九州大学学術情報リポジトリ
Kyushu University Institutional Repository
リグノセルロースの酵素糖化反応の活性化に関する 研究
高, 子晴
九州大学大学院生物資源環境科学府森林圏環境資源科学講座
https://doi.org/10.15017/25189
出版情報:Kyushu University, 2012, 博士(農学), 課程博士 バージョン:
権利関係:
1
.
Evaluation on the enhancing enzymatic hydrolysis of lignocellulose
Gao Ziqing
2012
2
CHPTER1 Introduction
1-1. Biomass ethanol
1-1-1. Structure and composition of lignocellulosic biomass 1-1-2. Biomass ethanol production process
1-1-3. The role of pretreatment in biological processing of cellulosic biomass 1-1-4. Biological pretreatment
1-2. Cellulase composition and function 1-3. β-glucosidases
1-3-1. Structures of β-glucosidases 1-3-2. Catalytic mechanisms 1-3-3. Function of β-glucosidases 1-3-4. Application of β-glucosidases 1-4. The aim of this study
CHPTER2
Screening the suitable wood rotting fungi for corn stover pretreatment
3
2-1. Introduction 2-2. Material and method 2-2-1. Fungal stains and isolation 2-2-2. Preparation of corn stover 2-2-3. Fungal screening
2-2-4. Saccharification
2-2-5. Identification of microorganism 2-3. Results
2-4. Discussion 2-5. Summary
CHPTER3
Physical and chemical characterizations of corn stover pretreated by G.
trabeum KU-41 3-1. Introduction 3-2. Material and method 3-2-1. Corn stover pretreatment
4
3-2-2. Chemical component analysis 3-2-3. Cellulose crystallinity
3-2-4. Initial adsorption capacity of cellulase 3-2-5. Scanning electron microscopy (SEM) 3-2-6. Specific surface area determination 3-3. Results
3-4. Discussion 3-5. Summary
CHPTER4
Screening the microorganism with high activity of β-glucosidases 4-1. Introduction
4-2. Material and method 4-2-1. Chemicals
4-2-2. Strains isolation
4-2-3. Screening of β-glucosidases-producing strains 4-2-4. Enzyme assay
5
4-2-5. Protein concentration determination 4-3. Results
4-4. Discussion 4-5. Summary
CHPTER5
Strain Identification 5-1. Introduction 5-2. Material and method
5-2-1. DNA extraction and PCR amplification from cultures 5-2-2. DNA sequencing
5-2-3. Phylogenetic Analysis 5-3. Results
5-4. Discussion 5-5. Summary
6
CHPTER6
Effect of different carbon source on β-glucosidases production in F.
proliferatum NBRC109045 6-1. Introduction
6-2. Material and method 6-2-1. Material
6-2-2. Electrophoresis and zymogram 6-3. Results
6-4. Discussion 6-5. Summary
CHPTER7
Partly purification of a glucose-tolerant β-glucosidase from F.
proliferatum NBRC109045 7-1. Introduction
7-2. Material and method 7-2-1. Growth condition
7-2-2. Effect of ammonium sulfate saturation to β-glucosidase activity
7
7-2-3. Test tube pilot experiment to determine starting conditions for ion-exchange chromatography
7-2-4. Determine optimal binding pH
7-2-5. Determine salt concentration for binding and elution 7-2-6. Enzyme purification
7-3. Results 7-4. Discussion 7-5. Summary
Conclusion References Acknowledge
8
CHPTER1 Introduction 1-1. Biomass ethanol
1-1-1.Structure and composition of lignocellulosic biomass
Lignocellulosic biomass is composed primarily of carbohydrate polymers (cellulose and hemicellulose) and phenolic polymer (lignin). Lower concentrations of various other compounds, such as proteins, acids, salts, and minerals, are also present. Cellulose and hemicellulose, which typically make up two-thirds of cell wall dry matter (dry matter: the portion of biomass containing no water), are polysaccharides that can be hydrolyzed to sugars and then fermented to ethanol. Bio-ethanol production is directly related to cellulose, hemicellulose, and individual sugar concentration in the feedstock.
Lignin cannot be used in fermentation processes; however, it may be useful for other purposes (Vermerris, 2008).
1-1-2. Biomass ethanol production
Biomass ethanol is the ethanol made from non-grain plant materials known as biomass. The bulk of most plants is fibrous material consisting of cellulose, hemicellulose and lignin. The bioconversion of biomass to ethanol requires hydrolysis of carbohydrate polymers, cellulose being changed to its constituent monomeric sugars
9
prior to microbial fermentation. Due to the complicated structure of the cell wall in biomass, additional pretreatment before hydrolysis is needed for biomass ethanol production compared to grain ethanol.
1-1-3. The role of pretreatment in biological processing of cellulosic biomass
In the context of biological processing of cellulosic biomass to sugars for fermentation to ethanol and other products, pretreatment generally refers to the disruption of the naturally resistant carbohydrate-lignin shield that limits the accessibility of enzymes to cellulose and hemicellulose (McMillan, 1994; Wyman et al., 2005; Holtzapple, 1993). However, the choice of pretreatment technology is not trivial and must take into account sugar-release patterns and solid concentrations for each pretreatment in conjunction with their compatibility with the overall process, feedstock, enzymes, and organisms to be applied (Aden et al., 2002; Wooley et al., 1999)
1-1-4. Biological pretreatment
Biomass pretreatment is an essential step in biomass ethanol production with high yield (Hendriks and Zeeman, 2009; Yang and Wyman, 2008). Many studies have been reported about different pretreatment methods for enhancing the digestibility of biomass (Keller et al., 2003; Kumar et al., 2009). Biological pretreatment that utilizes the metabolite of microorganism in nature to break up the cell wall of biomass for ethanol
10
production is a promising technology due to its advantages of having a low energy requirement and being friendly to the environment (Hakala et al., 2004; Zhang et al., 2007). Compared to chemical pretreatment, it is no need to recycle the chemical and does not bring exotic materials to environment. These reductions in the severity of pretreatment conditions could result in less biomass degradation and consequently lower inhibitor concentrations compared to conventional thermochemical pretreatment (Keller et al., 2003). Fungal pretreatment using wood-rot fungus is one of the most effective methods for enhancing the efficiency of enzymatic saccharification (Monrroy et al., 2011; Ray et al., 2010; Schilling et al., 2009; Taniguchi et al., 2005). Fissore et al (2010) evaluated a process of combined brown-rot decay –chemical delignification as a pretreatment for bioethanol production. The combination of brown-rot fungi and organosolv processes result in 210 ml ethanol/kg wood. Some thermochemical pretreatment methods have been performed for biomass ethanol production (Bobleter, 1994; Garrote et al., 1999; Kohlmann et al., 1995; Mosier et al., 2005; Yang and Wyman, 2008). However, biological pretreatment has been neglected. As is widely known, due to the ability to degrade lignin extensively, white-rot fungi (WRF) have received considerable attention for their potential to remove lignin for bio-ethanol pretreatment (Zhang et al., 2007). In contrast, brown-rot fungi (BRF) such as Gloeophyllum trabeum,
11
has different mechanisms for the degradation of wood. BRF rapidly depolymerize the cellulose and hemicellulose in wood with modified lignin in the brown residue: BRF degrade lignocellulose via a theorized two-part mechanism, with modification of the plant cell wall induced non-enzymatically and secretion of cellulases and hemicellulases likely occurring after modifications (Goodell et al., 1997; Hyde and Wood, 1997). The initial stages of decay are thought to involve Fenton chemistry (Fe2++H2O2) for the production of hydroxyl anions and radicals (Koenigs, 1974). The low molecular weight reactants, unlike enzymes, are small enough to penetrate the wood lignocellulose fabric, and have been shown in immunolabeling studies to be present throughout the S2 layer of the brown rot-degraded cell wall (Jellison et al., 1997). Cellulase production by brown rot fungi is different in that it is typically constitutive, not influenced by free glucose concentrations, and most often lacks exo-acting cellobiohydrolase (Martinez et al., 2009; Valášková and Baldrian, 2006). G. trabeum has the ability of fermenting sugar to ethanol (Rasmussen et al., 2010). The enzymatic and non-enzymatic mechanisms used by G. trabeum to degrade wood could potentially be employed for the bioconversion of other biomass, such as corn stover. The complicated structural modification of the cell wall plays a role in the initial degradation of BRF in the pretreatment for the purpose of bio-ethanol production.
