fig. 7A‒C).
Fig. 2. 6-OHDA誘発片側線条体破壊マウスの運動機能の評価.マウスの左側線条体に 生理食塩水もしくは6-OHDAを局所注入し,その14日後に行動解析を行った.A. 自発 運動量測定試験.マウスが赤外線センサーの下を通過する回数を測定した.B. ロータ ロッド試験.マウスが回転するロッドから落下するまでの時間を測定した.データは平 均値±標準誤差で示す [n = 8 (A), n = 10 (B)].*P < 0.05, vs. Sham群 (unpaired t-test).
A
0 50 100 150 200 250 300 350 5 rpm 16 rpm Lat en cy to fal l ( sec)*
0 5000 10000 15000 20000 25000 30000 Co un ts (/120 m in ) Sham 6-OHDAB
C
0 300 600 900 1200 1500 1800 15 30 45 60 75 90 105 120 Co un ts /5 m in Control 6-OHDA 0 Sham 6-OHDA Sham 6-OHDAFig. 3. 6-OHDA 誘発片側線条体破壊マウスの痛覚感受性の変化.マウスの左側線条体 に生理食塩水もしくは 6-OHDA を局所注入し,その 14 日後に行動解析を行った.なお, B‒D のデータにおいては,痛み刺激を左後肢に与えたマウスを ipsilateral 群, 右後肢に 与えたマウスを contralateral 群とした. A. ホットプレート試験.マウスを 49℃あるい は 55℃としたプレート上に置き,逃避反応を起こすまでの時間を測定した.B. プラン ター試験.マウスの後肢に熱光源を当て,逃避反応を起こすまでの時間を測定した.C. カプサイシン誘発疼痛試験.マウスの後肢足底にカプサイシン (1.6 µg/20 µl) を投与 し,その直後 5 分間の licking 及び lifting 時間を測定した.D. von Frey 試験.マウスの 後肢に von Frey フィラメントを用いて強度の異なる刺激を与え,逃避反応を起こす反 応閾値を測定した.データは平均値±標準誤差で示す (n = 10).*P < 0.05, ***P < 0.001, vs. Sham 群 (unpaired t-test).
A
B
C
D
0.0 0.5 1.0 1.5 2.0 Ipsilateral cotralateral W ithdr aw al thr es hol d (g) 0 30 60 90 120 150 Ipsilateral cotralateral Li cki ng /li ft in g tim e (sec) 0 3 6 9 12 Ipsilateral cotralateral W ith dr aw al lat en cy (sec)*
***
*
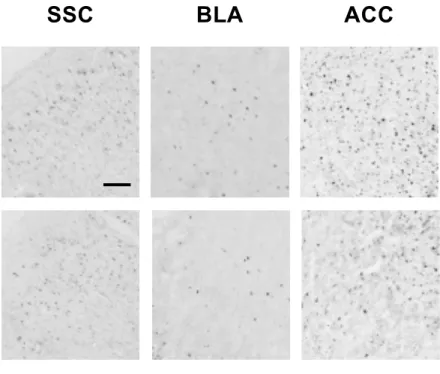
0 5 10 15 20 25 30 49°C 55°C W ith dr aw al lat en cy (sec) Sham 6-OHDA Contralateral Contralateral Contralateral 0Fig. 4. マウスの後肢足底へのカプサイシン投与による脳内 c-Fos 発現を示す典型的な 顕微鏡像.マウスの左側線条体に生理食塩水もしくは 6-OHDA を局所注入し,その 14 日後に右後肢足底へカプサイシン (1.6 µg) を投与した.投与 2 時間後に組織を固定し て脳薄切片を作製し,免疫染色法により体性感覚野 (SSC),扁桃体基底外側核 (BLA), 前部帯状回皮質 (ACC),中脳水道灰白質 (PAG),延髄吻側腹内側部 (RVM) および青斑 核 (LC)を含む切片の c-Fos 発現細胞を可視化した.Bar は 100 µm.