12
1-2. Cellulase composition and function
For microorganisms to hydrolyze and metabolize insoluble cellulose, extracellular cellulases must be produced that are either free or cell associated. Components of cellulase systems were first classified based on their mode of catalytic action and have more recently been classified based on structural properties (Henrissat et al., 1998).
Three major types of enzymatic activities are found: (i) endoglucanases or 1,4-β-D-glucan-4-glucanohy- drolase (EC 3.2.1.4), (ii) exoglucanases, including 1,4-β-D-glucan glucanohydrolases (also known as cellodextrinases) (EC 3.2.1.74) and 1,4-β-D-glucan cellobiohydrolases (cellobiohydrolases) (EC 3.2.1.91), and (iii) β-glucosidases or β-glucoside glucohydrolases (EC 3.2.1.21). Endoglucanases cut at
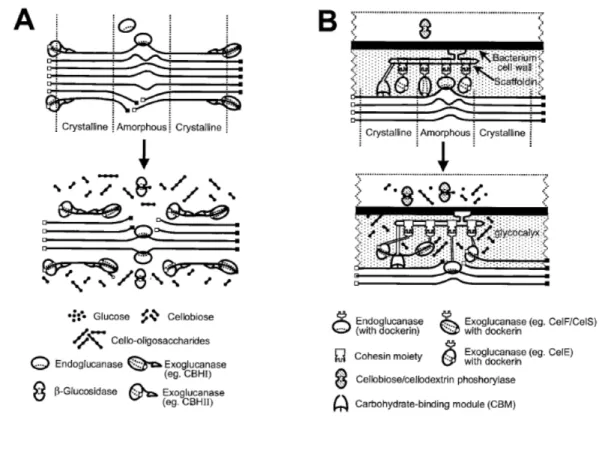
random at internal amorphous sites in the cellulose polysaccharide chain, generating oligosaccharides of various lengths and consequently new chain ends. Exoglucanases act in a processive manner on the reducing or nonreducing ends of cellulose polysaccharide chains, liberating either glucose (glucanohydrolases) or cellobiose (cellobiohydrolase) as major products. Exoglucanases can also act on microcrystalline cellulose, presumably peeling cellulose chains from the microcrystalline structure (Teeri, 1997). β-Glucosidases hydrolyze soluble cellodextrins and cellobiose to glucose (Fig.
1-1). Cellulases are distinguished from other glycoside hydrolases by their ability to
13
hydrolyze β-1,4-glucosidic bonds between glucosyl residues. The enzymatic breakage of the β-1,4-glucosidic bonds in cellulose proceeds through an acid hydrolysis mechanism, using a proton donor and nucleophile or base. The hydrolysis products can either result in the inversion or retention (double replacement mechanism) of the anomeric configuration of carbon-1 at the reducing end (Birsan et al., 1998; Withers, 2001).
1-3. β-glucosidases
1-3-1. Structures of β-glucosidases
β-Glucosidases play a role in the carbohydrate metabolism in many organisms by
acting on β-glycosidic linkages of cello-oligosaccharides containing β-d-1,4 glycosidic bonds. These enzymes are classified under 3 glycoside hydrolase (GH) families: GH1, 3 and 9, according to the classification by Coutinho and Henrissat (1999). β-glucosidases have various folding structures, but the overall fold of the catalytic domain is similar in each GH family (Fig 1-2). The families GH1 belong to the Clan GH-A, and they all have similar (b/a)8-barrel domains structure and their active sites contain two conserved carboxylic acid residues on b-strands 4 and 7, serving as the catalytic acid/base and nucleophile, respectively (Henrissat et al., 1995; Jenkins et al., 1995). GH9 enzymes have (a/a)6-barrel structures, Only a few GH9 proteins have been verified to be
14
β-glucosidases (Park et al., 2001; Qi et al 2008), as most proteins in this family are
endoglycosidases. The GH3 β-glucosidases and exoglucanases have a two-domain structure, a (b/a)8-barrel followed by an a/b sandwich comprising a 6-stranded b-sheet sandwiched between three a-helices on either side (Varghese et al., 1999).
1-3-2. Catalytic mechanisms
Glycoside hydrolases perform catalysis using two mechanisms, one with inversion and one with retention of chirality at the anomeric carbon (Park et al., 2002). Both of these mechanisms use a pair of acidic and nucleophilic residues, usually carboxylic acids, on either side of the sugar, approximately 5 Å apart in the retaining mechanism, and 10 Å apart in the inverting mechanism, in which a water molecule must fit between the catalytic base and the substrate. The GH9 β-glucosidases use an inverting mechanism, in which an activated water molecule makes a direct nucleophilic attack on the anomeric carbon to displace the aglycone in a single step, as shown in Fig. 1-3a (Qi et al., 2008). The catalytic base extracts a proton from the incoming water molecule while the catalytic acid protonates the leaving group aglycone. In contrast, most β-glucosidases that have been characterized (i.e., GH1 and GH3) are retaining enzymes, and they perform catalysis in two steps, glycosylation and deglycosylation (Fig. 1-3b).
In glycosylation, the aglycone departs with the donation of a proton from the catalytic
15
acid/base and nucleophilic attack of the catalytic nucleophile on the anomeric carbon to yield an α-linked covalent enzymeglycone intermediate. In the deglycosylation step, the process is reversed, as a water molecule attacks with basic assistance from the catalytic acid/base to displace the catalytic nucleophile from the glucose. Both the glycosylation and deglycosylation steps are thought to pass through oxocarbenium-ion-like transition states. The glucose of the incoming substrate has sometimes been observed to be distorted into a 1S3 skew boat as it moves toward the 4H3 half-chair shape in the first transition state, although in other structures it is poorly defined by the electron density, apparently due to high mobility (Davies et al., 2003; Czjzek et al., 2000). The structures of certain putative transition state mimics have also been solved in the active site and shown to have a structure close to the 4H3 half-chair, although others appeared to inhibit by mechanisms other than transition state mimicry (Davies et al., 2003; Gloster et al., 2004; Gloster et al., 2006)
1-3-3. Function of β-glucosidases
Mammals contain several β-glucosidases, including the family GH1 lactase-phloridzin hydrolase and cytoplasmic β-glucosidase, the GH30 human acid β-glucosidase (GBA1) and the bile acid β-glucosidase or GBA2. These enzymes are thought to play roles in metabolism of glycolipids and dietary glucosides. In addition, a
16
group of related family GH1 proteins is thought to play signaling functions. Although the Drosophila melanogaster genome contains only one GH1 gene, suggesting that insects may not have expanded this gene family at an early stage, other insects have adapted glycosides and glycoside hydrolases from the plants on which they feed for protection and digestive purposes (Zagrobelny et al., 2008). It is in plants that β-glucosidases have been found to play the widest array of biological functions, which
include roles in defense, symbiosis, cell wall catabolism and lignification, signaling, and plant secondary metabolism. Several putative β-glucosidases genes have been shown either to be induced by biotic or abiotic stress or to be necessary for successful response to the stress (Malboobi and Lefebvre, 1997; Van de Ven et al., 2000; Kawasaki et al., 2000; Thorlby et al., 2004; Lipka et al., 2005). Although much research has been done on β-glucosidases from microorganisms, most of it has focused on their application rather than their endogenous function. As such, most of the enzymes that have been studied in the context of their natural function are those involved in bioconversion to produce glucose from plant biomass, or in breaking through plant cell walls to establish pathogenic or symbiotic relationships (Gilbert et al., 2008). Bacterial β-glucosidases are often components of large complexes called cellosomes, contain polysaccharide degrading endoglucanases and carbohydrate binding proteins to localize the complex
17
and to the cellulose surface and the cell membrane (Doi and Kosugi, 2004; Carvalho et al., 2007). Alternatively, some microorganisms secrete soluble endoglycosidases and exoglycosidases for this function, including exoglucanases/β-glucosidases. Fungi, such as the white rot fungus Phanerochaete chrysosporium, may contain cytoplasmic β-glucosidases and extracellular exoglucanases, some of which may act in metabolism
of the organism’s own cell wall, in addition to plant cell wall biomass metabolism (Lymar et al., 1995; Igarashi et al., 2003; Tsukada et al., 2006).