SSC
BLA
ACC
Sham
6-OHDA
PAG
RVM
LC
Sham
6-OHDA
Fig. 5. 6-OHDA 誘発片側線条体破壊マウスの後肢足底へのカプサイシン投与による脳 内 c-Fos 発現変化.マウスの左側線条体に生理食塩水もしくは 6-OHDA を局所注入し, その 14 日後に右後肢足底へカプサイシン (1.6 µg) を投与した.投与 2 時間後に組織 をホルマリン灌流により固定し,脳薄切片を作製した.痛覚関連部位を含む切片におけ る c-Fos 発現細胞を免疫染色法により可視化して計数した.A. 体性感覚野 (SSC),B. 扁 桃体基底外側核 (BLA),C. 前部帯状回皮質 (ACC),D. 中脳水道灰白質 (PAG),E. 延髄 吻側腹内側部 (RVM),F. 青斑核 (LC).データは平均値±標準誤差で示す (n = 8).*P < 0.05, ***P < 0.001, vs. Sham 群 (unpaired t-test).
0 50 100 150 200 c-Fo s po si tive cel ls (n um be rs /m m 2) Sham 6-OHDA 0 20 40 60 c-Fo s po si tive cel ls (n um be rs /m m 2) 0 60 120 180 c-Fo s po si tive cel ls (n um be rs /m m 2)
**
0 30 60 90 120 c-Fo s po si tive cel ls (n um ber s/ ar ea )***
0 5 10 15 20 c-Fo s po si tive cel ls (n um ber s/ m m 2) 0 10 20 30 40 c-Fo s po si tive cel ls (n um be rs /a re a)**
A
B
D
E
C
F
Fig. 6. 6-OHDA 誘発片側線条体破壊マウスの痛覚過敏に対するパロキセチン (PXT) 急性投与の効果.マウスの左側線条体に生理食塩水もしくは 6-OHDA を局所注入し, その 14 日後に PXT (3, 10 mg/kg, i.p.) を投与した.PXT 投与 30 分後に行動解析を行 った.A. プランター試験.B. カプサイシン誘発疼痛試験.C. von Frey 試験.データは 平均値±標準誤差で示す (n = 8).*P < 0.05, **P < 0.01, ***P < 0.001, vs. Sham 群 (Sidak's multiple comparisons test).
0.0 0.5 1.0 1.5 2.0 0 3 10 W ithdr aw al thr es hol d (g) 0 30 60 90 120 150 0 3 10 Li ck in g/ lif tin g tim e (s ec ) 0 3 6 9 12 15 0 3 10 W ith dr aw al lat en cy (sec) Sham 6-OHDA
*
*
***
***
***
**
***
**
Paroxetine (mg/kg) Paroxetine (mg/kg) Paroxetine (mg/kg)A
B
C
0Fig. 7. 6-OHDA 誘発片側線条体破壊マウスの痛覚過敏に対するパロキセチン (PXT) 慢性投与の効果.マウスの左側線条体に生理食塩水もしくは 6-OHDA を局所注入し, その 14 日後から 2 週間 PXT (3, 10 mg/kg, i.p., 1 日 1 回) を投与した.PXT 投与 24 時間後に行動解析を行った.A. プランター試験.B. カプサイシン誘発疼痛試験.C. von Frey 試験.データは平均値±標準誤差で示す (n = 8).**P < 0.01, ***P < 0.001, vs. Sham 群; †P < 0.05, ††P <0.01, vs. vehicle 群 (Sidak's multiple comparisons test).
0 5 10 15 0 3 10 W ith dr aw al lat en cy (sec) Sham 6-OHDA 0.0 0.5 1.0 1.5 2.0 0 3 10 W ithdr aw al thr es hol d (g) 0 30 60 90 120 150 0 3 10 Li ck in g/ lif tin g tim e (s ec ) † *** † ††
***
**
Paroxetine (mg/kg) Paroxetine (mg/kg) Paroxetine (mg/kg)A
B
C
**
0考察
本研究では,6-OHDA により片側線条体を破壊したマウスにおいて,破壊線条体と反 対側後肢で熱刺激,化学刺激および機械刺激に対する痛覚過敏が生じていることを認め, また,この痛覚過敏に 5-HT 作動性下降性痛覚抑制系の機能低下が関与する可能性を示 した.さらに,5-HT 神経系の賦活作用を有する PXT の慢性投与が片側線条体破壊マウ スの痛覚過敏を軽減することを明らかとした. 6-OHDA 誘発片側線条体破壊マウスのスクリーニングは Iancu ら (2005) の基準をも とに行い,0.5 mg/kg のアポモルヒネ投与により毎分 4 回以上の回転運動を示すマウス を PD モデルマウスとして選別し,以降の実験に用いた.アポモルヒネ誘発回転運動は, 片側線条体破壊マウスにおいてアポモルヒネにより惹起される DA 神経破壊の反対側へ の回転運動で,黒質-線条体の DA 神経系の破壊を反映する表現型と考えられている.本 研究においても,基準以上のアポモルヒネ誘発回転運動を示した動物の一部を抽出して, 線条体の DA 神経脱落を DA 生合成酵素である TH を指標に免疫染色法により確認した ところ,TH の染色シグナルが著明に減少していること,すなわち,DA 神経系が変成・ 脱落していることが認められた. MPTP 投与により作製したヘミ PD モデル動物において協調運動の障害が認められて いる (Park et al., 2015).本研究においても,ロータロッド試験の 16 rpm での試行に おいて,6-OHDA 誘発片側線条体破壊マウスのロッドからの落下時間の短縮を認め, MPTP 投与と同様に 6-OHDA 投与によっても協調運動の障害が発現されることを確認 した.一方,自発運動量試験およびロータロッド試験の 5 rpm での試行において, 6-OHDA 誘発片側線条体破壊マウスにおける運動障害を認めなかった.これらの知見よ り,本実験に使用したマウスは,重篤な運動障害を示さない軽度のヘミ PD モデル動物 であるものと考えられた.6-OHDA 誘発片側線条体破壊動物における全身の痛覚変化については,これまでラッ トを用いた報告がなされており (Cao et al., 2016; da Rocha et al., 2013),Cao ら (2016) は,Hargreaves 試験での逃避潜時の短縮,ならびにホルマリン試験での第Ⅰ 相および第Ⅱ相における finching および licking 時間の増大を,また,da Rocha ら (2013) は,ホットプレート試験での逃避潜時の短縮を見いだしている.すなわち, 6-OHDA 誘発片側線条体破壊ラットが全身性に痛覚過敏を示すことが示されている.一 方で,マウスモデルを用いた本研究では,ホットプレート試験において 6-OHDA 群の 痛覚過敏を認めなかった.これまでの知見より,痛覚過敏の種差を科学的に論じること は不可能であるが,実験系の検出感度,あるいはモデル動物の筋力がラット/マウス間 で異なること,ならびに PD モデルとしての病状ステージが研究グループ間で異なるこ となどが,本研究と既報との差異になった要因であると考えられる. 本研究では,6-OHDA 誘発片側線条体破壊マウスにおける全身性の痛覚感受性変化を 認めなかったため,つぎに線条体破壊側半身と非破壊側半身を区別して痛覚感受性の変 化を解析した.これまでの研究においても,同様の手法により 6-OHDA 誘発片側線条 体破壊ラットの痛覚感受性変化が検討されているが,von Frey 試験において, contralateral の逃避閾値の上昇 (da Rocha et al., 2013) と ipsilateral の逃避閾値の低下 (Takeda et al., 2014),ホルマリン試験でのグルーミング行動において,contralateral の第Ⅰ相での増加と,ipsilateral の第Ⅰ相での低下と第Ⅱ相での増加 (Maegawa et al., 2015 ) が示されるなど,contralateral/ ipsilateral で痛覚鈍麻と痛覚過敏が混在して検 出されており,一定の見解は得られていない.本研究では,6-OHDA 誘発片側線条体破 壊マウスの線条体非破壊側 (contralateral) の後肢において,プランター試験での逃避 潜時の短縮,カプサイシン誘発疼痛試験での licking および lifting 時間の増大,von Frey 試験での逃避閾値の低下,すなわち,DA 神経非破壊側半身で熱刺激,化学刺激および 機械刺激に対する痛み感受性が増大していることを認めた.また本研究では,DA 神経