1-3-4. Application of β-glucosidases
β-Glucosidases have potential roles in various fields such as the food, pharmacology
and cosmetic industries and also in the valorisation of some products, due to the properties of this enzyme to convert and to synthesize biomolecules of high added value (Esen 1993). There are hundreds of different β-glucosidic flavor precursors in plants, and their hydrolysis often enhances the quality of the beverages and foods produced from them (Gϋnata, 2003; Esen, 2003). Aside from flavor enhancement, foods, feeds, and beverages may be improved nutritionally by release of vitamins, antioxidants, and other beneficial compounds from their glycosides (Opassiri et al., 2004). Indeed, β-glucosidase can either degrade or synthesize small carbohydrate polymers, depending on particular experimental conditions (Pérez et al. 2008).
18
As noted above, β-glucosidases are of interest for biomass conversion, since conversion of β-glucans, the largest source of biomass in the world. Limiting factors in conversion of cellulose to glucose for fermentation to alcohol include the inhibition of cellulases by oligosaccharides and the lack of adequate β-glucosidase production by certain microorganisms used for biomass breakdown. Thus, the identification and production of β-glucosidases, especially those with high glucose tolerance, has been of interest, and applicable β-glucosidases have been isolated from bacteria and fungi (Gillbert et al., 2008; Lymar et al., 1995)
1-4. The aim of this study
The bioconversion of renewable resources, especially lignocellulosic residues, into useful chemicals and fuel ethanol is a worldwide challenge for biotechnology.
Lignocellulose bio-refining generally involves three main steps: pretreatment, enzymatic hydrolysis, and fermentation. Commercialization of bio-ethanol is hindered by the high cost of pretreatment process and cellulase enzymes required for lignocellulose hydrolysis. Therefore, develop a suitable pretreatment method and screen the fungi or bacterial with high-efficient cellulase system are the necessary approach for successful commercialization.
In this study, we set forth the methods that enhancing the efficiency of bio-ethanol
19
production in two directions (1) develop biological pretreatment method that using brown-rot fungus to pretreat corn stover for bio-ethanol production. To gain a deeper understanding the brown-rot fungus pretreatment, the fungal-pretreated corn stover was evaluated in detail, including the lignin and structural carbohydrate contents, cellulose crystallinity, initial adsorption capacity of cellulase, SEM and specific surface area; (2) Isolate and screen microbles to produce considerable quantities of β-glucosidases. In the present study, a potential β-glucosidases-producing fungus NBRC109045 was isolated and identified as Fusarium proliferatum. Under optimized conditions, F. proliferatum produces β-glucosidases with an activity of 3.3 U/ml based on pNPG as substrate and an activity of 426 U/ml based on cellobiose as substrate. In this paper, we described ways that (a) isolating and screening microbes to produce considerable quantities of β-glucosidases; (b) modifying the method of zymogram to prove that different carbon
sources direct varied β-glucosidases expression in F. proliferatum; (c) assaying partial purification to prove high catalytic efficiency of β-glucosidase produced by F.
proliferatum growing in corn stover + wheat bran based medium.
20
FIGURE 1-1 Schematic representation of the hydrolysis of amorphous and microcrystalline cellulose by noncomplexed (A) and complexed (B) cellulase systems. The solid squares represent reducing ends, and the open squares represent nonreducing ends. Amorphous and crystalline regions are indicated. Cellulose, enzymes, and hydrolytic products are not shown to scale (Lynd et al., 2002)
21
FIGURE 1-2 Structures of β-glucosidase from different GH families. These include β-glucosidases or related enzymes from GH1 (Zea mays ZmGlu1, PDB code 1E1E), GH3 (Hordeum vulgare Exo I β-glucan glucohydrolase, PDB code 1EX1), GH5 (Candida albicans exo-β-(1, 3)-glucanase Exg exoglucanase, PDB code 1CZ1), GH30 (Homo sapiens, acid β-glucosidase/glucocerebrosidase GBA1, PDB code 2V3D), and GH9 (Vibrio parahaemolyticus, putative exoglucanase, PDB code 3H7L). The structural cartoons are colored in a spectrum from blue to red from their N- to C-termini, with the catalytic nucleophile and acid–base residues shown in stick for those enzymes in which they are known. The ligands shown are glucose in the GH3 barley ExoI and N-butyl-deoxyno- jirimycin in the GH30 human GBA1, both of which are shown with carbons in pink. The human GBA2 (bile acid β-glucosidase) shows low levels of sequence similarity to (α/α)6 enzymes, suggesting its catalytic domain may be similar to the GH9 structure. Drawn with Pymol (DeLano)
22
FIGURE 1-3 Retaining catalytic mechanisms of inverting and retaining β-glucosidases.
A: The inverting mechanism that is seen in family GH9 glycoside hydrolases, including β-glucosidases. A single displacement of the aglycone by the water leads to an anomeric carbon with inverted chirality.
B: The commonly accepted mechanism for hydrolysis with retention of anomeric configuration as seen GH Clan A and family GH3 β-glucosidases. The glucosyl moiety is distorted into an 1S3 skew boat upon binding to the enzyme in preparation to form the 4H3 half chair conformation of the proposed transition state (Rye and Withers, 2000;
Davies et al., 2003). The first step is glycosylation, in which the catalytic acid donates a proton to the leaving group, while the catalytic nucleophile attacks from the opposite side to form an α-linked intermediate. In the second, deglycosylation step, the catalytic base (the same carboxylate as the catalytic acid) extracts a proton from a water molecule, improving its nucleophilic power to attack at the anomeric carbon and displace the enzyme. Hydrolysis by either mechanism is equivalent in the organism, since mutarotation of the released glucose will lead to a racemic mixture of glucose in solution after a short time
23
CHPTER2
Screening the suitable wood rotting fungi for corn stover pretreatment 2-1. Introduction
The wood-rooting fungi including white-rot fungi (WRF) and brown-rot fungi (BRF) are the most important and efficient microorganisms involved in natural process of wood decomposition. This group plays an important ecological role and is widely distributed occurring in tropical and temperate environments in the world. Within this group, the species vary largely with regard to their relative cellulolytic versus ligninolytic capacity. These fungi possess hydrolytic enzymes like cellulases, pectinases, xylanases and, ligninolytic complex composed of lignin peroxidase (LiP) and manganese peroxidas (MnP) and laccase, which in general are triggered by their substrates.
Eriksson and Vallander (1982) reported as much as a 23% reduction in pulp-refining energy required by incubating spruce and pine chips for 2-wk with the white-rot fungus Phanerochaete chrysosporium at 35–40℃. It was suggested that fungal pretreatment disrupts the wood structure apparently by partially breaking down the lignin/ carbohydrate complex.
Sawada and his collaborators (1995) investigated the effects of fungal pretreatment
24
and steam explosion pretreatment on enzymatic saccharification of beech wood meal.
Their work indicates that fungal pretreatment followed by less severe steaming maximizes enzymatic saccharification. The investigators suggested that the lignin network covering the holocellulose (cellulose and hemicellulose) is broken down by successive fungal pretreatment and steam explosion pretreatments, which together maximize subsequent enzymatic saccharification of beech wood meal.
The concept that fungal pretreatment lowers the energy requirements of thermomechanical pulping of lignocellulosic biomass could potentially be applied to enzyme-based biomass conversion processes to lower the thermochemical pretreatment severity. This potential for fungal pretreatment can be explained by the ability of certain fungi to disrupt the plant cell wall chemistry, resulting in partial breakdown of the lignin/carbohydrate complex. Furthermore, fungal pretreatment can readily be incorporated into the feedstock handling area of biomass ethanol processes.
The purpose of this study is to choose the suitable wood-rotting fungi for corn stover pretreatment. Forty strains of wood-rot fungi (33 strains of WRF and 7 strains of BRF) were screened for corn stover pretreatment. A strain of brown-rot fungus KU-41 was selected for corn stover pretreatment due to its having the highest conversion of cellulose to glucose (CCG). A molecular biological identification showed that KU-41
25
was most closely related to Gloeophyllum trabeum.