破壊側半身においては認めなかった.本研究の成績より,6-OHDA 誘発片側線条体破壊 マウスでは黒質‒線条体のドパミン神経細胞の脱落により痛覚過敏が惹起されることが 示唆された. 末梢で受容した痛み刺激は,上行性痛覚伝達系と呼ばれる神経経路を介して伝達され て脳で認識される (Perl, 2011).すなわち,末梢での侵害刺激に起因する情報は脊髄後 角を経由して視床に伝わり,さらに情報は SSC へと伝達されて痛みとして認識されてい る.また,視床からの情報は BLA へも伝達され,痛みに対する不快感を惹起すると考 えられている (Yoshino et al., 2010).一方,上行性痛覚伝達系は脊髄後角において下行 性痛覚抑制系により制御を受けることが知られている (Perl, 2011).下行性痛覚抑制系 の神経経路は視床を起点とし,ACC,PAG を経由して脊髄後角へと投射されており (Xie et al., 2009),さらに,PAG から脊髄後角へは (1) RVM を経由する 5-HT 作動性神経経 路と (2) LC を経由する NA 作動性神経経路の 2 つの経路の存在が示されている (Millan, 2002).したがって,本研究では,6-OHDA 誘発片側線条体破壊マウスの痛覚過敏発現 に関わる神経基盤の探索として,SSC,BLA,ACC,PAG,RVM および LC の 6 つの脳 部位に着目して解析を行った. 核タンパクである c-Fos は,神経活動依存的に一過的に発現誘導されること (Coggeshall, 2005) より,その免疫組織学的検出によって複数の脳部位における神経活 動状況を視覚的に捉えることができる.末梢での痛み刺激受容を伝達する神経活動につ いては,後肢足底へのカプサイシン投与による痛み関連脳部位での c-Fos 発現誘導が示 されており(Horiguchi et al., 2013b),また,Horiguchi ら (2013b) はカプサイシン誘 発痛み関連行動と c-Fos 発現誘導との関連性を報告している.本研究では,まず予備検 討として,正常マウスの後肢足底へのカプサイシン誘発痛み刺激により SSC,BLA,ACC, PAG,RVM および LC で c-Fos が発現誘導されることを確認し (data not shown),さら に,6-OHDA 誘発片側線条体破壊マウスの痛覚過敏発現に関わる神経基盤の解析に応用
した.その解析により,片側線条体破壊マウスでは,線条体非破壊側 (contralateral) 後 肢へのカプサイシン刺激による線条体破壊側 (ipsilateral) 脳内での c-Fos 発現が,ACC, PAG および RVM の 3 部位で減少していること,一方で,SSC,BLA および LC の 3 部 位では差異が認められないことを明らかとした.上述した上行性痛覚伝達経路ならびに 下行性痛覚抑制経路における解析対象脳部位の関わりを考え合わせると,6-OHDA 誘発 片側線条体破壊マウスでは,5-HT 作動性下行性痛覚抑制系による制御が減弱している ことが示唆された.現時点において,線条体の DA 神経の変性・脱落が 5-HT 神経系の 神経活動を低下させる機序については全く不明であるが,進行性の PD 患者において, DA 神経系のみならず 5-HT 神経系が変性・脱落することが報告されており (Miguelez et al., 2014),痛覚過敏を含む PD の新たな薬物療法の開発へ向けて今後のその詳細の解明 が待たれるところである.また今後の詳細な分子機序の解明に向けては,脊髄の神経活 動変化についても解析対象に加えることが重要であろう.なお,PAG および RVM の 2 部位については,左脳/右脳の領域を明確に区別できなかったため,本研究では両側を 合わせて c-Fos 発現細胞を計数することにした.ヘミ PD モデルにおける痛覚刺激と神 経活動の関係について, Reyes ら (2008) は,6-OHDA 誘発片側線条体破壊ラットを 用いた検討において,破壊側 (ipsilateral) 後肢への機械刺激による PAG での c-Fos 発 現が対照動物と差異がないことを報告している.マウスを用いた本研究とは動物種や痛 み刺激の種類が異なるが,PAG および RVM における c-Fos 発現細胞の計数結果におい て,非破壊側 (contralateral) 領域を合わせたことの影響は極めて小さいものと思われ る. c-Fos 発現解析により,6-OHDA 誘発片側線条体破壊マウスにおける 5-HT 作動性下 行性痛覚抑制系の神経活動の低下が認められため,本研究ではつぎに,本マウスの痛覚 過敏に対する 5-HT 神経賦活薬の効果を検討した.5-HT 受容体は 5-HT1~5-HT7の 7 種 類のサブファミリーからなり,さらに 14 サブタイプと非常に多くの種類が存在する
(Hannon and Hoyer, 2008).したがって,本研究では各 5-HT 受容体サブタイプの作動 薬ではなく,5-HT トランスポーターの阻害により 5-HT 神経終末のシナプス間隙で 5-HT 濃度を上昇させる PXT (Gibiino and Serretti, 2012) を用いた.PXT の痛覚過敏へ の効果については,最近,坐骨神経結紮神経障害性疼痛モデルラットにおいて,PXT (10 mg/kg) の単回投与による機械的アロディニアの軽減作用が報告されている (Gai et al., 2014).一方,本研究で PXT は,熱刺激,化学刺激,機械刺激による 6-OHDA 誘発片 側線条体破壊マウスの痛覚過敏に対して,3 および 10 mg/kg の用量での単回投与では 効果は示さず, 10 mg/kg の用量での慢性投与によってのみ改善作用を認めた.これ らの差違が生じた理由として,動物種や病態モデルの違いが第一に挙げられるが,今回 用いた用量の PXT,とくに急性投与の PXT が PAG-RVM-脊髄後角の 5-HT 神経系を賦 活できたか否かは不明である.今後さらに,c-Fos 発現解析あるいはマイクロダイアリ シス法などにより,PXT 投与後の 5-HT 作動性下行性痛覚抑制系の神経活動変化を検証 する必要がある.PXT をはじめセロトニン再取り込み阻害薬 (SSRI) の臨床効果は,う つ病 (Hetrick et al., 2012),うつ病患者の不安症状 (Sanchez et al., 2014; Fava et al., 2000),ならびに PD 患者のうつ症状 (Richard et al., 2012) のいずれに対しても慢性投 与を必要とする.これまでの基礎研究より,SSRI の臨床効果の発現における慢性投与 の必要性については,5-HT 受容体の脱感作 (Yohn et al., 2017; Yamauchi et al. 2006), 神経栄養因子群をはじめとする種々の遺伝子発現変化 (Sillaber et al., 2008),ならびに 樹状突起伸展に関わる細胞内シグナル分子の活性化 (Xu et al., 2018) など様々な分子 機序の関与が示唆されている.したがって,PXT の痛覚過敏の改善効果に関わる分子機 序の解明には,5-HT 受容体の量的変化,神経栄養因子群の量的変化およびに神経の分 化・成熟に関わる細胞内シグナル分子の質的・量的変化に加えて,神経細胞の微細構造 変化の解析が大きな手助けとなることが考えられる. 以上,本研究では,6-OHDA の脳局所微量注入により片側線条体を破壊した PD モデ
ルマウスが,破壊線条体と反対側身体で熱刺激,化学刺激および機械刺激に対して痛覚 過敏を発現することを示し,その発現機序として 5-HT 作動性下行性痛覚抑制系の機能 低下が関与する可能性を見いだした.また,5-HT 神経系の賦活作用をもつ SSRI が,臨 床での抗うつ効果の発現と同様に,慢性投与によって PD モデルマウスの痛覚過敏を改 善することを示した.さらに興味深いことに,本研究で用いた PD モデルマウスの運動 機能障害は,自発運動では見られず,また,ロータロッド試験にて評価した協調運動に 関しても,運動負荷の大きい高速回転においてのみ認められた.近年,PD 患者の非運 動機能障害のなかでも,痛覚過敏は疾患発症の早期より発現することが報告されており (Yoritaka et al., 2013; Schapira et al., 2017),本研究で見いだした知見が,PD 発症早期 の痛覚過敏を反映することが強く示唆された.すなわち,6-OHDA 誘発片側線条体破壊 マウスは PD の痛覚過敏の病態解析に非常に有用であり,今後の新規薬物療法の開発へ の応用が期待される.