2-2. Material and method 2-2-1. Fungal stains and isolation
We selected 33 strains of WRF and 7 strains of BRW that decay wood significantly for use in corn stover pretreatment. Daedalea albida NBRC6510, Daedalea dickinsii NBRC31163, Fomitopsis insularis NBRC6887, Fomitopsis pinicola NBRC8705, Gloeophyllum trabeum NBRC6509 and Phlebia brevispora TMIC33929 were obtained
from the Biological Resource Center (Chiba, Japan) and Tottori Mycological Institute (Tottori, Japan). Coriolus hirstus YK-505 (Kondo et al., 1994a), Ceriporia lacelate MZ-340 and Phanerochaete sordida YK-624 (Kondo et al., 1994b) were obtained from stock cultures in our laboratory. KU-41 and other strains were isolated from decayed wood in a cedar forest at Morotuka, Higashiusuki district, Miyazaki, Japan. Stock cultures of the fungi were maintained on potato dextrose agar (PDA, Difco, Detroit, MI, USA) plates at 4ºC.
2-2-2. Preparation of corn stover
Corn stover was collected from Yingkou city in Liaoning Province in China. The
26
stover was then chopped air-dried and stored at room temperature. The constituents of corn stover (lignin: 23.3%, hemicellulose: 28.1%, cellulose: 35.4%, cell solubles: 12.4%, ash: 2.8%) were determined by following a detergent digestion protocol (Goering and Van Soest, 1970). Before pretreatment, the corn stover was milled and sized to 250 -350 μm, use water to remove the soluble content, then filtered, freeze-dried and subjected to treatment with wood-rot fungi.
2-2-3. Fungal screening
The mycelium was transferred to a new PDA medium in a 9-cm diameter Petri dish and incubated at 30ºC. Once the fungus covered most of the PDA plate, agar plates with mycelium were transferred to a sterile blender cup containing 25 ml of sterile water and homogenized for 30 s. One milliliter of the white-rot fungus homogenate was used to inoculate 10 ml of low-nitrogen basal III medium (Tien and Kirk, 1988) and 1 ml brown-rot fungus homogenate was inoculated into 10 ml potato dextrose broth (PDB) medium in 100-ml Erlenmeyer flasks. When the fungi covered the medium surface, the mixture was vortexed with 25 ml sterile water to mix it well, and 1 ml was inoculated into 5 g corn stover powder and then cultivated at 30ºC for 30 days (Keller et al., 2003).
Prior to fungal inoculation, the corn stover was sterilized in the autoclave for 20 min at
27
121ºC, and then 4 ml of sterilized water was added to the corn stover to keep the moisture content at 50%. Corn stover treated in the same conditions without fungal inoculation was used as the control. After pretreatment, the corn stover was washed to remove the soluble content with 250 ml water, then filtered and freeze-dried. Weight loss was determined as the difference between the weight of the corn stover at the beginning and at the end of the pretreatment according to the following formula: weight loss (%) = (W1-W2)/W1*100, where W1 is the weight of the sample before pretreatment, and W2
represents the weight of the sample after pretreatment. All experiments were performed in triplicate.
2-2-4. Saccharification
CCG was conducted to estimate the efficiency of the bio-pretreatment. Corn stover without pretreatment was used as the control. A 1 g corn stover sample with 0.7 g cellulase T3 (HBI Enzymes Inc. cellulase activity: 240 FPU/g; xylanase activity: 63.62 U/g) was added into 50 ml of 0.1 M sodium acetate buffer (pH 4.6) and incubated with gentle shaking (100 rpm) at 60ºC. After 48 h, the released of glucose was determined by a Biosensor BF-5 (Oji Scientific Instruments Co., Itd). All experiments were performed in triplicate. Upon evaluation of the effects of pretreatment, CCG was defined as the
28
percentage of cellulose in the raw material converted to glucose, taking into account the weight loss during pretreatment.
2-2-5. Identification of microorganism
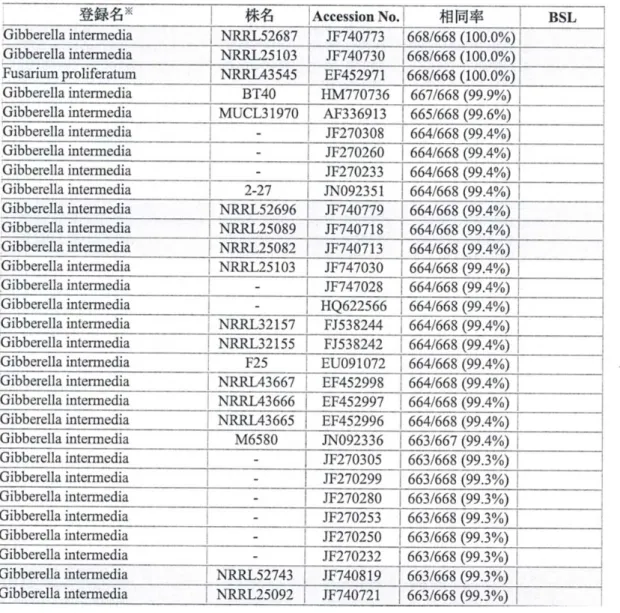
The ITS1-5.8-ITS2 ribosomal RNA gene of KU-41 was amplified by PCR using the primer set ITS1 primer (5'-TCCGTAGGTGAACCTGCGG-3') and ITS4 primer (5'-TCCTCCGCTTATTGATATGC-3'). The 626-bp amplicon obtained was cloned and sequenced. The sequences were proofread, edited, and merged into composite sequences using Clustalx-1.83.1. The fungus was determined to be most closely related to Gloeophyllum trabeum by comparing the strain with related strains in GenBank. The GenBank accession numbers of KU-41 are JF682770-JF682771 .
2-3. Results
Fungal stains and isolation Fungal screening
The CCG results of the pretreatment of corn stover with wood-rot fungi after 48 h of hydrolysis are shown in Fig. 2-1. Compared to the control, the CCG levels of corn stover pretreated with most white-rot fungi were not increased except in those pretreated
29
by Pycnoporus coccineus, W2 and W3, but this increase of CCG was not significant. We tried to use 7 strains of brown-rot fungi, which had the ability to decay wood significantly. The CCG of corn stover was not increased by pretreatment with these brown-rot fungi except for G. trabeum NBRC6509 and KU-41. In particular, the fungus KU-41 enhanced the CCG of corn stover. The CCG of pretreated corn stover was determined by the glucose yield and weight loss. Although some strains of wood-rot fungi have great degrading ability, they were not suitable for the corn stover pretreatment due to the simultaneous degradation of cellulose. Of these 40 strains of wood-rot fungi, KU-41 caused the greatest increase in CCG. Therefore, KU-41 was selected for further study.
Fungal identification
The ITS rDNA region of KU-41 showed the highest identity (99%) with G. trabeum, and it was named G. trabeum KU-41.
30
0 20 40 60
control KU-41 D. dickinsil NBRC31163 D. albida NBRC 6510 F. pinicola NBRC8705 F. insularis NBRC6887 G. trabeum NBRC6509 B-1 P. sordida YK-624 C. hirstus YK505 P. brevispora P. chrysosporium NBRC31249 P. coccineus C. lacerata MZ-340 W-1 W-2 W-3 W-4 W-5 W-6 W-7 W-8 W-9 W-10 W-11 W-12 W-13 W-14 W-15 W-16 W-17 W-18 W-19 W-20 W-21 W-22 W-23 W-24 W-25 W-26 W-27
The conversion of cellulose to glucose (%)
31
FIGURE 2-1 CCG of corn stover pretreated with wood-rot fungi after 48 hours hydrolysis. W means white-rot fungus unidentified; B means brown-rot fungus unidentified. CCG indicates glucose amount produced in the hydrolysis process/
cellulose amount in un-pretreated corn stover. Values are means ± SD of triplicate samples.
32
2-4. Summary
Forty strains of wood-rotting fungi (33 strains of white-rot fungi and 7 strains of brown-rot fungi) were screened for the pretreatment of bio-ethanol made from corn stover. Cellulase was used for cellulose saccharification from corn stover pretreated by wood-rotting fungi to test the efficiency of pretreatment. As a result, KU-41 strain of brown-rot fungus was selected for corn stover pretreatment due to the greatest increase in CCG. Therefore, KU-41 that identified as G. trabeum with using ITS rDNA gene sequences was selected for further study.