謝 謝 辞辞 本稿を終えるにあたり,本研究を行う機会を与えて頂き,終始懇切なる御指導,御鞭撻を 賜りました大阪大学大学院歯学研究科口腔科学専攻高次脳口腔機能学講座 (歯科麻酔学教 室) の丹羽均教授に謹んで感謝の意を表します. また,本研究の進行に際し常にご助言,御指導を戴いた大阪大学大学院歯学研究科口腔 科学専攻顎口腔病因病態制御学講座 (薬理学教室) の田熊一敞教授に深甚なる謝意を表し ます. 最後に本研究に対して御理解,御協力頂きました大阪大学大学院歯学研究科口腔科学専 攻高次脳口腔機能学講座 (麻酔学教室) ならびに顎口腔病因病態制御学講座 (薬理学教室) 教室員の皆様に熱く御礼申し上げます.
参考文献
Ago Y, Kawasaki T, Nashida T, Ota Y, Cong Y, Kitamoto M, Takahashi T, Takuma K, Matsuda T. SEA0400, a specific Na+/Ca2+ exchange inhibitor, prevents dopaminergic
neurotoxicity in an MPTP mouse model of Parkinson's disease. Neuropharmacology. 2011;61:1441‒1451.
Alonso Cánovas A, Luquin Piudo R, García Ruiz-Espiga P, Burguera JA, Campos Arillo V, Castro A, Linazasoro G, López Del Val J, Vela L, Martínez Castrillo JC. Dopaminergic agonists in Parkinson's disease. Neurologia. 2014;29:230‒241.
Bonin RP, Bories C, De Koninck Y. A simplified up-down method (SUDO) for measuring mechanical nociception in rodents using von Frey filaments. Mol Pain. 2014;10:26. doi: 10.1186/1744-8069-10-26.
Broen MP, Braaksma MM, Patijn J, Weber WE. Prevalence of pain in Parkinson's disease: a systematic review using the modified QUADAS tool. Mov Disord. 2012;27:480‒484.
Cao LF, Peng XY, Huang Y, Wang B, Zhou FM, Cheng RX, Chen LH, Luo WF, Liu T. Restoring spinal noradrenergic inhibitory tone attenuates pain hypersensitivity in a rat model of parkinson's disease. Neural Plast. 2016;ArticleID:6383240. doi: 10.1155/2016/6383240.
Coggeshall RE. Fos, nociception and the dorsal horn. Prog Neurobiol. 2005;77:299‒ 352.
Coulombe K, Saint-Pierre M, Cisbani G, St-Amour I, Gibrat C, Giguère-Rancourt A, Calon F, Cicchetti F. Partial neurorescue effects of DHA following a 6-OHDA lesion of the mouse dopaminergic system. J Nutr Biochem. 2016;30:133‒142.
da Rocha JT, Pinton S, Gai BM, Nogueira CW. Diphenyl diselenide reduces mechanical and thermal nociceptive behavioral responses after unilateral intrastriatal administration of 6-hydroxydopamine in rats. Biol Trace Elem Res. 2013;154:372‒ 378.
Fava M, Rosenbaum JF, Hoog SL, Tepner RG, Kopp JB, Nilsson ME. Fluoxetine versus sertraline and paroxetine in major depression: tolerability and efficacy in anxious depression. J Affect Disord. 2000;59:119‒126.
Gai BM, Bortolatto CF, Brüning CA, Zborowski VA, Stein AL, Zeni G, Nogueira CW. Depression-related behavior and mechanical allodynia are blocked by 3-(4-fluorophenylselenyl)-2,5-diphenylselenophene in a mouse model of neuropathic pain induced by partial sciatic nerve ligation. Neuropharmacology. 2014 ;79:580‒589. Gibiino S, Serretti A. Paroxetine for the treatment of depression: a critical update. Expert Opin Pharmacother. 2012;13:421‒431.