33
CHPTER3
Physical and chemical characterizations of corn stover pretreated by G.
trabeum KU-41 3-1. Introduction
The cellulose saccharification efficiency in corn stover pretreated by G. trabeum KU-41 was increased by 45%. (Chapter 2) The work introduced in this chapter is to find out the reasons why the cellulose saccharification was enhanced significantly.
Lignocellulosic material consists of mainly three different types of polymers, namely cellulose, hemicellulose and lignin, which are associated which each other (Fengel and Wegener, 1984).
In this study, physiochemical changes resulting from the pretreatments by G.
trabeum KU-41 with corn stover were determined. Chemical characterizations were performed to determine glucan, xylan and lignin for untreated and pretreated corn stover.
Physical characteristics including cellulose crystallinity, cellulase adsorption capacity of corn stover solids and specific surface area were measured.
3-2. Material and method 3-2-1. Corn stover pretreatment
Another strain of G. trabeum NBRC6430 obtained from the Biological Resource
34
Center (Chiba, Japan) was chosen to compare with the efficiency of corn stover pretreatment with G. trabeum KU-41. The biological pretreatment with G. trabeum KU-41 was carried out in a 250 ml Erlenmeyer flasks with 10 g corn stover powder and 40 ml distilled water (80% moisture content) for a 20-day pretreatment. The other conditions were the same as described in the fungal screening. The released glucose was determined by Biosensor BF-5. The xylose from enzyme hydrolysis was determined by HPLC (HITACHI, RI detector, Shodex Asahipak NH2P-50 4E column; eluent:
CH3CN/H2O=75/25; flow rate: 1 ml/min; temperature: 30ºC).
3-2-2. Chemical component analysis
The lignin composition in corn stover pretreated with G. trabeum was determined by the method published by the National Renewable Energy Laboratory (Sluiter et al., 2008). The carbohydrate composition was determined by total acidic hydrolysis followed by analysis of monomeric sugars by GC-MS (Mura et al., 2009). Corn stover pretreated by G. trabeum was milled and then subjected to acid hydrolysis. A 300 mg sample of pretreated corn stover was hydrolyzed with 72% sulfuric acid at 30ºC for 4 h.
The secondary hydrolysis was performed after dilution with water to a 3% sulfuric acid concentration by autoclaving at 121ºC for 60 min. Then the hydrolysate was filtered
35
through a glass filter. The acidic filtrate containing sugars was adjusted to a pH of 5-6 with barium hydroxide. The reduced monomeric sugars with NaBH4 were acetylated and then analyzed using a Shimadzu gas chromatography-mass spectrometer (GC-17A and GCMS-QP5050) with a glass capillary column, DB-5. The temperature program:
140 ºC for 3 min; 140ºC-280ºC, at a rate of 6 ºC /min; 280ºC-320ºC, at a rate of 10 ºC /min; 320ºC for 5 min.
3-2-3. Cellulose crystallinity
The crystallinity of the pretreated corn stover powder was measured by high resolution X-ray diffractometry (XD-D1 X-Ray diffractometer, Shimadzu, Japan). The measurement conditions were 40 kV and 40 mA. There were triplicate samples, a1, a2, and a3, pretreated with the same fungus, respectively. Each sample was scanned from 2θ=5° to 35° with a step size of 0.05 and tested three times to get the mean values ā.
Crystallinity (%) was defined as [(I002−Iam)/I002] × 100%, where I002 and Iam are the maximum intensity at 2θ=22.6º and the minimum intensity at 2θ=18.7º, respectively.
The crystallinity value of corn stover pretreated with certain strain of fungus is expessed as the mean ± SD for ā1, ā2 and ā3.
36
3-2-4. Initial adsorption capacity of cellulase
A total of 0.21 g of commercial cellulase (Cellulosin T3, HBI Enzymes Inc.) was added to 0.3 g corn stover pretreated with G. trabeum KU-41 and G. trabeum NBRC6430. The corn stover pretreated with fungus without the addition of cellulase was used for the control. The mixture was added to 15 ml, 100 mM sodium acetate buffer (pH 4.6). These reactions were placed at 4°C with 100 rpm to reach adsorption equilibrium and were stopped by centrifugation at 8000 rpm for 20 min. After 90 min, centrifugated at 8000rpm for 20min. Protein in the supernatant was measured by using the Bradford protein assay (Bio-Rad, Hercules, California, USA). Adsorbed cellulase was determined as the difference between the amount of protein initially added and the amount of un-adsorbed protein in the supernatant.
3-2-5. Scanning electron microscopy (SEM)
SEM pictures of untreated and pretreated corn stover (dried powder of particle size between 250 µm and 300 µm) were taken at magnifications of 500 times using a JSM-5600LV1 scanning electron microscope at 5 kV. Prior to taking the pictures, the samples were sputter- coated with a thin layer of gold.
37
3-2-6. Specific surface area determination
The specific surface area of the pretreated corn stover sample was estimated by the Brunauer-Emmett-Teller (BET) method (Brunauer et al., 1938).
3-3. Results
Pretreatment conditions and enzymatic hydrolysis
In the process of corn stover pretreatment with wood-rot fungi, moisture content and pretreatment period were important factors that affected the efficiency of pretreatment. The effect of moisture content on the pretreatment period and the CCG of corn stover pretreated with KU-41 was tested (data not shown). The highest CCG was obtained after 20-day pretreatment with 80% moisture content. The efficiency of pretreatment was evaluated by enzymatic hydrolysis. Fig. 3-1 shows enzymatic hydrolysis of the samples after 48 h. As expected, corn stover without pretreatment was capable of converting cellulose into glucose with an efficiency of 38.7%. High glucose yields were obtained when the corn stover was pretreated by KU-41 and NBRC6430.
The CCG levels increased by 47% and 42% over 20-day treatment with KU-41 and NBRC6430,respectively. The strain of G. trabeum KU-41 seemed to be the most promising fungus for biological pretreatment of corn stover.
38
Hemicellulose serves as a connection between lignin and cellulose fibrils and gives the whole cellulose-hemicellulose-lignin network more rigidity (Laureano-Perez et al., 2005). Xylan is the main component of hemicellulose in corn stover. In corn stover pretreated with KU-41 and NBRC6430, 32.0% and 31.4% xylan was transformed into xylose and released after 48 h of hydrolysis, in comparison with only 11.2% xylan transformation in the un-pretreated sample (Fig. 3-1). The xylose yield in the saccharification solution of pretreated corn stover was 2.8-fold greater than that of the un-pretreated sample suggested that during corn stover pretreatment with G. trabeum, the hemicellulose structure modification or breakage is beneficial for xylose release during enzymatic hydrolysis. An increase in xylose release led to an increase in cellulose accessibility, which consequently resulted in the glucose yield increasing in corn stover pretreated by G. trabeum KU-41and G. trabeum NBRC6430.
Compositional analysis
Compositional analysis of corn stover pretreated with fungus (Table 3-1) gave a good indication of the possible changes in the different components. As the pretreatment progressed, hemicellulose was the component that was removed in the greatest
39
proportion. Biodegradation of hemicellulose from corn stover was 43% with G. trabeum KU-41 and 43% with G. trabeum NBRC6430. As hemicellulose is connected with cellulose microfibrils, the prior removal of these can facilitate cellulose degradation.
The content of glucan decreased slightly, reaching removal percentages of 16.8% and 18.3% with G. trabeum KU-41 and G. trabeum NBRC6430, respectively. Consistent with the degradation mechanism of brown-rot fungi, lignin was essentially undegraded by the two strains of G. trabeum.
Crystallinity determination
Lignocellulosic biomass is mainly composed of cellulose (amorphous and crystalline), hemicellulose and lignin. Crystallinity is believed to affect enzymatic saccharification of cellulose (Laureano-Perez et al., 2005). However, due to the complex structure of the cell wall in lignocellulosic biomass, it is impossible to separate cellulose from other components completely or to measure the cellulose crystallinity directly.
X-ray measurement of CrI is still the most frequently-used method to estimate the effect of pretreatment on biomass crystallinity. The CrI values of pretreated corn stover samples here were determined by measuring the relative amount of crystalline cellulose in the total solid. The cellulose crystallinity (CrI %) in corn stover pretreated with G.