Hannon J, Hoyer D. Molecular biology of 5-HT receptors. Behav Brain Res. 2008;195:198‒213.
Hara Y, Ago Y, Taruta A, Hasebe S, Kawase H, Tanabe W, Tsukada S, Nakazawa T, Hashimoto H, Matsuda T, Takuma K. Risperidone and aripiprazole alleviate prenatal valproic acid-induced abnormalities in behaviors and dendritic spine density in mice. Psychopharmacology (Berl). 2017;234:3217‒3228.
Hara Y, Takuma K, Takano E, Katashiba K, Taruta A, Higashino K, Hashimoto H, Ago Y, Matsuda T. Reduced prefrontal dopaminergic activity in valproic acid-treated mouse autism model. Behav Brain Res. 2015;289:39‒47.
Hasebe S, Ago Y, Watabe Y, Oka S, Hiramatsu N, Tanaka T, Umehara C, Hashimoto H, Takuma K, Matsuda T. Anti-anhedonic effect of selective serotonin reuptake inhibitors with affinity for sigma-1 receptors in picrotoxin-treated mice. Br J Pharmacol. 2017;174:314‒327.
Hetrick SE, McKenzie JE, Cox GR, Simmons MB, Merry SN. Newer generation antidepressants for depressive disorders in children and adolescents. Cochrane Database Syst Rev. 2012;11:CD004851. doi: 10.1002/14651858.CD004851.pub3. Horiguchi N, Ago Y, Hasebe S, Higashino K, Asada K, Kita Y, Takuma K, Matsuda T. Isolation rearing reduces mechanical allodynia in a mouse model of chronic inflammatory pain. Pharmacol Biochem Behav. 2013a;113:46‒52.
Horiguchi N, Ago Y, Asada K, Kita Y, Hiramatsu N, Takuma K, Matsuda T. Involvement of spinal 5-HT1A receptors in isolation rearing-induced hypoalgesia in mice.
Psychopharmacology. 2013b;227:251‒261.
Iancu R, Mohapel P, Brundin P, Paul G. Behavioral characterization of a unilateral 6-OHDA-lesion model of Parkinson's disease in mice. Behav Brain Res. 2005;162:1‒10. Kaszuba BC, Walling I, Gee LE, Shin DS, Pilitsis JG. Effects of subthalamic deep brain stimulation with duloxetine on mechanical and thermal thresholds in 6OHDA lesioned rats. Brain Res. 2017;15:233‒241.
LeWitt PA. Levodopa therapy for Parkinson's disease: Pharmacokinetics and pharmacodynamics. Mov Disord. 2015;30:64‒72.
Maegawa H, Morimoto Y, Kudo C, Hanamoto H, Boku A, Sugimura M, Kato T, Yoshida A, Niwa H. Neural mechanism underlying hyperalgesic response to orofacial pain in Parkinson's disease model rats. Neurosci Res. 2015;96:59‒68.
Miguelez C, Morera-Herreras T, Torrecilla M, Ruiz-Ortega JA, Ugedo L. Interaction between the 5-HT system and the basal ganglia: functional implication and therapeutic perspective in Parkinson's disease. Front Neural Circuits. 2014;Article 21. doi: 10.3389/fncir.2014.00021.
Millan MJ. Descending control of pain. Prog Neurobiol. 2002 ;66:355‒474.
Park J, Lim CS, Seo H, Park CA, Zhuo M, Kaang BK, Lee K. Pain perception in acute model mice of Parkinson's disease induced by 1-methyl-4-phenyl-1,2,3,6-tetra- hydropyridine (MPTP). Mol Pain. 2015;17:11‒28.
Parikh V, Naughton SX, Yegla B, Guzman DM. Impact of partial dopamine depletion on cognitive flexibility in BDNF heterozygous mice. Psychopharmacology (Berl). 2016;233:1361‒1375.
Paxinos G, Franklin KBJ. The mouse brain in stereotaxic coordinate, compact 2nd ed. San Diego, CA, USA: Elsevier Academic Press;2004.
Perl ER. Pain mechanisms: a commentary on concepts and issues. Prog Neurobiol. 2011;94:20‒38.