40
trabeum KU-41(55.3±0.02) and G. trabeum NBRC6430 (55±1.37) was slightly
increased compared to the control (53.9±2.01). The slight increase of CrI in the pretreated samples suggested that the cellulose became more exposed after pretreatment.
However, all materials give rise to X-ray scattering, with the amorphous part including not only amorphous cellulose but also hemicellulose and lignin. Hemicellulose and lignin have been determined to have diffractograms similar to amorphous cellulose (Thygesen et al., 2005), giving wide unspecific peaks, which may affect the results.
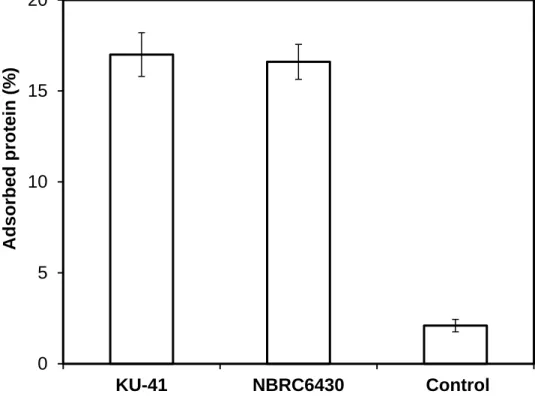
Initial adsorption capacity
Jeoh et al. (2007) claimed that the hydrolysis rate and/or yield is directly related to the amount of adsorbed enzymes. Fig. 3-2 shows the initial adsorption capacity of cellulase onto corn stover after pretreatment with G. trabeum. The pretreated corn stover showed a significant increase of the initial adsorption capacity. The initial adsorption capacity increased about 7-fold after pretreatment with G. trabeum. Lignin is believed to impede enzyme access to glucan chains by its binding and steric hindrance (Change and Holtzapple, 2000; Kim and Holtzapple, 2006). In this study, the lignin ratio in corn stover pretreated with G. trabeum KU-41 increased by 30% (Table 3-1), while the CCG was enhanced by 46% (Fig. 3-1). Evidently, the lignin content is not the
41
main reason of the increase of the cellulase initial adsorption capacity in corn stover pretreated with G. trabeum. Kumar and Wyman (Kumar et al., 2009) proposed that lignin did not directly control the cellulose accessibility but restricted the xylan accessibility, which in turn controlled access to cellulose. The xylose yield of pretreated corn stover increased 2.8-fold (Fig. 3-1). So we have reason to believe that the relationship among lignin, hemicellulose and cellulose changed such that the lignin influence was weakened in cellulose hydrolysis during the process of corn stover pretreatment with G. trabeum KU-41 and G. trabeum NBRC6430.
Microstructure analysis
SEM images of corn stover pretreated with fungus are shown in Fig. 3-3 at magnifications of 500 and 1000. The surface of the untreated corn stover was compact and rigid, suggesting that this property hinders the accessibility of cellulase to cellulose (Fig. 3-3E-F). More pores and cracks were created on the surface of the corn stover (Fig.
3-3A-B, 3-3C-D), and fungal pretreatment seemed to disrupt the biomass structure to some extent.
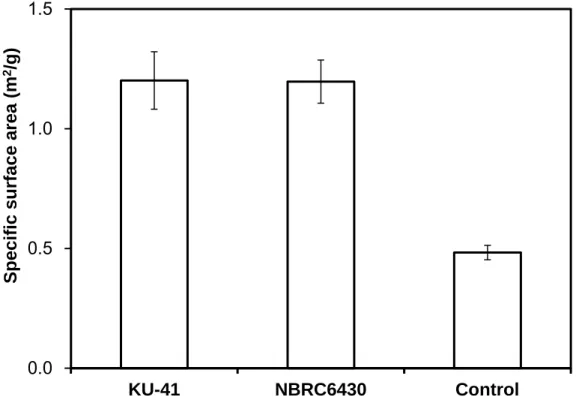
The specific surface area and pore distribution are generally considered to play
42
important roles in accessibility. It has been reported that the cellulase digestibility of pretreated biomass is limited by cellulose accessibility (Jeoh et al., 2007). Here (Fig.
3-4), a significant difference in the specific area between the control value and that of pretreatment samples was determined. The specific surface of corn stover pretreated with G. trabeum was increased 2.5-fold.
Based on comprehensive analysis of Fig. 3-3 and Fig. 3-4, the SEM demonstrated that the fungal pretreatment could cause severe degradation and evident damage to the intact cell structure. Both the selective biodegradation of cellulose and hemicellulose components by the fungus and fungal penetration might contribute to the pore production. Removal of hemicellulose increased the mean pore size of the substrate and therefore increased the probability of the cellulose to be hydrolyzed (Grethlein, 1985;
Palonen et al., 2004). Drying of the pretreated lignocellulose can cause a collapse in pore structure, resulting in decreased enzymatic hydrolysis (Grous et al., 1986).
Cellulosic particles have both external and internal surfaces. The internal surface area of porous cellulose particles depends on the capillary structure and includes intraparticulate pores (1–10 nm) as well as interparticulate voids (Marshall and Sixsmith, 1974). External surface area is closely related to shape and particle size, and can be
43
estimated by microscopic observation (Gilkes et al., 1992). Zhang and Lynd (2004) suggested that cellulase can get trapped in the pores if the internal area is much larger than the external area, which is the case for much lignocellulosic biomass. Pore production, which was a result of the pretreatment, led to a significant increase in specific surface area. These phenomena are probably the reasons that the CCG of corn stover pretreated with G. trabeum was enhanced compared to the control.
44
Table 3-1 Main composition of unpretreated and pretreated corn stover and ratio of component left after pretreatment
Pretreatment
Composition of pretreated corn stover (%)
Ratio of component left in pretreated corn stover (%)
G X L+A G X L+A
G. trabeum KU-41 39.1 12.0 33.0 83.2 57.0 96.1
G. trabeum NBRC 6430 38.4 12.0 33.3 81.7 57.0 97.0
Control 35.7 16.0 26.1 100.0 100.0 100.0
G: glucan.
X: xylan.
L+A: total lignin + ash.
45
FIGURE 3-1 CCG and xylose yield of pretreated corn stover after 48 hours hydrolysis.
Bar expresses CCG; line expresses xylose yield. Xylose yield means xylose amount produced during hydrolysis/xylose released by totally acid hydrolysis in pretreated corn stover. Values are means ± SD of triplicate samples.
0 10 20 30 40 50
0 10 20 30 40 50 60 70
KU-41 NBRC6430 Control
Xylose yield (%)
The conversion of celllulose to glucose (%)
46
FIGURE 3-2 Initial adsorption capacity of cellulase onto corn stover after pretreatment
with G. trabeum. Values are means ± SD of triplicate samples.
0 5 10 15 20
KU-41 NBRC6430 Control
Adsorbed protein (%)
47
48
FIGURE 3-3 SEM images of un-pretreated and pretreated corn stover samples with G.
trabeum. [A-B] pretreated sample with G. trabeum KU-41 500x (A), 1000x (B); [C-D] pretreated sample with G. trabeum NBRC 6430 500x (C), 1000x (D); [E-F] un-pretreated sample 500x (E), 1000x (F).
49
FIGURE 3-4 Specific surface area of corn stover pretreated and un-pretreated with G.
trabeum. Values are means ± SD of triplicate samples.
0.0 0.5 1.0 1.5
KU-41 NBRC6430 Control
Specific surface area (m2/g)
50
3-4. Discussion
Fungal pretreatment followed by thermochemical pretreatment could potentially lower the severity requirements of acid, temperature and pressure in thermochemical pretreatment (Keller et al., 2003). Lower pretreatment severity is expected to translate directly into lower chemical consumption, and because of lower temperature requirements, possibly lower steam cost. These reductions in severity could also reduce the capital costs and result in lower biomass degradation and consequently lower inhibitor levels. This potential of fungal pretreatment can be explained by the ability of certain fungi to disrupt the structure of the cell wall, resulting in partial breakdown of the lignin/carbohydrate complex. Lignin is a complex aromatic polymer based on phenylpropane units. It contains few hydrolysable bonds and so it is poorly susceptible to attack by hydrolytic enzymes. Lignin and its associated compounds have been found to inhibit cellulase action (Deshpande and Eriksson, 1984). There is evidence that lignin may retard cellulase action by adsorbing cellulase enzyme (Ishihara and Ishihara, 1979).