Rana AQ, Ahmed US, Chaudry ZM, Vasan S. Parkinson's disease: a review of non-motor symptoms. Expert Rev Neurother. 2015;15:549‒562.
Reyes S, Mitrofanis J. Patterns of FOS expression in the spinal cord and periaqueductal grey matter of 6OHDA-lesioned rats. Int J Neurosci. 2008;118:1053‒1079.
Richard IH, McDermott MP, Kurlan R, Lyness JM, Como PG, Pearson N, Factor SA, Juncos J, Serrano Ramos C, Brodsky M, Manning C, Marsh L, Shulman L, Fernandez HH, Black KJ, Panisset M, Christine CW, Jiang W, Singer C, Horn S, Pfeiffer R, Rottenberg D, Slevin J, Elmer L, Press D, Hyson HC, McDonald W; SAD-PD Study Group. A randomized, double-blind, placebo-controlled trial of antidepressants in Parkinson disease. Neurology. 2012;78:1229‒1236.
Sanchez C, Reines EH, Montgomery SA. A comparative review of escitalopram, paroxetine, and sertraline: Are they all alike? Int Clin Psychopharmacol. 2014;29:185‒ 196.
Schapira AHV, Chaudhuri KR, Jenner P. Non-motor features of Parkinson disease. Nat Rev Neurosci. 2017;18:435‒450.
Sillaber I, Panhuysen M, Henniger MS, Ohl F, Kühne C, Pütz B, Pohl T, Deussing JM, Paez-Pereda M, Holsboer F. Profiling of behavioral changes and hippocampal gene expression in mice chronically treated with the SSRI paroxetine. Psychopharmacology (Berl). 2008;200:557‒572.
Takeda R, Ishida Y, Ebihara K, Abe H, Matsuo H, Ikeda T, Koganemaru G, Kuramashi A, Funahashi H, Magata Y, Kawai K, Nishimori T. Intrastriatal grafts of fetal ventral mesencephalon improve allodynia-like withdrawal response to mechanical stimulation in a rat model of Parkinson's disease. Neurosci Lett. 2014;573:19‒23.
Tanaka T, Ago Y, Umehara C, Imoto E, Hasebe S, Hashimoto H, Takuma K, Matsuda T. Role of prefrontal serotonergic and dopaminergic systems in encounter-induced hyperactivity in methamphetamine-sensitized mice. Int J Neuropsychopharmacol.
2017;20:410‒421.
Xie YF, Huo FQ, Tang JS. Cerebral cortex modulation of pain. Acta Pharmacol Sin. 2009;30:31‒41.
Xu D, Sun Y, Wang C, Wang H, Wang Y, Zhao W, Bao G, Wang F, Cui Z, Jiang B. Hippocampal mTOR signaling is required for the antidepressant effects of paroxetine. Neuropharmacology. 2018;128:181‒195.
Yamauchi M, Miyara T, Matsushima T, Imanishi T. Desensitization of 5-HT2A receptor
function by chronic administration of selective serotonin reuptake inhibitors. Brain Res. 2006;1067:164‒169.
Yamawaki M, Kusumi M, Kowa H, Nakashima K. Changes in prevalence and incidence of Parkinson's disease in Japan during a quarter of a century. Neuroepidemiology. 2009;32:263‒269.
Yohn CN, Gergues MM, Samuels BA. The role of 5-HT receptors in depression. Mol Brain. 2017;10:28. doi: 10.1186/s13041-017-0306-y.
Yoritaka A, Shimo Y, Takanashi M, Fukae J, Hatano T, Nakahara T, Miyamato N, Urabe T, Mori H, Hattori N. Motor and non-motor symptoms of 1453 patients with Parkinson's disease: prevalence and risks. Parkinsonism Relat Disord. 2013;19:725‒ 731.
Yoshino A, Okamoto Y, Onoda K, Yoshimura S, Kunisato Y, Demoto Y, Okada G, Yamawaki S. Sadness enhances the experience of pain via neural activation in the anterior cingulate cortex and amygdala: an fMRI study. Neuroimage. 2010;50:1194‒ 1201.