Delignification is a method of increasing the biomass susceptibility of hydrolysis.
However, some papers reported that attempts to correlate the delignification of biomass to its increasing susceptibility to enzyme digestion were often inconclusive or contradictory depending on variables such as the type of substrate and the pretreatment
51
conditions (Fan et al., 1981; Millett et al., 1975; Sudo et al., 1976). Stranks (1961) has explained why the lignin-holocellulose relationship rather than the lignin concentration was responsible for the effect of the lignin on cellulose saccharification. Wong et al (Wong et al., 1988) have suggested that lignin distribution maybe as important as accessibility in determining the enzymatic digestibility of pretreated fibers by steam explosion. Gharpuray et al. (Gharpuray et al., 1983) also discussed the probable interdependent effects of lignin concentration and surface area on hydrolysis. The complex physical relationship between lignin and cellulose fibrils revealed by Ruel et al.
(1982) also supports the proposition that lignin distribution is more important to enzymatic hydrolysis than lignin concentration. Yelle et al. (Yelle et al., 2008) reported that brown-rot of spruce wood by Gloeophyllum trabeum resulted in a marked, non-selective depletion of all intermonomer side-chain linkages in the lignin. As shown in Table 3-1, the lignin ratio in corn stover pretreated with G. trabeum was increased compared to the control. But the CCG (Fig. 3-1) in corn stover pretreated with G.
trabeum was enhanced significantly. We believe that the structure of lignin in the
process of fungal pretreatment was modified despite the fact that amount was not decreased. The bonds between cellulose and lignin or hemicellulose and lignin were modified or broken (Filley et al., 2002), which may be one reason that the CCG in corn
52
stover pretreated with G. trabeum KU-41 was enhanced.
The association between the cellulose components and the hemicellulose appears to be of a physical nature, as there have been no reports of chemical bonds between cellulose and hemicellulose molecules. In a series of papers (Ishihara and Ishihara, 1979; Shimizu and Usami, 1978; Shimizu, 1980) on the effects of various pretreatments on enzymatic hydrolysis, it was found that in pretreatments which removed only lignin, the rate of digestion of the hemicellulosic component was similar to that of cellulose. This result suggested that the relationship between cellulose and hemicellulose was that one must be removed to facilitate an attack on the other. Increases in cellulose digestibility have been associated with decreased hemicellulose content; more than 80% of hemicellulose was degraded by a dilute sulfuric acid pretreatment (Yang and Wyman, 2008). In this study, 43% hemicellulose degradation was observed. One interpretation of these data is that removing xylan during pretreatment facilitated cellulase access to the cellulose, which resulted in increased digestibility. This concept has been discussed in the literature as relating changes in the porosity of the biomass sample to digestibility (Esteghlalian et al., 2001). We believe that the removal of lignin or hemicellulose or both serves one purpose: disruption of carbohydrate-lignin networking, which enhances enzyme adsorption, generally labeled as accessibility.
53
With lignocellulosic materials, crystallinity appears to be of lesser importance than the association between lignin (and hemicellulose) and surface area. Pretreatments such as ball milling reduced crystallinity but did not have a corresponding effect on hydrolysis (Gharpuray et al., 1983). When cardboard and newspaper were used, a decrease in crystallinity resulted in a significant increase in saccharification (Rivers and Emert, 1988). With cryomilled rice hulls, the relationship was not apparent (Sasaki et al., 1979), while alkali treatment of rice straw resulted in an increase in the crystallinity index from 40 to 52% and an increase in solubilization from 12 to 63% (Tanaka et al., 1979). It was also found that the crystallinity of cellulose in wood was not an inhibitory factor in their enzymatic hydrolysis experiments (Matsumura et al., 1977). The effects of various chemical pretreatments on a range of substrates have resulted in no change or an actual increase in crystallinity after the pretreatment, while the rate and extent of enzymatic hydrolysis were markedly increased. Puri (1984) concluded that any increase was probably due to the removal of amorphous material rather than any increase in the ordered region and that crystallinity was not as important a factor as generally believed.
In this study, Avicel [(R) RH-101 (Sigma)] was used as the sole carbohydrate for cultivating G. trabeum KU-41; the fungus grew very slowly and the crystallinity of Avicel did not change. This result was consistent with the finding that most brown-rot
54
fungi showed slow growth on crystalline cellulose despite their rapid degradation of cellulose in wood (Cohen et al., 2005). The present results indicate that the fungal pretreatment cleavages cellulose in the noncrystalline regions. Part of the amorphous cellulose degradation led to a change of the cellulose structure, which increased the surface area with access to cellulase in pretreated corn stover.
3-5. Summary
Although biotechnology presents important opportunities for achieving very low costs, pretreatment of naturally resistant cellulosic materials is essential if we are to achieve high yields from enzymatic hydrolysis. Fungus pretreatment with G. trabeum proved to be an effective way of increasing enzymatic hydrolysis of corn stover for bio-ethanol production. As is widely known, pretreatment with white-rot fungi could increase the enzymatic hydrolysis of biomass through lignin degradation. What happened in the process of pretreatment with brown-rot fungus is the crux of this paper.
The results showed that the pretreatment by G. trabeum had a partial defibrating effect on corn stover. Partial removal of xylan and modification of the structure of lignin resulted in disrupting the structure of the cell wall. The disruption of the structure of the cell wall increased the accessibility of cellulase to lignocellulose.
A future goal is to improve the efficiency of the fungus pretreatment method
55
described in this study by substantially reducing the cost and accelerating its commercial application. One approach that is attracting attention is the pretreatment of corn stover in vitro with the goal of a short pretreatment time. We believe that with further research and the development of technology, fungus pretreatment could achieve a real decrease in the cost of bio-ethanol production.
56
HPTER4
Screening the microorganism with high activity of β-glucosidases
4-1. Introduction
Biofuels derived from lignocellulosic biomass are emerging as promising alternatives to fossil fuels to meet the increasing global energy demands (Ragauskas et al., 2006). One of the key steps in bioconversion process is the enzymatic hydrolysis of the cellulose polymers in the biomass to monomeric sugars that are subsequently fermented to ethanol (Percival et al., 2006; Adsul et al., 2007). The three main categories of players in cellulose hydrolysis are cellobiohydrolases (or exo-1, 4-β-glucanases) (EC 3.2.1.91), endo-1, 4-β-glucanases (EC 3.2.1.4), and β-glucosidases (EC 3.2.1.21) (Beguin and Aubert, 1994). The endo-1, 4-β-glucanases randomly attack cellulose in amorphous zones releasing oligomers. The cellobiohydrolases liberate cellobiose from reducing and non-reducing ends. And finally β-glucosidases hydrolyze the cellobiose and in some cases the cellooligosaccharides to glucose (Ryu, 1980; Wood, 1985). Cellulose polymers are through sequential and cooperative actions of these enzymes degraded to
57
glucose. Cellobiohydrolases and endoglucanases are often inhibited by cellobiose, making β-glucosidases important in terms of avoiding decreased hydrolysis rates of cellulose over time due to cellobiose accumulation (Workman and Day, 1982). The low efficiency and high costs associated with the enzymatic hydrolysis process presents a major bottleneck in the production of ethanol from lignocellulosic feedstocks (Banerjee et al., 2010). For the enzymatic conversion of biomass to fermentative sugar on a commercial scale, it is necessary to have all cellulolytic components at the optimal level.
Since β-glucosidases activity is low in many microbial preparations used usually for the saccharification process (Enari, 1990). It is necessary to supply additional β-glucosidases to such reaction. In order to optimize the use of different biomasses, it is
important to identify new β-glucosidases with improved abilities on the specific biomasses as well as with improved abilities such as stability and high conversion rates.
The β-glucosidases can be arranged in three groups related to localization: intracellular, cell wall associated, and extracellular. Industrially, primarily the extracellular β-glucosidases are of interest (Soewnsen, 2010). The number of fungal species on earth
is estimated to 1.5 million of which as little as approximately 5% are known (Hawksworth, 1991; Hawksworth, 2001), a statement that calls for a more directed effort for unraveling the potential of unknown species found in nature. The
58
identification and characterization of new fungal species are often encountered in literature.
The object of this study is to screening microorganism with high cellulase activity.
The research group chose Jatropha wood chip, cassava wood chip, coconut shell, sugarcane, Jatropha stems and leaves, cassava stems and leaves and rice straw as cellulose resource and dipped in Cuc Phuong National Park and Ba Be National Park for degradation experiment. Then we separated and screened 18 strains with the ability of crystal cellulose degradation. During the 18 strains of fungi, a potent β-glucosidases-producing fungus SIID11460 was isolated.
4-2. Material and method
4-2-1. Chemicals
Unless specified otherwise, all chemicals were of analytical grade. Solubilized crystalline cellulose was obtained from Kyokuto Seiyaku Co., Ltd, Japan.
4-methylumbelliferyl-β-D-glucoside (MUG) was products of Sigma Chemical Co., (St.
Louis, Mo, USA). 4-Nitrophenyl-β-D-glucopyranoside monohydrate (pNPG) was bought from Tokyo Chemical Industry Co., Ltd, Japan. Corn stover was collected from Yingkou city in Liaoning Province in China. Wheat bran was obtained from private company.
59
4-2-2. Strains isolation
Wood chip of Jatropha carcass, branch and leaves of J. carcass, wood chip of Manihot esculenta, branch and leaves of M. esculenta, coconut shell, sugarcane, and rice straw
were used as lignocellulosic sources for degradation in Vietnamese National Park (Ba Be and Cuc Phuong). One month later, lignocellulosic sources were dug up. All strains that would be screened were isolated from degraded biomass samples and washed soil collected. Isolated strains were inoculated on solubilized crystalline cellulose (CC) plates and CMC plates to cultivate for two weeks (Deguchi et al. 2007). The microbes that could grow on CC and CMC were picked up and inoculated onto malt extract agar (MEA).
4-2-3. Screening of β-glucosidase-producing strains
The first step of screening
For primary screening, strains from MEA were plated on potato dextrose agar (PDA) medium in a 9-cm diameter Petri dish and incubated at 30ºC for 5 days. Then the colonies were inoculated on β-glucosidases screening agar containing 1% of CMC, 0.5% of MUG, 1.5% of agar, and Mandels salts (Daenen et al., 2008). The cultures were incubated at 30°C for 3 days. Then the plates were observed under UV light. Colonies
60
which showed fluorescence were sorted out. It is because methylumbelliferyl (MU) which was released from MUG by β-glucosidases can emit fluorescence when induced by UV light.
The second step of screening
For secondary screening, the mycelium of the β-glucosidase-producing isolates obtained from the primary screening was transferred to a new PDA medium in a 9-cm
diameter Petri dish and incubated at 30ºC. Once the fungus covered most of the PDA plate, agar plates with mycelium were transferred to a sterile blender cup containing 25 ml of sterile water and homogenized for 30 s. 10 ml of the fungal homogenate was used to inoculate into β-glucosidases secondary screening medium containing 1% corn stover + 1% wheat bran in 100 ml, pH 5.0 Mandels salts medium with KH2PO4 2 g l-1, (NH4)2SO4 1.4 g l-1, urea 0.69 g l-1, CaCl2·2H2O 0.3 g l-1, MgSO4·7H2O 0.3 g l-1 and 1 ml trace elements solution composed of MnSO4 1.6 g l-1, ZnSO4 2 g l-1, CuSO4 0.5 g l-1, CoSO4 0.5 g l-1 (Saibi et al., 2011) and incubated at 30 °C, 150 rpm for 5 days. Crude enzyme extract was obtained by centrifuging the liquid medium at 20,000 g for 20 (4°C) min and collecting the supernatant for confirming the β-glucosidases activity.
4-2-4. Enzyme assay
β-glucosidases activity towards p-nitrophenyl-β-D-glucopyranoside (pNPG) was
61
measured by the amount of p-nitrophenol (pNP) liberated from pNPG using a calibration curve at 410 nm (Cai et al., 1998). The reaction mixture contained 0.5 ml, 2 mM pNPG in 50mM sodium acetate buffer (pH 5.0) and an appropriately diluted enzyme solution 0.125 ml. After incubation at 45ºC for 10 min, the reaction was stopped by adding 1.25 mL, 1M Na2CO3 and the color that formed as a result of pNP liberation was measured at 410 nm. One unit of β-glucosidases activity was defined as the amount of enzyme liberating 1 μmol of pNP per minute under the assay conditions.
Specific activity is defined as the number of units per milligram of protein.
4-2-5. Protein concentration determination
Protein concentrations in the enzyme preparations were determined by the method of Bradford (Bradford, 1976) with reference to a standard calibration curve for bovine serum albumin (BSA).
4-3. Results
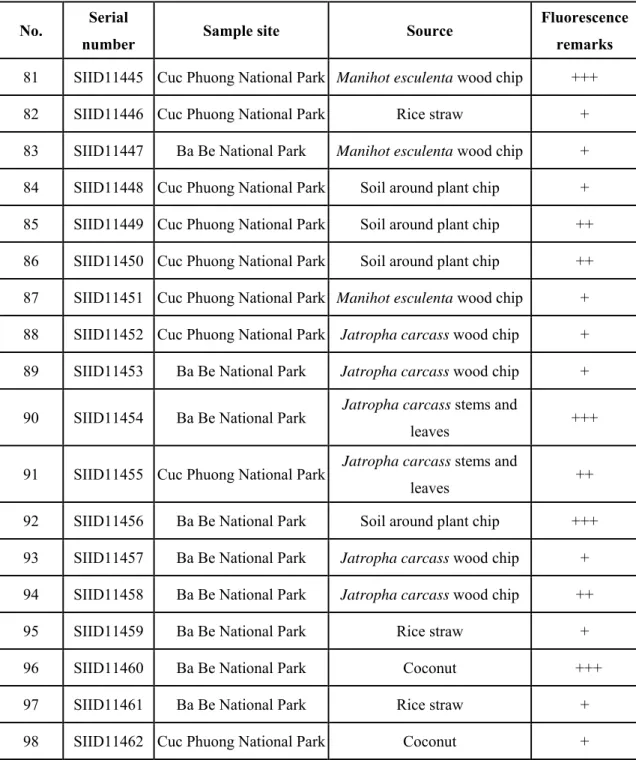
Screening of β-glucosidase-producing strain.
MUG released MU when MUG was catalyzed by β-glucosidases, and MU emitted fluorescence. In order to screen the best strain for β-glucosidases production, firstly the potential strains were cultivated in medium that contained MUG. Of these potential
62
strains, 4 strains showed the brightest fluorescence (Table 4-1). Next, these 4 strains were prepared in a medium that contained 1% of corn stover and 1% of wheat bran for five days. Of these 4 strains, SIID 11460 showed the highest activity of β-glucosidases.
Therefore, SIID 11460 was selected for further research.
63
Table 4-1 Screening of microorganism with β-glucosidases production.
No. Serial
number Sample site Source Fluorescence
remarks
81 SIID11445 Cuc Phuong National Park Manihot esculenta wood chip +++
82 SIID11446 Cuc Phuong National Park Rice straw + 83 SIID11447 Ba Be National Park Manihot esculenta wood chip + 84 SIID11448 Cuc Phuong National Park Soil around plant chip + 85 SIID11449 Cuc Phuong National Park Soil around plant chip ++
86 SIID11450 Cuc Phuong National Park Soil around plant chip ++
87 SIID11451 Cuc Phuong National Park Manihot esculenta wood chip + 88 SIID11452 Cuc Phuong National Park Jatropha carcass wood chip + 89 SIID11453 Ba Be National Park Jatropha carcass wood chip +
90 SIID11454 Ba Be National Park Jatropha carcass stems and
leaves +++
91 SIID11455 Cuc Phuong National Park Jatropha carcass stems and
leaves ++
92 SIID11456 Ba Be National Park Soil around plant chip +++
93 SIID11457 Ba Be National Park Jatropha carcass wood chip + 94 SIID11458 Ba Be National Park Jatropha carcass wood chip ++
95 SIID11459 Ba Be National Park Rice straw + 96 SIID11460 Ba Be National Park Coconut +++
97 SIID11461 Ba Be National Park Rice straw + 98 SIID11462 Cuc Phuong National Park Coconut + +++: with the brightest fluorescence
++: with brighter fluorescence +: with bright fluorescence